
Abstract
Forkhead box O1 (FoxO1) is a multifunctional initiator, mediator, and repressor of autoimmune diseases in an organ- or disease-specific manner. However, the role of FoxO1 in the salivary gland has not yet been elucidated. In this study, we discovered that FoxO1 and aquaporin 5 (AQP5) are both significantly downregulated in the patients with primary Sjögren syndrome, an autoimmune disease accompanying salivary gland dysfunction. Pharmacologic or genetic perturbation of FoxO1 in the rat salivary gland acinar cell line, SMG-C6, induced a significant downregulation of AQP5 expression, as observed in clinical specimens. There was a strong correlation between FoxO1 and AQP5 expression because FoxO1 is a direct regulator of AQP5 expression in salivary gland acinar cells through its interaction with the promoter region of AQP5. Serial injection of a FoxO1 inhibitor into mice induced a reduction of AQP5 expression in submandibular glands and, consequently, hyposalivation, which is one of the major clinical symptoms of primary Sjögren syndrome. However, there was no sign of inflammation or cell damage in the submandibular glands harvested from mice treated with the FoxO1 inhibitor. In conclusion, our findings indicate that FoxO1 in salivary gland tissue acts as a direct regulator of AQP5 expression. Thus, downregulation of FoxO1 observed in primary Sjögren syndrome is a putative mechanism for hyposalivation without the involvement of previously reported soluble factors in primary Sjögren syndrome patient sera.
Introduction
Forkhead box O1 (FoxO1) is an important transcriptional factor associated with various physiological functions such as vascular development (Hosaka et al. 2004), oxidative stress regulation (Tothova et al. 2007), and metabolism (Puig and Tjian 2005). The activities of FoxO1 are mainly regulated by phosphorylation, which inhibits its nuclear localization and thus becomes inactive (Rena et al. 2001). Recently, studies have shown an increased interest in the relationship between FoxO1 and autoimmune diseases. Deficiency of FoxO1 impairs function and differentiation of regulatory T cells and has been found to prevent or mitigate autoimmune diseases (Ouyang et al. 2010). In rheumatoid arthritis, FoxO1 is downregulated in peripheral blood mononuclear cells (Kuo and Lin 2007), and the majority of FoxO1 in synovial macrophages is in a phosphorylated form (Ludikhuize et al. 2007). However, roles of FoxO1 in salivary glands and salivary gland–manifesting autoimmune diseases have not yet been elucidated.
Primary Sjögren syndrome (pSS) is a chronic and systemic autoimmune disease exhibiting diverse clinical manifestations such as arthralgias, neuropathy, kidney dysfunction, vasculitis, gastrointestinal disorders, and secretory dysfunction in exocrine organs, including lacrimal and salivary glands (Baldini et al. 2012). Among these clinical presentations, hyposalivation significantly affects oral, systemic, and psychosocial health (Brito-Zeron, Kostov, et al. 2016; Vivino 2017). Since there is no fundamental cure for pSS, treatment is limited to the temporary stimulation of salivary secretion with pilocarpine or cevimeline for relief of oral dryness (Vivino 2017). Although the exact pathogenesis of pSS is still in debate, it is widely accepted that pSS can be considered an autoimmune epithelitis, suggesting that epithelial cells are the central regulators of the autoimmune response in pSS (Brito-Zeron, Baldini, et al. 2016). Activation of immune-competent molecules expressed by salivary gland epithelial cells (SGECs) induces focal lymphocytic infiltration and apoptosis of SGECs (Brito-Zeron, Baldini, et al. 2016). Other than the SGEC apoptosis, expressional or functional dysregulations of salivation-related proteins, including M3 muscarinic receptor and aquaporin 5 (AQP5), also largely contribute to the sicca symptom in pSS (Li et al. 2004; Alam et al. 2016).
AQP5 is the most important protein for salivation since it directly mediates the transcellular movement of water from the basal to apical or lumen side of acinar cells (Matsuzaki et al. 1999). In salivary gland acinar cells, the function of AQP5 is regulated by its localization and expression level through parasympathetic innervation (Hosoi et al. 2020). As described above, there have been numerous reports indicating that both AQP5 localization and expression are dysregulated in patients with pSS and animal models, primarily due to soluble factors in the sera (Delporte et al. 2016). However, only a few studies have examined the contributing factors for AQP5 dysregulation and hyposalivation in salivary gland tissue from patients with pSS. From a therapeutic perspective, it is important to identify tissue origin factors, because the local delivery of therapeutic molecules to salivary glands is significantly safer than systemic delivery, which possesses risks of systemic complications.
In this study, we report that FoxO1 directly regulates AQP5 expression in salivary gland acinar cells. In patients with pSS, both FoxO1 and AQP5 were downregulated compared with healthy subjects. Throughout in vitro and in vivo experiments, we found that FoxO1 functions as a transcription factor that binds to the promoter region of AQP5, and its downregulation or inhibition results in the downregulation of AQP5 expression and hyposalivation concomitantly.
Materials and Methods
Animals
Eight-week-old male ICR mice were used in this study. The animal experiment protocol for this study was approved by the Seoul National University Institutional Animal Care and Use Committee (approval number: SNU-160322-2-3).
Patient Sample Collection
After local anesthesia, an approximate 1-cm horizontal incision was made in the lower lip mucosa. Four to 5 minor salivary gland lobules were harvested and frozen for further evaluation. pSS was confirmed according to 2016 American College of Rheumatology/European League Against Rheumatism classification criteria (Shiboski et al. 2017). This study was approved by the Ethnic Committee of Kyungpook National University (2018-0010).
Genetic Perturbation of FoxO1
The lentiCRISPRv2 plasmid backbone (Sanjana et al. 2014) was used to construct a plasmid for CRISPR-mediated downregulation of FoxO1 (LC-FoxO1). A pair of oligos (5′-CACC GCGGCCGCAGCTGCCGCCAGT-3′ and 5′-AAACACTG GCGGCAGCTGCGGCCGC-3′) were treated with T4 polynucleotide kinase (NEB) and annealed using a Biometra TOne 96g (Analytik Jena). The annealed oligos were ligated into the plasmid with T4 DNA ligase (NEB). The pcDNA3 Flag FKHR plasmid was a gift from Kunliang Guan (Addgene plasmid 13507) (Tang et al. 1999).
Chemicals and Antibodies
AS1842856 was purchased from Calbiochem. Anti-AQP5 was obtained from US Biological and Alomone Labs. Anti-FoxO1a and anti–lamin B1 were purchased from Abcam and Thermo Fisher Scientific. Anti–β-actin was purchased from Sigma Aldrich. Anti-rabbit IgG was purchased from Cell Signaling.
Cell Culture and Transfection
The immortalized rat submandibular epithelial cell line, SMG-C6, was maintained as described previously (Quissell et al. 1997). LentiCRISPR-FoxO1 (LC-FoxO1) plasmid was transfected into SMG-C6 via electroporation using the Neon Transfection System (Invitrogen) according to the manufacturer’s instructions. LentiCRISPR-Green fluorescent protein (LC-GFP) plasmid was inserted into SMG-C6 as a control cell. Transfected SMG-C6 cells were selected with 4 µg/mL puromycin and maintained with puromycin-containing media. pcDNA3 Flag FKHR, LC-GFP, and LC-FoxO1–transfected SMG-C6 cells were selected and maintained in 4 µg/mL puromycin and 400 µg/mL G418.
Quantitative Real-Time Polymerase Chain Reaction
Total cell and tissue RNA were extracted with an RNA purification kit (Favorgen) and TRIzol reagent (Favorgen). Total RNA (50 ng) was reverse-transcribed using a First Strand cDNA Synthesis Kit (Thermo Fisher Scientific) according to the manufacturer’s instructions. Quantitative reverse transcription polymerase chain reaction (RT-PCR) was performed using the Luna qPCR master mix (NEB) and the Mic qPCR Cycler (Bio Molecular Systems). Relative messenger RNA (mRNA) levels were calculated as 2[Ct(β-actin) − Ct (gene of interest)] and the mRNA level of the control cells was set to 1. The primer sequences used in this study are listed in Appendix Table 1.
Protein Preparation and Immunoblot Analysis
Whole tissue and cell extracts were mixed with Laemmli buffer (60 mM Tris-HCl [pH 6.8], 2% (w/v) sodium dodecyl sulfate [SDS], 10% (v/v) glycerol and 0.02% (w/v) bromophenol blue), followed by sonication and heat denaturation. For the nuclear protein extraction, NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific) were used. The extracted proteins were subjected to SDS polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes. After blocking and antibody incubation steps, bands were detected using the D-Plus ECL Femto system (Dongin LS). Quantitation of Western blots was done with ImageJ software (National Institutes of Health).
Chromatin Immunoprecipitation
Cells were crosslinked with 1% formaldehyde followed by quenching with 125 mM glycine, treated with lysis buffer (1% SDS, 10 mM EDTA, and 50 mM Tris [pH 8.0]), sonicated, and mixed with dilution buffer (0.01% SDS, 1.1% Triton X-100, 1.2 mM EDTA, 16.7 mM Tris-HCl [pH 8.0], and 150 mM NaCl). Sheared DNA was incubated with antibodies overnight at 4°C. Lysates were incubated with Dynabeads Protein G (Thermo Fisher Scientific), and the precipitates were washed with SDS-carbonate (1% SDS and 0.1 M NaHCO3). The eluted samples were mixed with 5 M NaCl for reverse crosslinking. DNA was then purified after incubation with RNase A (Roche) and Proteinase K (Roche) using a QIAquick PCR Purification Kit (QIAGEN) following the manufacturer’s instruction. Enrichment was measured with a Mic qPCR Cycler (Bio Molecular Systems). The primer sequences are listed in Appendix Table 1.
Immunohistochemistry
Mouse submandibular glands (SMGs) were isolated and immediately fixed with 4% paraformaldehyde–phosphate-buffered saline (PBS) for 3 d at 4°C. For immunohistochemical staining of AQP5, fixed mouse SMGs embedded in paraffin wax block were sectioned and immunostained with anti-AQP5 antibody (1:1,000; Alomone Labs) following a standard protocol. Negative controls for AQP5 antibody were performed by staining E18 mouse embryonic kidney (Appendix Fig. 1) (Larsen et al. 2011).
Functional Assessment of Mouse Salivary Gland In Vivo
Murine saliva secretion was measured using the modified version of a previously described method (Varghese et al. 2018). Briefly, mice were anesthetized with a mixture of ketamine (25 mg/kg/mL; Yuhan Co, Ltd.) and xylazine (5 mg/kg/mL; Sigma Aldrich), and preweighted cotton pellets were firmly located under the tongue. Next, 300 µg/kg pilocarpine was intraperitoneally injected to stimulate saliva secretion. Saliva was collected for a total of 10 min following stimulation, and the weight difference in the saliva-absorbed cotton pellets was measured.
Gaussia Luciferase Assay
The rat AQP5 promoter reporter Gaussia luciferase (GLuc) plasmid (Genecopoeia, cat. RPRM44915-PG02) and TK- Renilla plasmid were cotransfected into LC-GFP and LC-FoxO1 SMG-C6 cells using Lipofectamine 3000 (Life Technologies), according to the manufacturer’s instructions, and cultured for 48 h. After harvesting the cultured media and cells, luciferase activity was measured using a secrete-pair Gaussian luciferase assay kit (Genecopoeia), a dual-luciferase reporter assay system (Promega), and a GloMax 20/20 Luminometer (Promega). The relative luciferase activity was determined as the ratio of Gaussia to Renilla activity.
Data Collection and Statistical Analysis
Statistical calculations were done by an unpaired t test or a one-way analysis of variance (ANOVA) with Tukey’s multiple comparison test using Prism 8.4.2 software (GraphPad Software). Statistical significance was set to *P < 0.05, **P < 0.01, ***P < 0.005, and ****P < 0.001.
Results
Simultaneous Downregulation of FoxO1 and AQP5 in pSS Patients
Expression levels of FoxO1 and AQP5 were examined in our sample sets of both healthy subjects (n = 9) and patients with pSS (n = 8). In minor salivary gland samples from patients with pSS, mRNA and protein levels of both FoxO1 and AQP5 were significantly decreased compared with those in healthy subjects (Fig. 1A–C; Appendix Fig. 2). A nonparametric Spearman correlation analysis revealed that the protein expression levels of FoxO1 and AQP5 were significantly correlated in both patients with pSS and healthy subjects (n = 17, P = 0.0017, r = 0.7157), which raises the possibility that FoxO1 regulates the expression of AQP5 (Fig. 1D).
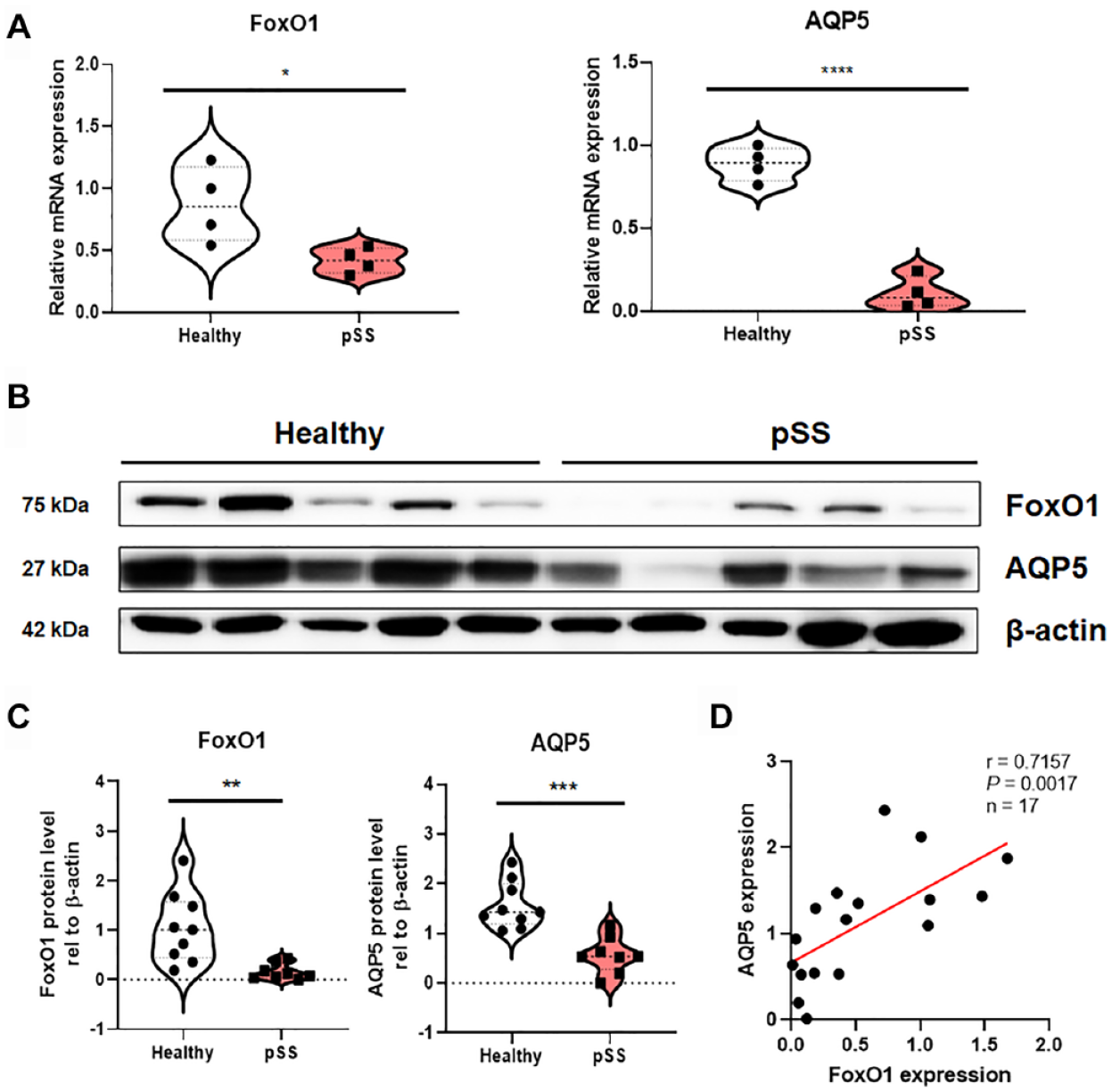
(
Chemical and Genetic Perturbation of FoxO1 Leads to Downregulation of AQP5
Inspired by the aforementioned results, we used chemical and genetic techniques to alter FoxO1 in SMG-C6 cells, an immortalized rat submandibular gland acinar cell line, to further prove a direct relationship between FoxO1 and AQP5 in salivary glands. AS1842856 is a chemical compound that selectively inhibits FoxO1’s transcriptional activity by directly binding to the active form without affecting the inactive form (Nagashima et al. 2010). Treatment with AS1842856 significantly decreased nuclear FoxO1, AQP5 mRNA, and protein levels in SMG-C6 cells in a dose-dependent manner (Fig. 2A–C). To confirm that the decreased AQP5 in AS1842856-treated SMG-C6 cells is not due to the off-target effects of the drug, we genetically disrupted FoxO1 function in SMG-C6 cells by using a CRISPR/Cas9 genome editing tool. Compared with the control (LC-GFP) cells, both total and nuclear FoxO1 expressions were significantly decreased in LC-FoxO1 cells, validating the knockout technique (Fig. 2E, F). As observed in AS1842856-treated cells, a significant downregulation of AQP5 was also detected in LC-FoxO1 cells compared with control cells (Fig. 2D–F). To further confirm the dependency of AQP5 expression on FoxO1, we artificially restored the expression of FoxO1 by transfecting LC-FoxO1 cells with FoxO1 plasmid (FKHR). FKHR-transfected LC-FoxO1 cells successfully restored FoxO1 expression compared with that in LC-GFP cells. Concordantly, AQP5 expression was also fully recovered compared with that in LC-FoxO1 cells transfected with an empty vector (EV) (Fig. 2G, H).
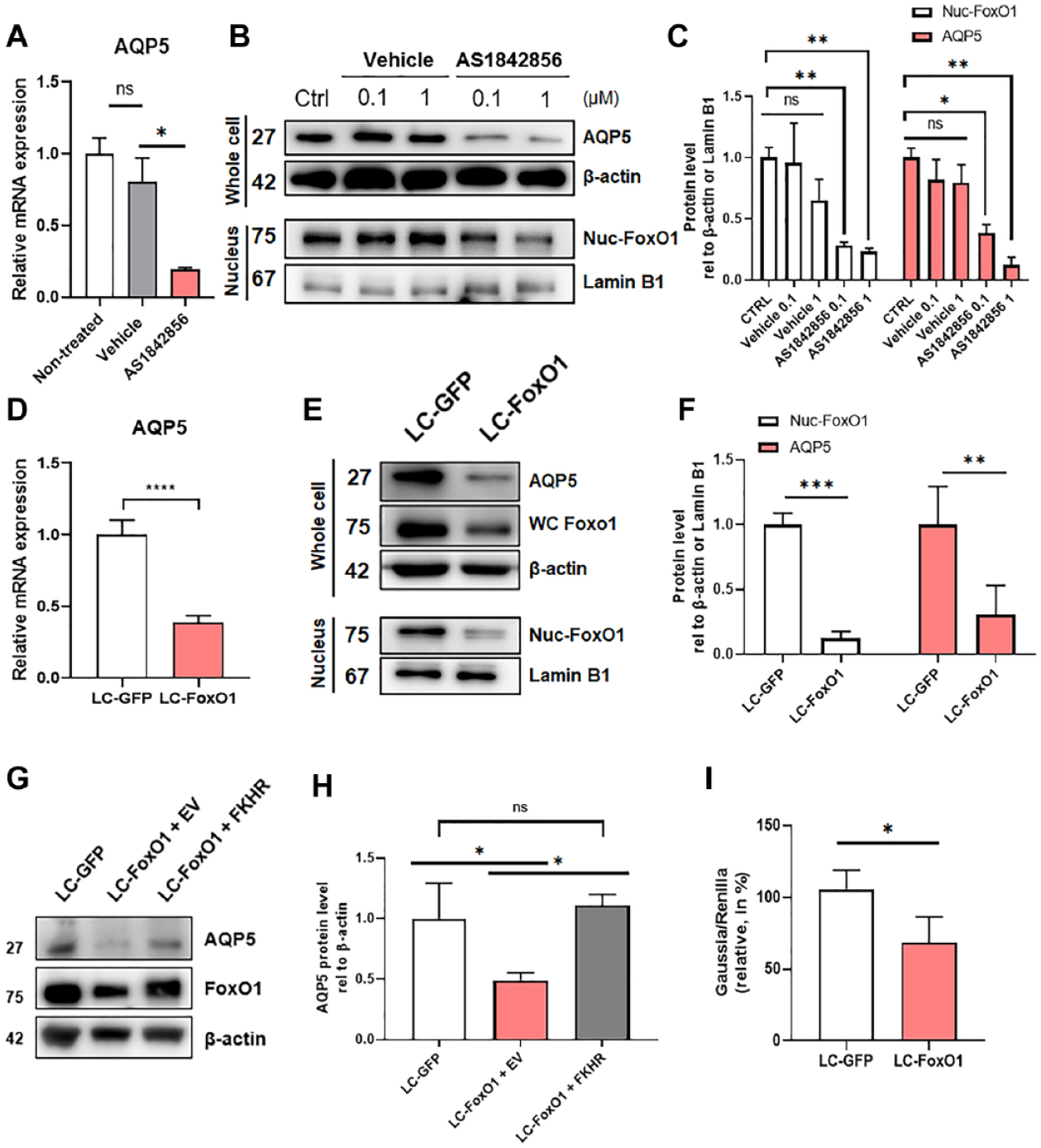
Chemical and genetic perturbation of FoxO1 in SMG-C6 cells. (
To determine whether the downregulation of AQP5 in FoxO1-perturbed cells resulted from the alteration of AQP5 mRNA stability, we performed an mRNA stability test using actinomycin D. No significant differences in AQP5 mRNA degradation rate were observed in control (LC-GFP) or LC-FoxO1 cells, indicating that FoxO1 had no effect on AQP5 mRNA stability (Appendix Fig. 3). Therefore, it is highly likely that the direct correlation between FoxO1 and AQP5 expression is because of the transcriptional activity of FoxO1. To prove this, the Gaussia luciferase (GLuc) AQP5 promoter plasmid was transfected into LC-GFP and LC-FoxO1 SMG-C6 cells to quantitatively measure the transcriptional activity of the AQP5 gene. As expected, a significant decrease in luciferase protein was observed in LC-FoxO1 cells compared with that in LC-GFP cells, providing further evidence that FoxO1 regulates the promoter region of the AQP5 gene (Fig. 2I).
Transcriptional Factor FoxO1 Interacts with a Promoter Region of AQP5
To confirm the interaction between FoxO1 and the AQP5 promoter, a chromatin immunoprecipitation (ChIP) was performed. Based on the previously reported response element sequences for FoxO1 binding (Wang et al. 2014), we analyzed AQP5 promoter sequences and selected 4 different regions for evaluating the interaction with FoxO1 (Fig. 3A; Appendix Fig. 4). Within the AQP5 promoter, 9 potential binding sequences for FoxO1 were identified (Appendix Fig. 4). One of these was located in the first region (AQP5(1)), 2 in the second region (AQP5(2)), and none of them in the third (AQP5(3)) or fourth region (AQP5(4)) (Appendix Fig. 4). As expected, a significant increase in FoxO1 binding was observed only for AQP5(2) of the vehicle (DMSO)-treated group, and it was significantly decreased in the 0.1-µM AS1842586-treated group (Fig. 3B). Similarly, a significant increase in FoxO1 binding was observed only in AQP5(2) of LC-GFP cells, and it was significantly decreased in LC-FoxO1 cells (Fig. 3C). To further prove that the AQP5(2) is the key FoxO1 binding site, we transfected LC-FoxO1 SMG-C6 cells with pGL3 vector containing region 2–deleted AQP5 promoter. As described above, when full promoter sequences of rat AQP5 were transfected, luciferase activity was decreased in LC-FoxO1 compared to LC-GFP SMG-C6 cells (Fig. 2I). However, when region 2–deleted promoter sequences of AQP5 were transfected, presence or absence of FoxO1 could not induce significant difference in luciferase activity in LC-GFP and LC-FoxO1 SMG-C6 cells (Appendix Fig. 5). These results indicate that FoxO1 functions as a transcriptional factor to upregulate the expression of AQP5 through a DNA-protein interaction.
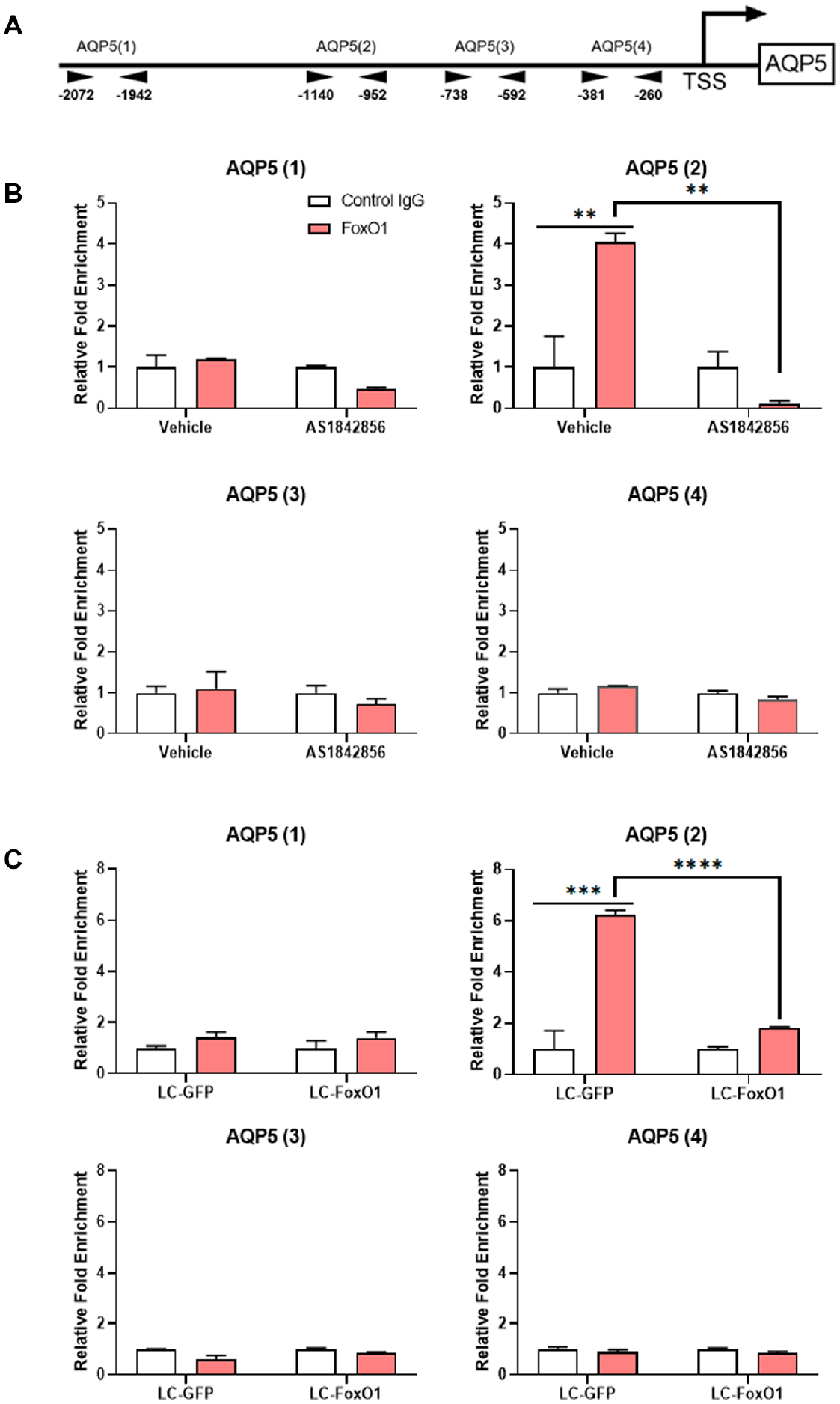
Interaction between FoxO1 and a promoter region of AQP5. (
Inhibition of FoxO1 Causes Hyposalivation and Downregulation of AQP5 In Vivo
As described above, decreased AQP5 expression and salivation are important pathophysiological characteristics of pSS. Therefore, we tested whether the inhibition of FoxO1 can recapitulate these pSS symptoms in vivo. First, we confirmed that mouse SMGs significantly express AQP5 and FoxO1 (Appendix Fig. 6). ICR mice were injected intraperitoneally with 15 mg/kg AS1842856 three times with a 2-d interval between treatments. Two days after the last injection, pilocarpine-induced salivary secretion was measured (Fig. 4A). The AS1842856-treated group exhibited an approximate 30% reduction in salivary secretion compared to the vehicle control (VC) group (Fig. 4B). As we observed in in vitro studies, mRNA and protein levels of AQP5 in murine SMGs were also significantly decreased in the AS1842856-treated group compared with the VC group (Fig. 4C–E). To confirm whether the hyposalivation with decreased AQP5 expression was due to acinar cell apoptosis or inflammation, terminal deoxynucleotidyl transferase-mediated dUTP nick end-labeling (TUNEL) assay and hematoxylin and eosin (H&E) staining were performed. Neither the VC nor the AS184 2856-treated group showed a significant number of TUNEL-positive cells and lymphocytic infiltration, suggesting that the inhibition of FoxO1 can induce hyposalivation, even in the absence of inflammation or overt acinar cell damage (Fig. 4F; Appendix Fig. 7). Immunohistochemical staining of AQP5 revealed that inhibition of FoxO1 via AS1842856 significantly reduced AQP5 expression in acinar cells without altering acinar area, acinar/ductal morphology, or salivary gland size (Fig. 4G, H). In summary, FoxO1-dependent regulation of AQP5 expression in salivary gland cells was reproduced in vivo as we successfully induced hyposalivation, a hallmark symptom of pSS.
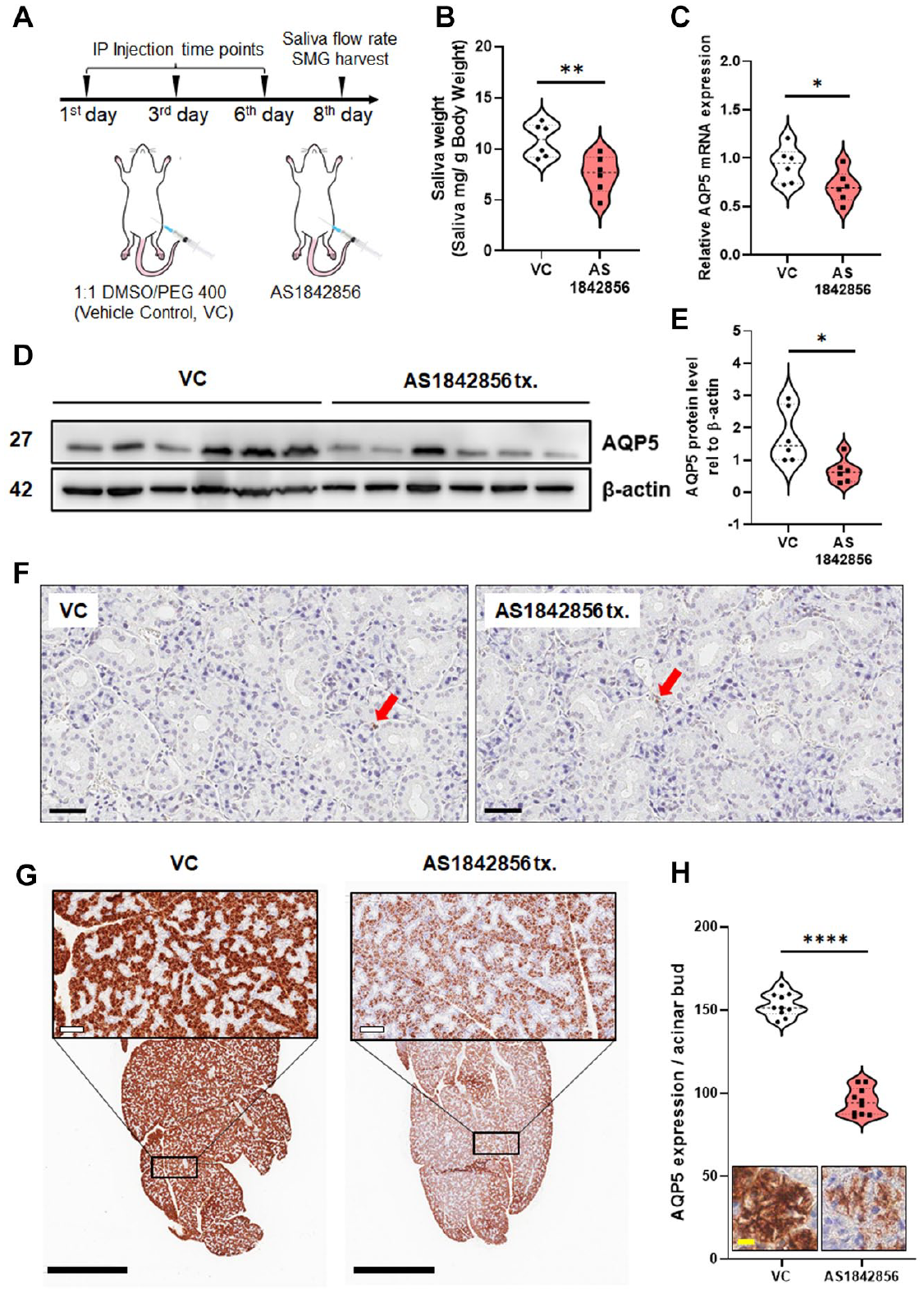
(
Discussion
In this study, we have experimentally demonstrated that FoxO1 can regulate AQP5 expression in salivary gland acinar cells. FoxO1 functions as a transcriptional activator of AQP5 expression by binding to the AQP5 promoter region. Based on the results, we have suggested a possibility that the decreased expression of FoxO1 in patients with pSS is a putative contributing factor for salivary gland dysfunction in pSS. Without treatment of classically defined pathological factors of pSS including pSS patient sera or pSS-related cytokines/autoantibodies, inhibition of FoxO1 was enough to reduce AQP5 expression in salivary gland epithelial cells. Furthermore, we confirmed that inhibition of FoxO1 in normal mice induced hyposalivation and downregulation of AQP5 in submandibular glands, recapitulating the characteristics of pSS. However, although a significant correlation between FoxO1 and pSS has been found in our study, it is necessary to collect diversified evidence to clearly prove our hypothesis. First, confirmation of FoxO1 expression levels in pSS mimicking animal models and additional gain-of-function study should be conducted to assess FoxO1’s contribution in pSS pathophysiology. Second, FoxO1 expression in other cell types, including immune cells, endothelial cells, and nerve cells of patients with pSS, should be comprehensively examined, since pSS is a complex multifactorial disease in which various cell types are involved (Brito-Zeron, Baldini, et al. 2016). Last, although the expression of FoxO1 is definitely decreased in salivary glands of patients with pSS, phosphorylation state of FoxO1 should also be examined, since it is the main regulatory mechanism for FoxO1’s activity (Rena et al. 2001).
Our study also raises the important question of whether downregulated FoxO1 is a causative factor for the onset of pSS or a consequence of pSS. If the systemic or salivary gland–specific downregulation of FoxO1 is a risk factor for the onset of pSS, the expression level of FoxO1 itself may be used as a biomarker for the early diagnosis of pSS. As mentioned above, FoxO1 is known as a regulator of the immune system. Although we did not observe any significant lymphocytic infiltration or acinar cell apoptosis in mice treated with FoxO1 inhibitors, FoxO1 downregulation might be one of induction signals for pSS development considering that our experiment is limited to the acute phase of FoxO1 inhibition. Otherwise, downregulated FoxO1 is simply a consequence of pSS rather than an inducer. In this case, one possible explanation is micro-RNA (miRNA)–mediated downregulation of FoxO1. It has been reported that miRNAs, such as miR-27a, miR-96, miR182, and miR370, target the 3′-untranslated regions of FoxO1 and thus downregulate FoxO1 expression in breast and prostate cancer cells (Sundaresan and Puthanveetil 2017). Since numerous miRNAs are significantly up- or downregulated in salivary gland epithelial cells of patients with pSS, posttranscriptional regulation of FoxO1 may reveal the mechanism of FoxO1 downregulation (Reale et al. 2018).
It is noteworthy that the difference in AQP5 expression between healthy subjects and patients with pSS is controversial. Xiao et al. (2011) reported that AQP5 expression in minor salivary glands biopsied from patients with pSS was significantly lower compared with that from healthy controls (n = 8). In contrast, Beroukas et al. (2001) reported that there was no difference in AQP5 expression and localization between healthy controls and patients with pSS (n = 10). Although the sample number used in the present study was less than that of the other 2 studies, both relied on immunofluorescence or immunohistochemical analysis, whereas we used quantitative RT-PCR and Western blot to simultaneously confirm AQP5 mRNA and protein expression. According to our results, both AQP5 mRNA and protein expression in the minor salivary glands of patients with pSS are significantly reduced compared with healthy controls. Our results are also supported by similar results from previous studies indicating reduced salivary gland AQP5 expression in nonobese diabetic mice, a widely used animal model for pSS (Soyfoo et al. 2018).
In conclusion, our data suggest that the expression of FoxO1 in salivary gland tissue is a possible contributing factor for the dysregulation of AQP5 and hyposalivation in pSS, independent from salivary autoimmune processes.
Author Contributions
S.M. Lee, S.W. Lee, contributed to conception, design, data acquisition, analysis, and interpretation, drafted the manuscript; M. Kang, contributed to data acquisition, analysis, and interpretation, critically revised the manuscript; J.K. Choi, contributed to data acquisition, critically revised the manuscript; K. Park, J.S. Byun, D.Y. Kim, contributed to conception, data acquisition, analysis, and interpretation, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
sj-pdf-1-jdr-10.1177_00220345211003490 – Supplemental material for FoxO1 as a Regulator of Aquaporin 5 Expression in the Salivary Gland
Supplemental material, sj-pdf-1-jdr-10.1177_00220345211003490 for FoxO1 as a Regulator of Aquaporin 5 Expression in the Salivary Gland by S.M. Lee, S.W. Lee, M. Kang, J.K. Choi, K. Park, J.S. Byun and D.Y. Kim in Journal of Dental Research
Footnotes
Acknowledgements
We thank Dr. Jae-Kwang Jung for his critical comments and helpful suggestions.
A supplemental appendix to this article is available online.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (No. 2017 R1A5A2015391 and No. 2020R1C1C1006757). This research was also supported by the Basic Science Research Program through the NRF funded by the Ministry of Education (No. 2019R1I1A2A01062430 and No. 2020R1A6A3A13075289).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.