
Abstract
A soft food diet leads to changes in the periodontal ligament (PDL). These changes, which have been recognized for more than a century, are ascribed to alterations in mechanical loading. While these adaptive responses have been well characterized, the molecular, cellular, and mechanical mechanisms underlying the changes have not. Here, we implicate Wnt signaling in the pathoetiology of PDL responses to underloading. We show that Wnt-responsive cells and their progeny in the PDL space exhibit a burst in proliferation in response to mastication. If an animal is fed a soft diet from the time of weaning, then this burst in Wnt-responsive cell proliferation is quelled; as a consequence, both the PDL and the surrounding alveolar bone undergo atrophy. Returning these animals to a hard food diet restores the Wnt signaling in PDL. These data provide, for the first time, a molecular mechanism underlying the adaptive response of the PDL to loading.
Introduction
The periodontal ligament (PDL), a fibrous connective tissue that anchors the tooth to the alveolar bone, is a mechanosensory tissue: it derives some of its mechanical strength from the helical structure of type I collagen, which accounts for >90% of its organic content (Liu et al. 1995). The fibers of the PDL form a meshwork and insert into bone and cementum as Sharpey’s fibers. The PDL sustains and transmits mechanical stresses (Storey 1973), and it readily adapts to changes in mechanical stimuli (Rios et al. 2008; Huang, Liu, et al. 2016). For example, decreased masticatory loading results in PDL thinning and bone loss (Denes et al. 2013). The molecular/cellular basis for this mechanoresponsiveness in the PDL was not known.
A number of cell types exist in the PDL, any of which may be mechanoresponsive. For example, most PDL space is occupied by fibroblasts, but there are osteoblasts and osteoclasts on the alveolar bone surfaces of the PDL and cementoblasts on the root surfaces of the PDL. Various in vitro analyses suggest that these cell types have some mechanoresponsive characteristics (Kim et al. 2007; Leucht, Kim, Currey, et al. 2007; Yan et al. 2012; Monnouchi et al. 2015). In addition, a population of undifferentiated mesenchymal stem/progenitor cells occupy the PDL space (Marchesan et al. 2011), and at least some of these stem/progenitor cells may be mechanoresponsive (Zhang et al. 2012; Wei et al. 2015; Zhang et al. 2016). In vitro, these PDL stem/progenitor cells can differentiate into osteoblasts, cementoblasts, and fibroblasts; whether they are responsible for mediating PDL mechanosensitivity was not known.
We sought to better understand how the PDL senses and responds to mechanical forces. This is clearly a broad question, and strategies to address this question can range from mathematical modeling (Tuna et al. 2014) and in vitro testing of the PDL (Oortgiesen et al. 2012) to genetic studies that disrupt PDL function (Lim et al. 2014; Takimoto et al. 2015). Like other investigators before us (Lin et al. 2013), we opted to examine in detail the in vivo changes that are elicited in response to physiologically relevant mechanical loads. For example, mechanical loading of the PDL begins around the time that an animal initiates mastication, and this event is accompanied by a remarkable reorganization of the PDL (Huang, Tang, et al. 2016). Throughout life, this organized structure is maintained and is ultimately responsible for the ability of the dentition to withstand masticatory forces on the order of hundreds of newtons (Natali 2003). The question that launched our study was as follows: What molecular pathways and cell populations in the PDL are triggered by functional loading of the dentition? By identifying some of the pathways involved in this adaptive response, we gained insights into 1) how these molecular pathways regulate the PDL stem/progenitor population and 2) what roles they perform in mechanically induced reorganization of the PDL.
Methods and Materials
Animals
All experimental protocols followed the ARRIVE guidelines (Animal Research: Reporting of In Vivo Experiments) and were approved by the Stanford Committee on Animal Research (13146). Axin2CreERT2/+;R26RmTmG/+ mice (018867 and 007576, respectively) and Axin2LacZ/+ mice (009120) were purchased from Jackson Laboratories and housed in a temperature-controlled environment with 12-h light/dark cycles. To induce Cre expression in Axin2CreERT2/+;R26RmTmG/+, tamoxifen (4 mg/ 25 g, body weight) was delivered intraperitoneally, and animals were sacrificed at indicated time points.
Histology Assays (Picrosirius Red, Pentachrome)
For picrosirius red staining, slides were stained with picrosirius solution (0.5 g, sirius red; 35780, Pfaltz & Bauer, Inc.; dissolved in 500 mL of saturated picric acid solution) and then viewed under polarized light. Pentachrome staining was performed as described (Leucht, Kim, Wazen, et al. 2007). In this staining, mature bone is stained yellow; osteoid and mineralized cartilage are stained green; nuclei are black; and cytoplasm is red.
Statistical Analyses
Results are presented as mean ± SD, with n equal to the number of samples analyzed. All statistical analyses were performed with GraphPad 5.0 (GraphPad Software). Histomorphometric results were based on Student’s t tests. Significance was attained at P < 0.05.
For feeding, micro–computed tomography (µCT), molecular analyses, Xgal staining, and finite element model, see the Appendix.
Results
Morphologic Changes in the PDL Are Associated with Masticatory Loading
During development, the disorganized PDL occupies space between the tooth bud and surrounding alveolar bone (Fig. 1A). Around this age (e.g., postnatal day 9 [P9]), when young animals are still nursing, the PDL space is largely composed of nonproliferating cells (Fig. 1B, C). The extracellular matrix (ECM) in the PDL space lacks an organized linear arrangement (Fig. 1D).

Morphologic changes in the PDL are associated with masticatory loading. (
Around the time when mice begin to consume hard chow, the PDL space undergoes a dramatic rearrangement: the PDL becomes a functional tissue populated by mitotically active cells (Fig. 1E, F), and the collagen-rich ECM orients itself perpendicular to the direction of masticatory force (Fig. 1G). In adult mice, this ECM arrangement is maintained (Fig. 1H, J) but mitotic activity is not (Fig. 1I), and the PDL becomes a quiescent tissue. Collectively, these data demonstrate that the PDL undergoes a morphologic transformation commensurate with its functional adaptation to loading.
Given the importance of Wnt signaling in the growth (Rooker et al. 2010) and maintenance (Yin et al. 2015) of the PDL, we wondered if this morphologic adaptation was accompanied by changes in the distribution of Wnt-responsive cells. A Wnt reporter stain (e.g., Axin2LacZ/+) was used to visualize Wnt-responsive cells in the PDL (Yuan, Pei, Zhao, Tulu, et al. 2018). At P9, suckling pups exhibited few Wnt-responsive cells in the PDL space (Fig. 1K). At P18, when pups had initiated mastication, Wnt-responsive cells were distributed throughout the PDL (Fig. 1L). In adulthood, the number of Wnt-responsive cells declined slightly (Fig. 1M).
We considered these data collectively: as mice progress from suckling to mastication, cell density in the PDL space increases. Cell density in the PDL then reaches a plateau, which is maintained into adulthood (blue line, Fig. 1N), suggesting that functional loading is a stimulus for PDL cell proliferation. Endogenous Wnt signaling paralleled this proliferative burst and then leveled off into adulthood. Together these observations suggested that Wnt signaling, as demonstrated by the presence of Wnt-responsive cells, was a link between the onset of masticatory loading and functional adaptation of the PDL to that load.
Masticatory Loading Triggers the Proliferation of Wnt-Responsive Cells That Contribute to the Functioning PDL
To test a direct role of Wnt-responsive cells in PDL response to functional loading, we used a lineage tracer strain of Wnt reporter mice (Yuan, Pei, Zhao, Tulu, et al. 2018) to identify and follow progeny of Wnt-responsive cells at different stages, including the initiation of mastication. Tamoxifen was first delivered to Axin2CreERT2/+;R26RmTmG/+ mice right before they were born (P0). This method initiates the recombination event that results in Wnt-responsive cells becoming GFP+ve (Yuan, Pei, Zhao, Li, et al. 2018; Yuan, Pei, Zhao, Tulu, et al. 2018). Wnt-responsive cells were examined at P1 and P15, when mice were still under suckling. At P1, the PDL space was narrow (Fig. 2A), and around 20% of the PDL cells were Wnt responsive (Fig. 2B). By P15, right before the mastication, this population was relatively stable (Fig. 2C, D, quantified in L). Mice began chewing hard foods at approximately P15. To “capture” those cells that became Wnt responsive following initiation of mastication, tamoxifen was delivered to Axin2CreERT2/+;R26RmTmG/+ mice for 3 d, P15 to P18. At P20, mice were sacrificed, and the histology of the PDL space and the distribution of Wnt-responsive cells was analyzed. At P20, the ECM in the PDL space had yet to become organized in response to mastication (Fig. 2E), and <10% of cells were Wnt responsive (Fig. 2F). In this subset of Wnt-responsive cells, however, the majority were proliferating (Fig. 2F′). By P30, when mice had been chewing hard food for ~2 wk, PDL fibers were oriented perpendicular to the direction of masticatory force (Fig. 2G), and progeny of the initial population of Wnt-responsive cells now occupied the entire PDL space (Fig. 2H, quantified in L). At this point, however, the majority of the Wnt-responsive progeny was no longer proliferating (Fig. 2H′). In adult mice, lineage tracing data suggested that the turnover of the Wnt-responsive cells are very slow (Fig. 2I–K, quantified in L).
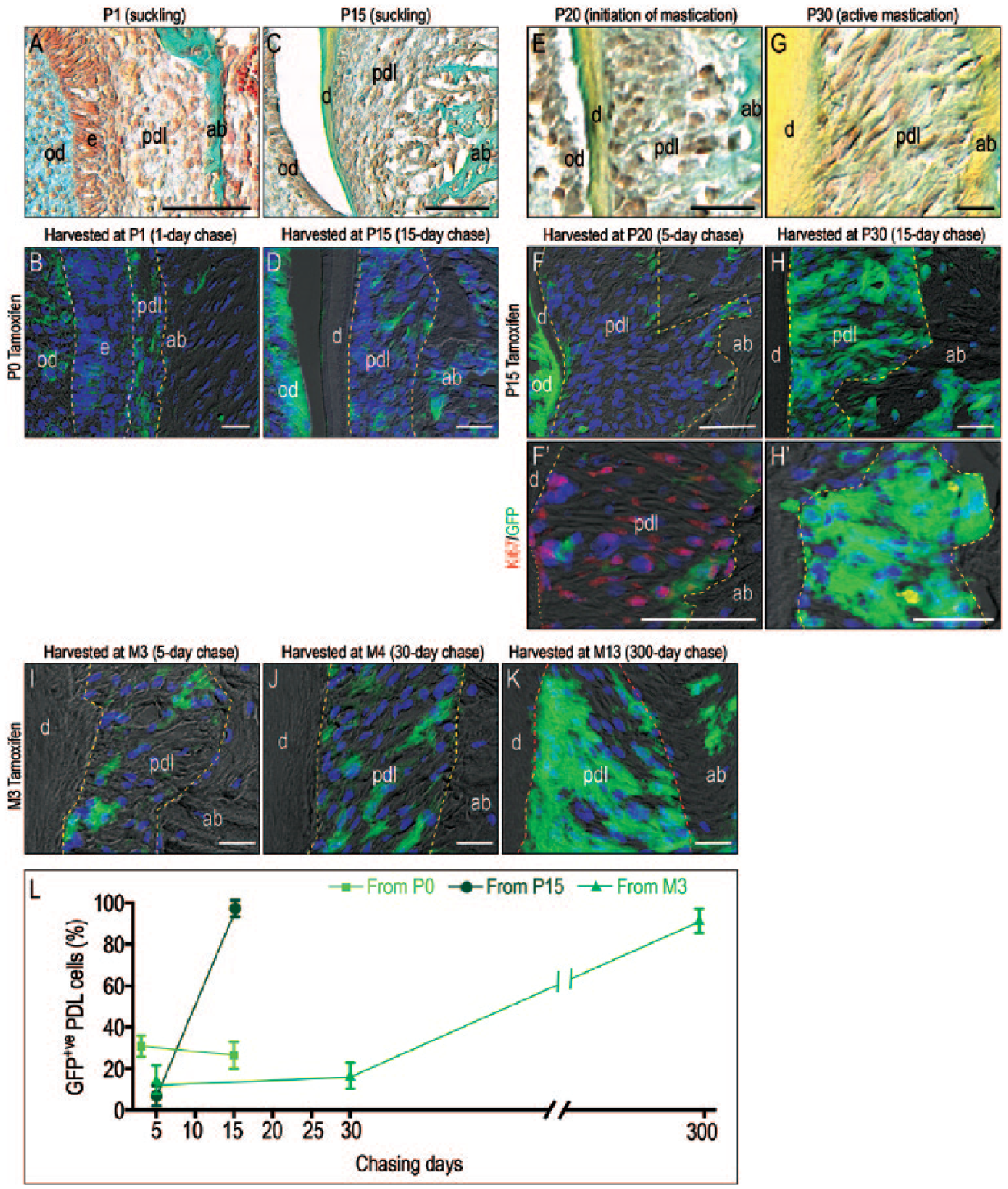
Mastication triggers the proliferation of Wnt-responsive cells that contribute to the functioning PDL. (
Thus, the transition from suckling to chewing hard food was accompanied by a burst in the proliferation of the Wnt-responsive progeny. Simultaneously, the PDL became organized into a functional tissue. Continued mastication, however, was not associated with continued Wnt pathway activation or with continued cell proliferation. The PDL maintained its functional organization but did so with very slow cell turnover.
Removing a Masticatory Load Results in the Loss of Wnt-Responsive Cells in the PDL
Thus far, our data were consistent with a model whereby loading of the PDL directly stimulates proliferation of a Wnt-responsive population in the PDL space, which is then responsible for the tissue’s functional adaptation to mastication. An alternative explanation, however, was that Wnt-responsive cells in the PDL expand in response to some other developmental event. To directly test whether mastication specifically triggered the proliferation of Wnt-responsive cells, we again used the lineage-tracing strain of mice.
Tamoxifen was delivered at P0 (Fig. 3A). We first ascertained the distribution of Wnt-responsive cells in the PDL space prior to tooth eruption. Wnt-responsive cells occupied the developing PDL space between the tooth root and the surrounding alveolar bone (Appendix Fig. 1A, A′). Coimmunostaining with cytokeratin 5 and GFP identified a subset of the Wnt-responsive progeny that was also positive for cytokeratin 5 (Appendix Fig. 1A′′). With continued root development, the population of Wnt-responsive cells did not increase in number (Appendix Fig. 1B, B′), but the cytokeratin 5/GFP population transitioned from a Hertwig’s epithelial root sheath morphology to a fragmented structure, reminiscent of the epithelial cell rests of Malassez (Appendix Fig. 1B′′).
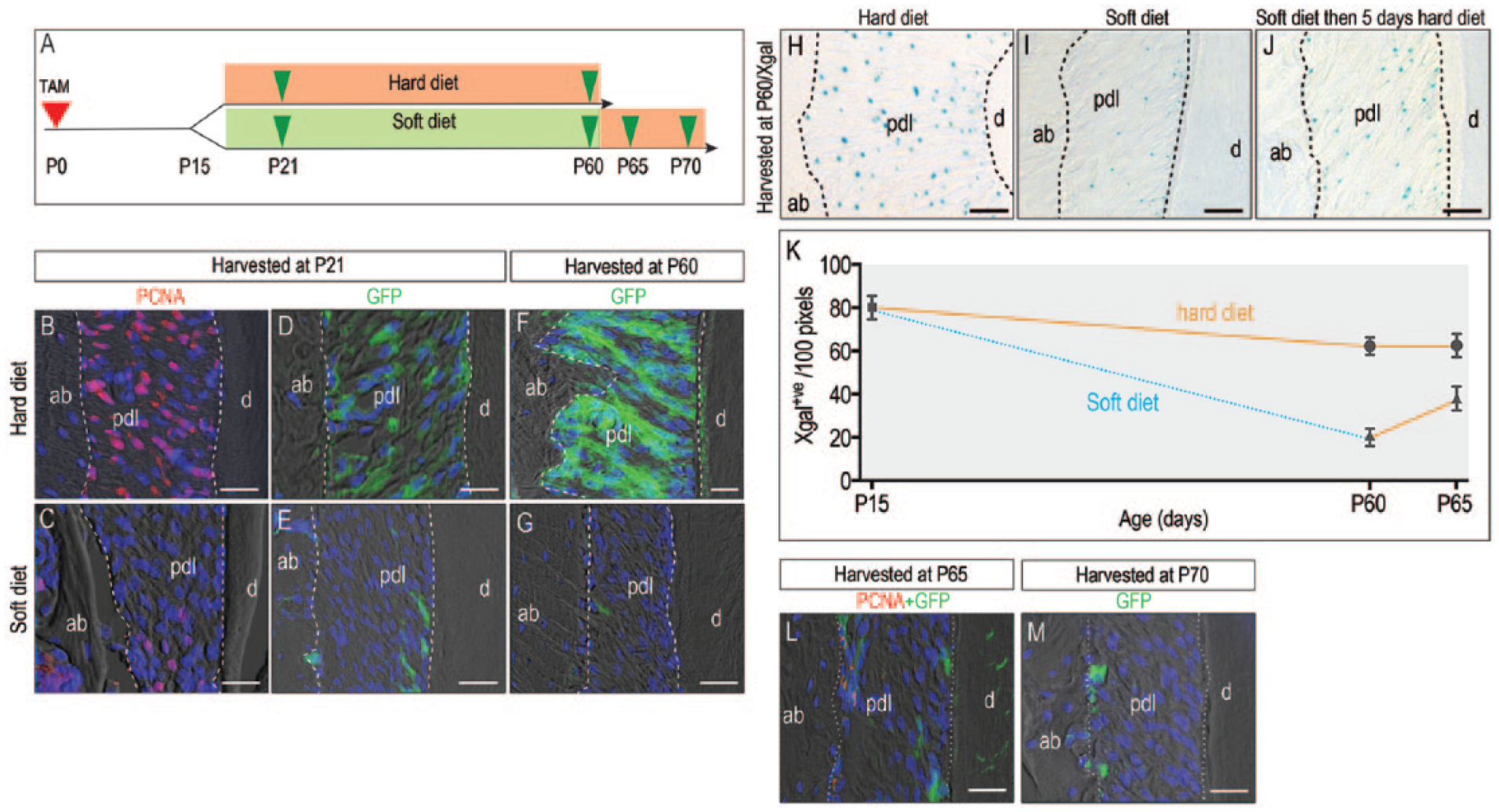
Wnt-responsive cells are lost when masticatory loading is eliminated. (
At P15, near the stage when mice are weaned, 2 experimental groups were formed according to diet: a normal hard food diet fed to one group and a soft food diet fed to the other group (Fig. 3A). When animals began chewing hard foods, PDL cells exhibited a proliferative burst (Fig. 3B); in those animals fed a soft diet, there was no proliferative burst (Fig. 3C).
The progeny of the initial Wnt-responsive population was then compared over time. After 6 d in the hard diet group, the proliferative burst resulted in a dramatic expansion of the Wnt-responsive population (Fig. 3D). In the soft diet group, the lack of a proliferative burst manifested as few Wnt-responsive cells in the PDL (Fig. 3E). These data demonstrated that it was masticatory force and not some other developmental event that was responsible for triggering the proliferation of Wnt-responsive cells in the PDL.
After 45 d in the hard diet group, Wnt-responsive progeny fully occupied the PDL (Fig. 3F). In the soft diet group, there were few Wnt-responsive cells in the PDL (Fig. 3G). The Wnt-responsive population coexpressed osterix, Runx2, and periostin (Appendix Fig. 2A–C). These data demonstrated that a hard diet was associated with a significantly larger population of these Wnt-responsive cells, which, based on the expression of these markers, appears to constitute a multipotential progenitor population capable of differentiating into a PDL fibroblast, an osteoblast, or a cementoblast.
How plastic is this Wnt-responsive population? We asked whether a change in diet could reactivate Wnt signaling, using Axin2 LacZ/+ reporter mice. Mice were fed a continuous hard diet (Fig. 3H), a continuous soft diet (Fig. 3I), or a soft diet followed by a hard diet (Fig. 3J). First, the distribution of Wnt-responsive cells was assessed by Xgal staining: the hard diet group had abundant Wnt-responsive progenitor cells occupying the PDL, while the soft diet group had minimal Wnt-responsive progenitors. In the group that had been shifted from a soft to a hard diet, the number of Wnt-responsive progenitor cells was increased relative to the soft diet control. Xgal+ve Wnt-responsive cells are quantified in Figure 3K. This result demonstrated that Wnt signaling changed in response to a change in the mechanical environment.
Did Axin2 descendants proliferate in response to the change in masticatory load associated with a transition from soft to hard food? To address this possibility, the lineage-chasing strain of Axin2CreERT2/+;R26RmTmG/+ mice was maintained on a soft diet for 45 d, as shown in Figure 3G, and then switched to hard diet for 5 and 10 d. Tissues were then analyzed with GFP and PCNA immunostaining, which revealed no obvious increase in the number of GFP+ve cells (Fig. 3L, M) and PCNA+ve cells (Fig. 3L). The lack of GFP/PCNA cells indicated that Axin2 descendants did not proliferate in response to the change in diet. Therefore, we conclude that the change to a hard diet clearly activated Wnt signaling (Fig. 3J), but the increase was not directly attributable to an increase in the mitotic activity of the original Axin2 descendants.
Mechanical Basis for Disuse Atrophy in the PDL
Thus far, our data showed that the Wnt pathway was activated by mechanical loading associated with a hard diet and that continued loading was required to maintain the Wnt-responsive progenitor cells in the PDL. Our data also suggested the ability of the PDL to rapidly respond to changes in the mechanical environment. These findings suggested that whatever factors were responsible for triggering Wnt signaling, they had to be dynamic in nature. Therefore, we considered how loading influenced the mechanical environment of the PDL.
A 3-dimensional volumetric rendering of the periodontium was produced from µCT data (Fig. 4A). The PDL (yellow, Fig. 4A′) was defined as the unmineralized space between the mineralized alveolar bone and tooth. A mesh was constructed from this 3D model (Fig. 4B), which was then imported into a finite element program (see Methods). Tensile and compressive strains were visualized in response to the load associated with a hard diet (Fig. 4C–C′′) and a soft diet (Fig. 4D–D′′). Both diets produced a similar spatial distribution of principal strains, but the peak strain values were ~10 times lower when the animal was fed a soft diet (compare Fig. 4C–C′′ with D–D′′).
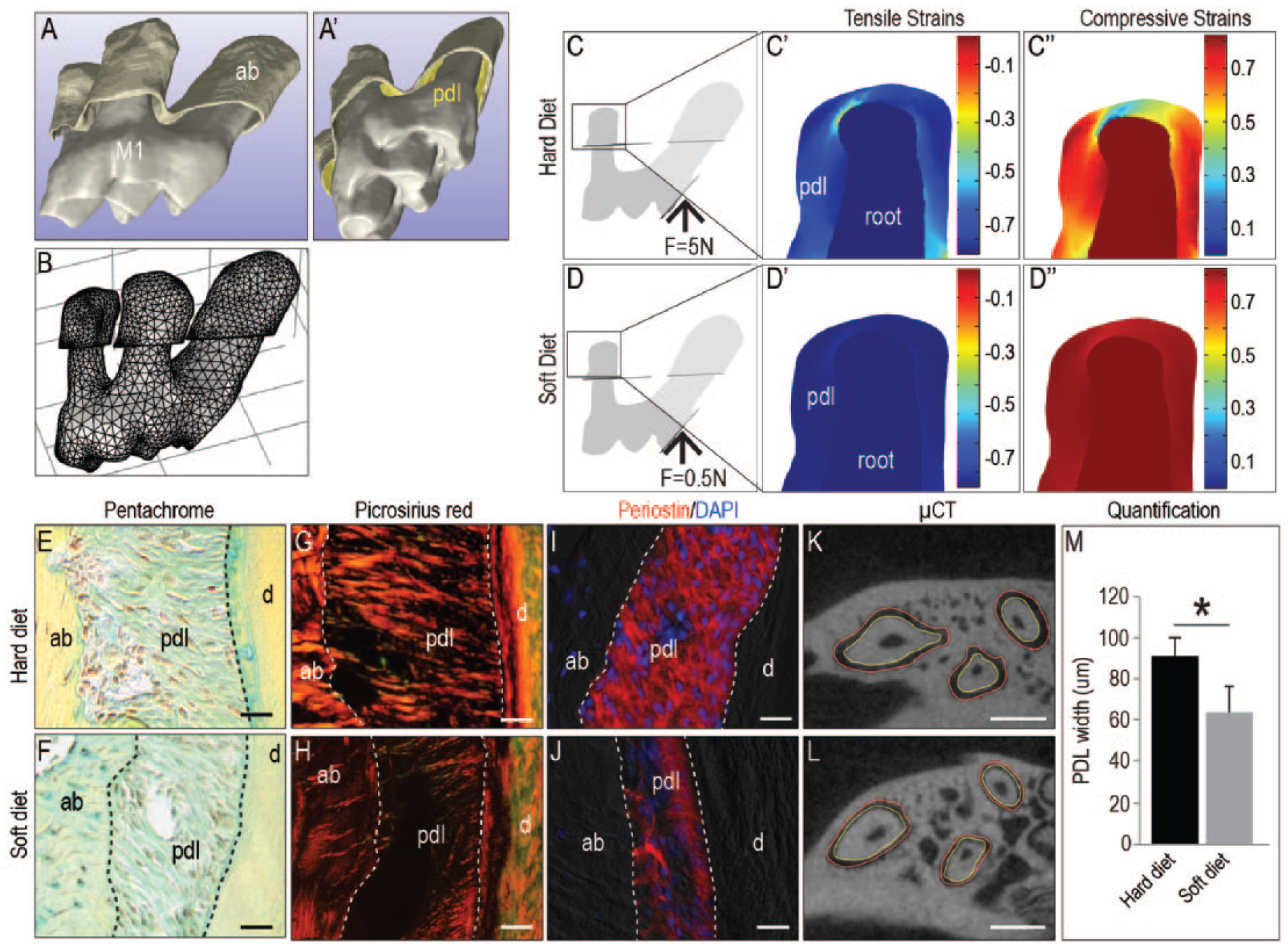
Underloading reduces tensile and compressive strains and results in PDL atrophy. (
Masticatory load maintains a level of tensile and compressive strains in the PDL, and these computational analyses suggest that a net strain condition is required for PDL homeostasis. The biological response to a decreased strain state was then examined. Although pentachrome staining did not reveal gross differences (compare Fig. 4E, F), picrosirius red staining did: When viewed under polarized light, picrosirius red staining identified the collagen-rich PDL in animals fed a hard diet (Fig. 4G). The collagen content in the PDL of animals fed a soft diet was significantly reduced (Fig. 4H). Periostin immunostaining was used to identify the PDL fibers; compared with that of animals fed a hard diet (Fig. 4I), the PDL width was reduced in animals fed a soft diet (Fig. 4J). Histomorphometric analyses of µCT data were employed to verify the change in PDL width at P60. From the edge of the alveolar bone (outlined in red, Fig. 4K, L) to the edge of the tooth surface (outlined in yellow), these analyses revealed a significant reduction in PDL width in animals on a soft diet (Fig. 4M). Collectively, these data demonstrate a functional link between 1) mastication-induced strain states in the PDL that trigger the proliferation of Wnt-responsive progenitor cells capable of generating PDL fibers and 2) the fibrous orientation of the PDL.
Discussion
Wnt-Responsive Cells Constitute an Essential Progenitor Population in the PDL
In this study, we demonstrated a causal relationship among the onset of masticatory function, the advent of Wnt signaling, and the organization of the PDL into a functional tissue (Figs. 1, 2). By taking advantage of the fact that the maxillary first molar erupts before mice have transitioned from suckling to chewing hard foods, we could observe the day-to-day changes in the organization of the PDL, and the reaction of Wnt-responsive PDL cells, to the first masticatory loads.
These analyses revealed a temporal correlation between the onset of masticatory function and the burst in proliferation of Wnt-responsive PDL cells (Figs. 1, 2). Simultaneously, the fibrous PDL became organized. Removing the masticatory force demonstrated a causal link between mastication and the burst in Wnt-responsive cell proliferation (Fig. 2): absent a hard diet, Wnt-responsive cells failed to become mitotically active and the PDL atrophied (Figs. 3, 4).
A Causal Relationship Exists among Mastication, the Proliferation of Wnt-Responsive Progenitors, and a Functional PDL
Two cellular events underlie PDL atrophy: proliferation in the PDL is eliminated, and Wnt signaling is nonexistent (Figs. 2, 3). The literature is replete with examples of how lack of loading leads to disuse atrophy of the skeleton (Jorgensen et al. 2000; LeBlanc et al. 2000; Takamoto et al. 2005; Tatara et al. 2014). An equally robust literature demonstrates that entheses, like the PDL, also undergo degeneration when subjected to disuse (Yasuda and Hayashi 1999; Wang et al. 2012; Kun-Darbois et al. 2015; Leung et al. 2015). These data imply that loading exerts a positive influence that maintains the collagenous ECM in an enthesis, which in turn ensures that the enthesis can function. Our data implicate Wnt signaling as the basis for this atrophy, as well as the recovery of the PDL in response to loading.
Why does the PDL narrow in response to reduced loads? We can speculate about the answer to this question by considering finite element data. It appears that under normal conditions, masticatory loads induce a level of stress and strain in the PDL that are required for its homeostasis (Fig. 4). When the masticatory load is reduced, the magnitudes of the stress and strain are also reduced. By changing the geometry of the PDL—for example, by narrowing the space—the homeostatic levels of stresses and strains are reestablished in the PDL. Thus, one can view the narrowing or atrophy of the PDL as a compensatory mechanism to maintain stress and strain levels in a physiologic (homeostatic) range. Similar changes are also observed in other ligamentous structures in response to reduced loading (Leigh et al. 2008). We speculate that the adaptive ability of the PDL relates to the Wnt-responsive population occupying this tissue. For example, when the PDL senses a force, nerve fibers within the tissue release calcitonin gene–related peptide (CGRP) and substance P (Hall et al. 2001; Jiang et al. 2016). It has been speculated that CGRP and substance P serve as vasodilators to stimulate plasma extravasation and leukocyte migration. Both CGRP (Mei et al. 2014; Zhou et al. 2016) and substance P (Mei et al. 2014) can activate Wnt signaling. Therefore, it is possible that masticatory forces are required to maintain CGRP and substance P, which in turn maintain Wnt-responsive cells in PDL space.
Conclusions
Mastication-induced strain maintains the PDL fibers (Fig. 4). In our mouse model, soft diet causes PDL atrophy, and this atrophy can be reversed when the animal is switched back to a hard food diet (Fig. 3). We demonstrated that a component of this adaptive response is dependent on a Wnt-responsive population identified in the PDL (Figs. 1–3). As this population diminishes in size with age (Fig. 1), the ability of the PDL to adapt to larger forces may decline as well. This hypothesis is currently being tested in our laboratory.
Author Contributions
X. Zhang, X. Yuan, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; Q. Xu, M. Arioka, contributed to data acquisition, analysis, and interpretation, critically revised the manuscript; L.A. Van Brunt, Y. Shi, contributed to data acquisition and analysis, critically revised the manuscript; J. Brunski, contributed to data analysis and interpretation, critically revised the manuscript; J.A. Helms, contributed to conception, design, data analysis, and interpretation, drafted and critically revised the manuscript.
Supplemental Material
DS_10.1177_0022034518817305 – Supplemental material for Molecular Basis for Periodontal Ligament Adaptation to In Vivo Loading
Supplemental material, DS_10.1177_0022034518817305 for Molecular Basis for Periodontal Ligament Adaptation to In Vivo Loading by X. Zhang, X. Yuan, Q. Xu, M. Arioka, L.A. Van Brunt, Y. Shi, J. Brunski and J.A. Helms in Journal of Dental Research
Footnotes
Acknowledgements
We thank Serdar Ustun Tulu for his contributions to generating the 3-dimensional .stl file representing the molar tooth used in the finite element model.
A supplemental appendix to this article is available online.
This work was supported by the National Institutes of Health (grant R01DE02400014) to J.A.H. and J.B. and by the China Scholarship Council (grant 201706245019) and West China Hospital of Stomatology (grant 2015-18) to X.Z.
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.