
Abstract
The metabolism and remodeling of alveolar bone are the most active among the whole skeletal system, which is related to the biological characteristics and heterogeneity of the bone mesenchymal stromal cells (MSCs). However, there is a lack of systematic description of the heterogeneity of MSC-derived osteoblastic lineage cells as well as their distinct osteogenic differentiation trajectory of alveolar bone. In this study, we constructed a single-cell atlas of the mouse alveolar bone cells through single-cell RNA sequencing (scRNA-seq). Remarkably, by comparing the cell compositions between the alveolar bone and long bone, we uncovered a previously undescribed cell population that exhibits a high expression of protocadherin Fat4 (Fat4+ cells) and is specifically enriched around alveolar bone marrow cavities. ScRNA-seq analysis indicated that Fat4+ cells may initiate a distinct osteogenic differentiation trajectory in the alveolar bone. By isolating and cultivating Fat4+ cells in vitro, we demonstrated that they possess colony-forming, osteogenic, and adipogenic capabilities. Moreover, FAT4 knockdown could significantly inhibit the osteogenic differentiation of alveolar bone MSCs. Furthermore, we revealed that the Fat4+ cells exhibit a core transcriptional signature consisting of several key transcription factors, such as SOX6, which are involved in osteogenesis, and further demonstrated that SOX6 is required for the efficient osteogenic differentiation of the Fat4+ cells. Collectively, our high-resolution single-cell atlas of the alveolar bone reveals a distinct osteogenic progenitor that may contribute to the unique physiological characteristics of alveolar bone.
Keywords
Introduction
As the thickened ridge of the jawbone that supports teeth, alveolar bone exhibits a unique developmental trajectory and morphological characteristics compared to the rest of the skeletal system (Rawlinson et al. 2009; Zhou et al. 2018). Alveolar bone originates from the neural crest, forms through intramembranous ossification, and exhibits a compact structure composed of plate-like trabeculae and limited bone marrow. In contrast, the femur originates from mesoderm, forms through endochondral ossification, and exhibits a porotic structure with rod-like trabeculae in the trabecular bone. In addition, due to occlusal stress stimulating change and odontogenic inflammatory response, the metabolism and remodeling of alveolar bone are the most active among the entire skeletal system (Hathaway-Schrader and Novince 2021). Consistent with these unique properties, alveolar bone is more resistant to bone loss and adipocyte accumulation than long bone when suffering from ovariectomy (Mavropoulos et al. 2007). These physiological and pathological differences between alveolar bone and long bone may be attributed to the different biological characteristics of the mesenchymal stromal cells (MSCs) from the 2 skeletal compartments, as it has been demonstrated that craniofacial bone MSCs show a strong proliferation rate and osteogenic differentiation capability but weak chondrogenic and adipogenic differentiation potential (Matsubara et al. 2005; Aghaloo et al. 2010). MSCs are remarkable heterogeneous cell populations that contain many subpopulations that correspond to stromal skeletogenic progenitor and precursor cells (Pittenger et al. 1999; Wolock et al. 2019). However, a systematic description of the heterogeneity of MSCs and MSC-derived osteolineage cells in alveolar bone is lacking. Moreover, whether the cellular composition of MSCs and osteogenic differentiation trajectory in alveolar bone differ from those in long bone has not been assessed.
Single-cell RNA sequencing (scRNA-seq) is a novel technology for high-throughput sequencing of messenger RNA (mRNA) at the single-cell level, which holds great potential for exploring the heterogeneity of MSCs in alveolar bone (Wen and Tang 2016). Recently, the bone marrow microenvironment of long bone has been characterized at single-cell resolution by several studies. The bone marrow stroma of long bone consisted of hematopoietic cells, MSCs, MSC-derived osteolineage cells, chondrocytes, fibroblasts, bone marrow–derived endothelial cells, and pericytes (Baryawno et al. 2019; Tikhonova et al. 2019; Baccin et al. 2020). Among the identified subpopulation of MSCs, several have been demonstrated to exert unique biological functions, such as the skeletal stem cell (Chan et al. 2015), the CTSK+ periosteal stem cell (Debnath et al. 2018), and the Adipoq+Plin1– adipogenic lineage precursors (Zhong et al. 2020). In a recent study, scRNA-seq has also been performed on mouse mandible to map the immune microenvironment in the murine alveolar bone (Lin et al. 2021). However, because of the high abundance of hematopoietic cells, which account for 90% of the whole isolated mandibular cells, the information gained on the nonhematopoietic alveolar bone marrow cells was limited.
In the present study, we aimed to perform an in-depth characterization of the cell types in alveolar bone, to identify the cellular taxonomy of the MSC-derived osteoblastic lineage cells, and to assess whether the alveolar bone contains unique osteoblastic subpopulations. To achieve these goals, we purified alveolar bone cells using fluorescence-activated cell sorting (FACS) followed by progressive depletion of the hematopoietic cells and generated the single-cell atlas of the nonhematopoietic cells in alveolar bone by performing scRNA-seq. Importantly, we revealed a previously undescribed Fat4+ cell population that is specifically enriched in the osteoblastic lineage cells of alveolar bone but not in long bone. We further demonstrated that the Fat4+ cells possess progenitor properties, with the core transcriptional features regulating osteogenic differentiation, which thus may represent a mesenchymal osteogenic progenitor population of the alveolar bone. This work sheds new light on the current understanding of alveolar bone remodeling in both physiological and pathological contexts.
Materials and Methods
Methods are described in the Appendix.
All animal experimental procedures were approved by the Animal Care Committee at Shanghai Ninth People’s Hospital, Shanghai Jiao Tong University School of Medicine (Shanghai, China) and performed according to institutional guidelines and the ARRIVE guidelines (Animal Research: Reporting In Vivo Experiments). Alveolar bones from 6- to 8-wk-old male C57 mice were harvested for the scRNA-seq generated by the 10X Genomics Chromium Controller Instrument and Chromium Single Cell 3′ V3 Reagent Kits (10X Genomics). The scRNA-seq data for long bone from 2 previously published studies (Baryawno et al. 2019; Baccin et al. 2020) were downloaded from GEO (GSE128423 and GSE122467). Integration analysis was according to a standard Seurat integration workflow (Stuart et al. 2019).
Results
Characterization of Alveolar Bone Single-Cell Atlas
To explore the cellular composition of the mouse alveolar bone, single-cell suspensions from C57BL/6 mice alveolar bone were prepared with enzymatic digestion followed by sorting viable CD45–Ter119– nonhematopoietic alveolar bone marrow cells. Sorted cells were profiled by droplet-based scRNA-seq (Fig. 1A). After filtering for the number of expressed genes and percentages of mitochondrial genes, a total of 5,273 cells with a median number of RNA features of 2,904 were selected for downstream analysis. Unbiased clustering of scRNA-seq profiles identified 20 clusters that exhibit distinct transcriptional signatures, including endothelial cells, skeletogenic cells, pericytes, neurons, and glia. Importantly, only ~12% of the cells in our data set were hematopoietic cells such as erythroid, T cells, and B cells, far less than the ~98% hematopoietic cells from a previously published alveolar bone scRNA-seq study (Lin et al. 2021), demonstrating a much more efficient enrichment of nonhematopoietic cells in alveolar bone (Fig. 1B, Appendix Figs. 1–2A).
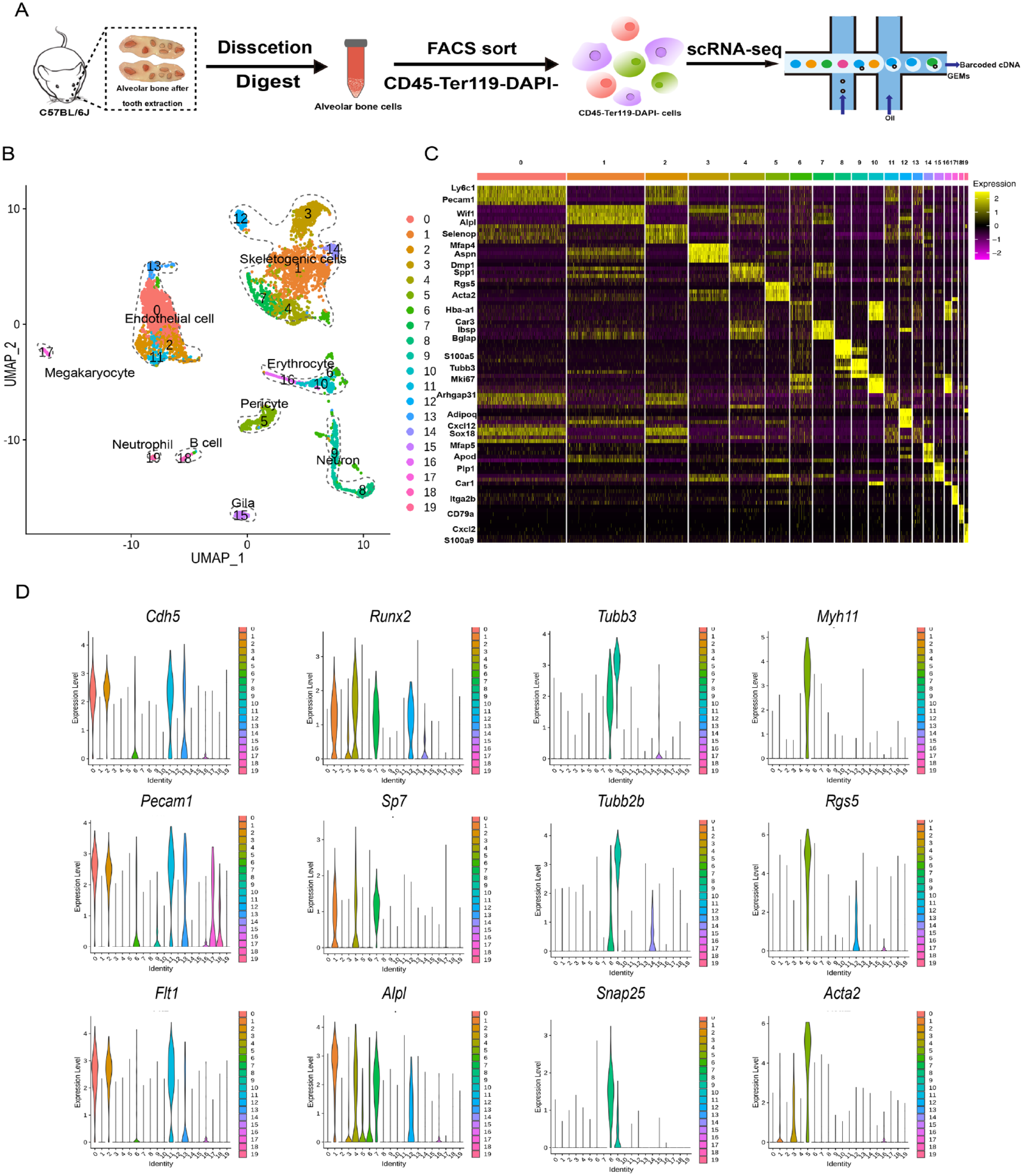
A single-cell atlas of alveolar bone. (
We characterized the most significant gene expression signatures associated with each cluster (Fig. 1C). In particular, the marker genes of the 4 major cell types in alveolar bone were 1) Cdh5, Pecam1, and Flt1i for endothelial cells (Rafii et al. 2016); 2) osteoblastic differentiation factors, such as Runx2, Sp7, and Alpl, for skeletogenic cells; 3) Myh11, Rgs5, and Acta2 for pericytes (Armulik et al. 2011); and 4) Tubb3, Tubb2b, and Snap25 for neurons (Jaglin et al. 2009) (Fig. 1D). Importantly, the skeletogenic cell population consisted of 6 closely related but distinct clusters, which may correspond to different transitional states during skeletogenesis (Fig. 1B–D). Our single-cell atlas of the alveolar bone cells provided unprecedented opportunities for unveiling the molecular underpinnings for the unique characteristics of the alveolar bone.
An Osteoblastic Lineage Cell Subpopulation Highly Expressing Fat4 Is Specifically Enriched in Alveolar Bone Skeletogenic Cells
To assess whether the cellular composition and/or transcriptional signatures of the skeletogenic cells in alveolar bone differ from those in long bone, we retrieved 2 previously published scRNA-seq data sets of long bone that similarly sorted for CD45–Ter119– nonhematopoietic cells to our study (Baryawno et al. 2019; Baccin et al. 2020) and integrated them with our alveolar bone data for comparative analysis. We selected the 7 alveolar bone clusters representing skeletogenic cells and pericytes, which may also play a part in skeletogenesis, for the integrated analysis (Fig. 1B). Unsupervised graph clustering on integrated skeletogenic cells generated 21 distinct clusters, which we further classified into 4 major cell types based on known marker genes used in previous scRNA-seq analyses on long bone (Fig. 2A, Appendix Fig. 2A, B). Although all 4 major cell types were present in both the alveolar bone and long bone, their fractions within the skeletogenic cell population varied greatly between the 2 bone types. In long bones, the proportions of osteoblastic and chondrocydic lineage cells were comparable. In contrast, the osteoblastic lineage cells and chondrocydic lineage cells accounted for ~70% and 7% of the skeletogenic cell population in alveolar bone, respectively (Fig. 2B, Appendix Fig. 2C). Such a prominent enrichment of osteogenic cells and depletion of chondrogenic cells was in accordance with the fact that alveolar bone is mainly formed through intramembranous ossification instead of the endochondral ossification process.
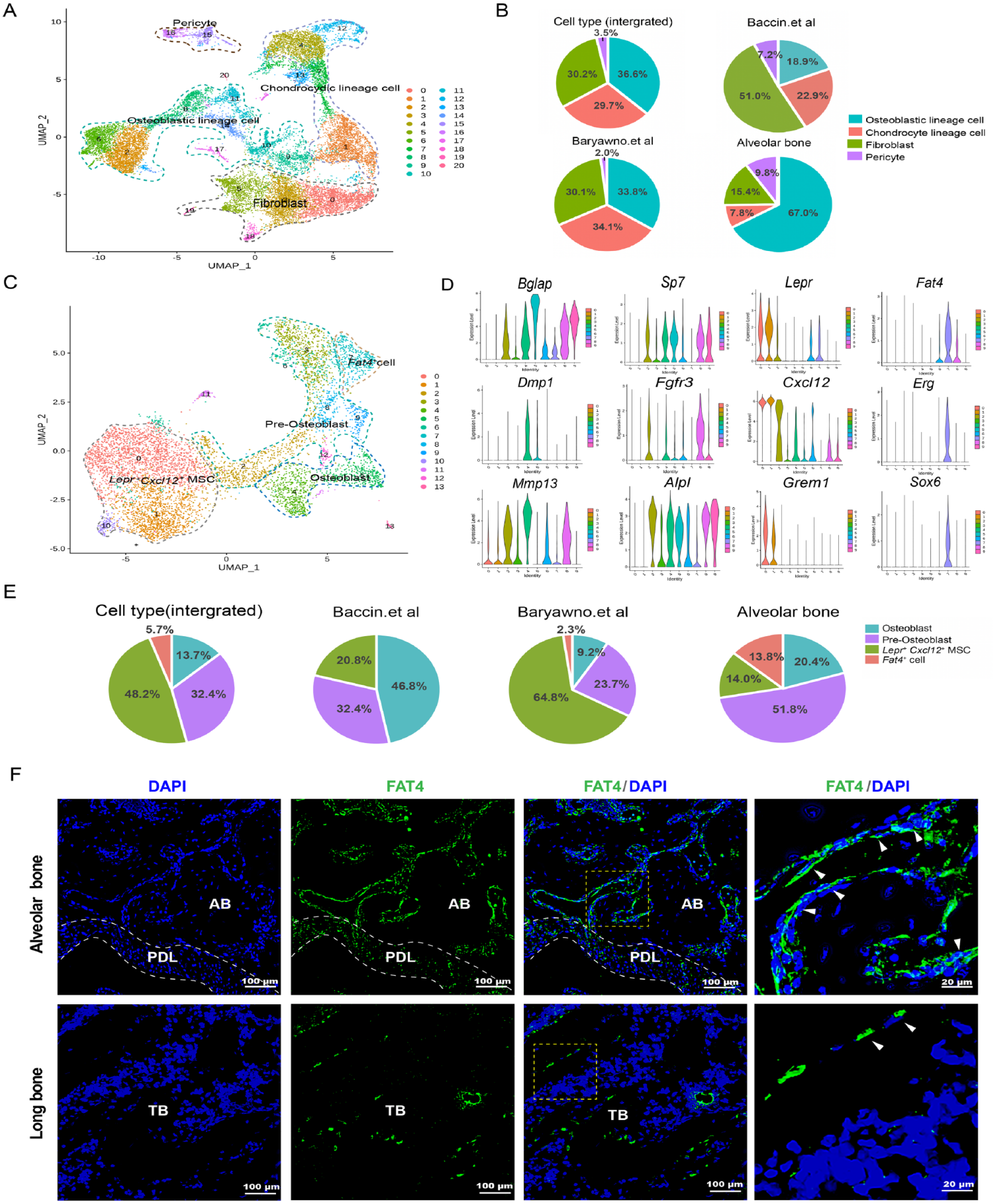
Differential composition of osteochondrogenic cell populations between alveolar bone and long bone reveals the enrichment of Fat4+ cells in alveolar bone. (
We further examined whether osteoblastic lineage cells in alveolar bone and long bone exhibit different cellular compositions, in particular, whether the alveolar bone contains unique osteoblastic subpopulations that may contribute to its distinct form of osteogenesis. Subclustering on the osteoblastic lineage cells from the integrated alveolar bone and long bone data sets resulted in 14 distinct subpopulations, among which 10 clusters contained at least 100 cells (clusters 0–9). These subpopulations could be classified into several major osteoblastic states based on the expression of typical markers of osteolineage differentiation along with the cell identity annotations from the previous long bone studies (Fig. 2D, Appendix Fig. 3A, B) (Baryawno et al. 2019; Baccin et al. 2020). First, Lepr+ mesenchymal stem cells (hereafter referred to as Lepr+ Cxcl12+ MSCs) were characterized by high expression of Lepr, Cxcl12, Kitl, Vcam1, and Grem1. Preosteoblasts were then annotated based on high expression of early osteoblast differentiation marker Runx2, Sp7, Alpl, and CD200, Fgfr3. Moreover, osteoblasts were identified by high expression of osteoblast maturation marker including Bglap, Dmp1, Ibsp, Col1a1, and Mmp13.
Notably, a distinct subpopulation of osteoblastic lineage cells characterized by exclusive expression of protocadherin Fat4, ETS transcription factor Erg, and sex-determining region Y-box Sox6 was identified (hereafter referred to as Fat4+ cells). Fat4+ cells exhibit relatively low expression of osteoblast maturation markers such as Bglap and Dmp1, osteoblast differentiation markers such as CD200 and Sp7 than preosteoblasts, and the markers for the former Lepr+ Cxcl12+ MSCs such as Lepr and Cxcl12 (Fig. 2D, Appendix Fig. 3A, B). Thus, Fat4+ cells may correspond to a mesenchymal osteoblastic lineage cell subpopulation that has not been described before. Importantly, we found that the Fat4+ cells are specifically enriched in the alveolar bone. Fat4+ cells accounted for nearly 15% of the osteoblastic lineage cells in the alveolar bone. In contrast, very few Fat4+ cells were present in long bone (2% and 0% in Baryawno et al. [2019] and Baccin et al. [2020], respectively) (Fig. 2E, Appendix Fig. 3C). Consistent with the scRNA-seq results, immunofluorescence staining showed that there were few Fat4+ cells in the long bone trabeculae. In contrast, considerably more Fat4+ cells were observed in mouse alveolar bone, where they were mainly located around the bone marrow cavities (Fig. 2F). The alveolar bone–specific enrichment of Fat4+ cells and their unique localization suggest these cells may be linked to the more active remodeling and greater osteogenic potential of the alveolar bone.
Fat4+ Cells Are Potential Mesenchymal Osteogenic Progenitors
We further investigated the role of Fat4+ cells during osteolineage differentiation by mapping them on the inferred differentiation trajectory of the integrated osteoblastic lineage cells. Lepr+Cxcl12+ MSCs were populated at the primary root of the pseudotime trajectory, with preosteoblasts and osteoblasts located at the ensuing segments along the trajectory, constituting the canonical osteolineage differentiation path. Intriguingly, Fat4+ cells diverged from the main differentiation path and formed a distinct branch. Branched expression analysis modeling (BEAM) analysis at the bifurcating point toward either the Fat4+ cells or osteoblasts (branch point 2 of Fig. 3A) revealed distinct gene expression profiles between the 2 branches (Fig. 3B).
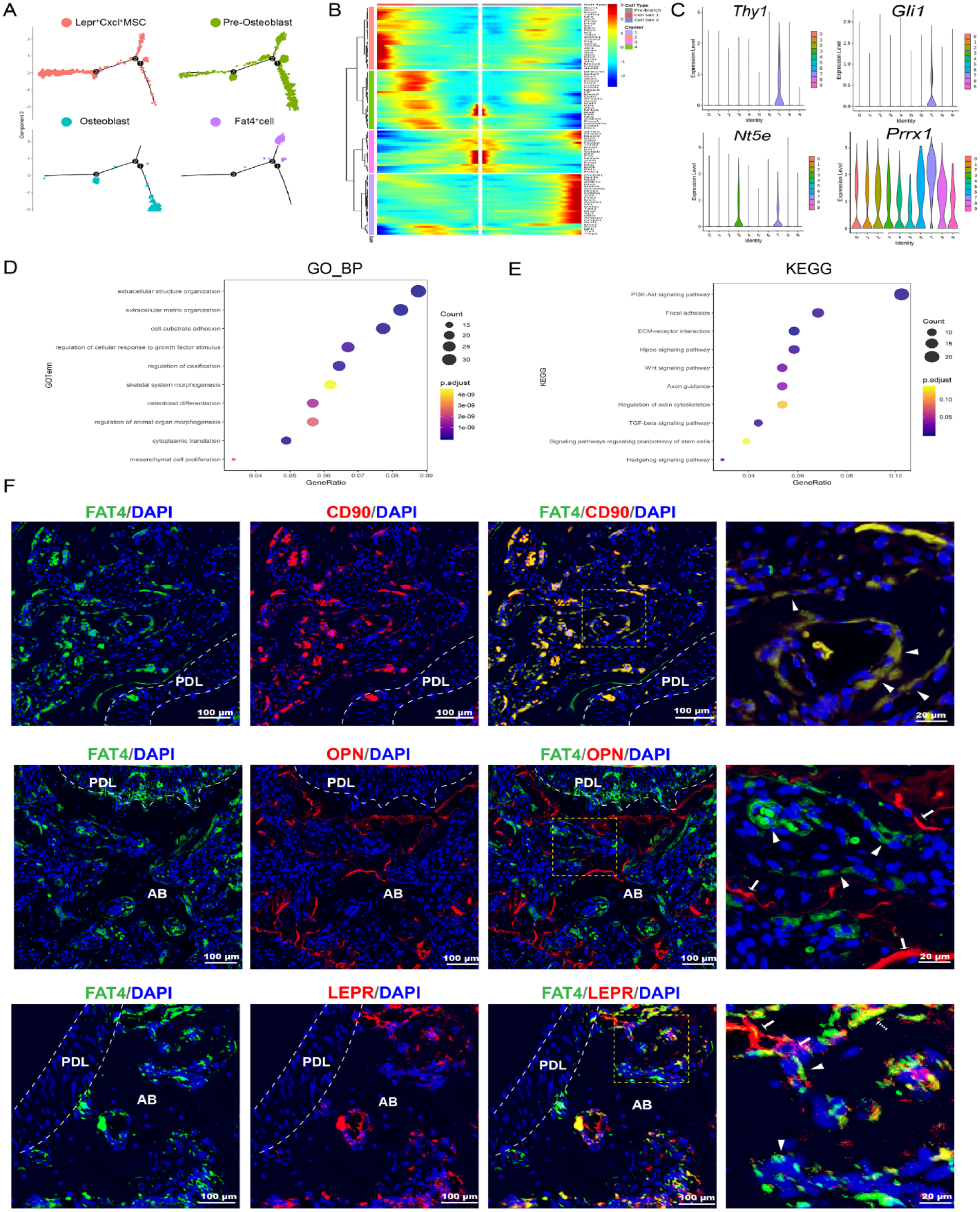
Fat4+ cells may initiate a distinct osteogenic differentiation trajectory in alveolar bone. (
While the Fat4+ cells might represent differentiated cells derived from the preosteoblasts, several lines of evidence suggested that these cells may represent a progenitor cell population that can initiate an alternative osteogenic differentiation trajectory in the alveolar bone. First, when analyzing the expression profile of MSC markers in the osteoblastic lineage cells, we found that several previously proposed stem cell markers such as Thy1 (CD90), Nt5e (CD105), Prrx1, and Gli1 were highly expressed in Fat4+ cells (Fig. 3C). Moreover, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses showed that Fat4+ cells were enriched for genes involved in extracellular structure organization, skeletal system morphogenesis, osteoblast differentiation, and PI3K-Akt, Hippo, and Wnt signaling pathway. These gene expression signatures suggested that the Fat4+ cells may be involved in alveolar bone development (Fig. 3D). Furthermore, immunofluorescent staining showed that Fat4+ cells were strongly coexpressed with typical MSC marker CD90, which was consistent with their expression profile in scRNA-seq analysis. In contrast, double staining of osteoblast marker OPN with FAT4 demonstrated that Fat4+ cells were located on the inner side of the bone marrow cavity relative to the osteoblasts. Moreover, whereas both Fat4+ cells and Lepr+ cells were located around the bone marrow, only a small fraction of cells exhibited coexpression of the 2 genes (Fig. 3E). Together, these results suggested that Fat4+ cells may represent a unique progenitor population of the alveolar bone.
Next, we directly demonstrated that the Fat4+ cells possess osteogenic differentiation capability. We isolated these cells from mouse primary alveolar bone cells or cultured mouse alveolar bone MSCs using cell surface FAT4 antibody (Fig. 4A–C). Concordant with the scRNA-seq results, flow cytometry analysis showed that ~3% of primary alveolar bone cells were Fat4+ cells, whereas only 0.01% of mouse long bone cells were Fat4+ (Fig. 4B). After verifying FAT4 expression of the sorting cells by immunochemical staining (Fig. 4D), these cells were cultivated in vitro to characterize their biological properties. Intriguingly, Fat4+ cells exhibited similar morphology to the canonical mouse MSCs, could form distinct colonies, and were capable of differentiating into osteoblasts and adipocytes (Fig. 4E–H). Moreover, to validate the osteogenic capacity of Fat4+ cells in vivo, ex vivo–expanded Fat4+ cells were transplanted subcutaneously into immunocompromised mice. Consistent with the in vitro differentiation assay, alizarin red staining demonstrated the formation of calcium nodules mediated by Fat4+ cells (Fig. 4I). Together, these results suggested the Fat4+ cells indeed exhibit characteristics of mesenchymal progenitors.
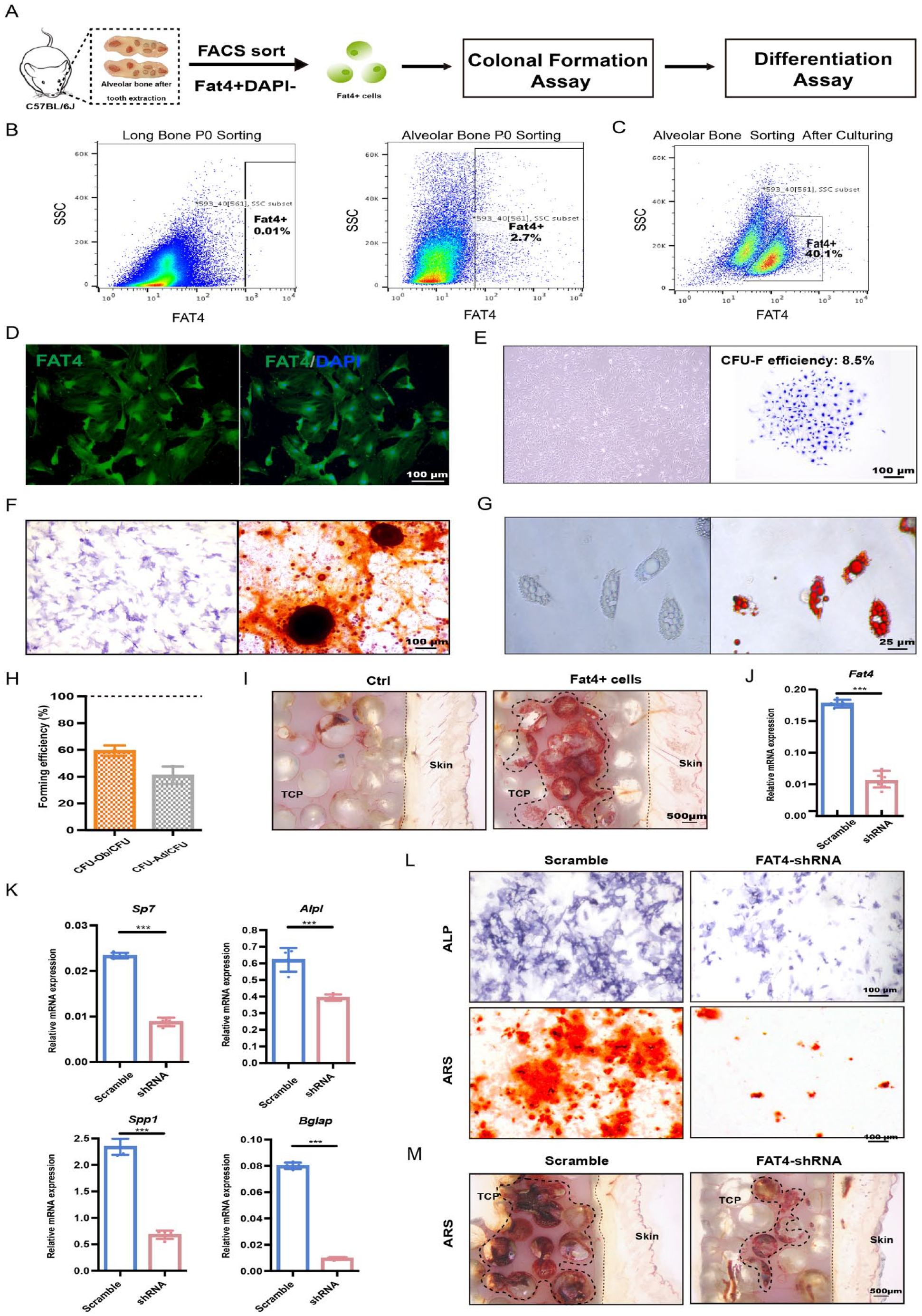
Biological performance of Fat4+ cells clarified it as an osteogenic progenitor. (
Furthermore, we knocked down the expression of FAT4 via lentivirus-expressed short hairpin RNA (shRNA) interference in alveolar bone MSCs. After confirming the inhibitory effect of shRNA (Fig. 4J), we analyzed the influence of FAT4 knockdown on the osteogenic differentiation of alveolar bone MSCs. Upon FAT4 knockdown, the mRNA levels of osteogenesis-related markers such as Sp7, Alpl, Spp1, and Bglap were notably reduced (Fig. 4K). Moreover, the ALP and alizarin red staining also showed decreased ALP activity as well as fewer bone-like nodules in the FAT4-shRNA group (Fig. 4L). In addition, fewer calcium nodules were detected in vivo when Fat4 expression was knocked down (Fig. 4M), suggesting the expression of FAT4 is required for the osteoblastic differentiation of alveolar bone–specific MSCs. Taken together, our results demonstrate that the Fat4+ cells are potential mesenchymal osteogenic progenitors that play a vital part in alveolar bone osteogenesis.
The Fat4+ Cells Exhibit Expression of Core Transcription Factors That Are Involved in Osteogenic Differentiation
We further explored the transcriptional regulatory networks underlying the progenitor properties of Fat4+ cells. By performing the SCENIC analysis, we assessed the activities of key transcription factors associated with the Fat4+ cells and inferred their transcriptional regulons, which consist of the transcription factors and their direct target genes. We found that the transcriptional activities of SOX6, ERG, IRX1, and PAX6 were particularly high in Fat4+ cells compared to the other osteoblastic subpopulations. These regulons of these transcription factors contain several key genes related to stem cell differentiation and osteogenesis regulation (Fig. 5A). Given that the SOX family transcription factors play critical roles in cartilage differentiation and skeletogenesis (Sarkar and Hochedlinger 2013), we wondered if SOX6 may contribute to the osteogenesis potential of alveolar bone MSCs. Immunochemical staining showed that SOX6 and FAT4 colocalized in alveolar bone cells, with the SOX6 signals enriched in the nucleus of Fat4+ cells (Fig. 5B). We next performed shRNA-mediated knockdown of SOX6 to investigate its influence on osteogenic differentiation. Indeed, the knockdown of SOX6 in alveolar bone MSCs (Fig. 5C) decreased the mRNA levels of downstream osteogenesis-related genes as well as the ALP activity and the formation of bone-like nodules (Fig. 5D–F). In summary, these data demonstrated that Fat4+ cells exhibit a core transcriptional signature consisting of several key transcription factors, such as SOX6, which may regulate the osteogenic differentiation potential of the Fat4+ cells, thereby contributing to alveolar bone osteogenesis.
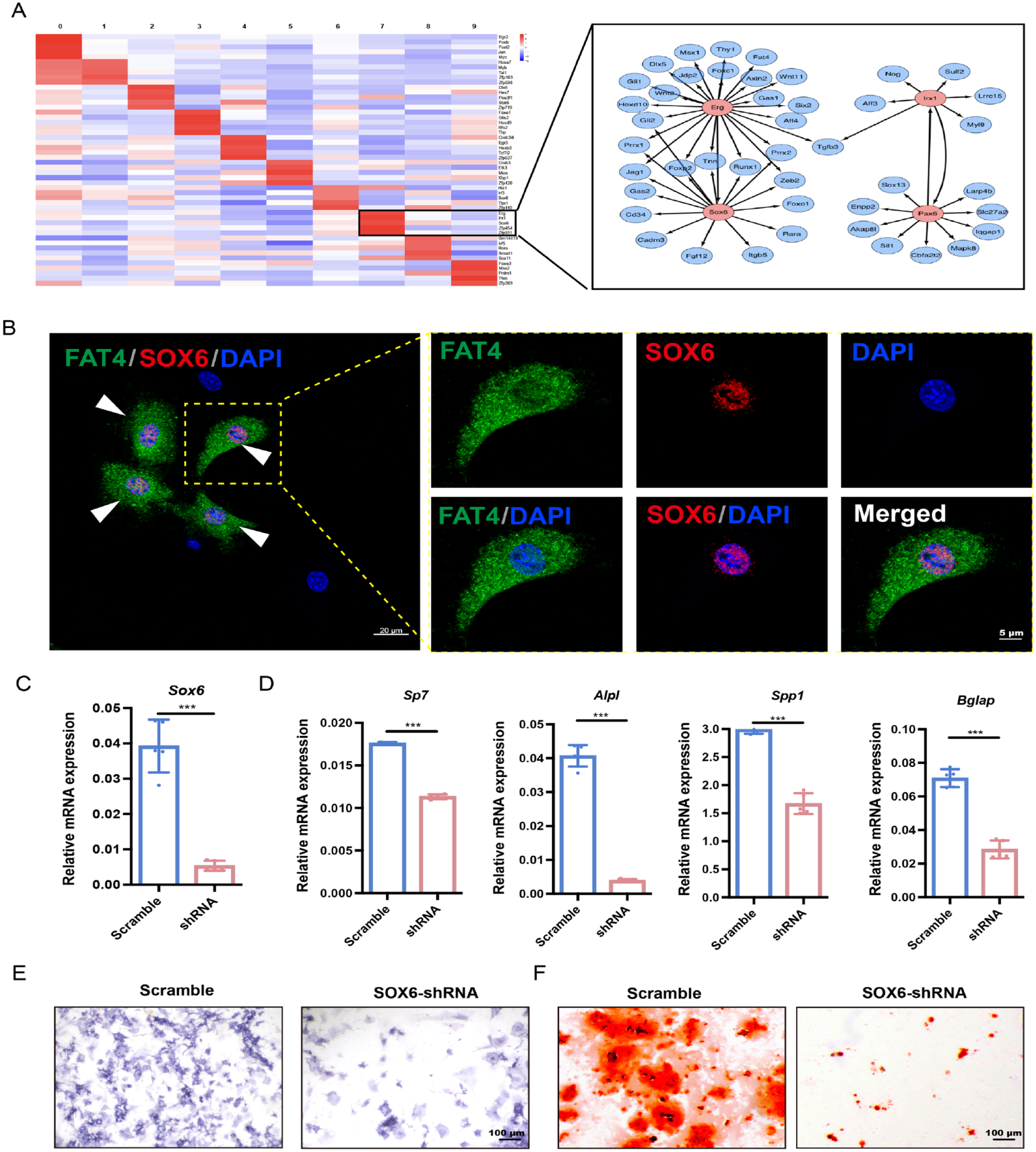
Fat4+ cells enriched for transcription factors regulating stem cell osteogenic differentiation. (
Discussion
MSCs from different developmental origins exhibit different physiological functions and significant heterogeneity. While unique physiological characteristics of alveolar bone have been well documented, there is a lack of systematic description of the heterogeneity of MSC-descendent osteoblastic lineage cells of alveolar bone. In this study, we constructed the single-cell atlas of the nonhematopoietic alveolar bone cells by scRNA-seq and revealed the differential composition of MSCs in alveolar bone and long bone. Combining scRNA-seq analysis, as well as in vivo and in vitro functional validation, we discovered a previously undescribed mesenchymal progenitor cell population, Fat4+ cells, which are particularly enriched in the alveolar bone, exhibit a core transcriptional signature linked to osteogenesis, and may initiate a distinct osteogenic differentiation trajectory of alveolar bone.
Advances in scRNA-seq have provided unprecedented opportunities for characterizing the bone marrow microenvironment. To date, there are 3 main strategies for obtaining MSCs and MSC-derived osteolineage cells in long bones for scRNA-seq: 1) the whole bone marrow single-cell suspension was used for sequencing (Lin et al. 2021), which was often used to capture the hematopoietic cells atlas; 2) using conditional fluorescence reporter-expressing mice, for example, Lepr-Cre (Tikhonova et al. 2019) and Col2-Cre (Zhong et al. 2020), followed by FACS, which was used for selectively purifying certain subsets based on known markers; and 3) single-cell suspension of bones, followed by sorting CD45– Ter119– nonhematopoietic and CD31– nonendothelial cells (Wolock et al. 2019) for sequencing. When it comes to alveolar bone, the very first attempt to construct the single-cell atlas of mouse mandibula has been performed using the first strategy (Lin et al. 2021). However, the nonhematopoietic cells accounted for only 1.74% of all the identified cells in this scRNA-seq data set, hampering the identification of different mesenchymal cell subpopulations. To construct more information-rich single-cell atlas of nonhematopoietic alveolar bone marrow cells, we adopted a different strategy to purify them by extracting the teeth, dissecting the ridge of jawbone, and crushing the alveolar bone, followed by enzymatic digestion to obtain single-cell suspension, and further enriching MSCs by efficiently sorting CD45–Ter119– nonhematopoietic cells. As a result, the nonhematopoietic cells now accounted for 93% of all cells in our scRNA-seq data set. The improved yield of mesenchymal cells led to the identification of 6 subsets of osteolineage cells, 4 subsets of endothelial cells, and 3 subsets of neurological cells and pericytes, greatly improving the resolution of the single-cell atlas of the alveolar bone MSCs.
Compared with long bone and other bones, alveolar bone exhibits exclusive biological characteristics. However, the distinct cell composition of alveolar bone osteolineage cells compared to long bone remains unclear. Of particular note, our scRNA-seq data revealed a newly identified osteolineage cell population that is characterized by the prominent expression of protocadherin Fat4 and transcription factor Sox6. The Fat4+ cells are particularly enriched in alveolar bone compared to long bone, are located on the inner side of the bone marrow cavity relative to osteoblasts, and express canonical MSC markers, suggesting they may represent a unique type of mesenchymal osteogenic progenitors enriched in alveolar bone. FAT4 has been demonstrated to control planar cell polarity, neuronal progenitor differentiation, and mesenchymal cell clustering, thus playing a role in the development of neural tube, bone, and gut (Mao et al. 2016; Betterman et al. 2020; Rao-Bhatia et al. 2020). In human, mutations of the protocadherins FAT4 result in Van Maldergem syndrome, which is characterized by craniofacial defects due to impaired osteogenesis of the maxilla (Cappello et al. 2013). Moreover, the SOX family transcription factors are well-established regulators of cell fate decisions during development. Among them, SOX6 cooperates with SOX9 and plays a key role in skeletogenesis (Ikeda et al. 2004; Guth and Wegner 2008). Indeed, by performing in vitro colony formation assay, as well as osteogenic and adipogenic induction assay, we have validated the mesenchymal progenitor properties of Fat4+ cells. Furthermore, our data demonstrated that knockdown of SOX6 or FAT4 could significantly inhibit the osteogenic differentiation of alveolar bone MSCs. In conclusion, the scRNA-seq analysis, combined with functional evidence, strongly suggested Fat4+ cells as distinct progenitor cells with osteogenic potential of alveolar bone. These data were in accordance with Crespo-Enriquez et al. (2019), who found that Fat4–/– mice showed severe craniofacial skeletal defects.
In summary, our high-resolution, high-information content single-cell atlas of the nonhematopoietic alveolar bone cells enabled the identification of an alveolar bone–enriched osteogenic progenitor cell population, Fat4+ cells. Future lineage-tracing studies will further elucidate the exact in vivo function and developmental trajectory of Fat4+ cells as a osteogenic progenitor. Our study revealed the unique cellular composition of alveolar bone MSCs and osteolineage cells, which may pave an avenue for developing therapeutic treatment of craniofacial deformities and periodontal diseases in the future.
Author Contributions
A. Jin, contributed to conception and design, data acquisition, analysis and interpretation, drafted and critically revised the manuscript; X. Gao, contributed to conception and data design, drafted and critically revised the manuscript; H. Xu, S. Sun, Y. Yang, X. Huang, X. Wang, Y. Liu, Y. Zhu, contributed to analysis and data interpretation, critically revised the manuscript; Q. Dai, Q. Bian, L. Jiang, contributed to conception and data design, critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345231159821 – Supplemental material for ScRNA-Seq Reveals a Distinct Osteogenic Progenitor of Alveolar Bone
Supplemental material, sj-docx-1-jdr-10.1177_00220345231159821 for ScRNA-Seq Reveals a Distinct Osteogenic Progenitor of Alveolar Bone by A. Jin, H. Xu, X. Gao, S. Sun, Y. Yang, X. Huang, X. Wang, Y. Liu, Y. Zhu, Q. Dai, Q. Bian and L. Jiang in Journal of Dental Research
Footnotes
Acknowledgements
The authors thank the bioimaging facility and flow cytometry lab of Shanghai Institute of Precision Medicine.
A supplemental appendix to this article is available online.
Declaration of Conflicting Interests
The authors declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by grants from the National Natural Science Foundation of China (82071083, 82271006, 82101048, 81870740, 81570950, 31970585, 32170544, 31801056); the Program of Shanghai Academic/Technology Research Leader (20XD1422300); the Natural Science Foundation of Shanghai (21ZR1436900, 22ZR1436700); Clinical Research Plan of SHDC (SHDC2020CR4084); Biomaterials and Regenerative Medicine Institute Cooperative Research Project, Shanghai Jiaotong University School of Medicine (2022LHB02); the Cross-Disciplinary Research Fund of Shanghai Ninth People’s Hospital, Shanghai Jiao Tong University School of Medicine (JYJC201902, JYJC202116); the Innovation Research Team of High-Level Local Universities in Shanghai (SSMU-ZLCX20180501, SHSMU-ZLCX20211700); the Research Discipline Fund no. KQYJXK2020 from Ninth People’s Hospital, Shanghai Jiao Tong University School of Medicine, and College of Stomatology, Shanghai Jiao Tong University; Original Exploration Project of Shanghai Ninth People’s Hospital, Shanghai Jiao Tong University School of Medicine (JYYC003); the Project of Biobank of Shanghai Ninth People’s Hospital, Shanghai Jiao Tong University School of Medicine (YBKB202216); and Two-Hundred Talent Project of Shanghai Jiao Tong University School.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.