
Abstract
Mucin in saliva plays a critical role in the hydration and lubrication of the oral mucosa by retaining water molecules, and its impaired function may be associated with hyposalivation-independent xerostomia. Age-dependent effects on salivary gland function and rheological properties of secreted saliva are not fully understood as aging is a complex and multifactorial process. We aimed to evaluate age-related changes in the rheological properties of saliva and elucidate the underlying mechanism. We performed ex vivo submandibular gland (SMG) and sublingual gland (SLG) perfusion experiments to collect saliva from isolated glands of young (12 wk old) and aged (27 mo old) female C57BL/6J mice and investigate the rheological properties by determining the spinnbarkeit (viscoelasticity). While fluid secretion was comparable in SMG and SLG of both mice, spinnbarkeit showed a significant decrease in SLG saliva of aged mice than that of young mice. There were no significant differences in GalNAc concentration between young and aged SLG saliva. Liquid chromatography/tandem mass spectrometry analysis of SLG saliva revealed that (Hex)1 (HexNAc)1 (NeuAc)1 at m/z 793.31 was the most abundant O-glycan structure in SLG saliva commonly detected in both mice. Lectin staining of salivary gland tissue showed that SLG stained strongly with Maackia amurensis lectin II (MAL II) while Sambucus nigra agglutinin (SNA) stained little, if any, SLG. The messenger RNA expression of St3gal1 that encodes an α-2,3 sialic acid sialyltransferase SIAT4-A showed a decrease in SLG of aged mice, confirmed by a Western blot analysis. Lectin blot analysis in SLG saliva revealed that the relative signal intensity detected by MAL II was significantly lower in aged SLG. Our results suggest that spinnbarkeit decreases in SLG of aging mice due to downregulation of sialic acid linked to α-2,3 sialic acid sialyltransferase expression.
Introduction
Population aging is a global phenomenon. The world population over 65 y of age is estimated to be 16% by 2050, accounting for 1 in 6 people (Raftery et al. 2012). As life expectancy increases, oral health care becomes crucial to maintain quality of life (Iwasaki et al. 2021), with saliva playing a critical role in maintaining oral health (Dodds et al. 2005; Iwasaki et al. 2018). Clinical research and animal studies have demonstrated an association between aging and xerostomia, the subjective sensation of dry mouth (Miyagi et al. 2019; Proctor and Shaalan 2021). Estimates show that 30% of adults over 65 y have xerostomia (Ship et al. 2002). Although xerostomia is associated with decreased salivary flow in the stimulated or unstimulated state, recent studies have revealed altered rheological properties of saliva in patients with xerostomia (Chaudhury et al. 2015, 2016). Rheological properties (viscosity, elasticity, stickiness) are essential salivary characteristics that enable a thin layer of residual saliva to coat mucosal surfaces by water retention (Osailan et al. 2011) with gel-forming mucins. Decrease or structural changes in gel-forming mucins can influence the rheology of saliva, leading to hyposalivation-independent dryness in older patients (Wolff and Kleinberg 1998). The mouse sublingual gland (SLG) presents the mucous acinar cell phenotype secreting viscous saliva (Culp et al. 2004). Recent RNA-sequence analysis of 3 major salivary glands in mice revealed differentially expressed genes encoding mucins in each gland (Gao et al. 2018). Among the transcripts encoding secreted high-molecular-weight, gel-forming mucins (i.e., Muc2, Muc5ac, Muc5b, and Muc6), Muc19 was the only large secreted mucin transcript in adult mouse SLG, with little expression in the submandibular gland (SMG) and parotid gland (PG) (Das et al. 2009). Thus, SLG is a promising target for investigating the effect of aging on mucous cell phenotypes to elucidate altered salivary characteristics. However, collecting and assessing secreted SLG saliva is challenging because the fluid amount is too little for in vivo experimental analysis (Kondo et al. 2015). Secretion is dependent on environmental factors, including body temperature, blood pressure, and, essentially, the central nervous system input (Turner and Sugiya 2002), all likely to be affected by aging (Pettan-Brewer and Treuting 2011). In SMG and SLG ex vivo experiments, surgical removal of the glands and perfusion are performed to collect ductal saliva and determine the effects of aging. This study explores the mechanism of age-related xerostomia by examining the changes in rheological properties of saliva with aging using ex vivo experiments.
Materials and Methods
Animals
Young female C57BL/6J mice (12 wk old; n = 11) and aged female mice (27 mo old; n = 9) were purchased from CLEA Japan, Inc. Both mice were kept in the same facility and maintained with an identical diet CE2 (CLEA Japan, Inc.). Mice were housed in micro-isolator cages with ad libitum access to laboratory chow and water during 12-h light and dark cycles; reagents were purchased from Sigma-Aldrich unless otherwise indicated. All animal procedures were approved by the Animal Committee of Kyushu Dental University (approval number 19-027). All experiments followed the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines.
Ex Vivo Perfusion of Submandibular Glands and Sublingual Glands
Ex vivo perfusion of SMG and SLG was performed (Kondo et al. 2019). Mice were anesthetized by intraperitoneal injections of chloral hydrate (400 mg/kg of body weight), following ligation of all branches of the common carotid artery (except the artery supplying the SMG and SLG). Using a microscope (SZX7; Olympus), both glands were removed and placed in a perfusion chamber at 37°C. SMG and SLG were perfused through the common carotid artery and stimulated by muscarinic receptor agonist carbachol (CCh; 0.3 µM) and β-adrenergic agonist isoproterenol (IPR; 1.0 µM). The concentration of the agonist-induced fluid secretion was based on previous reports (Mukaibo et al. 2018). We collected secreted saliva from SMG and SLG independently into each capillary tube. The perfusion solution was composed of 4.3 mM KCl, 120 mM NaCl, 25 mM NaHCO3, 5 mM glucose, 10 mM HEPES, 1 mM CaCl2, and 1 mM MgCl2 (pH 7.4, gassed with 95% O2 and 5%CO2).
Spinnbarkeit (Viscoelasticity) Property Measurement of Collected Saliva
The rheological properties of collected saliva were determined using the Neva Meter (IMI-0501; Ishikawa Iron Works) (Chaudhury et al. 2015, 2016). A freshly collected 25 µL saliva sample was loaded on the measuring dish, and the measuring device tip automatically descended until it touched the sample. Saliva was stretched with a constant stretching rate of 10 mm/s. The length of saliva at the point where the tug broke was measured using an optical laser running along the measuring tip. Seven consecutive measurements were performed, and the average of 5 measurements (excluding the maximum and minimum value) was used as the sample’s spinnbarkeit (mm). Saliva samples were incubated with 50 units of α2-3,6,8 neuraminidase (New England Biolabs #P0720S) for 30 min at 37°C, followed by spinnbarkeit measurement. All measurements were performed immediately after saliva collection at room temperature.
O-Glycan Quantitative Assay
We used a tear mucin assay kit (Cosmo Bio Co.) to measure the O-glycan concentration in saliva following the manufacturer’s instructions. The O-glycans were β-eliminated from the core protein under alkaline conditions and fluorescently labeled by binding of 2-cyanoacetamide (2-CNA) at the reduced terminal of the N-acetylgalactosamine (GalNAc) (Honda et al. 1980). The fluorescence intensity was obtained by measuring the excitation wavelength at 336 nm and the fluorescence wavelength at 383 nm using a fluorescent plate reader (Infinite 200Pro; Tecan).
Quantitative Polymerase Chain Reaction
Quantitative polymerase chain reaction (qPCR) was performed as described previously (Mukaibo et al. 2018) (details in the Appendix and primer sequences in Appendix Table 1). The messenger RNA (mRNA) levels were normalized to the abundance of Eef2 messages (Kouadjo et al. 2007).
Histochemical Staining
Immunofluorescent staining was performed (Mukaibo et al. 2018), and staining with lectin probes is shown in the Appendix.
Liquid Chromatography/Tandem Mass Spectrometry of O-Glycans in Collected Saliva
In total, 6 µL of each collected SLG saliva from young mice (n = 5) and aged mice (n = 5) was processed (Zeba Micro spin desalting columns; Thermo Fisher Scientific) to obtain 5-µL desalted samples of young and aged mice, each pooled into 25 µL. Each sample (10 µL) was further analyzed using the LCMS-IT-TOF (ion-trap time-of-flight mass spectrometer combined with liquid chromatogram) system (Shimadzu Corp.) (Kameyama et al. 2019). Glycan compositions were determined using the GlycoMod tool (Cooper et al. 2001) (procedure and system settings in the Appendix).
Protein Isolation and Western Blot Analysis
SLG lysate was prepared, and Western blot analysis was performed (see Appendix). In brief, 16 µg SLG lysate was separated in sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene difluoride (PVDF) membranes. The membranes were probed with polyclonal rabbit anti-SIAT4-A antibody (LifeSpan BioSciences) and, subsequently, monoclonal anti–β-actin antibody (Sigma).
Lectin Blot and Western Blot Analysis for Saliva Samples
Collected SLG saliva was desalted, separated by SDS-PAGE, and transferred to PVDF membranes. The membranes were probed with either Maackia amurensis lectin II (MAL II) or goat anti-Muc19 antibody. The signals were captured by Image Lab (Version 2.0.1; Bio-Rad) and quantitatively analyzed (see Appendix).
Statistical Analysis
Results are presented as the mean ± SE. Student’s t test and 1-way analysis of variance (ANOVA) were performed, followed by Bonferroni’s post hoc test to compare 2 groups or multiple groups, using Prism 8 software version 8.4.3 (GraphPad Software). P values less than 0.05 were considered statistically significant.
Results
Fluid Secretion of SMG and SLG Was Comparable in Young and Aged Mice
Isolated mouse SMG and SLG were perfused and stimulated with muscarinic receptor agonist CCh (0.3 µM) and β-adrenergic agonist IPR (1.0 µM), respectively, to examine fluid secretion. The measured flow amount in 30 min was comparable in young and aged mice (Fig. 1A, B) in both glands. Notably, the flow rate in the first 10 min in aged SMG was significantly lower than in young SMG (Fig. 1A). When normalized to gland weight, SMG flow rate in 10 min and total flow amount significantly decreased, whereas SLG did not show significant differences in flow rate/flow amount (Appendix Fig. 1A, B).
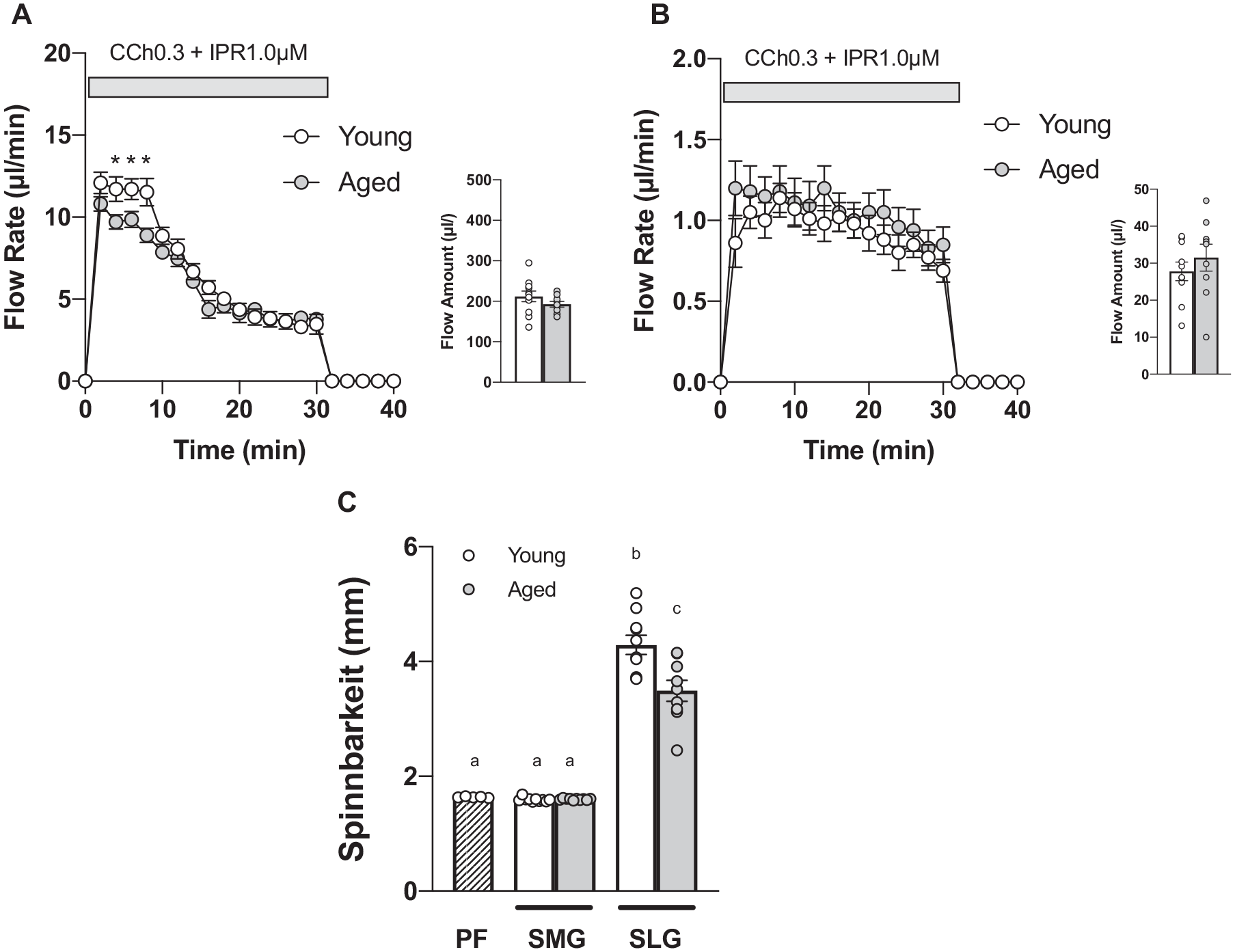
Fluid secretion was intact in aged submandibular gland (SMG) and sublingual gland (SLG), while spinnbarkeit decreased in aged SLG saliva. Mouse SMG and SLG were surgically removed, and each isolated gland was perfused with muscarinic receptor agonist carbachol (CCh; 0.3 μM) plus β-adrenergic receptor agonist isoproterenol (IPR; 1.0 μM) to induce saliva secretion. (
SLG Saliva of Aged Mice Showed Less Viscoelasticity Than That of Young Mice
Using a Neva Meter, collected saliva was measured for its viscoelasticity, determined as spinnbarkeit. Aged SLG saliva showed significantly lower spinnbarkeit values (18.6%) than young SLG saliva. SMG saliva of young and aged mice showed spinnbarkeit comparable to the perfusate (PF) (Fig. 1C). SLG saliva spinnbarkeit (after neuraminidase [NM] digestion) decreased by 23.7% and 16.6% in young and aged mice, respectively (Table).
Overview of Examined Properties of Saliva and Salivary Glands in Young and Aged Mice.
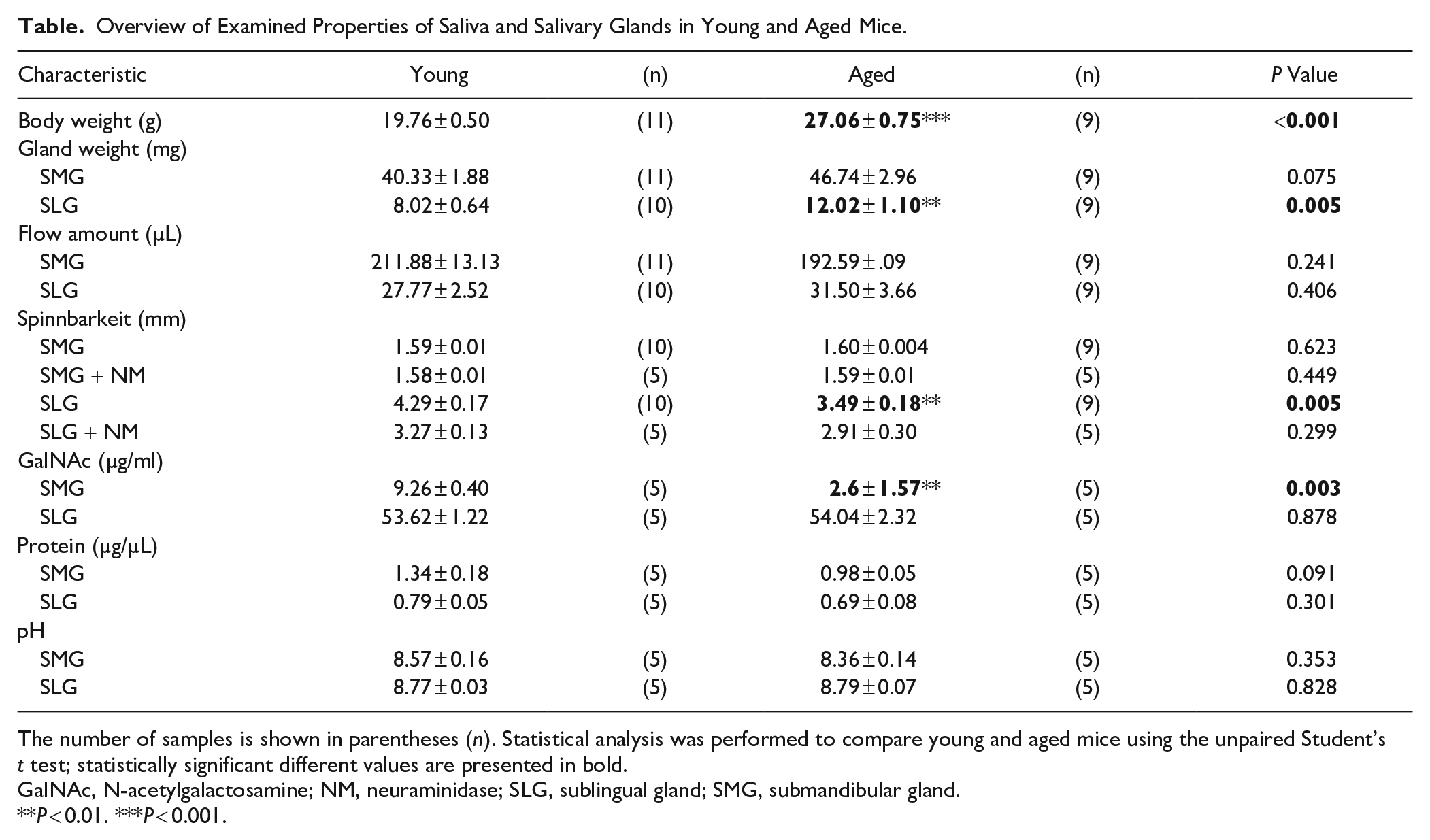
The number of samples is shown in parentheses (n). Statistical analysis was performed to compare young and aged mice using the unpaired Student’s t test; statistically significant different values are presented in bold.
GalNAc, N-acetylgalactosamine; NM, neuraminidase; SLG, sublingual gland; SMG, submandibular gland.
P < 0.01. ***P < 0.001.
GalNAc Concentration in SLG Saliva Was Comparable in Young and Aged SLG Saliva
Results showed that aged SLG saliva was less viscous than young SLG saliva. As glycan contents account for 80% of the mucin molecular mass (Bansil and Turner 2006), we initially hypothesized that O-glycan decreased in aged SLG saliva compared to young SLG saliva. GalNAc concentration was measured in young and aged SLG saliva to test the hypothesis. While GalNAc concentration was markedly higher in SLG than in SMG, there was no significant difference between young and aged mice SLG (Fig. 2A).
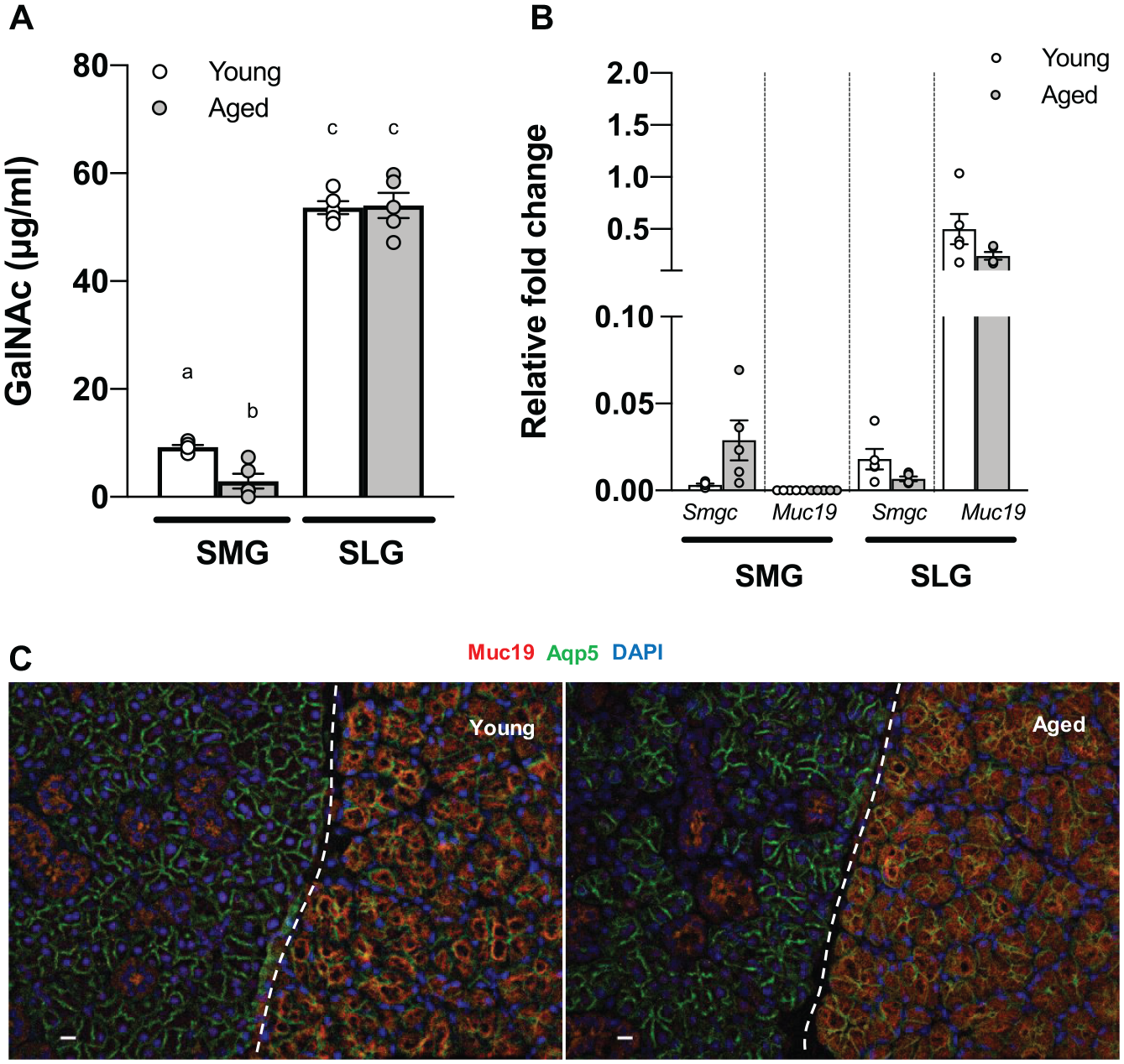
N-acetylgalactosamine (GalNAc) concentration, Smgc, and Muc19 messenger RNA (mRNA) expression and Muc19 localization were comparable in young and aged sublingual gland (SLG). (
Muc19 Localization and Gene Expression of Smgc and Muc19 SLG Were Comparable in Aged and Young Mice
Previous studies have shown the switch in gene splicing from Smgc to Muc19 during early mucous cell cytodifferentiation at postnatal 28 d (Das et al. 2009). Therefore, we analyzed the mRNA expression of both genes in young and aged SMG and SLG to determine any alteration in Muc19 gene expression for Smgc in aged glands. However, Smgc and Muc19 gene expressions were comparable in young and aged SLG, whereas Muc19 was almost undetectable in SMG (Fig. 2B). We examined the differences in Muc19 localization between young and aged SLG using immunohistochemistry. While the staining was negative in SMG acinar cells, Muc19 was exclusively targeted in SLG acinar cells. The localization was comparable in young and aged mice SLG, where Muc19 showed diffusion within the acinar cells (Fig. 2C). Negative control images are shown in Appendix Figure 3.
Sialo-Glycan Is the Most Abundant Glycan in SLG Saliva
To assess the glycan structure of collected SLG saliva, we used a liquid chromatography/tandem mass spectrometry (LC-MS/MS) analysis. High-performance liquid chromatography (HPLC) showed the highest peak at the same retention time (Fig. 3A). MS/MS analysis of peaks demonstrated that the most abundant O-glycan in young and aged mice SLG saliva was (Hex)1 (HexNAc)1 (NeuAc)1 at m/z 793.31 (Fig. 3B). The top 5 O-linked glycan peaks other than the highest peak in each sample are presented in Appendix Figure 4. We determined sialic acid composition in SLG by staining young SLG tissues with lectin MAL II, which detects α2,3-linked sialic acid linked to either N- or O-glycans. Consistent with immunofluorescent staining, SLG acinar cells showed stronger staining with MAL Ⅱ than SMG acinar cells. In contrast, Sambucus nigra agglutinin (SNA), which preferentially binds α2,6-linked sialic acid, stained little (if any) tissue in SLG, whereas SMG showed stronger staining (Fig. 4A, B).
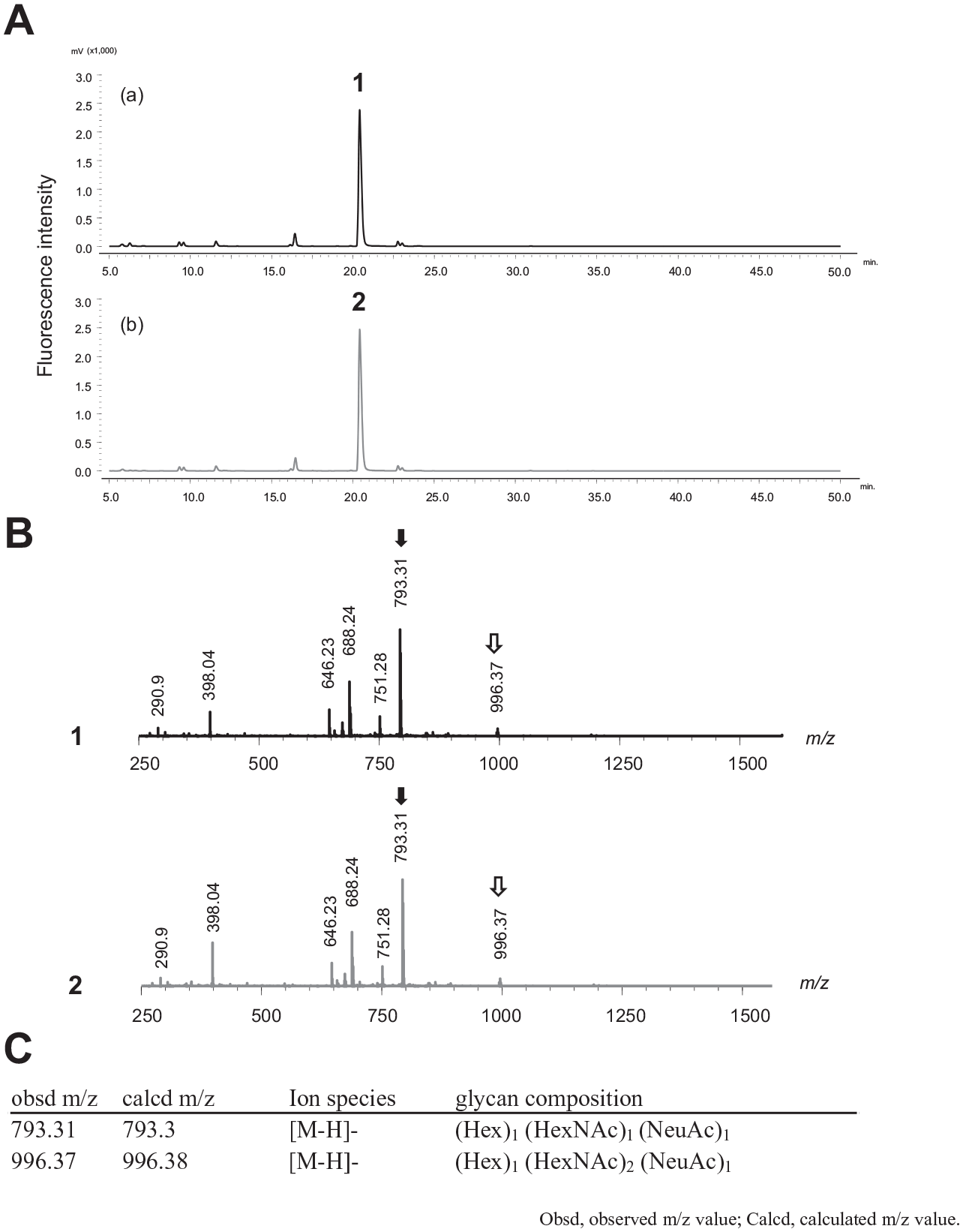
Analysis of O-linked glycans in aged and young sublingual gland (SLG) saliva using liquid chromatography/tandem mass spectrometry (LC-MS/MS). (
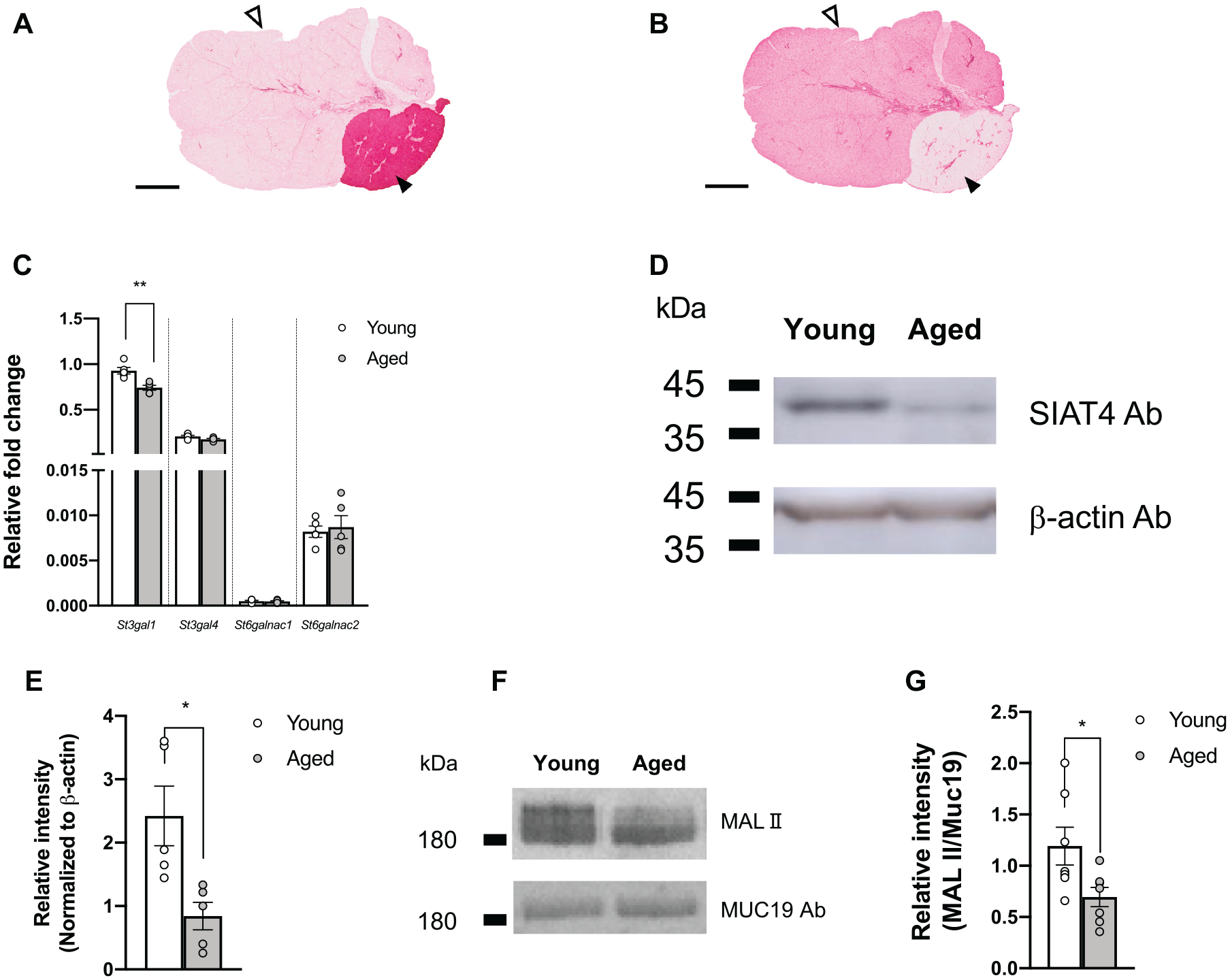
Distribution of gland-specific sialic acid detected using lectin and comparison of α-2,3 sialyltransferase messenger RNA (mRNA) and protein level. (
St3gal1 mRNA and SIAT4-A Protein Expression Was Significantly Lower in Aged SLG Than Young SLG
We further investigated the possible sialyltransferase genes that synthesize the sialo-glycan structure (Hex)1 (HexNAc)1 (NeuAc)1 in LC-MS/MS. Previous reports documented 6 subtypes of β-galactoside α-2,3 sialyltransferase from St3gal1 to St3gal6 expressed in SLG and SMG (Gao et al. 2018). Among those, St3gal1 and St3gal4 are highly expressed in SLG, mediating the transfer of sialic acid (N-acetyl-neuraminic acid; Neu5Ac) to core 1 O-glycan (Kono et al. 1997; Gao et al. 2018). We compared the expression levels between young and aged SLG using qPCR analysis. Results showed that St3gal1 expression was significantly decreased by 20% in aged SLG compared to young SLG. In contrast, the expression levels of potent sialyltransferase genes N-acetylgalactosaminde α-2,6 sialytransferase St6galnac1 and St6galnac2 were comparable in young and aged SLG (Fig. 4C). To examine the protein expression levels of SIAT4-A (molecular mass of 39 kDa) (UniportKB P54751) encoded by St3gal1 between young and aged SLG, 16 μg of whole-cell lysis was separated by SDS-PAGE, followed by Western blotting. Consistent with the St3gal1 mRNA expression pattern, SIAT4-A antibody-detected protein bands at approximately 40 kDa showed a 0.11-fold decrease in aged SLG compared to young SLG (Fig. 4D). To directly compare α-2,3 sialic acid in collected SLG saliva, we performed lectin blotting and Western blot analysis and quantitatively compared the signal intensity (MALII/Muc19) between aged and young SLG saliva. Aged SLG saliva showed significantly lower signal intensity than young SLG saliva (Fig. 4F, G, Appendix Fig. 5).
Discussion
We performed an ex vivo salivary gland experiment to evaluate the effect of aging on the rheology of SLG saliva. Spinnbarkeit of SLG saliva in aged mice was significantly decreased compared to young mice (Fig. 1C). Mucin provides viscosity to saliva (Park et al. 2007). Reports show that SLG predominantly secretes gel-forming mucin in rodents. Therefore, we focused on SLG and secreted saliva to identify the mechanism of age-dependent change in the rheological properties of aged mice. SMG served as a representative control to verify or corroborate SLG results. First, we examined the changes in fluid secretion with aging.
Consistent with a previous report (Miyagi et al. 2019), ex vivo SMG flow rate in 10 min decreased in aged mice, and Aqp5 mRNA expression reduced in aged SMG (Appendix Fig. 2A). In SLGs, the fluid secretion was comparable in aged and young mice (Fig. 1B) with the comparable gene expression levels involved in fluid secretion (Aqp5, Slc12a2, and Tmem16A) in aged and young SLG (Appendix Fig. 2B). Next, we used the Neva Meter to measure the rheological properties of saliva. Studies show that spinnbarkeit correlates with viscosity (Gohara et al. 2004) and is used to evaluate the rheology (viscoelasticity) of saliva (Chaudhury et al. 2015, 2016).
Results showed that spinnbarkeit decreased by 18.6% in aged SLG saliva compared to young SLG saliva (Fig. 1C). In contrast, the spinnbarkeit of SMG saliva was not significantly different from that of perfusate, suggesting little, if any, secretion of gel-forming mucin in SMG saliva. Reports show differential expression of Muc10 /Prol1 and Muc13 in mouse SMG relative to PG and SLG (Gao et al. 2018). The observations suggest that both mucins are membrane bound and not associated with the spinnbarkeit in saliva (Iida et al. 2019). Notably, gel-forming mucin Muc19 mRNA expression was almost absent in SMG (Fig. 2B).
Studies show bicarbonate concentration and pH as factors affecting salivary viscoelasticity (Vijay et al. 2015), possibly playing a role in decondensation of mucin granules by competing with fixed anions in mucin for Ca2+ (Chen et al. 2010). Bicarbonate is regulated by duct cells through the cystic fibrosis transmembrane regulator (CFTR) channel at the apical membrane of the duct cells in mice (Catalan et al. 2011; Gustafsson et al. 2012). Duct area in SLG and SMG accounts for approximately 10% and 33%, respectively (Kondo et al. 2015), suggesting an insignificant effect of bicarbonate on SLG saliva viscoelasticity in SLG. Notably, salivary pH measurement in SLG of young and aged mice was comparable (Table).
O-glycan contributes up to 80% of dry mucin molecular mass in secreted saliva (Zalewska et al. 2000), providing salivary viscoelasticity and determining lubrication and hydration by water retention in the O-glycan structure (Park et al. 2007; Inoue et al. 2008). We first hypothesized that decreased secretion of O-linked glycan could contribute to spinnbarkeit decrease in SLG saliva of aged mice compared to young mice. However, there was no significant difference in GalNAc concentration in SLG saliva between young and aged mice (Fig. 2A), implying that core protein Muc19 was intact in aged SLG saliva. Muc19 is the only gel-forming mucin exclusively expressed in SLG saliva of mice (Das et al. 2009; Culp et al. 2015). Muc19 is a splicing variant of Smgc with exon 1 in common and differentially spliced and switched from Smgc to Muc19 (Smgc, exons 2–18; Muc19, exons 19–60) at postnatal day 28 (Das et al. 2009). In line with the GalNAc concentration, there was no significant difference between young and aged SLG for Muc19 and Smgc (Fig. 2B). Muc19 localization was comparable in SLG of young and aged mice, predominantly in the acinar cells (Fig. 2C). Muc19 was hardly observed in SMG acinar cells, consistent with qPCR analysis results. LC-MS/MS was used to investigate the glycoprotein composition in SLG saliva. Results showed that the (Hex)1 (HexNAc)1 (NeuAc)1 structure containing core 1 glycan accounted for more than 80% (Fig. 3) among all peaks observed, with sialic acid adding to the reducing end of the glycan. Studies show a correlation between sialic acid and saliva viscoelasticity (Gottschalk and Thomas 1961; Ito et al. 1988; Chaudhury et al. 2015), implying that sialic acid could affect salivary viscoelasticity. Consistent with these results, SLG saliva treated with NM, which cleaves α-2,3 and α-2,6 sialic acids, showed that spinnbarkeit was markedly reduced by 23.7% and 16.6% in young and aged mice, respectively. However, spinnbarkeit was comparable in young and aged SLG saliva after treatment (Table), suggesting that sialic acid plays a pivotal role in providing viscoelasticity to saliva. SIAT4-A, encoded by St3gal1, is an enzyme that transfers sialic acid to the reducing end of glycan in the core 1 structure, and SIAT4-A was considered a candidate protein highly expressed in SLG. Six types of St3gal genes have been documented, from St3gal1 to St3gal6. Of these, St3gal1 and St3gal4 showed the highest expression in salivary glands of mice (Kono et al. 1997). Consistent with this report, lectin probe staining showed that MAL II, specific for α-2,3 sialic acid, clearly stained SLG, while little if any staining was identified in SMG. However, SNA, specific for α-2,6 sialic acid, stained little SLG (Fig. 4A, B). Consistent with qPCR results, SIAT4-A expression decreased at the protein level (Fig. 4D, E), suggesting that lowered St3gal1 expression in aged SLGs causes decreased SIAT4-A expression in aged mice. Notably, the estimation of α-2,3 sialic acid per unit Muc19 protein (MAL II/Muc19) decreased in aged SLG saliva (Fig. 4F, G). Collectively, reduced SIAT4-A in aged mice SLG may lower levels of α-2,3 sialyltransferase activity, with a consequent decrease in saliva viscoelasticity. There are few reports on age-related changes in the rheological properties of saliva (Pushpass et al. 2019). This study is the first to clarify the relationship between aging and structural alterations in mucin. Results showed that SLG, not SMG, is responsible for the rheological properties of saliva, which seem to be closely related to oral microbiota composition (Culp et al. 2015). Thus, alterations in the rheology of saliva may be associated with caries and periodontal disease in the elderly. MUC19 is undetectable in human saliva, and it remains unclear whether MUC19 contributes to gel-forming mucins. If the sialo-glycan structure of mucins in human saliva changes with aging, as seen in Muc19, it may provide new therapeutic strategies for xerostomia in the elderly. Moreover, the prevalence of xerostomia is higher among older females (Han et al. 2015), and SLG displays a stronger sex bias compared to SMG and PG in rodents (Mukaibo et al. 2019). As we used female mice in this study, further study is needed to clarify whether there are sexually dimorphic effects on the rheological properties of saliva.
Author Contributions
M. Yamada, T. Mukaibo, contributed to conception, design, data acquisition, analysis, and interpretation, drafted the manuscript; C. Masaki, contributed to conception, design, data acquisition, analysis, and interpretation, critically revised the manuscript; T. Munemasa, T. Nodai, Y. Kondo, contributed to data acquisition and interpretation, critically revised the manuscript; R. Hosokawa, contributed to conception, design, data acquisition and interpretation, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
sj-pdf-1-jdr-10.1177_00220345221076071 – Supplemental material for Altered Rheological Properties of Saliva with Aging in Mouse Sublingual Gland
Supplemental material, sj-pdf-1-jdr-10.1177_00220345221076071 for Altered Rheological Properties of Saliva with Aging in Mouse Sublingual Gland by M. Yamada, C. Masaki, T. Mukaibo, T. Munemasa, T. Nodai, Y. Kondo and R. Hosokawa in Journal of Dental Research
Footnotes
Acknowledgements
We thank Dr. James E. Melvin for the critical reading of the manuscript and Dr. Takeshi Sato of Nagaoka University of Technology for technical guidance on lectin blot analysis.
A supplemental appendix to this article is available online.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by a grant from the Japan Society for the Promotion of Science KAKENHI (21K09983).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.