
Abstract
Stem and progenitor cells play important roles in the development and maintenance of teeth and bone. Surface markers expressed in bone marrow–derived mesenchymal stem cells are also expressed in dental tissue–derived stem cells. Mouse skeletal stem cells (mSSCs, CD45−Ter119−Tie2−CD51+Thy−6C3−CD105−CD200+) and human skeletal stem cells (hSSCs, CD45−CD235a−TIE2−CD31−CD146−PDPN+CD73+CD164+) have been identified in bone and shown to play important roles in skeletal development and regeneration. However, it is unclear whether dental tissues also harbor mSSC or hSSC populations. Here, we employed rainbow tracers and found that clonal expansion occurred in mouse dental tissues similar to that in bone. We sorted the mSSC population from mouse periodontal ligament (mPDL) tissue and mouse dental pulp (mDP) tissue in the lower incisors by fluorescence-activated cell sorting (FACS). In addition, we demonstrated that mPDL-derived skeletal stem cells (mPDL-SSCs) and mDP-derived skeletal stem cells (mDP-SSCs) have similar clonogenic capacity, as well as cementogenic and odontogenic potential, but not adipogenic potential, similar to the characteristics of mSSCs. Moreover, we found that the dental tissue–derived mSSC population plays an important role in repairing clipped incisors. Importantly, we sorted the hSSC population from human periodontal ligament (hPDL) and human dental pulp (hDP) tissue in molars and identified its stem cell characteristics. Finally, hPDL-like and hDP-like structures were generated after transplanting hPDL-SSCs and hDP-SSCs beneath the renal capsules. In conclusion, we demonstrated that mouse and human PDL and DP tissues harbor dental stem cells similar to mSSCs and hSSCs, respectively, providing a precise stem cell population for the exploration of dental diseases.
Introduction
The development and maintenance of teeth and bone rely on stem or progenitor cells that give rise to multiple stem cell lineages, such as odontoblasts and cementoblasts (Morsczeck and Reichert 2018; Blau and Daley 2019). Dental tissue–derived and bone-derived stem cells have some similarities. For example, both dental pulp–derived stem cells (DPSCs) and bone marrow–derived stem cells (BMSCs) express the same cell markers, such as CD146 and CD73 (Gronthos et al. 2000; Tormin et al. 2011; Breitbach et al. 2018; Walker et al. 2019). Moreover, periodontal ligament stem cells (PDLSCs) and periosteum-derived stem cells (PSCs) express the same cell markers, such as α-SMA (Roguljic et al. 2013; Ortinau et al. 2019). However, single marker-labeled stem cells are highly heterogeneous as the markers are broadly expressed in multiple tissue types in a manner independent of developmental stage (Birbrair 2019). Hence, it is critical to identify a highly pure stem cell population in dental tissues based on multiple cell surface markers.
The CD45−Ter119−Tie2−CD51+Thy−6C3−CD105−CD200+ population was identified as mouse skeletal stem cells (mSSCs) in growth plates of long bone (Chan et al. 2015). This population can give rise to bone, cartilage, or stromal cells and plays an important role in bone development, fractures, distraction osteogenesis, and cartilage regeneration (Tevlin et al. 2017; Mizuhashi et al. 2018; Ransom et al. 2018; Murphy et al. 2020; Ambrosi et al. 2021). Consequently, the CD45−CD235a−TIE2−CD31−CD146−PDPN+CD73+CD164+ population was recently identified as human skeletal stem cells (hSSCs) in growth plates of human fetal bone and plays an important role in repairing bone fractures (Chan et al. 2018). Cell surface markers are similarly expressed in both skeletal and dental tissue–derived mesenchymal stem cell (MSC) populations. Thus, we attempted to sort mSSC and hSSC populations in mouse and human dental tissues and identify their functions.
In this study, a rainbow system was used to evaluate postnatal clonal expansion in dental tissues and bone in vivo to determine whether mesenchymal tissues in the tooth share a common progenitor. In addition, the mSSC and hSSC populations were sorted from mouse and human dental tissues by fluorescence-activated cell sorting (FACS), and the clonogenic capacity and cementogenic, odontogenic, chondrogenic, and adipogenic potential were evaluated. Finally, the ability of dental-derived mSSC populations to repair clipped incisors and of human molar–derived hSSC populations to generate human dental tissue–like structures in vivo was evaluated.
Materials and Methods
Mice
Rosa26-Confetti (Rainbow, Jax, 013731) mice on the C57BL/6J (CD45.2) background were crossed with Ubc-ERT2-Cre (Jax, 008085) mice to obtain Rainbow+/−;Ubc-ERT2-Cre+/− mice for tracing clonal expansion. The mice received an intraperitoneal injection of tamoxifen (Sigma, T5648) at 75 mg/kg body weight at postnatal day 3 (P3) or P35, and tissues were harvested at P35 or P60. Wild-type (WT) C57 mice underwent lower incisor clipping. NPG mice on a nonobese diabetic (NOD) background (Weitonglihua Company) were used for transplantation experiments. For mouse experiments, each group contained 3 mice of the same age with no selection based on sex, and the researchers who performed the analyses were blinded to group allocation in accordance with ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines 2.0. All procedures involving animals and their care were approved by the Medical Ethics Committee of Wuhan University (MLIC202044).
Histological Processing
Samples were fixed in 4% formaldehyde for 12 h at 4°C and then decalcified with 10% (w/v) EDTA for 2 wk. More details are provided in the Appendix Materials and Methods.
Immunohistochemistry and Confocal Microscopy
For immunohistochemistry, sections were incubated with primary antibodies against COL1A1 (Boster, 1:200) and OCN (Boster, 1:200) overnight at 4°C and then with the secondary antibody for 1 h at 37°C. By confocal microscopy, cyan fluorescent protein (CFP) and green fluorescent protein (GFP) could not be detected in Rainbow mice because genetic recombination occurs less frequently, as previously reported (Newton et al. 2019). Yellow fluorescent protein (YFP) and red fluorescent protein (RFP) were detected in sections from our Rainbow mice. More details are provided in the Appendix Materials and Methods.
Isolation of Mouse Dental Tissue–Derived mSSCs by FACS
Mouse mandibles were separated from P30 to P35 or P60 mice (3–5 mice comprised 1 pooled sample). Cell suspensions were acquired from mouse periodontal ligament (mPDL) and mouse dental pulp (mDP) tissues in lower incisors by digestion in 2 mg/mL collagenase type I (SCR103; Sigma) at 37°C for 30 min under constant agitation. Then, the cells were stained with anti-mouse antibodies and analyzed with a BD LSRFortessa X-20 or sorted with a BD FACSAria III. More details are provided in the Appendix Materials and Methods.
Isolation of Human Dental Tissue–Derived hSSCs
Normal impacted third molars (n = 20) were collected from 20 individuals aged 19 to 29 y at the School and Hospital of Stomatology, Wuhan University, following the guidelines approved by the Medical Ethics Committee of School & Hospital of Stomatology, Wuhan University (2020/B52). Human periodontal ligament (hPDL) tissues were gently separated from the surface of the root using a blade, and human dental pulp (hDP) tissues were separated from molars broken by a hammer. Cell suspensions were acquired by digestion in 2 mg/mL collagenase type I (SCR103; Sigma) at 37°C for 30 min under constant agitation. Then, the cells were stained with anti-human antibodies and analyzed or sorted by flow cytometry. More details are provided in the Appendix Materials and Methods.
Clonogenicity and Osteogenic, Chondrogenic, and Adipogenic Potential
Clonogenicity was evaluated by staining with 0.1% crystal violet. Osteogenesis was evaluated by Alizarin red staining, chondrogenesis was evaluated by Alcian blue staining, and adipogenesis was evaluated by Oil red O staining. More details are provided in the Appendix Materials and Methods.
Incisor Clipping Experiments
The left lower incisors of P30 mice were clipped by scissors, and the right incisors were untreated as the controls. More details are provided in the Appendix Materials and Methods.
Real-Time Quantitative Polymerase Chain Reaction
Real-time quantitative polymerase chain reaction (PCR) was performed using a SYBR PCR Master Mix Kit (Vazyme). More details are provided in the Appendix Materials and Methods.
Single-Cell RNA-Sequencing Data Analysis
Single-cell RNA-sequencing (RNA-seq) data (GSE146123) were analyzed by dimplot and the featureplot function of the Seurat package to identify clusters and evaluate gene expression.
Transplantation of hPDL and hDP Skeletal Stem Cells
hPDL skeletal stem cells (SSCs) or hDP-SSCs were sorted by FACS, and 5 × 105 cells at passage 1 were resuspended in 2 µL Matrigel (Corning, CB40234A) and then transplanted using a plunger beneath the renal capsule of 8 weeks old immunodeficient NPG mice under anesthesia with isoflurane. The mice recovered in a new cage with a homeothermic blanket and antibiotic-containing water. These procedures were performed in accordance with the specifications of an approved animal protocol (Medical Ethics Committee of School & Hospital of Stomatology, Wuhan University, 2020/B52). Four weeks posttransplantation, the tissues were analyzed by Masson and immunohistochemical staining according to the manufacturer’s instructions.
Statistics
Statistical analysis was performed using GraphPad Prism (GraphPad Software). A paired Student’s t test was used to calculate statistical significance. Values of P < 0.05 are considered to indicate statistical significance; *P < 0.05, **P < 0.01, and ***P < 0.001 are used to indicate statistically significant differences between 2 groups.
Results
Putative Clonal Expansion in Dental Tissues Resembles That in Bone
Recently, SSCs were identified in growth plates of long bones by rainbow tracers. Due to the similarity of skeletal tissues and dental tissues, we crossed Rainbow mice with Ubc-ERT2-Cre mice to determine whether dental tissues and skeletal tissues show similar clonal expansion (Fig. 1Ai–iii). After tracing from P3 to P35 during tooth development, we observed putative clonal expansion with RFP or YFP in the middle region of lower incisors similar to that in growth plates of the femur (Fig. 1Bi–ii). Tracing from P3 to P60 revealed that clones were distributed in the mPDL and mDP of the lower incisors and molars (Fig. 1Ci–ii). To visualize long-term progenitors in dental tissues, we traced clonal expansion from P35 to P60 and found clones with fewer cells in lower incisors and molars as well (Fig. 1D, E), indicating lower clonal activity in dental tissues upon reaching the adult stage.
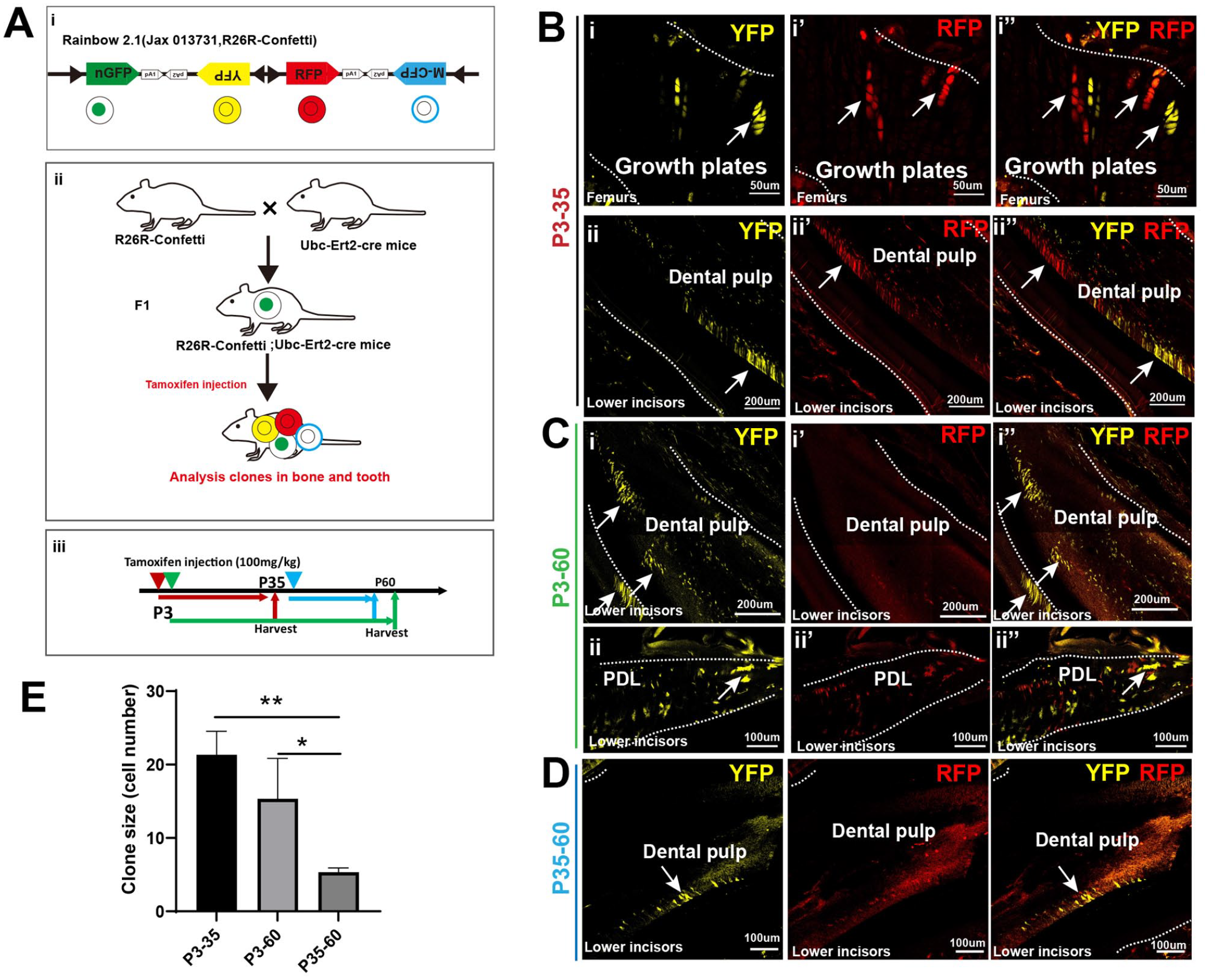
Postnatal clonal expansion visualized in mouse dental tissues was similar to that in bone. (
Identification of Mouse Dental Stem Cells in Lower Incisors That Resemble Putative mSSCs
Previous studies have identified clones in growth plates as mSSCs (CD45−Ter119−Tie2−CD51+Thy−6C3−CD105−CD200+). We aimed to sort this population from mPDL and mDP in lower incisors and identify their functions. Flow cytometry showed that both mPDL and mDP tissues harbor putative mSSC populations similar to those in growth plates of femurs (Fig. 2A). In vitro experiments showed mPDL-SSCs and mDP-SSCs generated other subpopulations and have high clonal ability compared with subpopulations (Appendix Figure 2). Notably, we found that clones generated from both mPDL-SSCs and mDP-SSCs were similar to mSSCs, indicating the clonogenic capacity of mPDL-SSCs and mDP-SSCs (Fig. 2B, C). To investigate differentiation, the osteogenic, chondrogenic, and adipogenic potentials were evaluated. We found that both mPDL-SSCs and mDP-SSCs generated mineralized matrix and cartilage matrix to an extent similar to putative mSSCs (Fig. 2D–G). In contrast, mPDL-SSCs and mDP-SSCs did not have adipogenic potential similar to that of mSSCs (Fig. 2H).
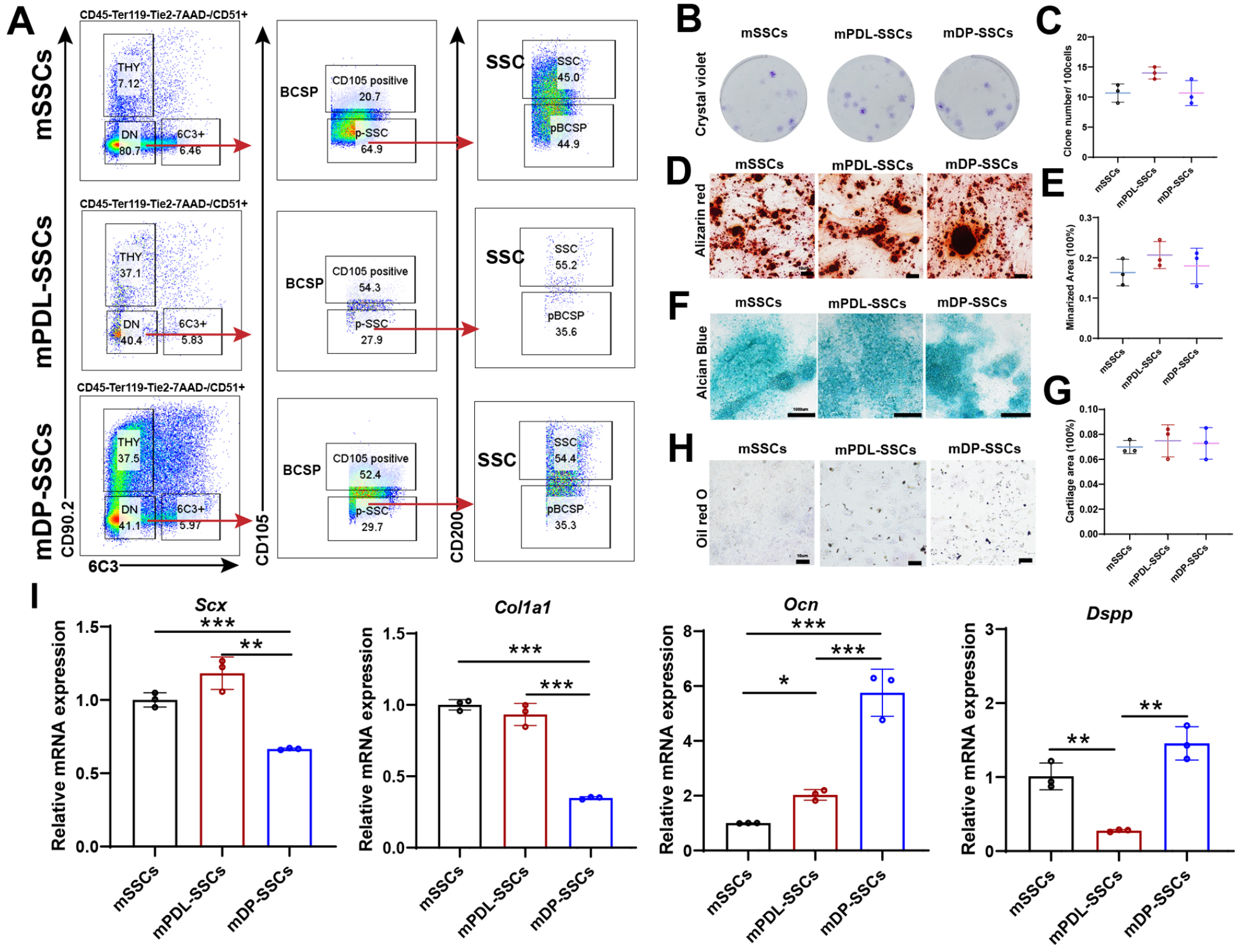
Identification of mouse dental tissue–derived dental stem cells that are similar to mouse skeletal stem cells (mSSCs). (
To verify the tissue specificity of the mSSC populations, we examined the expression of Scx, Col1a1, Dspp, and Ocn in mSSCs, mPDL-SSCs, and mDP-SSCs. We found that the expression of Scx and Col1a1, which have been reported to be highly expressed in periodontal ligament (PDL) tissue and bone (Roguljic et al. 2013; Takimoto et al. 2015; McKenzie et al. 2017), was increased in mPDL-SSCs and mSSCs compared with mDP-SSCs (Fig. 2I). In addition, immunofluorescence showed decreased expression of Col1a1 in mDP-SSCs (Appendix Fig. 3). In contrast, the expression of Dspp and Ocn was increased in mDP-SSCs compared with mPDL-SSCs and mSSCs, which may be related to its function in dentin formation (Suzuki et al. 2009) (Fig. 2I).
Mouse Dental Tissue–Derived Putative mSSCs Contribute to Repairing Clipped Incisors
Next, we investigated the repair of clipped incisors by the above-identified dental stem cells. First, we characterized CD51+CD200+ cells in mouse incisors based on public single-cell RNA-seq analysis (Krivanek et al. 2020) and found that CD51+CD200+ cells were located in the Gli1+ cluster that identified as dental stem cells (Men et al. 2020) (Fig. 3A–C). To evaluate the change in the percentage of dental stem cells during development, the mPDL-SSC and mDP-SSC populations at P30 and P60 were examined by FACS. We found that the percentage of mDP-SSCs at P60 had a decreasing trend compared with that at P30 (Fig. 3D). Second, we clipped the left lower incisors of P30 mice and then injected EdU for 3 d to verify the proliferative activity of dental cells. We found that the length of EdU+ epithelial cells increased in clipped incisors (Fig. 3E), and the dental tissue–derived mSSC population increased to a greater extent in clipped incisors than in unclipped incisors (Fig. 3F, G). This finding indicates that dental tissue–derived mSSCs play an important role in the repair of clipped incisors.
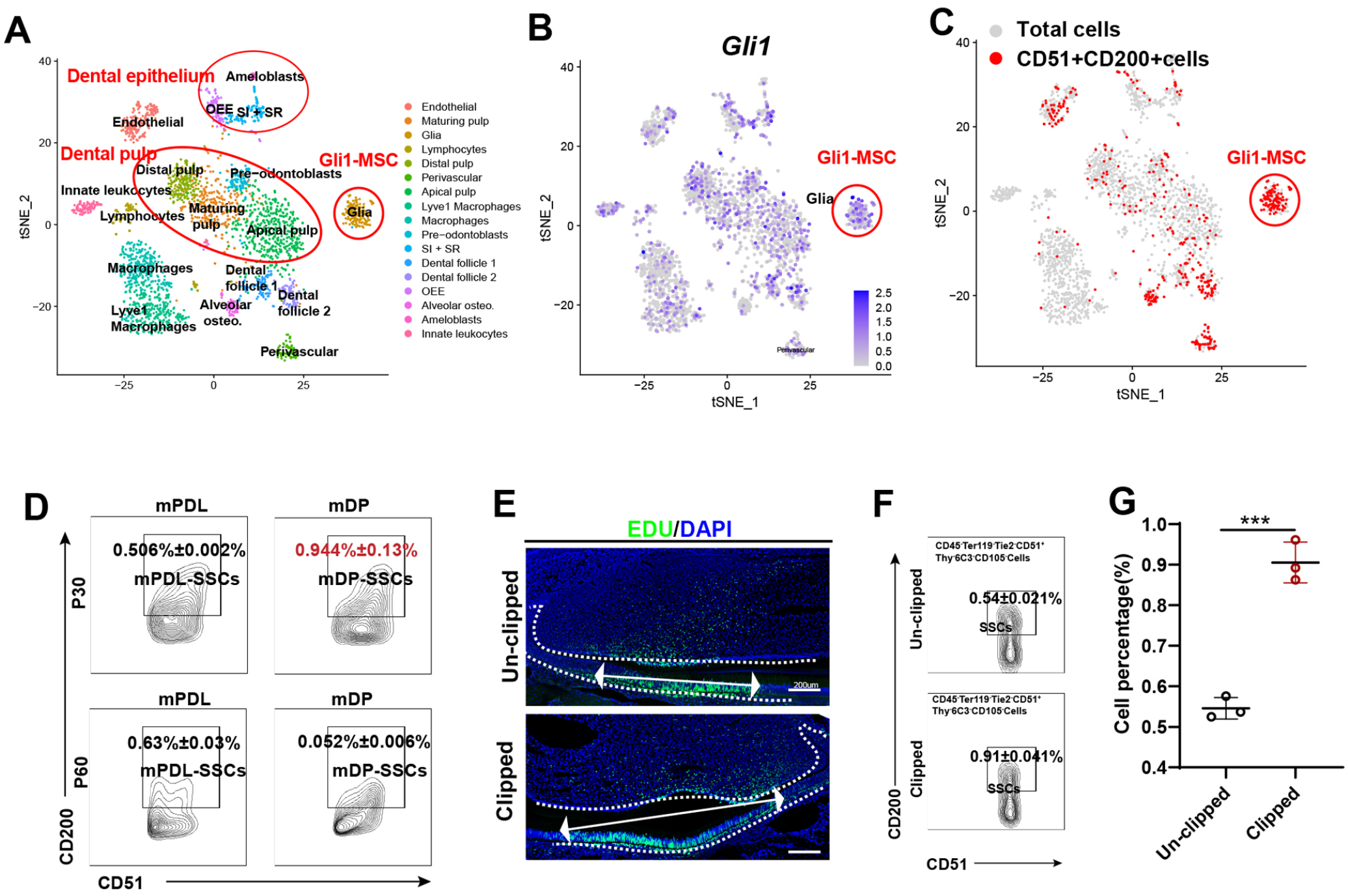
Regeneration of clipped incisors by mouse dental stem cells similar to mouse skeletal stem cells (mSSCs). (
Identification of Human Dental Stem Cells from Molars That Resemble Putative hSSCs
With the identification of hSSCs (CD45−CD235a−TIE2−CD31−CD146−PDPN+CD73+CD164+), we tried to sort this population from human molars. We sorted this population from hPDL and hDP tissue in molars by flow cytometry (Fig. 4A, B). Compared with the cartilage and osteogenic subpopulations, hSSCs and BCSPs (PDPN+CD146+) exhibited high clonal activity (Appendix Fig. 4). By analyzing the proportions of SSCs and the CD146+ population in hPDL and hDP, we found a higher frequency of hPDL-SSCs and hDP-SSCs than the CD146+ cells in molars (Fig. 4C). In addition, the percentage of hSSCs and the CD146+ population were higher in hPDL tissue than in hDP tissue (Fig. 4C).
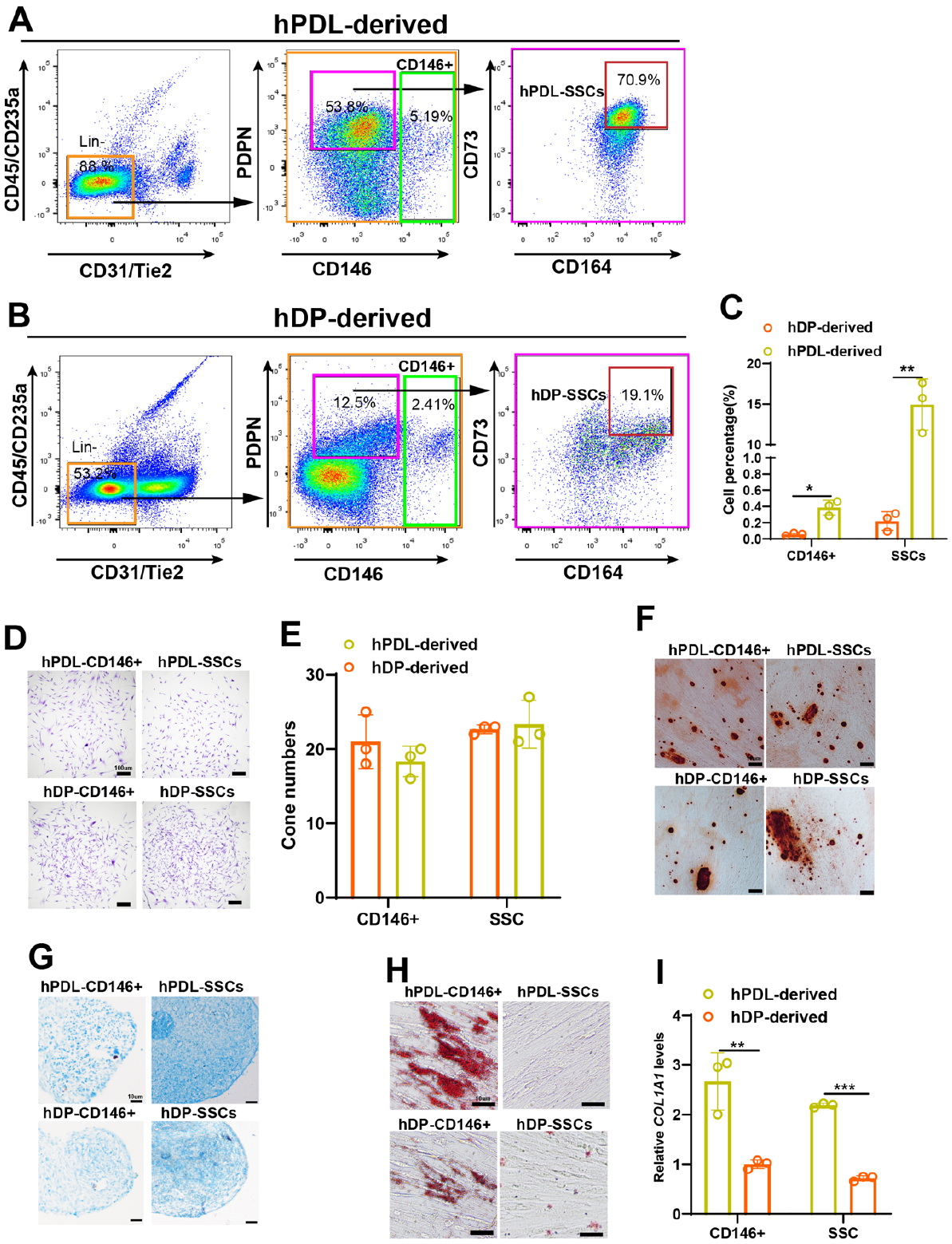
Identification of human dental stem cells similar to putative human skeletal stem cells (hSSCs) from human periodontal ligament (hPDL) and human dental pulp (hDP) tissues in molars. (
Next, we compared hDP-SSCs or hPDL-SSCs with the CD146 population, previously identified as hPDLSCs, and found that they had the same clonogenic capacity (Fig. 4D, E). To test the differentiation potential of these cells, we sorted these populations and cultured them in conditioned medium. We observed that both hPDL-SSCs and hDP-SSCs had osteogenic or chondrogenic potential similar to that of the hPDL- or hDP-CD146+ population but did not have adipogenic ability similar to that of hSSCs (Fig. 4F–H). In addition, the PDL-related gene COL1A1 was increased in hPDL-SSCs and the CD146 population but not in the hDP-derived population (Fig. 4I). Together, these results suggest that hPDL and hDP tissues house the putative hSSC population.
Generation of hPDL-Like and hDP-Like Tissues by Human Dental Tissue–Derived SSCs In Vivo
To verify whether hPDL-SSCs and hDP-SSCs can generate hPDL-like and hDP-like tissues in vivo, we transplanted ex vivo expanded hSSCs into the renal capsule of immunodeficient NPG mice (Fig. 5A). Four weeks after transplantation, hPDL-SSCs generated hPDL-like tissues with collagen fibers and cementum, similar to native PDL tissues from human molars, as assessed by Masson staining (Fig. 5B, C). In addition, we observed the collagen fibers in the hPDL-like tissues were positive for COL1A1 and OCN, similar to native human PDL tissues (Fig. 5B, C). Importantly, we also observed hDP-like tissues and dentin-like structures, similar to native tissues, as shown by Masson staining (Fig. 5D, E). Immunohistochemistry showed the hDP-like tissues were positive for COL1A1, and mature mineralized bone- or dentin-like structures were stained red and were positive for OCN, which was similar to native hDP tissues (Fig. 5D, E).
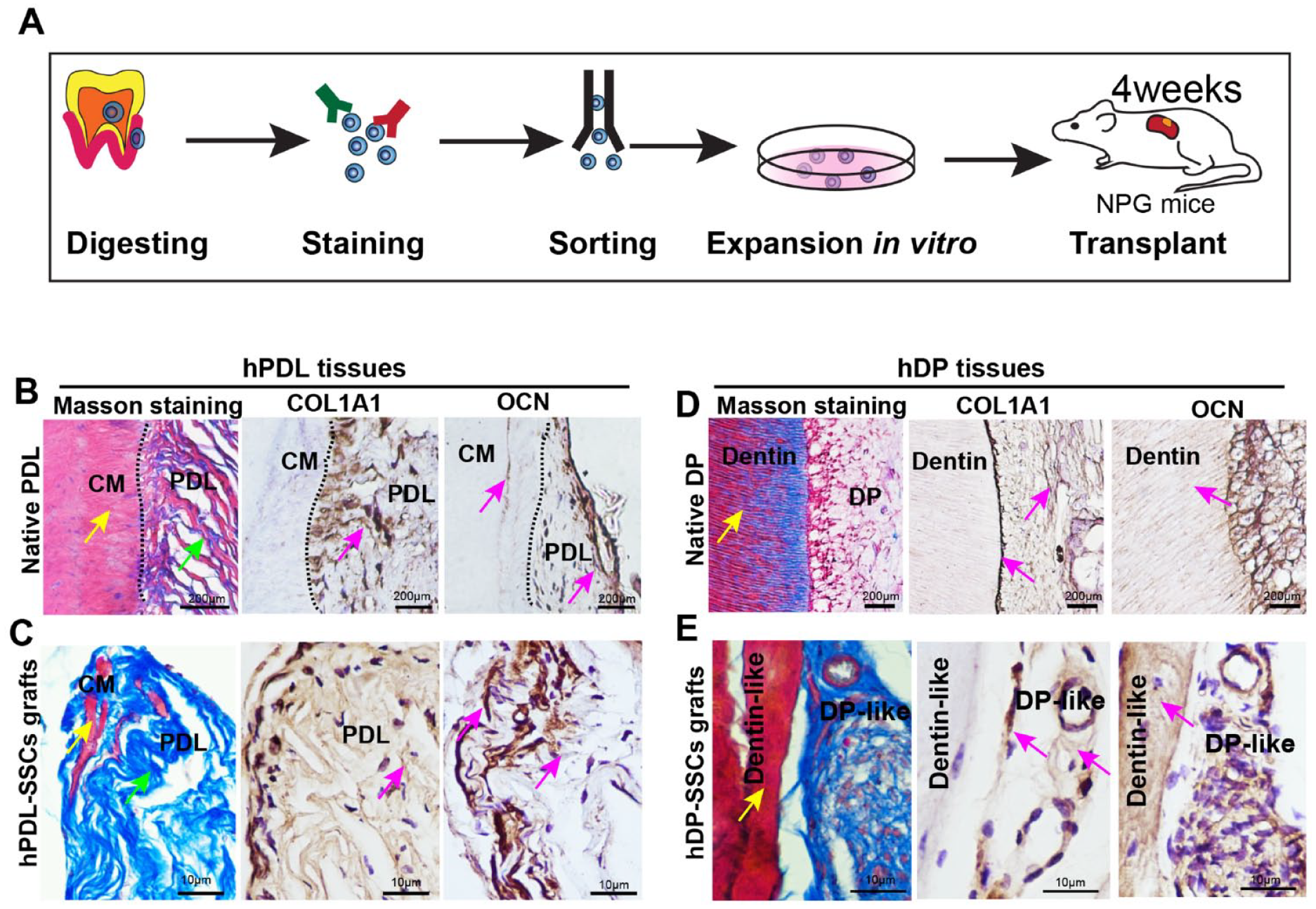
Generation of human periodontal ligament (hPDL)–like and human dental pulp (hDP)–like tissues by human dental stem cells in vivo. (
Discussion
This study identified a putative mSSC population (CD45−Ter119−Tie2−CD51+Thy−6C3−CD105−CD200+) in mPDL and mDP tissues of mouse incisors. This population possesses clonogenic capacity and cementogenic, odontogenic, and chondrogenic potential but not adipogenic potential. In addition, this population plays an important role in repairing clipped incisors in vivo. Importantly, we also identified a putative hSSC population (CD45−CD235a−TIE2−CD31−CD146−PDPN+CD73+ CD164+) in hPDL and hDP tissues of molars. This population possesses clonogenic capacity and cementogenic, odontogenic, and chondrogenic potential, similar to the well-defined CD146+ population, and gives rise to hPDL-like and hDP-like tissues in vivo.
Mouse and human SSCs differ from classic MSCs in the following ways. First, mouse and human SSCs exist in the growth plate of cartilage tissue, but MSCs exist in bone marrow (Chan et al. 2015; Kfoury and Scadden 2015). Second, the surface markers and isolation methods are different. Currently, there are various markers for MSCs, such as Lepr, Cxcl12, and CD73 (Abbuehl et al. 2017; Agarwal et al. 2019; Pineault et al. 2019), while mSSCs are positive for CD200 and CD51, and hSSCs are positive for PDPN, CD73, and CD164. SSCs are sorted by FACS using a combination of surface markers. Third, mouse and human SSCs give rise to bone and cartilage but not fat, which is similar to the population we isolated from dental tissues. Last, the markers used to sort SSCs from humans and mice are different (Chan et al. 2018).
There are some challenges to labeling SSCs using single markers and locating mSSCs. For example, some skeletal stem cell–related genes, such as Pthrp (Mizuhashi et al. 2018), Grem1 (Worthley et al. 2015), and Sox9 (Kuwahara et al. 2019), and dental stem cell–related genes, such as Plp (Kaukua et al. 2014), Ng2 (Feng et al. 2011), Gli1 (Zhao et al. 2014), and Sox2 (Sanz-Navarro et al. 2018), are frequently used to trace stem cells. However, these genes are known to be broadly expressed by multiple tissue types and are almost ubiquitously expressed within skeletal or dental lineages independent of developmental stage (Ambrosi et al. 2019). Frustratingly, precise localization of SSCs is difficult at the histological level because these cells are sorted by a set of 8 cell surface markers. However, we believe that Gli1 is a well-defined stem cell marker demonstrated by single cell RNA-seq in our results and previous genetic lineage tracing (Zhao et al. 2014; Men et al. 2020). In line with our study, a recent study reported that CD200 was expressed in dental follicles and PDL fibroblasts (Takahashi et al. 2019), which supports the notion that dental tissues have an mSSC population.
Mouse SSCs have been shown to reside in different tissues, such as the periosteum and cranial sutures (Debnath et al. 2018; Menon et al. 2021). Therefore, we sorted this population from dental tissues by using mSSC or hSSC markers and found that these cells resided in not only PDL tissue but also DP tissue. Indeed, many stem cell markers were expressed in both bone tissue–derived stem cells and dental-derived stem cells. For example, recent studies have identified α-SMA as a surface marker of periosteal stem cells and of PDL progenitor cells (Roguljic et al. 2013; Debnath et al. 2018). The markers CD105, CD73, and CD90 in bone marrow–derived MSCs are also expressed in dental pulp–derived MSCs (Tsai et al. 2011; Breitbach et al. 2018). In addition, Plp1- or Sox10-ERT2-Cre–derived neuronal glia, NG2-Cre–derived pericytes, and Gli1-expressing cells contribute to pulp cells and odontoblasts in the incisors. All these markers are also widely expressed in bone marrow–derived MSCs (Mendez-Ferrer et al. 2010; Feng et al. 2011; Kaukua et al. 2014; Zhao et al. 2014).
SSCs play an important role in dental repair and regeneration. We found that the proportion of dental tissue–derived mSSCs increased in clipped incisors, indicating that the activity of stem cells was enhanced to repair damaged dental tissues. In bone fractures, the number of mSSCs increased in the callus. In addition, we found that SSCs gave rise to hPDL-like and hDP-like tissues similar to previously identified hPDLSCs and hDPSCs (Gronthos et al. 2000; Seo et al. 2004), indicative of regeneration capacity. However, our hPDL-like tissues were less mature, perhaps due to transplantation time or the lack of hydroxyapatite/tricalcium phosphate ceramic particle induction.
Author Contributions
J.F. Liang, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; J. Wang, contributed to data analysis, drafted the manuscript; Y.T. Ji, L.T. Han, contributed to data acquisition, drafted the manuscript; Q. Zhao, contributed to data analysis, drafted and critically revised the manuscript; R.J. Miron, Y.F. Zhang, contributed to conception and design, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345221084199 – Supplemental material for Identification of Dental Stem Cells Similar to Skeletal Stem Cells
Supplemental material, sj-docx-1-jdr-10.1177_00220345221084199 for Identification of Dental Stem Cells Similar to Skeletal Stem Cells by J.F. Liang, J. Wang, Y.T. Ji, Q. Zhao, L.T. Han, R.J. Miron and Y.F. Zhang in Journal of Dental Research
Footnotes
A supplemental appendix to this article is available online.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Science Foundation of China (82025011).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.