
Abstract
Oral mucosal homeostasis is achieved by complex immunologic mechanisms, orchestrating host immunity to adapt to the physiologic functions of the various specialized niches in the oral cavity. Dental implants introduce a novel mucosal niche to the immune system to deal with. Nevertheless, the immune mechanisms engaged toward implants and whether they have broader effects are not well defined. Using a murine model, we found an accumulation of neutrophils and RANKL-expressing T and B lymphocytes in the implant-surrounding mucosa, accompanied by local bone loss. Surprisingly, the presence of implants had an impact on remote periodontal sites, as elevated inflammation and accelerated bone loss were detected in intact distant teeth. This was due to microbial dysbiosis induced by the implants, since antibiotic treatment prevented bone loss around teeth. However, antibiotic treatment failed to prevent the loss of implant-supporting bone, highlighting the distinct mechanisms mediating bone loss at each site. Further analysis revealed that implants induced chronic lymphocyte activation and increased mRNA expression of IFN-α and accumulation of IFN-α–producing plasmacytoid dendritic cells, which we previously reported as bone-destructive immune responses. Collectively, this study demonstrates that implants have a strong and broad impact on oral mucosal homeostasis, inducing periodontal bone loss in a niche-specific manner that is both microbiota dependent and independent.
Keywords
Introduction
The oral cavity is a complex ecosystem involving constant host-bacteria interactions that shape local immune responses to maintain mucosal homeostasis (Hajishengallis et al. 2011). This delicate balance is often dysregulated, resulting in uncontrolled immune responses that could lead to oral and systemic pathologies (Hovav 2014; Hajishengallis et al. 2016). Periodontitis represents one of the most common inflammatory diseases that culminate in the destruction of the periodontal apparatus (Armitage 1996). It has been established that bacterial plaque is the main etiologic factor mediating excessive immune responses and, consequently, periodontal bone destruction (Van Dyke et al. 1993). Furthermore, a recent study reported that under physiologic conditions, ongoing damage to the oral epithelium due to masticatory forces facilitates local expansion of Th17 cells and induces alveolar bone destruction in a microbiota-independent manner (Dutzan et al. 2017). The importance of the oral epithelium in regulating mucosal homeostasis and bone loss was also demonstrated in mice lacking the protein growth arrest–specific 6 (GAS6; Nassar et al. 2017). Expression of GAS6 is upregulated in the oral epithelium postnatally by the microbiota, where it acts as a central regulator of host-bacteria interplay. These observations suggest that alveolar bone loss around teeth can be mediated by various mechanisms under physiologic or inflammatory conditions.
Peri-implantitis is an additional oral inflammatory disease involving bone destruction around dental implants (Berglundh et al. 2011; Berglundh et al. 2018) and is considered a major global health concern (Derks and Tomasi 2015). Although the primary etiology of this disease associates with the microbiota, we have shown that titanium implants by themselves can elevate the local inflammatory milieu and dysregulate mucosal homeostasis. This effect is mediated by titanium ions released from the implants, which impair the development of Langerhans cells that function as regulators of oral mucosal immunity (Heyman et al. 2018). Nevertheless, despite the assumption that peri-implantitis and periodontitis share etiologic factors and hence are treated similarly, periodontitis generally responds well to bacterial plaque control while peri-implantitis is much less responsive (Renvert et al. 2008). Moreover, whereas antibiotic treatment is beneficial in the treatment of periodontitis, its efficacy in peri-implantitis is debatable (Javed et al. 2013; Romanos et al. 2015). This suggests that although the clinical signs of both diseases are similar, the immune mechanisms and the role of microbiota in each disease are likely to be different.
Using a murine model of dental implants, we found in the current study that besides increasing local inflammation and bone loss, implants are capable to induce oral microbial dysbiosis and enhance inflammation and bone loss in remote teeth. This enabled us to dissect the mechanisms mediating bone loss around implants and teeth under physiologic conditions. Our data suggest that bone loss in each niche involves distinct immunologic and microbial mechanisms, highlighting the specificity and complexity of the oral immune system.
Materials and Methods
Mice
Female BALB/c mice (4 to 5 wk old) were purchased from Envigo. The animals were housed in the specific pathogen–free unit. All experimental procedures were reviewed and approved by the Institutional Animal Care and Use Committee of the Hebrew University-Hadassah Medical Center and followed the ARRIVE guidelines.
Extractions and Implant Placement
The model design and surgery protocols used for this study were previously described (Heyman et al. 2018). For details, see Appendix.
Isolation and Processing the Gingiva and Peri-implant Mucosal Tissues
The mucosal tissues were processed as previously reported (Heyman et al. 2018) and are described in the Appendix.
RNA Extraction and Quantitative Real-time Polymerase Chain Reaction
RNA was isolated and processed via quantitative real-time polymerase chain reaction (RT-qPCR; see Appendix).
Antibodies and Reagents
Antibodies and reagents are described in the Appendix.
Micro–computed tomography Analysis
For micro–computed tomography scans and bone analysis for teeth and implants, see Appendix.
Cytokine Secretion by Cultured Splenocytes
The supernatants of cultured splenocytes were analyzed by ELISA (see Appendix).
Cultivation of Oral Microbiota
The oral cavity of each mouse was swabbed for 30 s, and the samples were plated as outlined in the Appendix.
Microbiome Sequencing and Analysis
Oral bacterial DNA was extracted and used for Illumina sequencing, as described in the Appendix, with data analysis and diversity-related statistical tests.
Broad-Spectrum Antibiotic Model
Four antibiotics and an antifungal treatment were administrated to the mice. For details, see Appendix.
Statistical Analysis
In each experiment, the mice were independently tested and data expressed as mean ± SEM. Statistical tests were performed with the Student t test. P < 0.05 was considered significant.
Results
Dental Implants Promote Bone Loss around Remote Teeth
We have previously shown that titanium dental implants impair local development of Langerhans cells, resulting in elevated inflammation at the implant vicinity (Heyman et al. 2018). To examine if the presence of such dysregulated local immunity affects the implant-supporting bone, we evaluated bone volume at various time points after implant placement using micro–computed tomography (Fig. 1A). As demonstrated in Figure 1B and C, we detected gradual bone loss at 4 and 10 wk after implant placement as compared with 2-wk controls. We then asked whether the implants were promoting alveolar bone loss around remote teeth. For this, we assessed the bone around the contralateral teeth 10 wk after implant placement; as a control, the same area in naïve mice was examined (Fig. 1D, E). Indeed, micro–computed tomography analysis revealed a significant reduction in bone volume around the contralateral teeth of implanted mice as compared with the teeth of nonimplanted mice. Since receptor activator of NF-κB ligand (RANKL) and its antagonist osteoprotegerin (OPG) are key regulators of pathophysiologic bone remodeling (Grimaud et al. 2003), we calculated the ratio of RANKL and OPG mRNA levels using RT-qPCR in the tissue. In concurrence with the bone loss results, the highest RANKL:OPG ratio was found around implants (peri-implant mucosa), whereas in the contralateral teeth (gingiva), the ratio was lower but still significantly higher than the teeth of naïve mice (control; Fig. 1F). Collectively, these findings suggest that dental implants undergo spontaneous bone loss with time. Moreover, the presence of the implant accelerates alveolar bone loss around remote teeth.
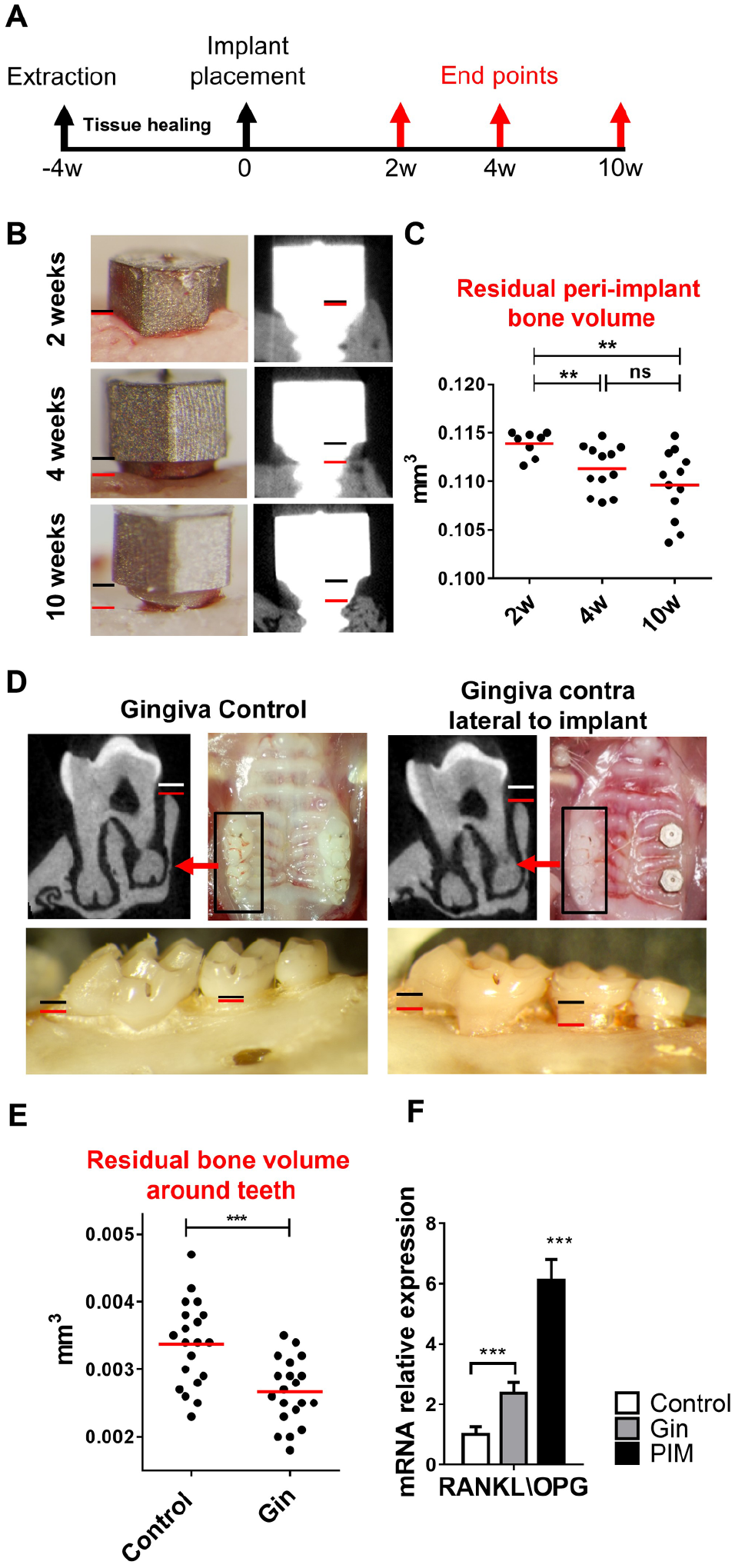
Bone resorption around implants and contralateral teeth following implant placement. (
Implants Dysregulate Immune Homeostasis around Remote Teeth
We next characterized gingival leukocyte subpopulations that are known to regulate periodontal bone remodeling (Mizraji et al. 2017). The gating strategy is provided in Appendix Figure 1A. Four weeks after implant placement, total leukocytes (CD45+) were significantly elevated around implants (peri-implant mucosa) and, to a lesser extent, in the contralateral teeth (gingiva) in comparison with gingiva of naïve mice (control; Fig. 2A). The same pattern was observed in the lymphoid lineage, with higher frequencies of T cells (CD3+), B cells (B220+), T-helper cells (CD3+CD4+), and T-regulatory cells (CD3+CD4+FoxP3+) in the implant and contralateral teeth as compared with naïve control (Fig. 2B, C). Since expression of RANKL on lymphocytes was reported to affect osteoclastogenesis and facilitate bone loss (Chen et al. 2014), we evaluated its expression on T and B cells. Concurring with our earlier results regarding RANKL/OPG mRNA expression (Fig. 1F), RANKL expression on gingival CD4+ T cells was elevated in implant and contralateral teeth; nevertheless, RANKL-expressing B cells increased only around implants (Fig. 2C). With regard to myeloid cells, neutrophils and Ly6Chi and Ly6Clo monocytes present at higher frequencies only around implants (Fig. 2D). To further examine the immunologic state of the mentioned tissues, we quantified by RT-qPCR the expression of certain immunologic genes 4 wk after implant placement. As depicted in Figure 2E, the expression of IFN-γ, IFN-α, and IL-1β was significantly upregulated in the peri-implant mucosa and contralateral teeth as compared with naïve mice. IL-17A and IFN-β expression, however, was elevated only in the peri-implant mucosa. Of note, no differences were found in the expression of genes involved in the recruitment of inflammatory cells, such as TNF-α, VCAM-1, ICAM-1, and P/E-selectins (Appendix Fig. 1B). Taken together, implant placement results in a sustained high-leukocyte infiltration not only locally but at distant gingival sites, representing a modified homoeostatic state rather than acute inflammation. Furthermore, the differences in leukocyte subpopulations among implants and contralateral teeth suggest the development of distinct immune responses at each site that might regulate bone loss differently.
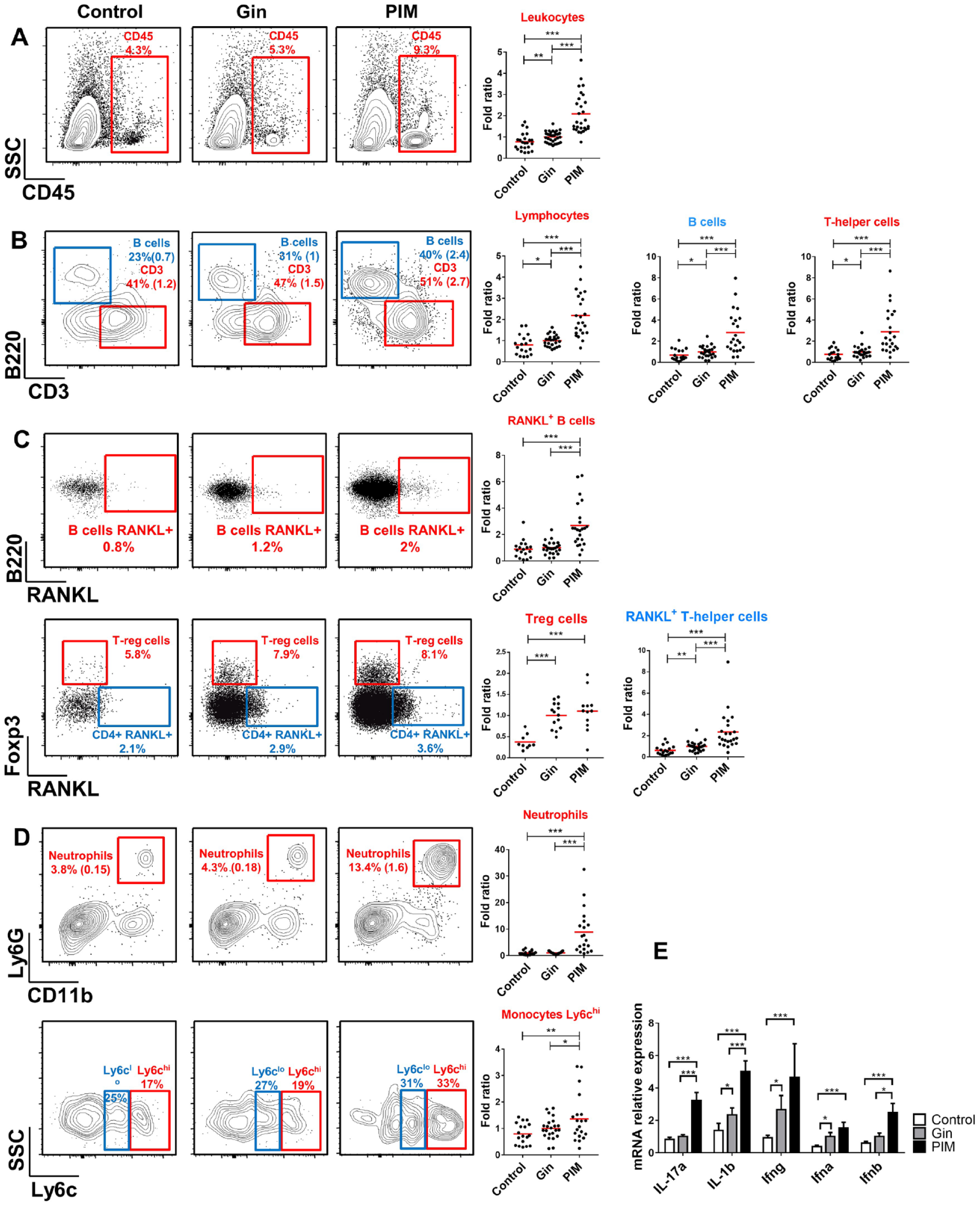
Implant placement dysregulated immune homeostasis in the tissue surrounding the implant and the contralateral teeth. Different immune cell populations were quantified in the gingiva of nonimplanted mice (control), contralateral gingiva of implanted mice (Gin), and peri-implant mucosa (PIM) per flow cytometry (FACS). Representative FACS plots show the frequencies of (
Ongoing Elevated Levels of Plasmacytoid Dendritic Cells and Cytokine Production in Implanted Mice
The presence of dysregulated gingival immunity several weeks after implant placement may imply that the implants induce prolonged activation of the immune system. To address this point, we collected the gingiva-draining lymph nodes (LNs) of naïve and implanted mice for analysis of dendritic cells (DCs). Appendix Figure 2 depicts our gating strategy employed to identify different DCs in the LNs. Increased frequencies of total DCs (CD11c+MHCII+) were found in the LNs of implanted mice as compared with naïve control (Fig. 3A). This elevation was mainly attributed to accumulation of plasmacytoid dendritic cells (pDCs; MHCIIloCD11cintCD11b−B220+PDCA1+Siglec-H+) rather than migratory DCs (CD11cintMHCIIhi) arriving from the gingiva. However, further analysis of pDCs revealed that in implanted mice, a larger fraction of these cells was CD4 negative, a phenotype previously reported to represent migratory pDCs with a capacity to secrete IFN-α (Yang et al. 2005). Indeed, using quantitative RT-qPCR, we were able to detect upregulated and sustained expression of IFN-α in the LNs of implanted mice as compared with the LNs of naïve mice (Fig. 3B). On this regard, we recently reported that elevated IFN-α production in the draining LNs induces chronic activation of T cells that facilitate bone loss (Mizraji et al. 2017). To examine whether such chronic activation is taking place also upon implant placement, we cultured splenocytes from 4-wk implanted mice with no external stimulation and measured spontaneous cytokine secretion using ELISA. Indeed, higher levels of IFN-γ and IL-10 were found in the supernatants of implanted mice as compared with naïve control, indicating that splenocytes of implanted mice are chronically activated (Fig. 3C). These findings suggest that production of IFN-α by pDCs play a role in dysregulating immunity in implanted mice via ongoing activation of the gingiva and development of bone-destructive immune responses.
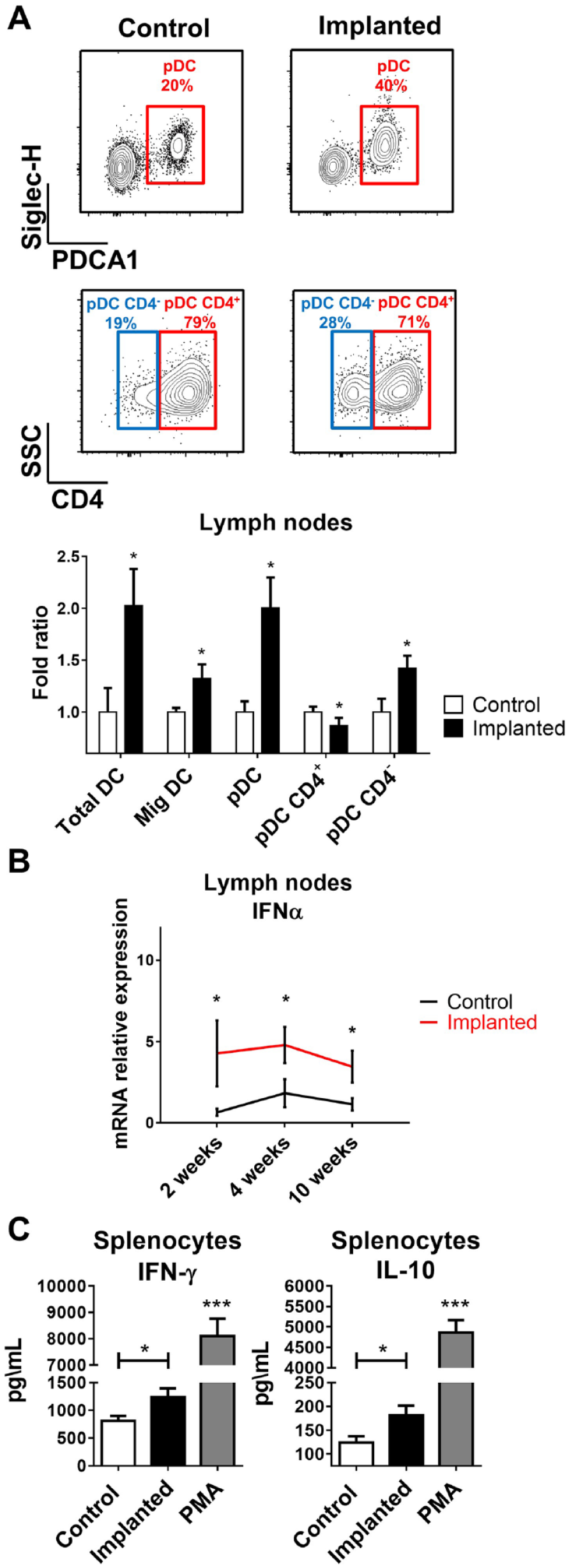
Increased proportions of plasmacytoid dendritic cells (pDCs) and cytokine production in the lymph nodes and spleen of implanted compared with nonimplanted mice (Control). (
Implant Placement Results in Oral Microbial Dysbiosis
Considerable alteration of oral mucosal immunity is known to cause oral microbial dysbiosis that is often associated with pathologic consequences (Hajishengallis et al. 2011; Nassar et al. 2017). We thus examined if dysregulation of gingival immune responses by implants is also capable of altering the oral microbiota. For this, we sampled oral microbiota from the same mice before implant placement and 4 wk after. As demonstrated in Figure 4A, the levels of cultivated anaerobic and aerobic bacteria were significantly higher in implanted mice as compared with the level that they had prior to implant placement. Further analysis revealed that following implant placement, the mice had a significant increase in the diversity of oral microbiota (Fig. 4B), which also varied significantly from the microbiota that they harbored prior to implant placement (Fig. 4C). Detailed taxonomic analysis showed that while preimplanted mice were primarily colonized by the bacterial phylum Firmicutes, implant placement facilitates the expansion of Proteobacteria and Bacteroidetes on the expense of Firmicutes (Fig. 4D). On a family level, implant placement decreases the relative abundance of Streptococcaceae by 2-fold, whereas other bacteria families were expanded, such as S24-7 (Bacteroidetes), Pasteurellaceae (Proteobacteria), Clostridiales-2 (Firmicutes), Enterobacteriaceae (Proteobacteria), Bacteroidaceae (Bacteroidetes) Lactobacill-aceae (Firmicutes), and more (Fig. 4E). Of note, the relative proportions of known periodontal pathogenic families, such as Prevotellaceae (Bacteroidetes), Fusobacteriaceae (Fusobacteria), and Porphyromonadaceae (Bacteroidetes), were increased after implant insertion (Fig. 4E). To understand more accurately the impact of immune dysregulation in the tissue around implants and contralateral teeth on local oral microbiota, we sampled the microbiota specifically in these sites using small microbrushes under full anesthesia and tongue retraction. Absolute abundance of key bacterial families was then analyzed by RT-qPCR of the 16S gene. Following a second taxonomic analysis, we were able to demonstrate similar changes in the microbiome of the contralateral teeth and implant groups after implant insertion (Appendix Fig. 3A). Concurring with these results, Prevotellaceae and Lactobacillaceae were increased while Streptococcaceae decreased around implants and the contralateral teeth in comparison with samples taken in an identical manner from preimplanted mice (Fig. 4F, Appendix Fig. 3B). It should be also mentioned that in correlation to the intensity of the immune dysregulation, the absolute abundance of Prevotellaceae and Streptococcaceae around implants was significantly different than that observed in the contralateral teeth. Interestingly, the changes in microbial diversity observed in implanted mice resembled microbial dysregulation observed in the oral cavity of mice lacking GAS6, a key regulator of oral mucosal homeostasis (Nassar et al. 2017). In line with this notion, we found significant reduction of GAS6 expression in the gingiva surrounding the implants and, to a lesser extent, at the contralateral gingiva as compared with the gingiva of the same mice prior to implant placement (Fig. 4G). Since GAS6-madiated effect is influenced by the ability of the expended bacteria to utilize reactive oxygen species generated under inflammatory conditions for anaerobic respiration, we also measured the expression of reactive oxygen species–producing genes iNOS and DUOX2. Indeed, the expression of both genes was upregulated in the gingiva of implanted mice and contralateral gingiva (Fig. 4H). Finally, we asked whether the oral dysbiosis induced by implants has any impact on gut microbiota. As depicted in Appendix Figure 3C, 10 wk after implant placement, considerable alteration in the diversity of the gut microbiota was detected. In conclusion, implant placement leads to expansion of bacteria that are known to expand under inflammatory conditions, while the magnitude of dysbiosis correlates to the level of inflammation induced by the implant locally or remotely. This process is likely to be mediated by the capability of the implants to activate the oral mucosa via downregulation of GAS6 expression. Additionally, the capacity of oral implants to alter the gut microbiota over time demonstrates their vast systemic influence on the host microbial and immune system.
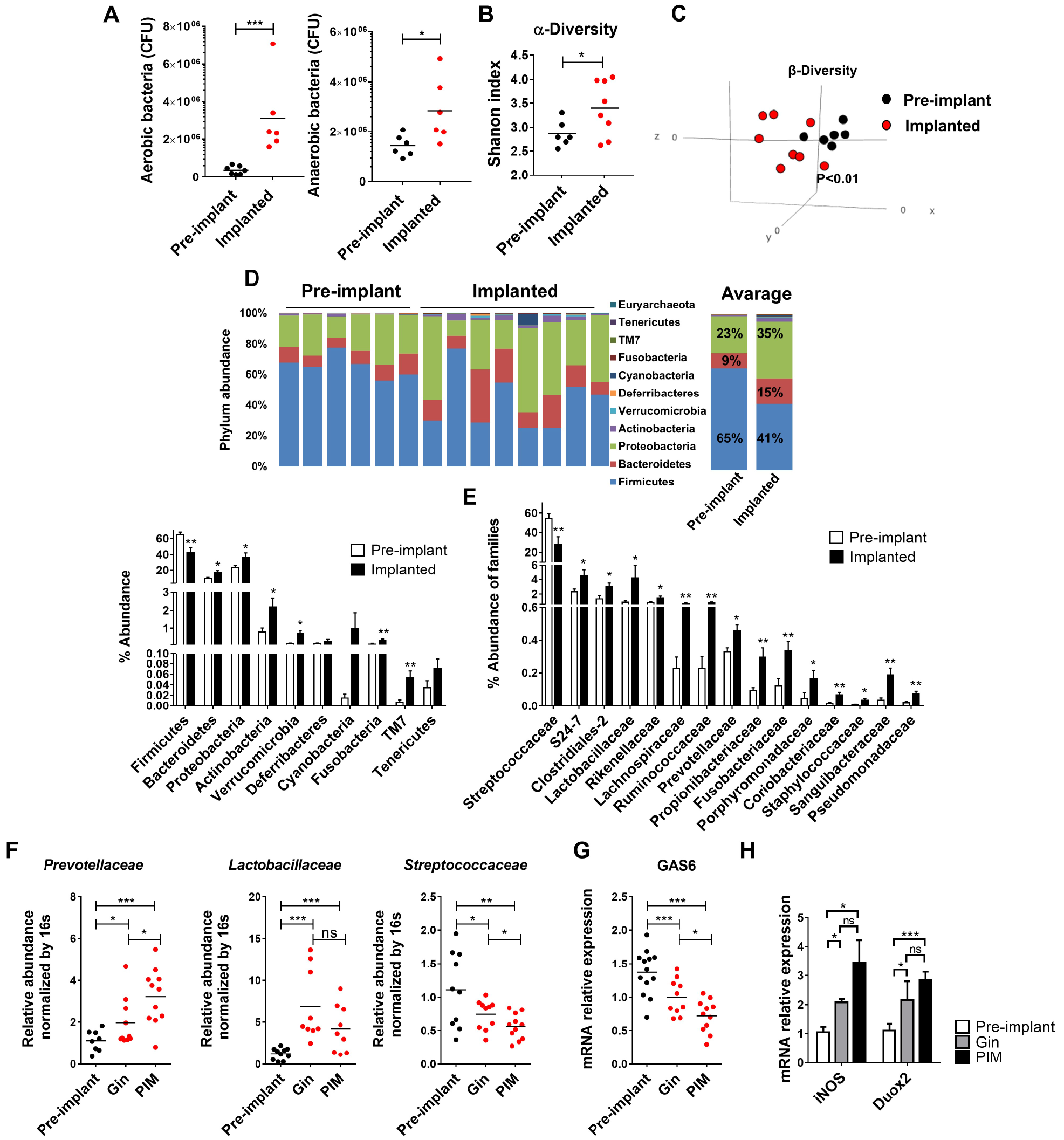
Bacterial dysbiosis in the oral cavity following implant placement. (
Bone Loss around Implants but Not Contralateral Teeth Is Microbiota Independent
To dissect the role of microbiota during periodontal bone loss in implanted mice, we administered the mice with broad-spectrum antibiotics and antifungal treatment before and after implant placement, as illustrated in Figure 5A. Examination of oral microbiota during the treatment confirmed a massive reduction in the bacterial load, and the remaining bacterial population displayed significant variation from the original microbiota (Appendix Fig. 4A–F). We next quantified residual bone level around implants and revealed that the reduction in the microbiota had no impact on bone destruction, since treated mice lost implant-supporting bone similar to untreated mice (Fig. 5B). On a contrary, alveolar bone loss in the contralateral teeth was significantly inhibited by the treatment, highlighting the central role of the microbiota in this process (Fig. 5C). As these results imply that local immunity plays a critical role in bone loss around implants, we assessed implant immune responses during the treatment. Total leukocytes as well as B and CD4+ T lymphocytes, expressing RANKL or not, were considerably reduced due to the treatment (Fig. 5D). Nevertheless, the percentage of neutrophils was not altered by the treatment. Evaluation of DCs in gingiva-draining LNs showed a reduction of migratory DCs upon antibiotic treatment, whereas CD4− pDCs retained their high percentages in implanted mice (Fig. 5E). Spontaneous secretion of IFN-γ levels by splenocytes was not affected by the antibiotic treatment, indicating that chronic activation of T cells in implanted mice is microbiota independent (Fig. 5F). In summary, although implants and contralateral teeth have dysregulated immunity and microbiota, the mechanisms mediating bone loss at each site are distinct. Whereas bone loss in contralateral teeth is microbiota dependent, around implants this process is microbiota independent and likely to involve chronic immune activation mediated by IFN-α secreting pDCs.
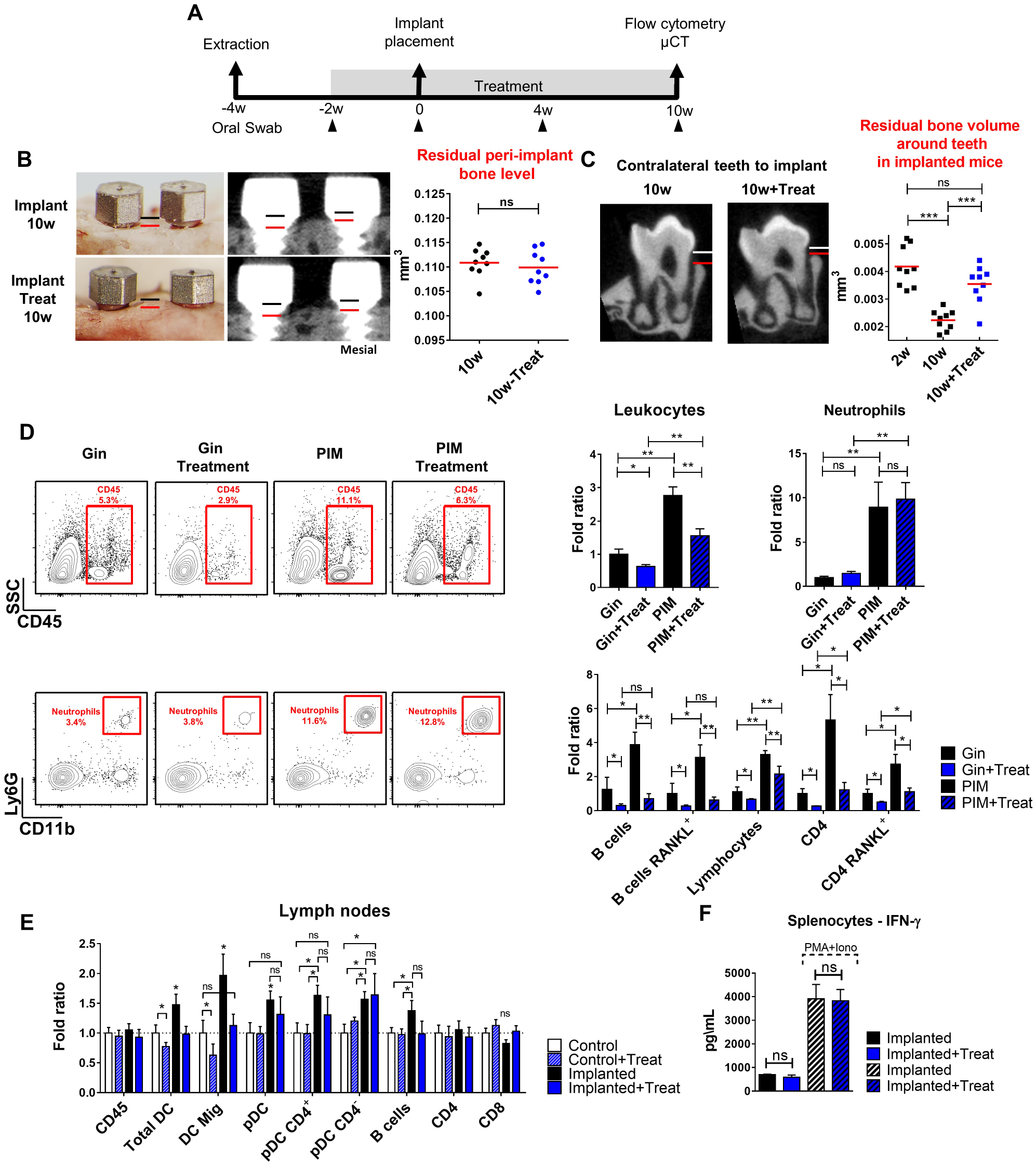
Bone loss around implants but not contralateral teeth is microbiota independent. (
Discussion
This study expands our previous observations demonstrating that, besides dysregulating local immunity (Heyman et al. 2018), dental implants have a broader influence on oral mucosal immunity. The data demonstrate that implant-associated immune responses dysregulate oral microbiota, which in turn alters gingival immunity at distant sites and facilitates alveolar bone loss. Such remote bone loss, however, involves distinct mechanisms than bone loss around implants, as it is induced by the microbiota and can be prevented by antibiotic treatment. Destruction of alveolar bone due to a dysbiotic microbiome was reported in experimental and human periodontitis and was mediated by expansion of IL-17–producing T cells (Th17) and recruitment of neutrophils (Dutzan et al. 2018). A study further reported that such Th17 cells are in fact FoxP3+ T cells that were converted locally to Th17 and thus termed exFoxP3Th17 cells (Tsukasaki and Takayanagi 2019). This, however, does not seem to be the mechanism of remote bone loss in the present study, since 1) IL-17 expression and neutrophils were not elevated in contralateral gingiva and 2) Foxp3+ T cells were elevated. This is also the case of implant-specific bone loss, because IL-17 and neutrophils that were considerably increased around implants led to a dysbiotic microbiota and not vice versa. Therefore, bone loss induced by dental implants, either locally or remotely, is mediated by an alternative mechanism that differs from periodontitis-associated bone destruction.
A possible mechanism to implant-associated bone loss could be the prolonged secretion of type I interferons (e.g., IFN-α and IFN-β), which was previously shown to induce chronic inflammation and tissue destruction (Wilson et al. 2013). Furthermore, using a model of repetitive infections with Porphyromonas gingivalis, we previously reported protracted and high levels of pDCs and type I interferons, which resulted in elevated T-cell activation, high RANKL expression, and alveolar bone loss (Mizraji et al. 2017). This pathologic process was reversed by depletion of type I interferons, demonstrating the deleterious impact of unrestrained expression of these cytokines on oral immunity. Accordingly, the durable high levels of pDCs and IFN-α/β in the LNs and the upregulation of gingival IFN-α/β in implanted mice might explain the chronic activation of T and B cells and their elevated RANKL expression that facilitate bone loss around implants. Elevation of RANKL-expressing T cells was also detected in the contralateral gingiva as well as upregulation of IFN-γ mRNA levels. This raises the possibility that excessive Th1 responses rather than Th17 cells mediate bone loss in remote sites as previously suggested (Teng et al. 2005; Arizon et al. 2012; Mizraji et al. 2018). Nonetheless, gingival IFN-α/β levels were shown to be negatively regulated by GAS6 expressed in epithelial cells (Nassar et al. 2017), and in agreement with this notion, reduced expression of GAS6 was detected in the gingiva of implanted mice. It is worth mentioning that activation of pDCs could be mediated by the reported capacity of particles and titanium ions released by the implant to dysregulate the oral microbiota (Souza et al. 2020). This might lead to changes in the accessibility of bacterial DNA that can be sensed by TLR9 in pDCs and induce secretion of type I interferons (Colonna et al. 2004).
The nature of oral microbiota is known to reflect the immunologic status of the oral mucosa (Hajishengallis et al. 2012; Costalonga and Herzberg 2014). Indeed, we found in implanted mice an expansion of oral bacteria that are capable to utilize inflammation by-products for anaerobic respiration (Winter et al. 2013; Winter and Baumler 2014). In fact, the dysbiosis observed in implanted mice resembles that found in Gas6−/– mice, further supporting a role to the GAS6–type I interferon axis in our system (Nassar et al. 2017). Detailed taxonomic analysis also revealed that microbial dysregulation is niche specific, as the predominant changes in the microbiota were found around implants as compared with remote gingiva. Nevertheless, the relatively moderate microbial changes observed in remote sites were sufficient to accelerate alveolar bone loss, highlighting the delicate equilibrium of bone remodeling in the periodontium. It is not completely clear yet which mechanisms are induced in remote sites by the microbiota that facilitate bone loss. However, based on cytokine expression in the gingiva and lymphocyte infiltration, it is likely that Th1 immune responses represented by IFN-γ are involved in this process rather than Th17 responses. Interestingly, besides altering the oral microbiota, dental implants induce microbial dysbiosis in the gut. This is in line with a previous study demonstrating that microbial dysbiosis initiated by periodontal pathogens is capable of dysregulating gut microbiota (Nakajima et al. 2015), highlighting the systemic impact of the oral microbiota in the body.
The higher accumulation of leukocytes and expression of proinflammatory cytokines around implants as compared with normal teeth are in concurrence with human studies (Nowzari et al. 2008; Nowzari et al. 2012; Yaghobee et al. 2014; Gurlek et al. 2017; Obadan et al. 2018). Of note, these studies showed aggressive immune responses, quick uncontrolled tissue destruction, and bone loss around implants as compared with teeth (Salvi et al. 2017), similar to the situation that we found in mice. While the aim of this study was to address the role of implant placement on oral mucosal homeostasis and bone loss, the distinct immunologic mechanisms induced around implants and remote teeth might shed light on the different responsiveness of human periodontitis and peri-implantitis to treatment (Renvert et al. 2008). It can be assumed that peri-implantitis, in contrast to periodontitis, is not responding well to mechanical and antibiotic treatments because bone loss taking place around implants is not entirely microbiota dependent. While our results are based on experiments performed in a murine model and should be cautiously interpreted to humans, they may have clinical implications suggesting that bone loss in human peri-implantitis is likely to be mediated by 2 factors: first, bacterial infection that can be treated with antibiotic; second, host destructive immunity against the implant that is microbiota independent and thus cannot be prevented by antibiotic or plaque control. As such, future treatment against peri-implantitis might include immunotherapeutic approaches in addition to the regular treatment. Our study suggests that such approaches could target type I interferon signaling that is likely to be involved in implant bone loss in a microbiota-independent manner.
Author Contributions
O. Heyman, contributed to conception, data acquisition, and analysis, drafted and critically revised the manuscript; Y. Horev, N. Koren, O. Barel, I. Aizenbud, Y. Aizenbud, contributed to data acquisition, critically revised the manuscript; M. Brandwein, contributed to data analysis and interpretation, drafted and critically revised the manuscript; L. Shapira, contributed to conception and data interpretation, critically revised the manuscript; A.H. Hovav, A. Wilensky, contributed to conception, design, data analysis, and interpretation, drafted and critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
DS_10.1177_0022034520920577 – Supplemental material for Niche Specific Microbiota-Dependent and Independent Bone Loss around Dental Implants and Teeth
Supplemental material, DS_10.1177_0022034520920577 for Niche Specific Microbiota-Dependent and Independent Bone Loss around Dental Implants and Teeth by O. Heyman, Y. Horev, N. Koren, O. Barel, I. Aizenbud, Y. Aizenbud, M. Brandwein, L. Shapira, A.H. Hovav and A. Wilensky in Journal of Dental Research
Footnotes
Acknowledgements
We thank MIS Implants Technologies for manufacturing the microimplants.
A supplemental appendix to this article is available online.
This work was supported in part by Israel Science Foundation grant 2369/18 to A.W.
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.