
Abstract
In humans, teeth are replaced only once, and the successional dental lamina (SDL) of the permanent tooth is maintained in a quiescent state until adolescence. Recently, we showed that biomechanical stress generated by the rapid growth of the deciduous tooth inhibits SDL development via integrin β1–RUNX2 signaling at embryonic day 60 (E60) in miniature pigs. However, the mechanism by which RUNX2 regulates SDL initiation within the SDL stem cell niche remains unclear. In the current study, we transcriptionally profiled single cells from SDL and surrounding mesenchyme at E60 and identified the landscape of cellular heterogeneity. We then identified a specific fibroblast subtype in the dental follicle mesenchyme between the deciduous tooth and the SDL of the permanent tooth (DFDP), which constitutes the inner part of the niche (deciduous tooth side). Compared with traditional dental follicle cells, the specific expression profile of DFDP was identified and found to be related to biomechanical stress. Subsequently, we found that RUNX2 could bind to the enhancer regions of Fbln1 (gene of fibulin-1), one of the marker genes for DFDP. Through gain- and loss-of-function experiments, we proved that the biomechanical stress–mediated RUNX2–fibulin-1 axis inhibits the initiation of SDL by maintaining SDL niche homeostasis.
Keywords
Introduction
Most vertebrates renew their teeth multiple times throughout life, whereas humans renew their teeth only once. Replacement teeth develop from the successional dental lamina (SDL), an epithelial band embedded in the mandible (Wu et al. 2020). Epithelial stem cells in the SDL differentiate into all cell lineages in the enamel organ of mice (Juuri et al. 2013). However, how SDL epithelial stem cells maintain quiescence or are activated in the mandibular microenvironment remains unclear. The stem cell niche is the cellular microenvironment in which stem cells reside. It governs their stemness and adaptability (Ruiz et al. 2014; Chacon-Martinez et al. 2017). The stem cell niche of the SDL, which we discovered previously upon examining alligator tooth renewal (Wu et al. 2013), refers to the mesenchymal microenvironment surrounding the SDL in the mandible. Although RNA sequencing of the SDL and surrounding mesenchyme has been conducted in bearded dragons (Salomies et al. 2019), the cell–cell or cell–extracellular matrix (ECM) interactions within the niche regulating the SDL stem cells in large mammals are largely unknown.
Miniature pigs, a large mammal model, have diphyodont dentition, similar to humans. Our previous work used the replacement of the deciduous canine (DC) with the permanent canine (PC) in miniature pigs as a model, demonstrating that the SDL of the PC separated from the DC at embryonic day (E) 60 and then remained in a quiescent state until DC eruption at E90 (Wu et al. 2020). We also found that the biomechanical stress generated by the differential growth rates of the DC and alveolar socket inhibits SDL initiation via the integrin β1–RUNX2 pathway, which is expressed in the dental follicle mesenchyme between the DC and SDL of the PC (Wu et al. 2020).
In the current study, we aimed to investigate how biomechanical stress–mediated RUNX2 regulates SDL initiation within the SDL stem cell niche. We transcriptionally profiled single cells from SDL and surrounding mesenchyme at E60 and identified the landscape of cellular heterogeneity. We then identified a specific fibroblast subtype in the dental follicle mesenchyme between the deciduous tooth and the SDL of the permanent tooth (DFDP) that constitutes the inner part (deciduous tooth side) of the niche. We then demonstrated that DFDP secretes the ECM protein fibulin-1, which acts downstream of RUNX2. In this manner, the biomechanical stress–mediated RUNX2–fibulin-1 axis maintains the resting stage of the SDL and may inhibit PC initiation in the niche.
Materials and Methods
Detailed materials and methods are included in the Appendix.
Pregnant Wuzhishan pigs from the Institute of Animal Science of the Chinese Agriculture University were used in this study. All procedures were reviewed and approved by the Animal Care and Use Committee of the Capital Medical University in Beijing, China (permit number: AEEI-2018-186). This study complied with the ARRIVE (Animal Research: Reporting of In Vivo Experiments) 2.0 guidelines.
Single-cell RNA sequencing (scRNA-seq) data (10x Genomics Chromium) were profiled and quantified using the Cellranger V4.0.0 (10x Genomics) pipeline with the Sscrofa11.1 genome as reference. Quality control process, normalization, clustering, differential gene expression, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis, and visualization were performed using Seurat (V4.0.6), clusterProfiler, and ggplot2. Spatial transcriptomics was performed using the Visium Spatial Gene Expression assay (10x Genomics), and the accession number for the section is “PN: 2000233; SN: V10L23-019-A1: 210923B_E60DC_C.” The analysis scripts have been made available on the GitHub repository (https://github.com/QiongLi1996/E60-DC_C), and technical details are listed in the Appendix.
The mechanical force (3 kPa) was applied on the mandible slice using the Flexcell FX-5000 Compression System (Flexcell International). The sources of all reagents are listed in Appendix Table 1. The fibulin-1 recombinant protein (R&D Systems, cat. 9007-FB-050) and the anti–fibulin-1 antibody (Abcam, cat. ab175204) were used in this research.
Results
Identification of Specific Fibroblast Subtype in the DFDP
Our previous study showed that SDL of the PC separated from the DC at E60 and then remained in a quiescent state until DC eruption at E90 (Wu et al. 2020). To investigate which cell types regulate the SDL and how the SDL changes its expression profile, we performed spatial transcriptomics (ST) and scRNA-seq on the mandibular bone tissue of miniature pig embryos at E60. The SDL and the surrounding mesenchyme were micro-dissected into 1 sample (Fig. 1A). As the width and length of the SDL were approximately 60 µm and 700 µm, respectively, it was not possible to harvest enough cells from 1 sample. Therefore, in this study, we mixed 12 samples dissected from 12 embryos at E60 into 1 mixed sample and digested them into a single-cell suspension (Fig. 1A). Clustering was performed using Uniform Manifold Approximation and Projection (UMAP), and 5 clusters were identified: oral epithelial, SDL, fibroblast, endothelial, and myeloid (Fig. 1B; Appendix Fig. 1A). We found that KRT8, KRT14, PITX2, SOSTDC1, and PI15 were the marker genes of the SDL (Appendix Fig. 1A). SOSTDC1 has been proved to be expressed on the SDL in our previous work (Wu et al. 2020). The identified marker genes, KRT8 and PI15, were confirmed by immunofluorescence (Appendix Fig. 2E, F′).
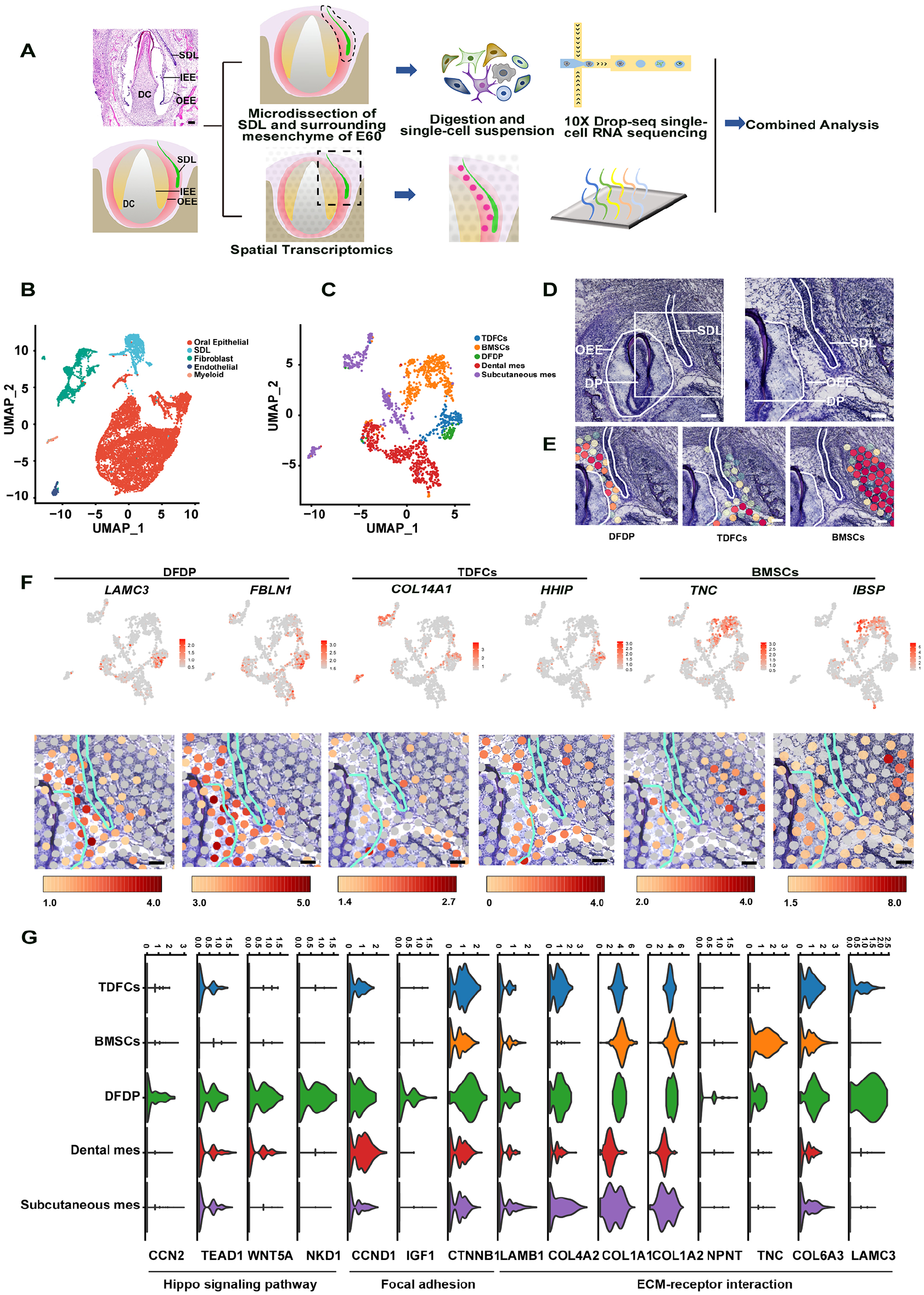
Identification of specific fibroblast subtype in the dental follicle mesenchyme between the deciduous tooth and the successional dental lamina (SDL) of the permanent tooth (DFDP). (
To elucidate the heterogeneity of the microenvironment of the SDL, we then subclustered the fibroblast population in Figure 1B. Approximately 1,673 fibroblast cells were clustered into 5 fibroblast subpopulations (Fig. 1C; Appendix Fig. 1B). To confirm the clustering results, we mapped the scRNA-seq data to the mandible section using ST (Fig. 1D, E; Appendix Fig. 1C–G), and the results showed a distinct distribution of different subpopulations. There were 2 major fibroblast subpopulations close to the SDL of PC: the DFDP and traditional dental follicle cells (TDFCs). The DFDP was located at the dental follicle mesenchyme between the DC and SDL of the PC, while TDFCs were located at the other parts of the dental follicle of the DC (Fig. 1E). The other 3 fibroblast subpopulations, BMSCs, dental mes, and subcutaneous mes, were mapped to the alveolar bone, dental mesenchyme area, and subcutaneous mesenchyme area, respectively (Fig. 1E; Appendix Fig. 1F, G). The marker genes of these subtypes are shown in Appendix Fig. 1B. The representative marker genes LAMC3 and FBLN1 for the DFDP, COL14A1 and HHIP for TDFCs, and TNC and IBSP for BMSCs were shown with feature plot analysis of scRNA-seq and ST spot (Fig. 1F; Appendix Fig. 1B). Furthermore, the immunostaining result of marker genes of COL26A1, RUNX2, COL14A1, AKAP12, and IBSP confirmed the mesenchymal cell subtypes (Appendix Fig. 2A–D, G).
As the SDL was surrounded by the DFDP and TDFCs, we then set out to determine which cell type senses the mechanical stress and regulates the behavior of the SDL. We further analyzed the differences between the DFDP and TDFCs. The upregulated and downregulated genes in the DFDP compared to TDFCs were visualized using a volcano map (Appendix Fig. 3A). We found that ECM genes (COL26A1, LAMC3, and FBLN1) were among the top 10 genes of the DFDP (Appendix Fig. 3A), confirming our previous finding that ECM molecules are the main components of the SDL stem cell niche (Wu et al. 2013). KEGG pathway enrichment analysis revealed 17 pathways enriched in the DFDP compared to the TDFCs. Among them, ECM–receptor interaction (ranking 1st), focal adhesion (ranking 2nd), and Hippo signaling pathway (ranking 12th) were related to biomechanical stress (Appendix Fig. 3B, Appendix Table 2). All genes participating in the 3 pathways are listed in Fig. 1G, including focal adhesion molecules, ECM–receptor interaction molecules, and Hippo signaling pathway molecules. The results showed the molecules of these biomechanical stress–related pathways upregulated in the DFDP. Thus, we identified a specific fibroblast subtype, DFDP, in the dental follicle mesenchyme between the DC and SDL of the PC that was regulated by biomechanical stress at E60.
Fibulin-1 Acts as a Downstream Molecule of RUNX2 in the DFDP
Previously, we found that the biomechanical stress generated by the differential growth rates of the DC and alveolar socket inhibits SDL initiation via the integrin β1–RUNX2 pathway, which is expressed in the dental follicle mesenchyme between the DC and SDL of the PC (Wu et al. 2020), where the DFDP is located (Appendix Fig. 2B; Appendix Fig. 3C, D). As ECM molecules COL26A1, LAMC3, and FBLN1 are highly expressed in the DFDP (Appendix Fig. 3A), we then investigated whether RUNX2 regulates the quiescence or activation state of SDL stem cells via one of them. By analyzing the chromatin immunoprecipitation with high-throughput sequencing (ChIP-seq) data of mouse mesenchyme stem cells (GSE79813) (Meyer et al. 2016), we found that Runx2 binds to the intronic regions of Fbln1 by identifying ChIP-seq peaks (Fig. 2A). Previous studies suggested that enhancers exhibit a characteristic chromatin “signature” of monomethylation of histone H3 lysine 4 (H3K4me1) in the absence of significant trimethylation (H3K4me3) (Bulger and Groudine 2011). Moreover, one of the cis-regulatory motifs recognized by the CCAAT/enhancer binding protein (C/EBP) family is the enhancer core homology (Landschulz et al. 1988). We thus reanalyzed the ChIP-seq data sets targeting the DNA binding sites of C/EBPβ and C/EBPα, as well as histone modification sites with H3K4me1 and H3K4me3, to determine whether the intronic regions were enhancers (Meyer et al. 2016). We found that the binding sites of RUNX2 coincide with the binding sites of C/EBPα, C/EBPβ, and H3K4me1, but not H3K4me3 (Fig. 2A). It suggested the intronic RUNX2 binding sites in the Fbln1 gene were probably enhancers. Through conservation analysis, we found that these enhancers of Fbln1 are conserved between mouse and pig (Appendix Fig. 4). To examine whether fibulin-1 acts downstream of RUNX2, we transduced dental follicle cells with a RUNX2 overexpression or control lentivirus vector and assessed the expression level of fibulin-1 in the supernatant via proteomic analysis. After culturing for 48 h, secreted proteins in the supernatant were screened. The analysis revealed 32 upregulated proteins in the RUNX2 overexpression group, and fibulin-1 was at the top of the list (Fig. 2B; Appendix Table 3). Thus, we believe that fibulin-1 is a downstream molecule of RUNX2 in the DFDP. Although RUNX2 could also bind to the potential enhancer regions of Col26a1 and Lamc3, the ChIP-seq signals of these 2 genes were lower than that of Fbln1 (Fig. 2A; Appendix Fig. 5). Thus, these results indicated that RUNX2 could upregulate the Fbln1 expression through binding to the enhancers of Fbln1, and thereby we selected Fbln1 for the further study.
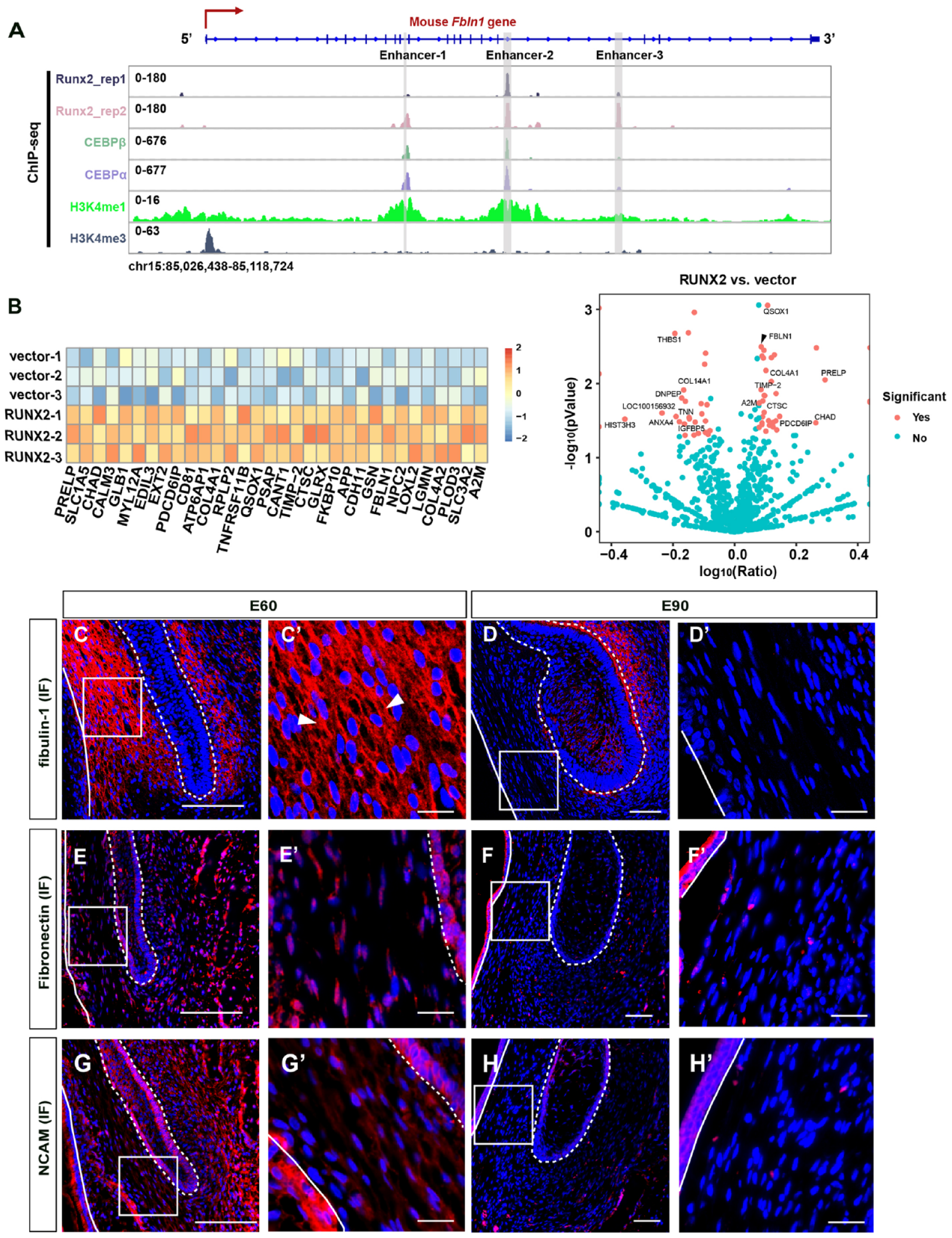
Fibulin-1 acts as a downstream molecule of RUNX2 in the dental follicle mesenchyme between the deciduous tooth and the successional dental lamina (SDL) of the permanent tooth (DFDP). (
The scRNA-seq and ST results showed that the expression level of FBLN1 in DFDP was substantially higher than that in TDFCs at E60 (Fig. 1F). Furthermore, immunofluorescence (IF) staining confirmed this expression pattern at E60 (Fig. 2C). The expression level of fibulin-1 in the DFDP decreased significantly from E60 to E90, whereas no significant difference in the TDFCs was observed from E60 to E90 (Fig. 2C–D′). The dramatic decrease in DFDP suggests a fundamental role of fibulin-1 in maintaining the homeostasis of the SDL niche at E60.
To determine whether other ECM molecules exhibited similar expression dynamics from E60 to E90, we assessed the expression levels of fibronectin (Lukjanenko et al. 2016; Dalton and Lemmon 2021) and neural cell adhesion molecule (NCAM) (Wu et al. 2013), which have been demonstrated to function in the stem cell niche. We found that the expression levels of these 2 molecules were high in both the DFDP and TDFCs at E60 but decreased at E90 (Fig. 2E–H′).
Fibulin-1 Is Regulated by Biomechanical Stress–Mediated RUNX2 Axis in the DFDP
Previously, we micro-dissected the mandible slice containing the DC and SDL of the PC and cultured it in vitro for 2 d. We found that the SDL was enlarged and started developing in only 2 d in vitro (Wu et al. 2020). Subsequently, we investigated whether fibulin-1 was also regulated by the biomechanical stress–mediated RUNX2 axis. E60 mandibular slices were cultured with static compression of 3 kPa using the FX-5000 Compression System, and then we examined fibulin-1 expression levels. RUNX2 expression was significantly upregulated in the 3-kPa group and downregulated in the 0-kPa group (Fig. 3A–C). In the 3-kPa group, fibulin-1 was highly expressed in both the DFDP and TDFCs as assessed by IF staining, which resembled its expression pattern at E60. However, almost no IF signal was detected in the DFDP of the 0-kPa group, which resembled the expression pattern at E90 (Fig. 3D–E′).
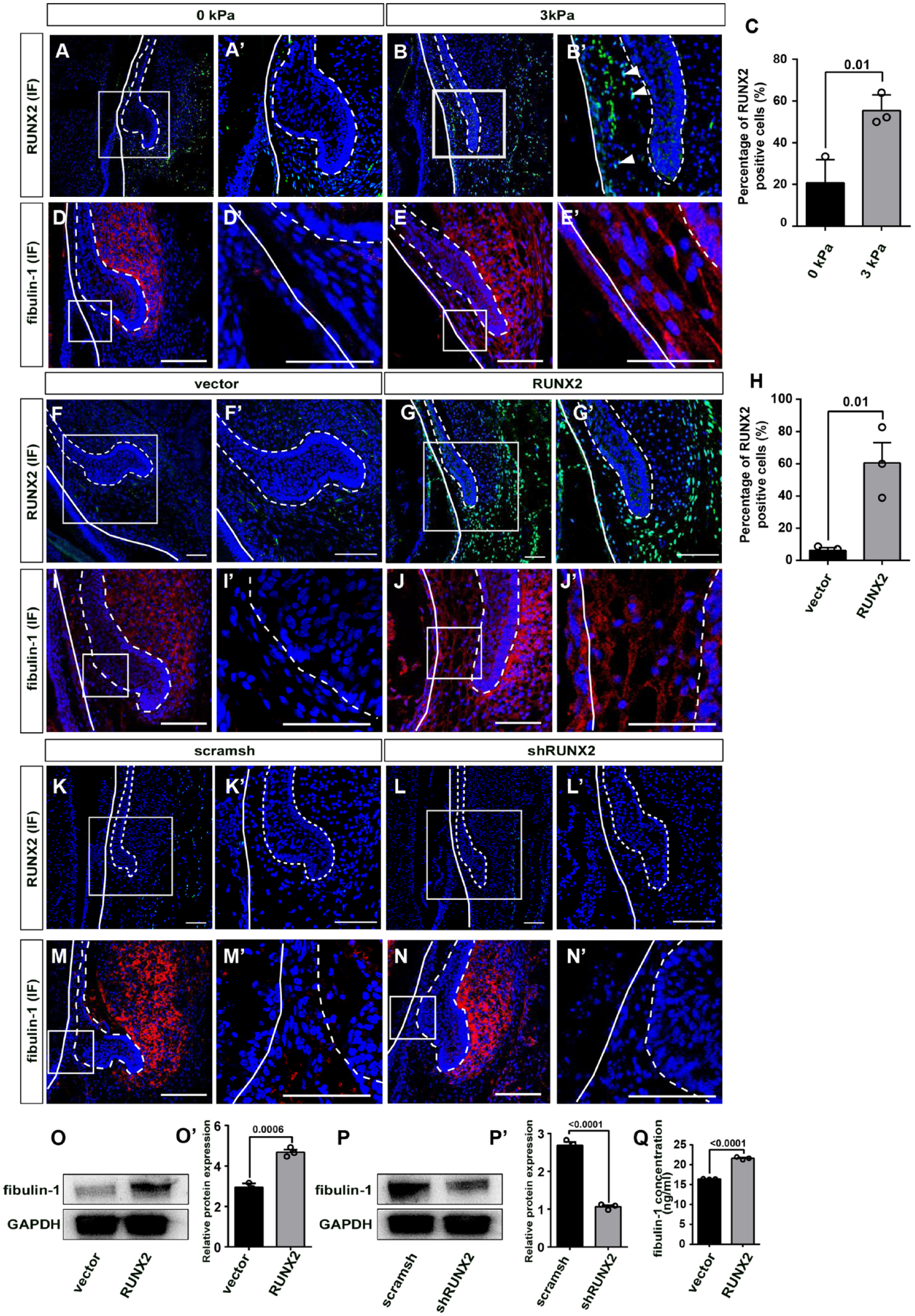
Biomechanical stress–mediated RUNX2 regulates fibulin-1 expression during permanent canine (PC) initiation stage. (
We then examined fibulin-1 levels in cultured mandibular slices transduced with RUNX2 overexpression (RUNX2 group) or control lentivirus vector (vector group) and RUNX2 knockdown (shRUNX2 group) or knockdown control lentivirus vectors (scramsh group). RUNX2 expression was significantly upregulated in the RUNX2 group (Fig. 3F–H) and was downregulated in the shRUNX2 group (Fig. 3K–L′). The results showed that fibulin-1 was upregulated in the RUNX2 group, resembling the expression pattern at E60 (Fig. 3J). However, almost no IF signal was detected in the vector, shRUNX2, or scramsh groups, which resembled the fibulin-1 expression pattern at E90 (Fig. 3I, M–N′).
To confirm these findings at the cell level, dental follicle cells were transduced with RUNX2 overexpression or knockdown lentivirus vectors and their respective controls. The results showed that fibulin-1 expression was significantly upregulated in the RUNX2 overexpression group and downregulated in the RUNX2 knockdown group (Fig. 3O–P′; complete Western blot gels are shown in Appendix Fig. 6). The RUNX2 overexpression group had higher levels of fibulin-1 in their supernatant compared to the control group, as assessed by enzyme-linked immunosorbent assay (Fig. 3Q).
These results indicate that fibulin-1 is regulated by the biomechanical stress–mediated RUNX2 axis in the DFDP.
Fibulin-1 Inhibits PC Initiation, Probably by Maintaining ECM Niche Homeostasis
We investigated whether fibulin-1 influenced PC initiation. Gain- and loss-of-function studies were performed by incubating cultured E60 mandibular slices with fibulin-1 recombinant protein or fibulin-1 neutralizing antibody. After in vitro culturing for 48 h, the SDL of the PC entered the early cap stage in the control and fibulin-1 neutralizing antibody groups (Fig. 4A, B). However, cells remained in a quiescent state in the fibulin-1 recombinant protein group (Fig. 4C). This demonstrated that fibulin-1 stabilizes the niche and is required to maintain the quiescent state of the SDL.
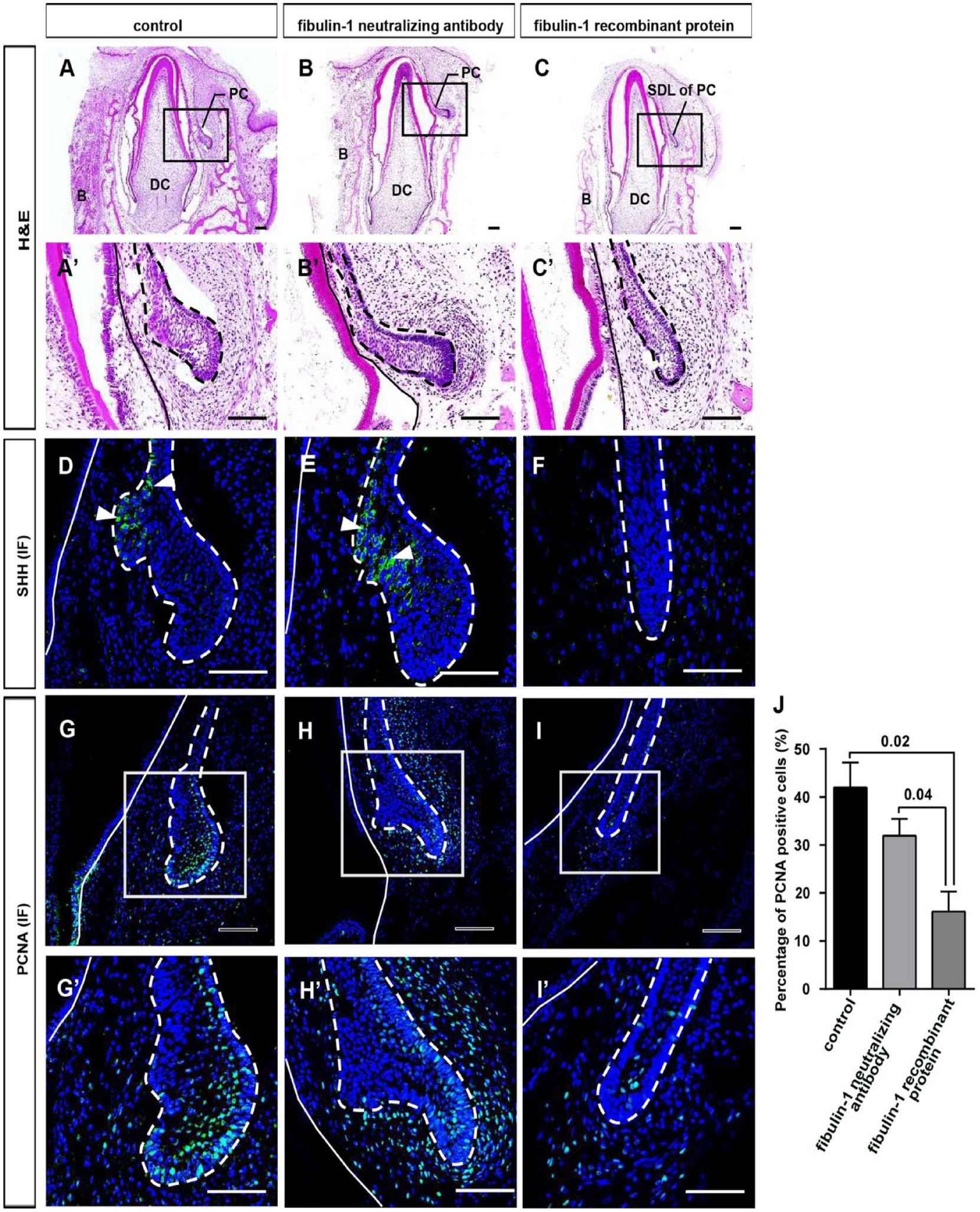
Fibulin-1 inhibits permanent canine (PC) initiation. (
To examine whether the enlarged SDL had differentiated into the enamel organ, we assessed the expression of SHH, a marker of the primary enamel knot. In cultured mandibular tissue, SHH was expressed on the inner side of the SDL in the control and fibulin-1 neutralizing antibody groups. However, SHH was not expressed in the fibulin-1 recombinant protein group (Fig. 4D–F). To investigate the proliferation status of the cultured E60 mandible slices, IF staining of proliferating cell nuclear antigen (PCNA) was performed. This result showed that PCNA-positive cells were significantly higher in control and fibulin-1 neutralizing antibody groups compared to the fibulin-1 recombinant protein group (Fig. 4G–J).
Activation of the canonical Wnt pathway in the dental epithelium indicates the initiation of tooth development. We examined the expression of 2 key molecules, CTNNB1 and lymphoid enhancing factor 1 (LEF1), showing that CTNNB1 RNA and LEF1 protein levels significantly increased in the SDL of the control and fibulin-1 neutralizing antibody groups compared to the fibulin-1 recombinant protein group (Fig. 5A–G). The results indicated that the SDL had differentiated into the enamel organ in both the control and fibulin-1 neutralizing antibody groups.
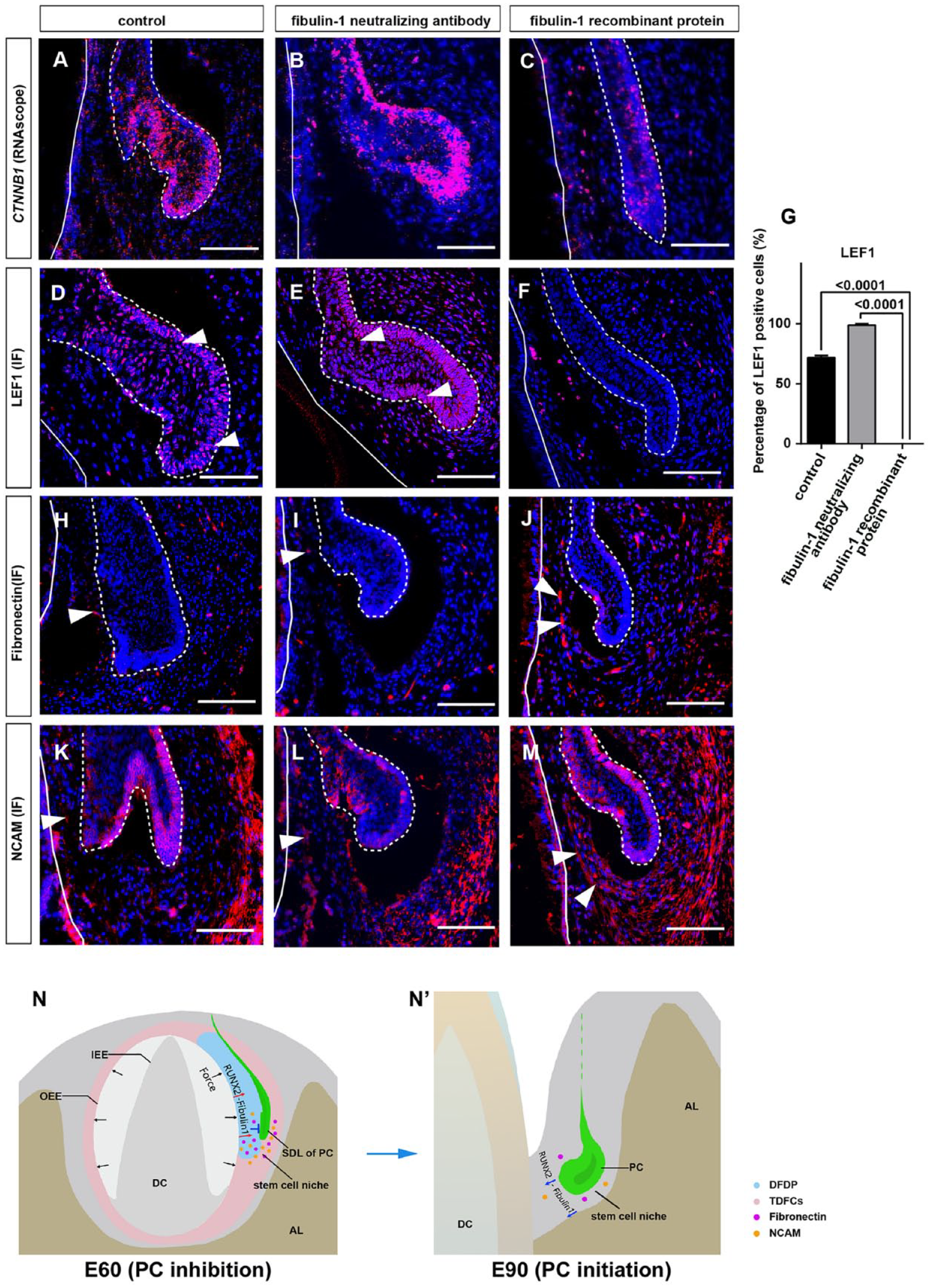
Fibulin-1 regulates SDL initiation via maintaining extracellular matrix (ECM) homeostasis. (
We also studied the expression patterns of ECM molecules fibronectin and NCAM in the control, fibulin-1 neutralizing antibody, and fibulin-1 recombinant protein groups. The results showed that the expression levels of both molecules were significantly decreased in the DFDP in the control and fibulin-1 neutralizing antibody groups, resembling that of E90 (Fig. 5H, I, K, L). However, the expression levels of both molecules were partially maintained in both the DFDP and TDFCs in the fibulin-1 recombinant protein group, which resembled that of E60 (Fig. 5J, M).
These findings suggested that fibulin-1 maintains niche homeostasis in the DFDP.
Discussion
In the current study, we further investigated the regulatory mechanisms underlying mesenchymal RUNX2 expression within the niche. First, the landscape of cellular heterogeneity of the SDL and surrounding mesenchyme at E60 was identified using a combination of scRNA-seq and ST. A specific fibroblast subtype named the DFDP was found in the mesenchyme between the DC and SDL of the PC. We then found that RUNX2 bound to the enhancer regions of Fbln1, a marker gene for the DFDP. Through gain- and loss-of-function experiments, we proved that the biomechanical stress–mediated RUNX2–fibulin-1 axis regulates the initiation of SDL by maintaining the SDL niche homeostasis (Fig. 5N, N′).
Specific Niche Mesenchymal Cells Exert Critical Effects on the Quiescent State of the SDL
The stem cell niche consists of stem cell progeny, mesenchymal/stromal and immune cells, and niche-specific ECM (Takeda et al. 2011; Morgner et al. 2015; Ali et al. 2017; Roberts et al. 2017). In the niche, progenies provide essential niche signals to stem cells (Sato et al. 2011), and mesenchymal and immune cells produce the necessary signals. For example, mesenchymal cells in the dermal papilla secrete bone morphogenetic proteins and WNTs, which regulate the activated or quiescent state of hair follicle stem cells in mice (Plikus et al. 2008). The stem cell niche of the SDL refers to the mesenchymal microenvironment surrounding it in the mandible. Previously, we found that ECM molecules, NCAM, and tenascin-C are the main components of the SDL niche in alligators (Wu et al. 2013). In this study, we identified that a specific fibroblast subtype (DFDP), constituting the inner part (deciduous tooth side) of the SDL niche, upregulates biomechanical stress–related ECM genes and maintains the quiescent state of the SDL by secreting fibulin-1.
Mechanical Inputs Upregulate the Expression Level of Fibulin-1, Thereby Stabilizing the ECM Niche of the SDL
Several studies have indicated that niche-specific ECM controls stem cell differentiation (Alcaraz et al. 2008). Rigidity and other physical properties of the ECM regulate organ morphogenesis by affecting cell fate. Cells are coupled to ECM proteins in the niche via adhesion molecules. This coupling helps to maintain stem cell position, which affects cell fate decisions and self-renewal in their niche (Rompolas et al. 2013; Morita et al. 2021). However, the specific ECM molecule that regulates stem cells remains unknown. In our study, fibulin-1 was shown to be an important ECM protein in the SDL niche regulating the epithelial stem cell fate.
Macroscale mechanical inputs can be transmitted to ECM proteins, such as fibronectin, via the integrin cell surface receptor, after which they are transformed into nanoscale forces (Galbraith et al. 2002; Petridou et al. 2017). For example, mechanical cues regulate the stability of focal adhesion complexes to activate mechanoresponsive signals such as mitogen-activated protein kinases (Ras homolog [RHO] family member A, and RHO-associated protein kinase 1) (Vining and Mooney 2017). We previously identified that, at E60, the development of the DC exerted biomechanical stress on the SDL and its niche. In the present study, we further confirmed that biomechanical stress transmits signals to the ECM and mesenchymal cells through the RUNX2–fibulin-1 pathway.
Previously, we found that β-catenin and Lef1 were expressed in the dental follicle mesenchyme between deciduous teeth and the SDL of permanent teeth at E60 (Wu et al. 2020), indicating the activation of the canonical Wnt pathway in the DFDP. However, in this study, we found that the Wnt inhibitors APCDD1 and DKK3 were at the top of the gene expression list for the DFDP (Appendix Fig. 3A). The presence of Wnt inhibitors suggests that there is an elaborate balance of Wnt signaling in the DFDP. Whether and how Wnt signaling interacts with fibulin-1 need to be further investigated.
Conclusion
Our study suggests that fibulin-1, secreted by a specific fibroblast subtype located between the SDL and deciduous tooth, is a key ECM protein involved in niche homeostasis. Biomechanical stress–mediated RUNX2–fibulin-1 regulation emphasizes the need for future studies on the timing of organ renewal or regeneration.
Author Contributions
G. Li, contributed to data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; Z. Shen, contributed to data analysis, drafted and critically revised the manuscript; Q. Li, X. Lin, J. Wang, B. Zhao, Y. Feng, L. Feng, W. Guo, contributed to data acquisition, critically revised the manuscript; X. Li, contributed to data interpretation, critically revised the manuscript; L. Hu, J. Wang, C. Zhang, Z. Fan, contributed to data analysis, critically revised the manuscript; S. Wang, X. Wu, contributed to data conception and design, drafted and critically revised the manuscript. All authors gave their final approval and agree to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345231182052 – Supplemental material for Fibulin-1 Regulates Initiation of Successional Dental Lamina
Supplemental material, sj-docx-1-jdr-10.1177_00220345231182052 for Fibulin-1 Regulates Initiation of Successional Dental Lamina by G. Li, Q. Li, Z. Shen, X. Lin, X. Li, J. Wang, B. Zhao, Y. Feng, L. Feng, W. Guo, L. Hu, J. Wang, C. Zhang, Z. Fan, S. Wang and X. Wu in Journal of Dental Research
Footnotes
Acknowledgements
We thank Kun Jiao for performing the animal surgery.
A supplemental appendix to this article is available online.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Chinese Research Unit of Tooth Development and Regeneration, CAMSI Innovation Fund for Medical Sciences (no. 2019-12M-5-031); National Natural Science Foundation of China (grant no. 82030031 and 91649124 to S. Wang and no. 82022014 to X. Wu); Beijing Municipal Science & Technology Commission (no. Z181100001718208); Beijing Municipal Education Commission (no. 119207020201); Beijing Hospitals Authority of Hospitals’ Mission Plan (code SML20151401); and Beijing Municipality Government grants (Beijing Scholar Program-PXM2018_014226_000021, PXM2018_193312_000006_0028S643_FCG, PXM2019_014226_000011, PXM2020_014226_000005; Z181100001718208).
Data Availability
The scRNA-seq and spatial transcriptomics data can be found in Gene Expression Omnibus (GSE210143, GSE210142). The ChIP-seq data were obtained from NCBI GEO (GSE79813). Proteomic analysis data can be found in iProX under the accession of IPX0006452002.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.