
Abstract
Bone sialoprotein (gene: Ibsp; protein: BSP) is a multifunctional extracellular matrix protein present in bone, cementum, and dentin. Accumulating evidence supports BSP as a key regulator of mineralized tissue formation via evolutionarily conserved functional domains, including a C-terminal integrin-binding Arg-Gly-Asp (RGD) domain implicated in extracellular matrix–cell signaling. Ablation of Ibsp in mice (Ibsp−/−) results in impaired bone growth and mineralization and defective osteoclastogenesis, with effects in the craniofacial region including reduced acellular cementum formation, detachment of the periodontal ligament (PDL), alveolar bone hypomineralization, and severe periodontal breakdown. We hypothesized that BSP-RGD plays an important role in cementum and alveolar bone formation and mineralization, as well as periodontal function. This hypothesis was tested by replacing the RGD motif with a nonfunctional Lys-Ala-Glu (KAE) sequence in (IbspKAE/KAE) mice and OCCM.30 murine (IbspKAE) cementoblasts. The RGD domain was not critical for acellular or cellular cementum formation in IbspKAE/KAE mice. However, PDL volume and thickness were increased, and significantly more tartrate-resistant acid phosphatase–positive osteoclasts were found on alveolar bone surfaces of IbspKAE/KAE mice versus wild type mice. PDL organization was disrupted as indicated by picrosirius red stain, second harmonic generation imaging, dynamic mechanical analysis, and decreased asporin proteoglycan localization. In vitro studies implicated RGD functions in cell migration, adhesion, and mineralization, and this was confirmed by an ossicle implant model where cells lacking BSP-RGD showed substantial defects as compared with controls. In total, the BSP-RGD domain is implicated in periodontal development, though the scale and scope of changes indicated by in vitro studies indicate that other factors may partially compensate for and reduce the phenotypic severity of mice lacking BSP-RGD in vivo.
Introduction
Bone sialoprotein (gene: Ibsp; protein: BSP) is a multifunctional extracellular matrix (ECM) protein of the SIBLING family (small integrin-binding ligand N-linked glycoprotein) present in bone, cementum, and dentin (Ganss et al. 1999; Fisher and Fedarko 2003). Accumulating evidence supports BSP as a key regulator of mineralized tissue formation via evolutionarily conserved functional domains, including an N-terminal collagen-binding domain, polyglutamic (polyE) acid sequences that nucleate hydroxyapatite, and a C-terminal integrin-binding Arg-Gly-Asp (RGD) domain. In BSP, the RGD domain mediates cell attachment and migration via αvβ3 and αvβ5 integrins (Bellahcène et al. 2000; Rapuano et al. 2004; Karadag and Fisher 2006) and modulates osteoblast/osteoclast formation and activity via αvβ3 and the FAK-MAPK (focal adhesion kinase–mitogen-activated protein kinase) signaling pathway (Valverde et al. 2005; Gordon et al. 2009). However, studies on the BSP-RGD domain have been limited to in vitro assays, emphasizing the need for in vivo experiments to investigate physiologic functions.
Ablation of Ibsp in mice (Ibsp−/−) impairs long bone growth and mineralization and osteoclastogenesis (Malaval et al. 2008; Holm et al. 2015). Yet, BSP plays arguably more critical roles in the craniofacial region. As an ECM protein linked to differentiation of cementoblasts and osteoblasts and mineralization of cementum and alveolar bone, BSP is positioned to play important functions in preserving the periodontal complex. Ibsp−/− mice exhibit reduced acellular cementum formation, detachment of the periodontal ligament (PDL), alveolar bone hypomineralization, and severe periodontal breakdown (Foster et al. 2013, 2015; Soenjaya et al. 2015). The mechanisms underlying these dentoalveolar defects remain unclear, in part because the respective contributions of BSP domains remain uncertain.
To test the importance of the BSP-RGD domain in dentoalveolar development and function, the RGD motif was replaced by a nonfunctional Lys-Ala-Glu (KAE) sequence (Bellahcène et al. 2000; Gordon et al. 2009) in mice and OCCM.30 cementoblasts. We hypothesized that BSP-RGD functions in cell-ECM interactions were important in cementum and alveolar bone formation and function.
Materials and Methods
Mice
Animal studies were approved by the Animal Care and Use Committee (National Institutes of Health). Previous analyses of Ibsp−/− mice used a 129/CD1 mouse genetic background (Malaval et al. 2008; Foster et al. 2013, 2015). The majority of new genetically engineered mouse models are prepared on an inbred standardized C57BL/6 background (Appendix); therefore, Ibsp−/− mice on a C57BL/6 genetic background were engineered (Appendix Fig. 1). For continuity, these mice and appropriate wild type (WT) controls were compared with previously described Ibsp−/− mice on a 129/CD1 background (Foster et al. 2013, 2015). IbspKAE/KAE mice were generated on a C57BL/6 background by CRISPR/Cas9 gene editing to replace the RGD domain of BSP with a nonfunctional KAE sequence (Appendix Fig. 2). New mouse models were generated by Taconic Biosciences and sequenced to confirm deletion/substitution. Mice were euthanized, and mandibles were harvested at 10 d postnatal (dpn; active Ibsp mRNA expression in cementoblasts and alveolar bone osteoblasts), 14 dpn (prior to molar eruption), 26 dpn (shortly after first molar eruption), and 60 dpn (>1 mo occlusion). Ibsp mRNA was analyzed at 10 dpn, an active period of periodontal development when cementoblasts and osteoblasts highly express Ibsp (MacNeil et al. 1995). Additional details are in the Appendix.
Micro–computed tomography
Hemimandibles for micro–computed tomography were fixed in 10% neutral buffered formalin (n = 3 to 8/genotype), scanned in a µCT 50 scanner (Scanco Medical), and analyzed as previously described (Chavez et al. 2021). Additional details are in the Appendix.
Histology
Hemimandibles for histology were fixed in Bouin’s solution for 24 h, decalcified in an acetic acid–formalin–sodium chloride solution, and paraffin embedded for serial 5-μm coronal sections (Foster et al. 2015). Hematoxylin and eosin, tartrate-resistant acid phosphatase (TRAP), picrosirius red, immunohistochemistry, and in situ hybridization stains (n = 3 to 5/genotype) were described previously (Foster et al. 2018).
Second Harmonic Generation Microscopy
Hemimandibles for second harmonic generation (n = 3/genotype) were fixed in 2% paraformaldehyde and 2% glutaraldehyde in phosphate buffer for 24 h and plastic embedded for sawing to the first molar mesial root by a low-speed diamond saw (Bueheler; Naveh et al. 2018). Additional details are in the Appendix.
Dynamic Mechanical Analysis
Unfixed hemimandibles (26 dpn; n = 6 or 7/genotype) underwent dynamic mechanical analysis to measure static stiffness (K), hysteresis (W), dynamic stiffness (K*), elastic stiffness (K′), loss stiffness (K′′), and energy dissipation (tan δ), as previously described (Zhang et al. 2020). Additional details are in the Appendix.
Cell Culture
A homozygous Ibsp knock-in cementoblast cell line (IbspKAE), where the RGD motif was replaced by a nonfunctional sequence, was generated from the well-established immortalized murine cementoblasts (OCCM.30) as a WT parent strain (D’Errico et al. 1999) with CRISPR/Cas9 at the Transgenic Core Facility (National Heart, Lung, and Blood Institute, National Institutes of Health). Ibsp knockout cells (IbspΔNterm; Ao et al. 2017) were used as a comparison. Additional details are in the Appendix and in Appendix Figure 3. Migration, adhesion, and mineralization assays are detailed in the Appendix and in Appendix Figure 4.
Ossicle Implant Model
An ossicle implant model tested the ability of cells to promote in vivo mineral nodule formation (Ao et al. 2017) in collagen-elastin sponges implanted subcutaneously in SCID mice (NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJ; n = 4/genotype). Additional details are in the Appendix.
Statistical Analysis
Results are expressed as mean ± standard deviation. Data were analyzed with t test for independent samples or 1-way analysis of variance with post hoc Tukey test (Prism 7.04; GraphPad Software). For all tests, α = 0.05.
Results
Altered Periodontal Structures in the Absence of the BSP-RGD Motif
Previous data on global BSP ablation used Ibsp−/− mice on a mixed 129/CD1 genetic background (Malaval et al. 2008; Foster et al. 2013, 2015). Ibsp−/− and IbspKAE/KAE mice described here were generated on a C57BL/6 genetic background to provide an established inbred mouse model. Given reports that genetic background affects mouse phenotypes (Iwaniec et al. 2006; Mukhopadhyay et al. 2012) and prior to focused studies on the role of the BSP-RGD motif in modulating the periodontia, we comparatively analyzed the dental and periodontal effects of Ibsp knockout on both genetic backgrounds at 60 dpn. In brief, when compared with WT controls, Ibsp−/− mice on a C57BL/6 background exhibited reduced acellular cementum and PDL detachment, significantly reduced alveolar bone proper (ABP) volume, and incisor alterations associated with defective eruption, but they had an overall milder mineralization phenotype than Ibsp−/− mice on a 129/CD1 background. Additional details are in the Appendix and in Appendix Figures 5 and 6.
To define the functions of the BSP-RGD motif, we genetically edited the RGD domain to be replaced by the nonfunctional KAE motif and analyzed homozygous IbspKAE/KAE mice on the C57BL/6 background. No differences were found in enamel or dentin/cementum volumes or densities by micro–computed tomography analyses of molars in IbspKAE/KAE versus WT mice at 14, 26, or 60 dpn (Appendix Fig. 7). In periodontal tissues, mandibular bone and ABP volume and density were not altered in IbspKAE/KAE mice (Fig. 1A–E). However, PDL volume was significantly increased in IbspKAE/KAE versus WT at 26 and 60 dpn (10% and 18%, respectively; P < 0.01 and P < 0.001), mirrored by 20% increased PDL thickness at 60 dpn (P < 0.05). Notably, IbspKAE/KAE mice did not share incisor alterations with Ibsp−/− mice, with a 5% reduction in incisor volume (P < 0.01) and 85% reduction in incisor pulp volume (P < 0.0001; Appendix Fig. 5), but rather showed normal overall volume and pulp volume at 60 dpn (Fig. 1F).
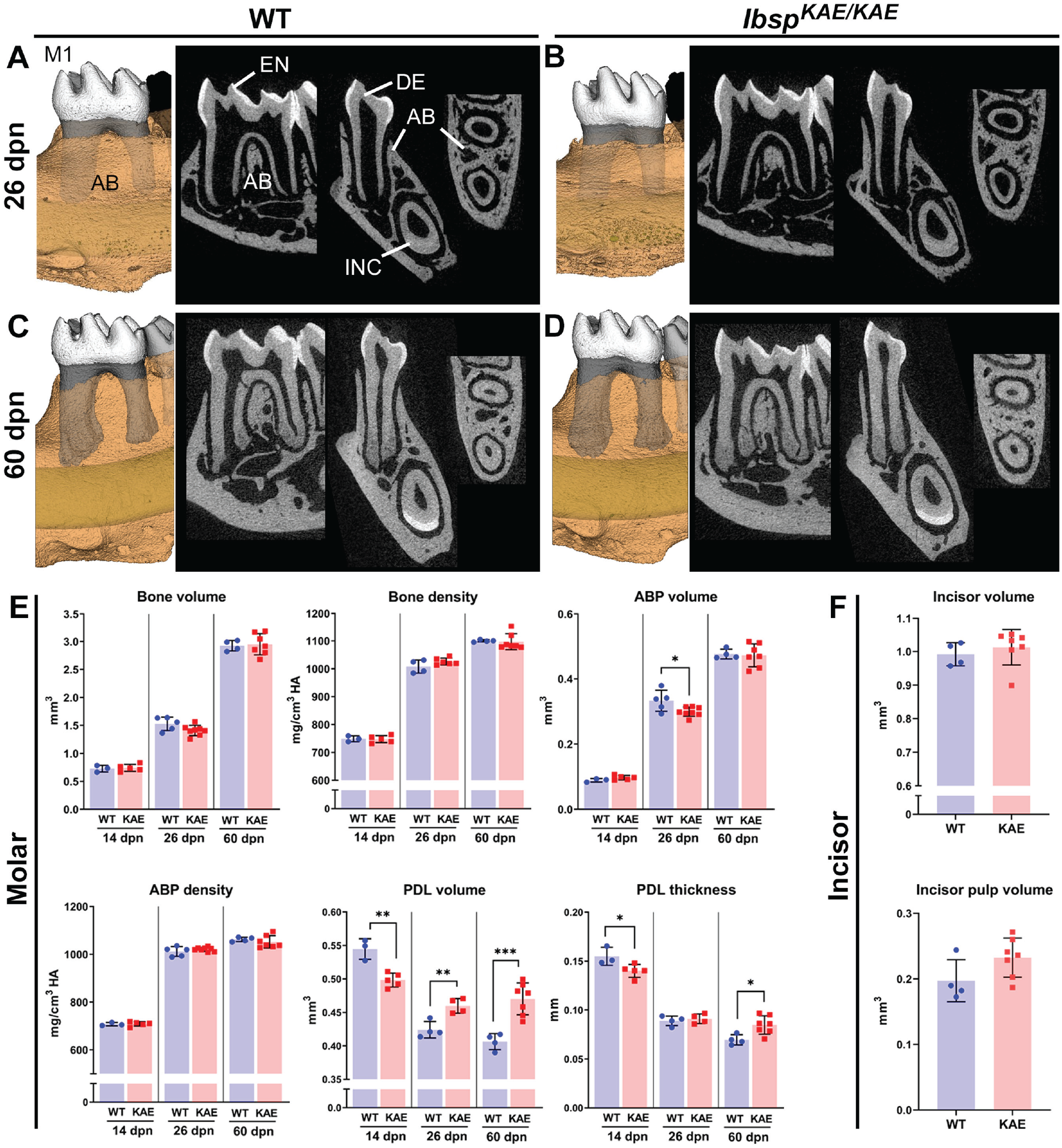
Altered periodontal structures in the absence of the BSP-RGD motif. (
In situ hybridization at 10 dpn during active periodontal formation indicated unchanged numbers and density of Ibsp+ cementoblasts on root surfaces and osteoblasts on alveolar bone surfaces (Fig. 2A–C). Immunohistochemistry at 60 dpn confirmed normal localization of BSP in mature and functional cementum and alveolar bone of IbspKAE/KAE versus WT mice (Fig. 2D). Histology showed normal tissue morphology, including no acellular or cellular cementum defects and normal PDL attachment in IbspKAE/KAE mice (Fig. 2E–H).
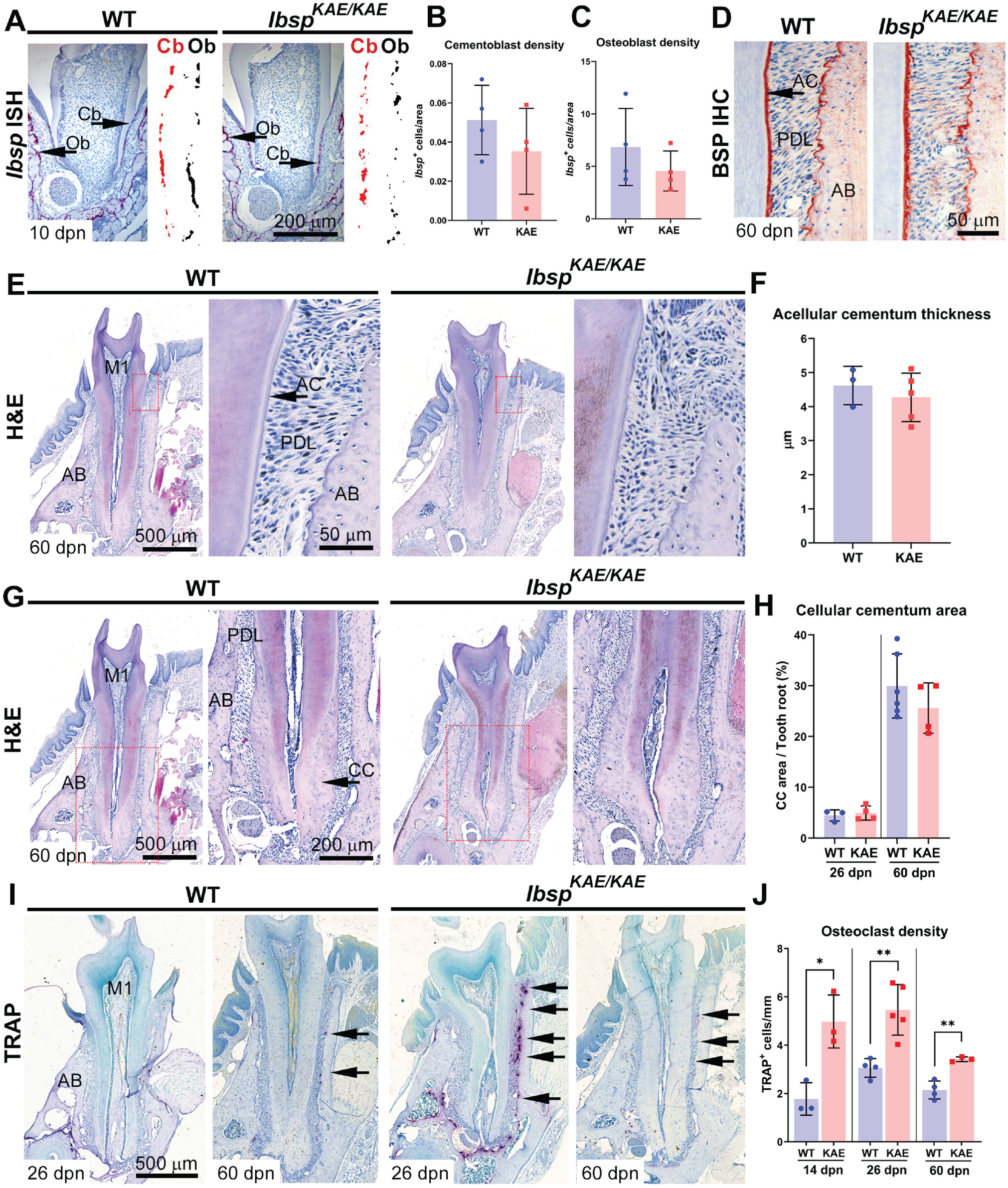
Normal cementum development but increased osteoclast numbers in the absence of BSP-RGD. (
Altered PDL volume and thickness in IbspKAE/KAE mice prompted us to analyze tissues for the presence of TRAP+ osteoclast-like cells that may promote alveolar bone resorption. TRAP staining revealed a 60% to 180% increase in osteoclast cell density on the alveolar bone surfaces of IbspKAE/KAE versus WT mice at 14 (P < 0.05), 26 (P < 0.01), and 60 dpn (P < 0.01; Fig. 2I, J). Osteoclast/odontoclast cells were not observed along tooth root surfaces from IbspKAE/KAE or WT mice.
Altered PDL Structure and Mechanical Properties in the Absence of the BSP-RGD Motif
Altered PDL parameters in IbspKAE/KAE mice prompted further analysis of the PDL region. When compared with WT well-organized PDL collagen fibers, with insertion of Sharpey’s fibers into cementum, IbspKAE/KAE mice had disorganized PDL fiber orientation at 14, 26, and 60 dpn (Fig. 3A). Second harmonic generation microscopy, used to image fibrillar collagen under confocal microscopy, revealed highly organized PDL collagen fibrils (green signal) in 26-dpn WT, while IbspKAE/KAE tissues displayed relatively thin, sparse, and disoriented fibrils (Fig. 3B).
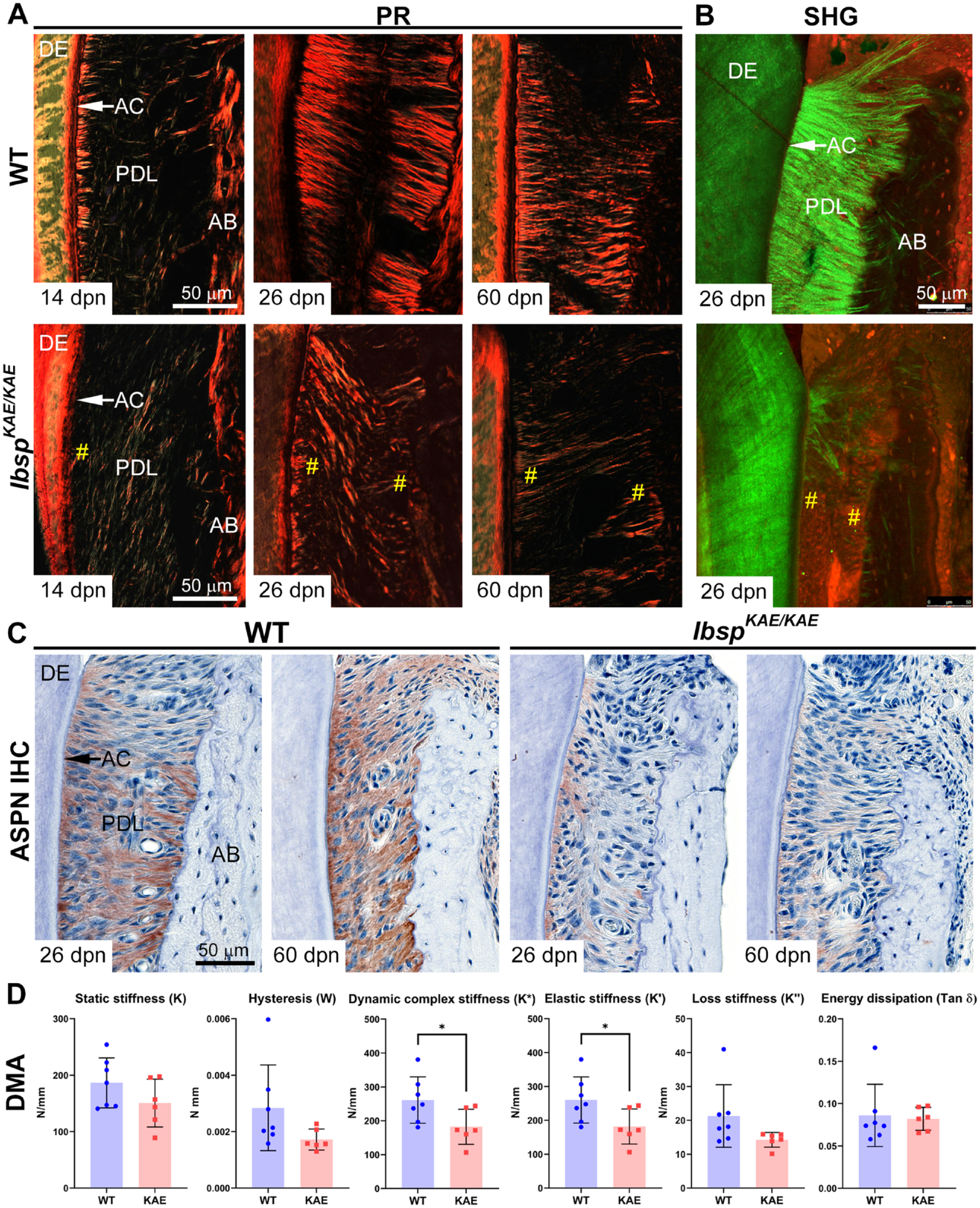
Altered periodontal ligament (PDL) structure and mechanical properties in the absence of the BSP-RGD motif. (
Immunohistochemistry for a panel of PDL protein and proteoglycan markers revealed decreased staining for asporin within the PDL, with slightly higher expression near the root, in IbspKAE/KAE versus WT at 26 and 60 dpn (Fig. 3C). Localization and abundance of decorin, lumican, biglycan, and periostin appeared unchanged in IbspKAE/KAE versus WT periodontal tissues (Appendix Fig. 8).
Given the altered structure and composition of IbspKAE/KAE versus WT periodontal tissues, we performed dynamic mechanical analysis to test mechanical properties of the PDL. When compared with WT, IbspKAE/KAE mice exhibited significantly lower dynamic stiffness (K*) and elastic (storage) stiffness (K′; P < 0.05 for both), while other dynamic mechanical analysis parameters were not significantly different (Fig. 3D).
The RGD Domain of BSP Regulates In Vitro Cell Migration, Adhesion, and Ossicle Mineralization
Altered PDL structure, composition, and mechanical properties associated with lack of the BSP-RGD motif prompted us to investigate in vitro cell functions. IbspKAE OCCM.30 immortalized murine cementoblasts carrying the inactivating RGD mutation were compared with previously described OCCM.30 cells harboring a large N-terminal deletion (IbspΔNterm; Ao et al. 2017). While no changes were noted in cell proliferation at 24 h between cell types, IbspKAE and IbspΔNterm cementoblasts exhibited a 30% to 40% reduced migration rate, with no significant difference between them, as compared with WT cells (Fig. 4A, B).
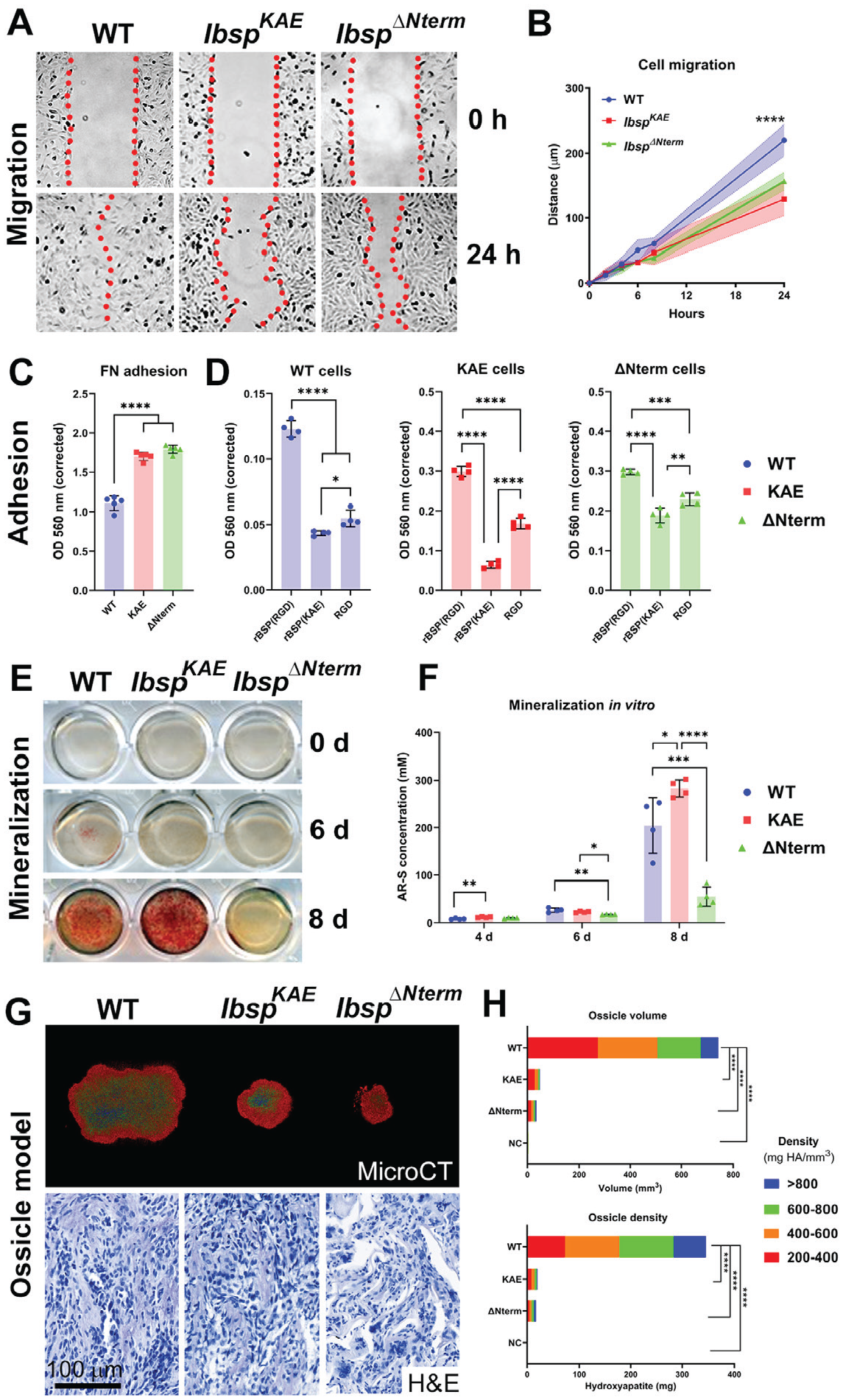
The RGD domain of BSP regulates in vitro cell migration and adhesion and in vivo mineralization. IbspKAE OCCM.30 immortalized murine cementoblasts carrying the inactivating RGD mutation were compared to wild type (WT) cells and cells harboring a large N-terminal deletion (IbspΔNterm). (
The ability of cells to adhere to dishes precoated with the RGD motif–containing ECM protein, fibronectin (FN) indicated that IbspKAE and IbspΔNterm cells had greater sensitivity to and increased adherence on FN versus WT cells (Fig. 4C). Because of differences in cell-ECM adhesion to FN, cells were tested on dishes precoated with recombinant BSP (rBSP), rBSP-KAE, or RGD peptide. WT, IbspKAE, and IbspΔNterm cells all showed significantly less adhesion to rBSP(KAE) versus rBSP(RGD; P < 0.0001 for all comparisons). All cells showed relatively greater adhesion to RGD versus rBSP(KAE), though IbspKAE showed the greatest sensitivity and response versus the other cell types (Fig. 4D).
IbspKAE cells promoted mineral nodule formation equivalent to WT cells, while IbspΔNterm cells exhibited 70% reduced mineralization capacity by alizarin red staining (Fig. 4E, F). The in vivo ossicle implant model revealed that IbspKAE and IbspΔNterm cells produced smaller ossicles versus WT cells (P < 0.0001; Fig. 4G). No differences were detected between IbspKAE and IbspΔNterm implants. Negative controls (no cells) exhibited minimal volume and mineral density. Hematoxylin and eosin staining of decalcified nodules showed similar distribution of cells and ECM in WT and IbspKAE cementoblasts, while IbspΔNterm cells were present in a matrix appearing like osteoid/cementoid, as previously described (Ao et al. 2017).
Discussion
BSP plays a key role in bone homeostasis and is essential for cementum formation and periodontal function. We examined the functional importance of the BSP-RGD domain with multimodal in vivo and in vitro approaches, replacing RGD with a nonfunctional KAE sequence. While the RGD domain was not critical for cementum or alveolar bone mineralization, periodontal structure, organization, composition, and function were affected in mice (Appendix Tables 1 and 2). In vitro studies implicated RGD functions in cell migration, adhesion, and mineralization, as confirmed by an ossicle implant model where cells lacking BSP-RGD showed substantially defective mineralization versus controls. In total, the BSP-RGD domain is implicated in periodontal development, though in vitro studies indicate that other factors may partially compensate for and reduce the phenotypic severity of mice lacking BSP-RGD.
Functions of the BSP-RGD Motif in Dentoalveolar Development
The most striking dentoalveolar effect of global BSP knockout in mice is the lack of functional acellular cementum, causing PDL detachment and severe periodontal breakdown, also contributing to incisor malocclusion in mice on a 129/CD1 background (Foster et al. 2013, 2015; Soenjaya et al. 2015) and with a milder phenotype on the C57BL/6 background analyzed here. Key initial findings in mice lacking the BSP-RGD domain were presence of acellular cementum, with no observed PDL detachment, periodontal destruction, or incisor malocclusion. These collective observations suggest that RGD-associated functions are not responsible for the major cementum and alveolar bone hypomineralization phenotypes arising from BSP ablation, thereby pointing to other BSP domains, including the N-terminal hydrophobic collagen-binding motif and polyE mineral-nucleating sequences, with additional potential contributions from phosphorylated serines and glycosylation (Tye et al. 2005; Xu et al. 2017).
While a cementum phenotype was not identified, these studies revealed a vital role for BSP-RGD in periodontal tissues by multiple lines of evidence. As a marker for cementoblasts and osteoblasts contributing to cell differentiation and tissue mineralization, BSP is positioned to function in PDL attachment and periodontal integrity. However, it was surprising that primary changes were detected in PDL organization and composition, rather than in cementum and bone. Analyses revealed decreased PDL volume and thickness prior to tooth eruption in the absence of BSP-RGD. While root formation, alveolar bone remodeling, and organization of the PDL from the dental follicle are understood to be intimately connected developmental processes (Radlanski et al. 2015), little is known about cellular regulators—for example, how dental follicle cell fates are determined to contribute to cementoblasts, PDL fibroblasts, or alveolar bone osteoblasts. These early PDL differences may reflect altered differentiation, migration, and/or accumulation of dental follicle cells to root and/or bone surfaces. Conversely, we found increased PDL parameters at later ages and decreased ABP in the absence of BSP-RGD. Increased numbers of osteoclasts were present on alveolar bone surfaces of IbspKAE/KAE versus WT mice, and osteoclast resorption likely contributes to the observed PDL and ABP changes. Though the PDL was structurally altered, IbspKAE/KAE maintained periodontal attachment, structure, and function, within the limits of this study. Ibsp−/− and IbspKAE/KAE mice share the presence of increased numbers of osteoclasts on alveolar bone, but they differ in that Ibsp−/− mice demonstrate reduced osteoclast differentiation and function in long bones (Malaval et al. 2008; Boudiffa et al. 2010), with evidence that BSP contributes to osteoclast differentiation in vitro (Valverde et al. 2005) and in vivo (Valverde et al. 2008). We hypothesize that altered periodontal structures in mice with BSP knockout or RGD mutation attract and activate more osteoclast precursors, overriding inherent cell-autonomous functional defects. While at later ages, altered periodontal structures in IbspKAE/KAE mice might be a factor in attraction of osteoclasts, increased osteoclast numbers even at 14 dpn, prior to molar eruption, suggest that altered occlusal forces are not primarily responsible. Work is ongoing to analyze the skeletal phenotype of the IbspKAE/KAE mice and investigate effects on osteoclastogenesis in vitro.
Picrosirius red stain and second harmonic generation microscopy revealed reduced organization of PDL in IbspKAE/KAE mice versus controls. Mechanical testing revealed differences at a posteruption age, with lower dynamic and storage stiffness reflecting PDL changes, with additional contributions from disordered PDL organization. Though IbspKAE/KAE teeth show more displacement under loading, they still bear chewing forces and maintain energy dissipation comparable to WT.
Previous studies have highlighted the importance of small leucine-rich proteoglycans (SLRPs) in PDL function and collagen organization, affecting cell mechanosensory functions. Mice genetically ablated for SLRP fibromodulin and biglycan exhibited disrupted PDL collagen fibrillogenesis, overexpression of asporin, hyperactive TGFβ/BMP signaling, and increased numbers of osteoclasts on alveolar bone (Wang et al. 2014). In our studies we noted decreased expression of asporin in IbspKAE/KAE mice versus WT mice in a gradient fashion, with greater localization in proximity to the root surface, while the other SLRPs and periostin were unaltered. Asporin was reported to be a negative regulator of mineralization when overexpressed in PDL cells (Kajikawa et al. 2014), while conflicting reports indicate a positive effect on mineralization (Lee et al. 2011; Xu et al. 2015; Zhan et al. 2019). Asporin overexpression/treatment in fibroblasts was shown to increase collagen calcification and prevent mechanosensory-induced ECM remodeling (Liu et al. 2021). These accumulated findings indicate a role for asporin in cell-ECM interactions, including within the periodontal complex, though further studies are required.
The RGD Domain of BSP Controls Migration and Adhesion of Cementoblasts In Vitro
Periodontal alterations in mice lacking BSP-RGD prompted us to explore functions of the domain in vitro. We mutated BSP-RGD to KAE via CRISPR/Cas9 in a well-established immortalized cementoblast cell line, OCCM.30 cells (D’Errico et al. 1999), comparing them with WT OCCM.30 cells and IbspΔNterm cells featuring a large N-terminal deletion (Ao et al. 2017). Migration and adhesion assays revealed a marked effect of the absence of the RGD domain, suggesting key roles for this domain in cell interactions with the surrounding ECM and environment. IbspKAE cells showed increased adhesion to FN and a hypersensitive response to RGD versus rBSP(KAE). Other conserved motifs in BSP are known to be involved in cell-ECM interactions, most notably the collagen-binding domain (Tye et al. 2005), and IbspΔNterm cells (lacking that domain an additional N-terminal residues) showed altered adhesion to all ECM proteins tested.
IbspKAE cementoblast mineralization appeared undiminished when compared with WT by alizarin red staining, supporting the BSP polyE domains (and possibly other regions) as the primary modulators of mineralization (Goldberg et al. 2001). However, the ossicle implant model indicted severe defects in IbspKAE cells, reflected as reduced ossicle volume and hydroxyapatite deposition. Considering the combined in vivo and in vitro data, these contrasting results do not appear to be from impaired mineralization per se but may implicate altered cell-ECM interactions, such as those highlighted by impaired migration and adhesion assays and suggested by previous studies on the BSP-RGD domain (Gordon et al. 2009). In vivo observations showing reduced PDL at preeruption ages that progressed to increased PDL with deranged organization at later ages fit with this hypothesis. Additionally, caution is always warranted in use of in vitro mineralization assays, as they are subject to numerous variables (Bonewald et al. 2003), with cell density being relevant in functions of Ibsp knockout cells (Bouet et al. 2015).
In summary, we identified the BSP-RGD domain as a contributor to periodontal development and function. BSP-RGD has been implicated in osteoblast differentiation in vitro (Gordon et al. 2009), in line with other studies that point to the biological importance of RGD domains in a variety of proteins and contexts. These studies represent a first step toward understanding functions of BSP-RGD, with additional studies needed to expand beyond the limitations of animal numbers and to identify specific molecular functions in distinct cells and tissues.
Author Contributions
K. Nagasaki, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; M.B. Chavez, A. Nagasaki, contributed to conception, design, data acquisition, analysis, and interpretation, critically revised the manuscript; J.M. Taylor, M.H. Tan, M. Ma, M.E. Thew, contributed to data acquisition and analysis, critically revised the manuscript; E. Ralston, contributed to data acquisition, analysis, and interpretation, critically revised the manuscript; D.-G. Kim, contributed to data analysis and interpretation, critically revised the manuscript; M.J. Somerman, B.L. Foster, contributed to conception, design, data analysis, and interpretation, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345221100794 – Supplemental material for The Bone Sialoprotein RGD Domain Modulates and Maintains Periodontal Development
Supplemental material, sj-docx-1-jdr-10.1177_00220345221100794 for The Bone Sialoprotein RGD Domain Modulates and Maintains Periodontal Development by K. Nagasaki, M.B. Chavez, A. Nagasaki, J.M. Taylor, M.H. Tan, M. Ma, E. Ralston, M.E. Thew, D.-G. Kim, M.J. Somerman and B.L. Foster in Journal of Dental Research
Footnotes
Acknowledgements
We thank Dr. Harvey Goldberg (Western University) and Dr. Larry Fisher (National Institute of Dental and Craniofacial Research/National Institutes of Health [NIH]) for discussions and provision of reagents for these studies, Dr. Chengyu Liu and Dr. Fan Zhang at the Transgenic Core Facility (National Heart, Lung, and Blood Institute/NIH) for their advice and generating the cell lines, and Dr. Davide Randazzo at the Light Imaging Section (National Institute of Arthritis and Musculoskeletal and Skin Diseases/National Institutes of Health) for slide scanning.
A supplemental appendix to this article is available online.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by the Japan Society for the Promotion of Science to A. Nagasaki (JSPS Research Fellowship for Japanese Biomedical and Behavioral Researchers at the National Institutes of Health); by the National Institute of Dental and Craniofacial Research/National Institutes of Health to support for M.B. Chavez (T32DE014320); by the National Institute of Dental and Craniofacial Research/National Institutes of Health to M.B. Chavez (F30DE030358); by the National Institute of Dental and Craniofacial Research/National Institutes of Health to B.L. Foster (R01DE027639); and by intramural funding from the National Institute of Arthritis and Musculoskeletal and Skin Diseases/National Institutes of Health to M.J. Somerman.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.