
Abstract
Multiple genetic and environmental etiologies contribute to the pathogenesis of cleft palate, which is the most common of the inherited disorders of the craniofacial complex. Insights into the molecular mechanisms regulating osteogenic differentiation and patterning in the palate during embryogenesis are limited and needed for the development of innovative diagnostics and cures. This study used the Pax9-/- mouse model with a consistent phenotype of cleft secondary palate to investigate the role of Pax9 in the process of palatal osteogenesis. Although prior research has identified the upregulation of Wnt pathway modulators Dkk1 and Dkk2 in Pax9-/- palate mesenchyme, limitations of spatial resolution and technology restricted a more robust analysis. Here, data from single-nucleus transcriptomics and chromatin accessibility assays validated by in situ highly multiplex targeted single-cell spatial profiling technology suggest a distinct relationship between Pax9+ and osteogenic populations. Loss of Pax9 results in spatially restricted osteogenic domains bounded by Dkk2, which normally interfaces with Pax9 in the mesenchyme. Moreover, the loss of Pax9 leads to a disruption in the normal osteodifferentiaion of palatal osteogenic mesenchymal cells. These results suggest that Pax9-dependent Wnt signaling modulators influence osteogenic programming during palate formation, potentially contributing to the observed cleft palate phenotype.
Keywords
Introduction
The Wnt/β-catenin signaling pathway has an important regulatory role in the development and homeostasis of the skeleton (Krishnan et al. 2006; Joeng et al. 2017; Huybrechts et al. 2020), including the craniofacial complex. We and others have shown that Wnt signaling pathway genes play crucial roles in palatogenesis, specifically driving mesenchymal cell proliferation and differentiation (Jia, Zhou, Fanelli, et al. 2017; Jia, Zhou, Wee, et al. 2017; Li et al. 2017; Jia et al. 2020; Janeckova et al. 2023; Pina et al. 2023). Intracellular mediators of canonical Wnt signaling, such as β-catenin, promote lineage specification of mesenchymal stem cells and the differentiation of osteogenic precursors into functional osteoblasts and osteocytes (Glass et al. 2005; Cruciat and Niehrs 2013; Kim et al. 2013). Notably, mutations in coding sequences of Wnt factors result in skeletal dysplasias with clinical phenotypes that involve isolated alterations in bone mass (Regard et al. 2011; Martinez-Gil et al. 2022). Genome-wide association studies have implicated PAX9, the paired homeodomain-containing transcription factor, in craniofacial morphogenesis (Shaffer et al. 2016), hinting at the plausibility of genetic variants in this region influencing normal facial variation.
Pax9-/- mice offer a valuable resource for studies on palate development as they consistently demonstrate defects in elongation, delayed elevation, failed contact, and fusion of the palatal shelves (Peters et al. 1998; Jia, Zhou, Fanelli, et al. 2017). We have used Pax9-/- mice to demonstrate a unique molecular relationship between the Wnt signaling pathway and Pax9 within the posterior domain of the developing palate (Jia, Zhou, Fanelli, et al. 2017; Jia et al. 2020). Our results suggest that Pax9 shares a functional relationship with Dkk1 and Dkk2, specific modulators of Wnt signaling during secondary palate development (Jia et al. 2020). It is known that the Dkk1 and Dkk2 proteins block the binding of Wnt ligands to the low-density lipoprotein receptor–related protein (LRP) 5 and 6 at the cell surface (Semenov et al. 2001). This competitive inhibition of Wnt effector molecule function directly antagonizes the activation of Wnt signal transduction (Tamai et al. 2000). Further evidence of the Dkk’s role in Wnt signaling modulation is shown in our prior work, wherein the genetic reduction of Dkk1 during palatogenesis corrected secondary palatal clefts in Pax9-/- mice with restoration of Wnt signaling activities (Jia, Zhou, Fanelli, et al. 2017; Jia et al. 2020). Furthermore, ChIP-qPCR assays showed that Pax9 directly binds to regions near the transcription start sites of Dkk1 and Dkk2 (Jia et al. 2020). Thus, we concluded that the molecular mechanisms underlying Pax9’s role in modulating Wnt signaling activity during palate development is likely to involve the inhibition of Dkk1 and Dkk2 expression.
The advancement of single-cell and spatial transcriptomics technologies provides us with valuable tools to further explore the relationship between Pax9 and Wnt signaling. Here, we demonstrate that the global loss of Pax9 disrupted the spatial expression domains of key Wnt modulator genes—in particular, Dkk2—at critical phases of secondary palate formation. We present a Wnt-centered framework in high spatial resolution for osteogenic patterning and differentiation, the balance of which depends on the regulatory influence of Pax9. This study expands contemporary understanding of the functional roles played by Pax9 and its overlapping Wnt modulators to bring about differentiation and patterning morphogenesis of the secondary palate.
Materials and Methods
Animals
All animal procedures were approved by the National Institutes of Health, National Institute of Child Health and Human Development Animal Care and Use Committee, under Animal Study Protocol #21-031. C57BL/6J mice were obtained from the Jackson Laboratory. An inbred strain of female C57BL/6 mice was used for all experiments. Pax9+/− mice were provided by Dr. Rulang Jiang (Cincinnati Children’s Hospital) and generated as described previously (Zhou et al. 2013). Timed pregnancies were conducted via vaginal plug identification, with day 0.5 indicating date of identification. To ensure accurate developmental stage comparison, embryos comparatively analyzed were of the same litter, with the same C57BL/6 background. Genotyping of all mice included in this study was performed by Transnetyx, Inc. This study conforms to the ARRIVE 2.0 guidelines.
Single-Nucleus RNA + ATAC Sequencing (Multiome-RNA+ATAC-seq)
In this study, we generated a new single-nucleus gene (snRNA) and transposase-accessible chromatin (snATAC) multiome sequencing data set of microdissected Pax9-/- secondary palate tissue from embryonic day (E) 13.5 (n = 3 pooled biological replicates) for direct comparison with our previously generated multiome data set on normal secondary palate tissue from E13.5 and E15.5 C57BL/6 mouse embryos (Pina et al. 2023). To properly compare the new multiome data set to those generated in our prior study, all methodological steps were replicated as described previously (Pina et al. 2023).
Single-Nucleus RNA + ATAC-seq Bioinformatics Analysis
Raw fastqs were aligned to mm10 genome build using the standard Cellranger multiomics settings and imported to ArchR (v.1.0.1). Doublets were identified and filtered, and cells were filtered for minimum 4 transcription start sites (TSS) enrichment, 2,500 fragments per cell. Dimensionality reduction was performed using latent semantic indexing (LSI) based on the cell by fragment matrix and cell by gene matrix, and clusters were identified. Peaks were called based on original clustering, discarding reads from promoters (2,500 bp ± TSS) and exons. Cluster assignments were confirmed using canonical marker genes based on gene expression and gene ontology enrichment of marker genes identified using the getMarkerFeatures function of ArchR. Statistical significance and strength of enrichments were determined using t test, grouping cells by cluster. Detailed scripts and bioinformatic methodology of snRNA-seq analysis can be found on the Cotney Lab GitHub (https://github.com/emmawwinchester/mousepalate).
Micro–Computed Tomography (µCT) 3D Reconstruction and Segmentation
Whole heads from E15.5 samples (n = 3 biological replicates per genotype) fixed overnight (10% NBF) were scanned using a ScanCo µCT 50 ex vivo cabinet system (Scanco Medical) in 70% ethanol using the following parameters: 70 kVp, 85 µA, 0.5-mm AI filter, 900-ms integration time, and 10-µm voxel size. Three-dimensional rendering and segmentation of reconstructed files were performed using 3D Slicer software (slicer.org) using the semiautomatic segmentation function “Islands,” with standardized thresholding and island size.
Xenium In Situ mRNA Localization and Analysis
Xenium in situ subcellular mRNA detection technology was used as previously reported following all manufacturer guidelines and specifications (Janesick et al. 2023). We custom designed a 350-plex targeted gene panel (Appendix Table 1) to detect mRNA expression for cell-type identification as well as signaling pathway interactions selected and curated primarily based on single-cell atlas data we generated in our previous work on palate development (Pina et al. 2023). For this assay, we formalin fixed and paraffin embedded 3 biological replicates of Pax9-/- and 3 biological replicates of Pax9+/+ mouse embryos at embryonic day (E) 14.5. Whole heads were processed, embedded, and sectioned (4 µm) to the level of the first molar. All 6 samples were assayed in a single batch under identical conditions. The post-Xenium hematoxylin and eosin staining followed demonstrated protocol CG000160 from 10× Genomics, Inc. All analyses were generated and exported from the 10× Genomics open-access software, Xenium Explorer v1.2, following previously published guidelines from the manufacturer. Regions of interest were defined and standardized between sections analyzed, as were number of cells per frame. Features were filtered (mean object counts >1.0) and retained the top N features by L2FC for each cluster.
Results
A Pax9-Enriched Subcluster Is Transcriptionally Distinct from Other Mesenchymal Cell Populations in the Developing Palate
First, we further analyzed our recently published data sets assessing cell type–specific gene expression and binding motif enrichment using integrated snRNA and snATAC sequencing of micro dissected wild-type secondary palate tissue from C57BL/6 mouse embryos at E13.5 and E15.5 of development (Pina et al. 2023). Initial clustering and gene ontology enrichment analysis of marker genes from these populations determined the presence of 6 general cell types: epithelium, mesenchyme, muscle cells, neural cells, endothelium, and blood cells (Fig. 1A). Further isolated analysis of the mesenchyme identified 8 subtypes of this population. Canonical marker genes separate these cells into 4 broad categories: osteogenic cells, chondrogenic cells, generalized mesenchyme, and Pax9+ mesenchyme (Pina et al. 2023).
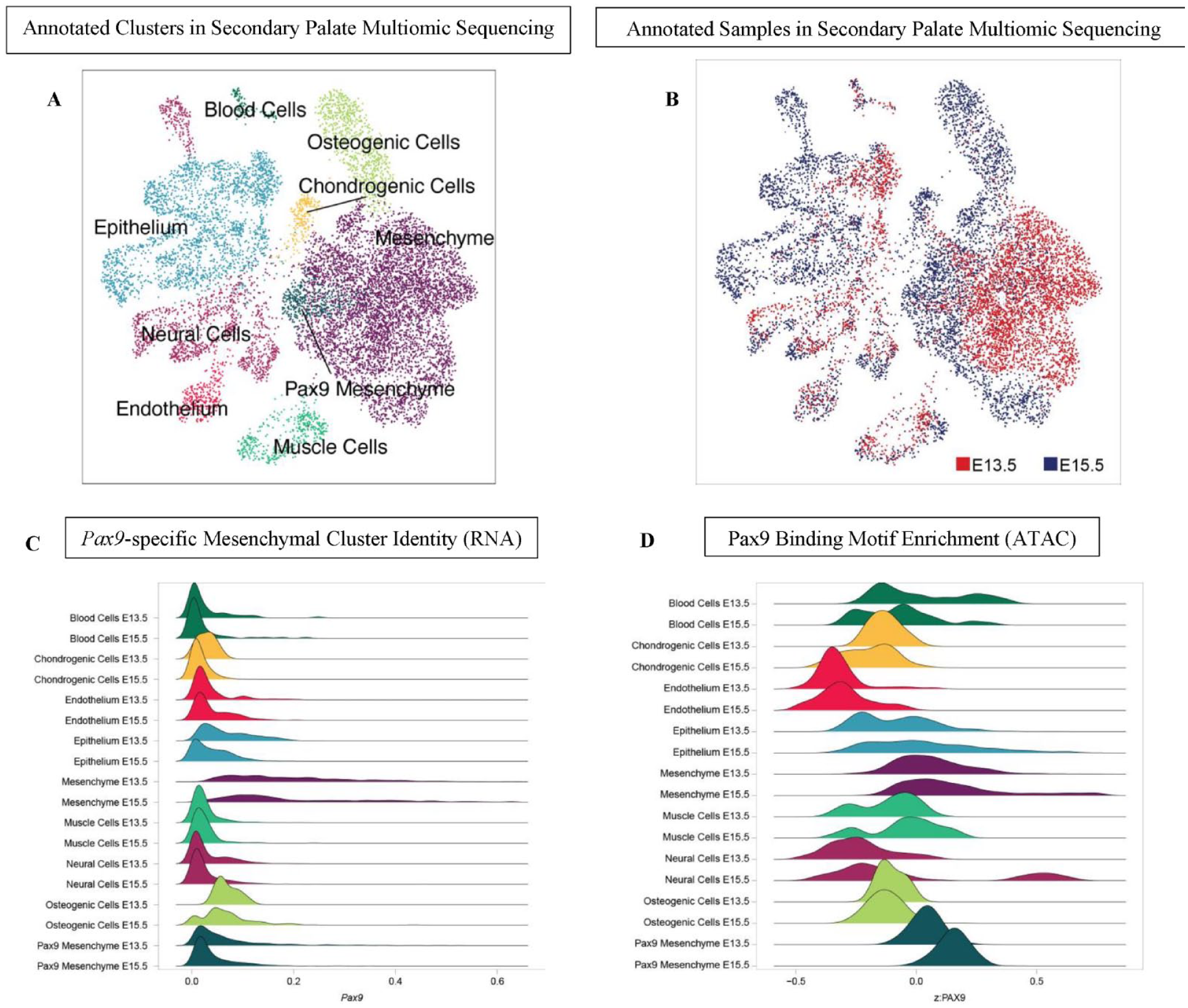
Single-nucleus multiomic sequencing (snRNA+ATAC-seq) of normal secondary palate identifies Pax9-enriched cell cluster distinct from other mesenchymal populations. (
When compared with all cell types in the developing palate, we observed that the population of Pax9+ mesenchymal cells demonstrated a 3.62 log2-fold enrichment of Pax9 expression (FDR 3.01e-04) (Fig. 1B). Similarly, we noted a bias of the Pax9 motif within peaks accessible within the Pax9 mesenchyme cluster, indicating bias of the Pax9 motif within regions of accessible chromatin open in this population (Fig. 1C, Appendix Fig. 1). Hence, Pax9 may influence osteogenic programming via Wnt signaling modulation of adjacent cell types within the developing palate mesenchyme, which exert paracrine effects on neighboring osteogenic cells.
Pax9 Is Needed for Complete Wnt-Mediated Palatal Osteogenesis
Next, we sought to understand the effects of global loss of Pax9 on the development of the secondary palate osteogenic-enriched population. µCT revealed 3D insufficiency of the mesial ossification front of the palatine bone at E15.5, despite indications of mature bone (Fig. 2A). Coronal cross-sectional analysis exhibited dense, spongy bone in Pax9-/- embryos (Fig. 2A). To better assess the downstream molecular consequences of the global loss of Pax9 expression in the secondary palate, we generated a new multiome-seq data set of murine E13.5 Pax9-/- palatal shelves to directly compare to our previously generated data set of normal secondary palate development at the same time point. Integrated multiome-seq analysis of wild-type and Pax9-/- data sets revealed a relatively greater proportion of early and late osteoprogenitor cells in E13.5 Pax9-/- secondary palate compared with the E13.5 wild-type secondary palate samples (Fig. 2B). Furthermore, the expression domain of key Wnt signaling modulators, Dkk1 and Dkk2, was notably expanded in the Pax9-/- secondary palate (Fig. 2C). Together, these morphological and molecular features of the Pax9-/- secondary palate could indicate a disruption in Wnt signaling dynamics—evidenced by the relative increase in expression of direct Wnt antagonists—which correlates to an alteration of the palatal ossification processes. This may suggest a critical role for Pax9-dependent Wnt signaling in maintaining proper palatal bone formation during development.
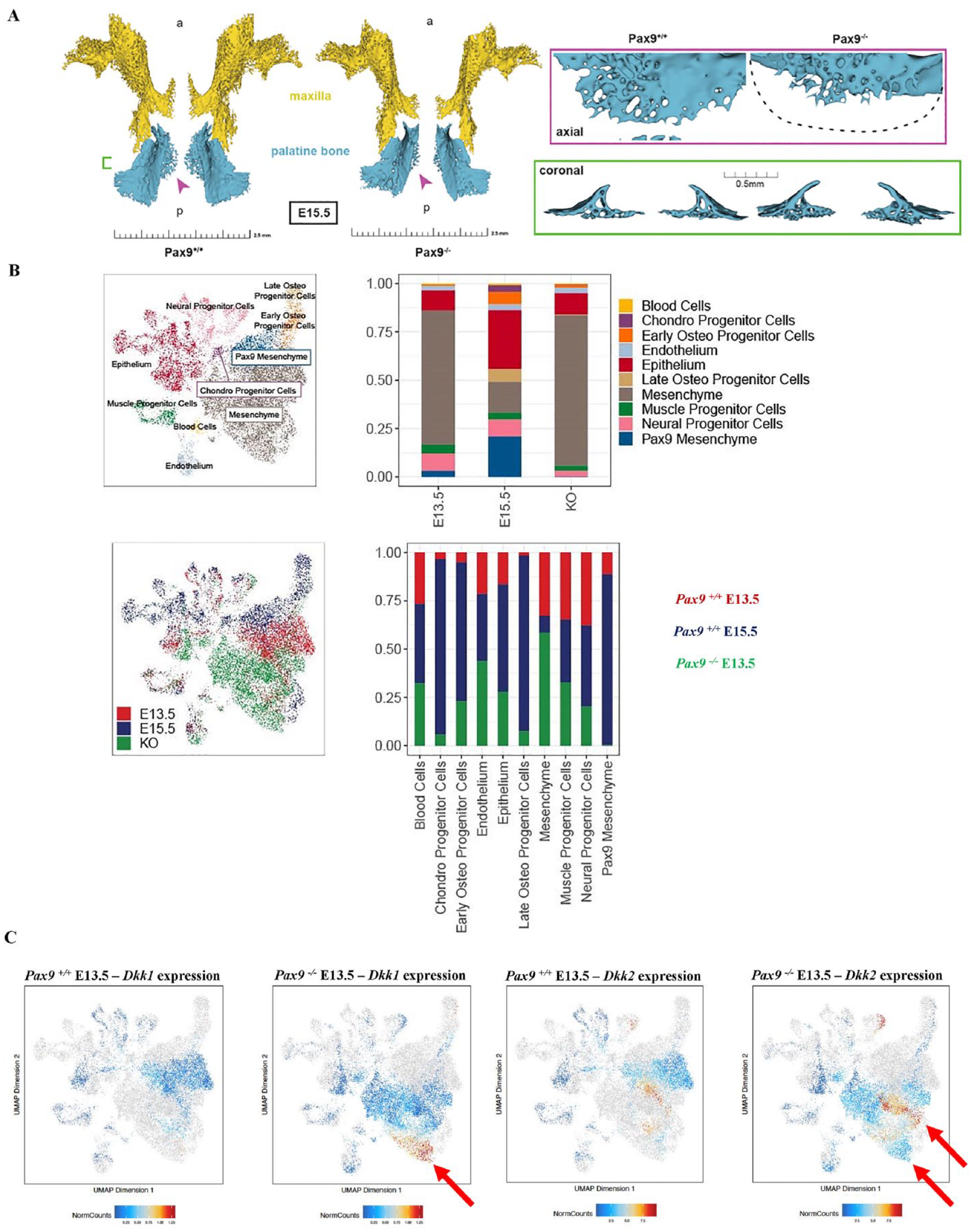
Loss of global Pax9 results in failure of palatal bone outgrowth and midline fusion, with a notable increase in Wnt modulator and osteoprogenitor cell expression. (
Wnt Inhibitory Genes Are Upregulated in the Palate in the Absence of Pax9
To study whether Pax9 regulates the commitment of palatal mesenchymal progenitor cells to the osteogenic lineage through Wnt signaling, we performed highly multiplex targeted single-cell spatial profiling analyses (Xenium in situ, 10× Genomics, Inc.). Age-matched (E14.5) whole embryonic heads from wild-type and Pax9-/- murine litters (n = 3 biological replicates per group) were included. As the first application of such spatial transcriptomic technology in developmental biology, we custom designed a panel of 350 unique marker gene probes (Appendix Table 1) to dissect differential cell types and effector-ligand signaling interactions within the secondary palate with and without the functional presence of Pax9.
Spatial transcriptomic analysis revealed 28 different cell clusters in both wild-type and Pax9-/- samples (n = 3 biological replicates per genotype were run and analyzed) (Fig. 3A, B). Transcriptomic signature differences were identified in differential expression analysis between wild-type and Pax9-/- samples. Clusters in the secondary palate region of interest were identified based on spatial localization within the coronal sections, and genes were analyzed within these clusters specifically. Clusters with enriched genes related to epithelium, osteogenic mesenchyme, extracellular matrix, and ciliated cells (both epithelium and mesenchyme) were identified for each sample analyzed in each group (Fig. 3C, Appendix Table 2). Notably, there were 2 distinct clusters (1, 7) identified in the Pax9+/+ palate samples related to epithelial cell marker gene expression, while there was only 1 distinct cluster (7) present in the Pax9-/- samples. We noted that epithelial expression of Wnt5a and Wnt7b was present in abundance in the Pax9+/+ palate epithelium, while neither was found to be enriched in cluster 7 of the Pax9-/- samples. In the osteogenic mesenchyme-associated clusters, the Pax9-/- palate demonstrated 5 (2, 8, 9, 10, 12) unique clusters of enriched genes, compared with just 4 (2, 3, 6, 10) clusters in the wild-type palate. Notably, Wnt signaling modulators, such as Sfrp2 differentially expressed in clusters 2 and 9, were identified to be enriched in the Pax9-/- palate osteogenic mesenchyme in the wild-type palate samples. Similarly, the extracellular matrix clusters revealed enhanced Wnt modulator expression as well as the appearance of expanded defining marker genes in the absence of Pax9, including the early osteoprogenitor cell marker, Crabp1. Finally, in the primary cilia-associated clusters, several genes showed a differential downregulation in the absence of Pax9, including Dynlrb2, Foxj1, Capsl, Mapk13, Krt7, Tuba1b, and Wnt7b. Taken together, across all cell types identified within the secondary palate, our in situ analysis provides additional evidence of a concordant genetic relationship between Pax9 and Wnt signaling during palate development (Fig. 3D).
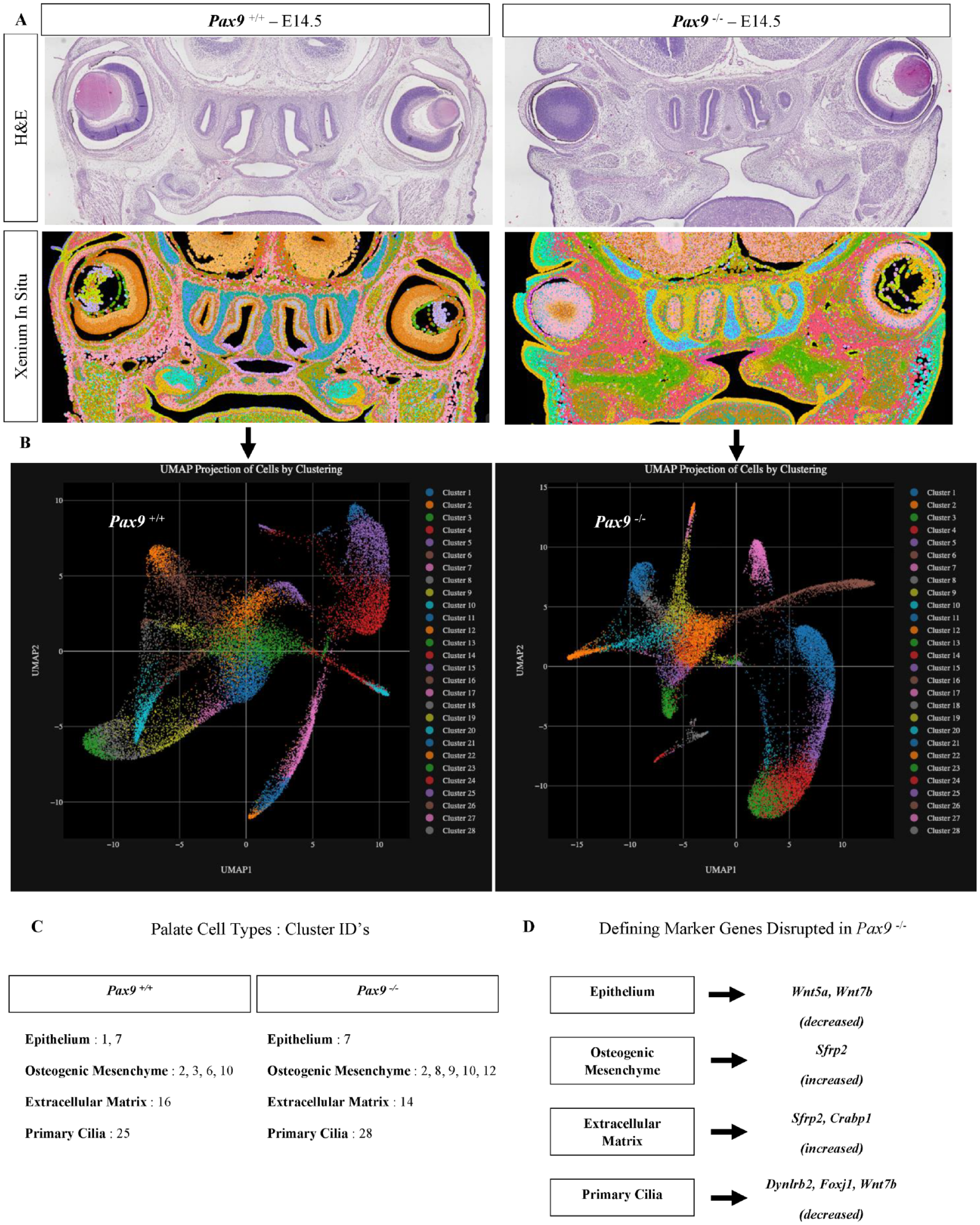
Highly multiplex in situ mRNA localization in normal and Pax9-/- palatal shelves reveal disruption of spatial patterning and mesenchymal progenitor cell differentiation. (
Disrupted Palatal Bone in Pax9-/- Cleft Is Bounded by Dkk2
Validating and spatially resolving our observations from the multiome-seq analysis, we observed a notable increase in the number of Dkk1 and Dkk2 transcripts localized within the Pax9-/- cleft palate mesenchyme (Fig. 4A). Interestingly, Dkk1’s expression domain is localized primarily within the osteogenic zones, while Dkk2’s expression bounds the mesial border of midline osteogenic extension of the palatine bone (Fig. 4B). We further validated the expression profiles of Dkk1, Dkk2, and Sp7 initially identified via Xenium in situ analysis in subcellular spatial resolution using our well-established RNAscope multiplex in situ hybridization protocol (Appendix Fig. 2).
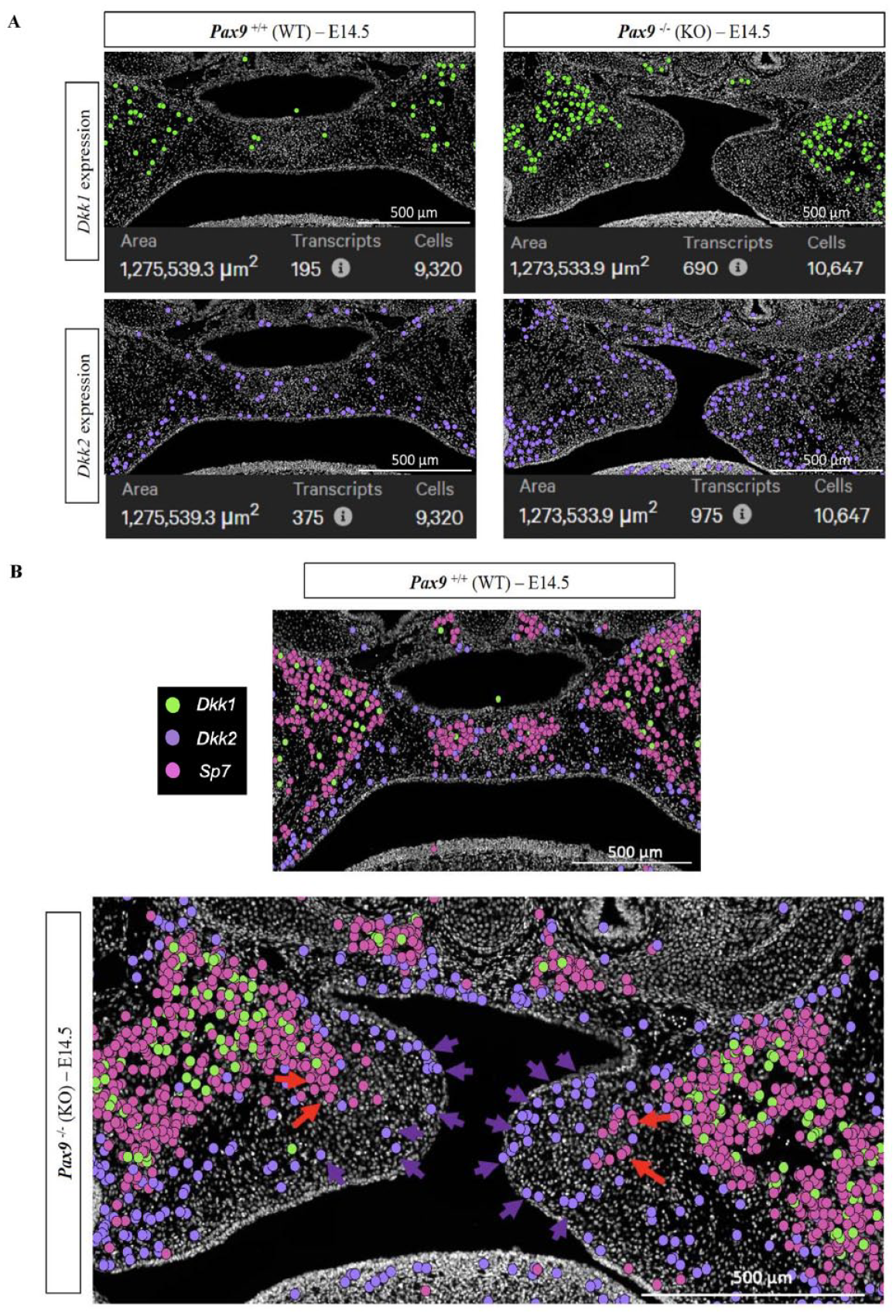
Spatial dysregulation of Wnt modulator expression within Pax9-/- palatal shelves. (
Upon further query of differentially expressed marker genes from overlapping signaling pathways included in our custom panel, we noted spatial alteration in expression domains of Fgfr1, Gli1, Bmp2, and Bmp4 in the Pax9-/- palatal shelves, notably in the mesial aspect of the elevated shelves, the domain of Pax9 enriched expression in normal palatal shelves (Appendix Fig. 3, Appendix Tables 3 and 4). Given the likely observed alteration in osteodifferentiation dynamics in the Pax9-/- palate compared with the normal palate, we further analyzed a spectrum of osteogenic differentiation marker genes to capture differential changes in osteoprogenitors (Six2), preosteoblasts (Runx2), osteoblasts (Sp7), immature osteocytes (Dmp1), and mature osteocytes (Sost) (Appendix Fig. 4). Finally, we differentially analyzed the expression pattern of the patterning molecule, Osr1, to further assess the potential spatial patterning effects with and without the presence of the patterning morphogen, Pax9. Our Xenium in situ analysis identified a differential increase in Osr1 expression in Pax9-/- palatal shelves, particularly in the mesial oral domain (Appendix Fig. 5). This higher spatial resolution of specific mesenchymal compartment gene enrichment extends and enlightens our prior studies on these molecules in situ, suggesting that the loss of Pax9 may lead to impaired osteogenic differentiation, as relatively higher transcript counts of less mature markers (Six2, Runx2, and Sp7) were localized within the Pax9-/- palatal shelves (Appendix Table 5). Moreover, the mesial enrichment of Dkk2 (alongside overlapping pathway effector deficiencies and the absence of extension of Sp7 toward the midline as is seen in the normal palate) may hint at its functional role within the Pax9-/- cleft palate, actively antagonizing Wnt-driven osteogenic extension to the midline.
Discussion
Pax9 is a master orchestrator of patterning and morphogenesis throughout the body, from the thymus, ultimobranchial bodies, limb bud, and tooth organ to the axial skeleton (Peters et al. 1998; Peters et al. 1999). This highly conserved transcription factor has been shown to functionally interact with a number of mesenchymal factors and signaling pathways required for palate development to ensue, which act in concert to regulate cell proliferation and differentiation of cranial neural crest–derived mesectoderm (Neubuser et al. 1997; Hilliard et al. 2005; Ichikawa et al. 2006; Jia, Zhou, Fanelli, et al. 2017; Li et al. 2017; Jia et al. 2020). The role of Pax9 in patterning during development of multiple organ systems, including the palate, is clearly established. However, prior to the present study, this has not been explored sufficiently in the context of palate osteogenesis.
The resultant cleft palate phenotypes in the setting of either premature (Mori-Akiyama et al. 2003; Sweat et al. 2020) or delayed (Baek et al. 2011) palate osteogenesis exemplify the importance of precise temporal onset and spatial regulation of intramembranous ossification. Intriguingly, and supportive of our hypothesis that Wnt-mediated osteogenic programming is disrupted in the absence of Pax9, a premature ossification and cleft palate phenotype is described in Sox9-/- (Mori-Akiyama et al. 2003) and Six2-/- mice (Sweat et al. 2020). Sweat et al. (2020) further noted a direct upstream regulatory relationship between the osteoprogenitor transcription factor, Six2, and Pax9, supporting a predicted role for Pax9 in progression of palate osteogenesis. The spatial transcriptomic alterations in Pax9-/- secondary palate cell clusters—specifically, the upregulation of Dkk2 surrounding the midline extension of palatal bone—provide evidence of Pax9’s functional role in regulating the differentiation and maturation of specific palatal mesenchymal progenitor cells. The spatial patterning disruption in the absence of Pax9 in the palatal bone is further delineated in the present work with the finding that a key osteogenic patterning molecule, Osr1, is upregulated in the mesial oral domain of the Pax9-/- palatal shelves. As Osr1 functions in orchestrating appropriate skull closure and cranial bone formation by negative regulation, this upregulation could help explain the disrupted osteogenic differentiation as well as midline extension of growth in the Pax9-/- cleft palate.
Also of notable intrigue was the finding that certain primary cilia-related genes were downregulated in the Pax9-/- cleft palate—including one of the novel reported genetic markers from our prior study of normal palatogenesis, Dynlrb2 (Pina et al. 2023)—as there is evidence of canonical Wnt signaling pathway involvement in primary cilia function (Janeckova et al. 2023). However, beyond the scope of the present work, more extensive functional studies on the connection between Pax9 and palatal mesenchymal osteoprogenitors in orchestrating their respective signaling pathways and cellular differentiation networks are needed.
The differential upregulation of Dkk1 and Dkk2 within the Pax9-/- secondary palate mesenchyme is supportive of our previous foundational work in which these 2 molecules were targeted with small-molecule Wnt agonist therapeutics for the correction of embryonic cleft secondary palate (Jia, Zhou, Fanelli, et al. 2017). Wnt signaling is known to directly influence Runx2 transcription through its proximal promoter TCF binding site (Gaur et al. 2005). Our prior work highlighted this concept in the release of overall Wnt inhibition through Wise genetic deletion and Wnt-agonist treatment to be sufficient to rescue the cleft palate and restore osteogenic domains to varying extents (Jia et al. 2017; Li et al. 2017). Importantly, small-molecule Wnt agonist IIIc3a’s inhibition of Dkk1, Dkk2, and Dkk4 was 20% more effective to correct cleft palate in utero than WAY262611 inhibition of Dkk1 alone (Jia, Zhou, Fanelli, et al. 2017).
Given the 3D nature of the developing a secondary palate, contemporary 2D methods to study genetic regulators notably limit our ability to ascertain their spatial profiles comprehensively. While this is the first application of such in situ highly multiplex targeted single-cell spatial profiling technology in developmental biology, it will be necessary to continue its use (alongside other emerging spatial technologies) to study multiple domains of the palate. Furthermore, additional functional studies to dissect the roles of Dkk1 and Dkk2 in the normal and abnormal palate during development will provide further foundational knowledge requisite to advance targeted therapeutic strategies.
In summary, this study addresses the perturbation of osteogenic patterning and differentiation in relation to Wnt signaling mediators in the Pax9-/- genetic model of secondary cleft palate. We generated new multiomic and in situ spatial transcriptomic data sets of normal and abnormal palatogenesis to characterize the disrupted midline osteogenic extension of the palatine bone in the Pax9-/- cleft secondary palate, alongside its perturbation in Wnt signaling dynamics, pointing to Pax9’s likely role in patterning and tuning Wnt-driven osteogenesis in the palate. We highlight for the first time, a spatial dysregulation of Wnt modulators and their overlapping pathway effectors in the Pax9-/- palate, specifically identifying Dkk2 enriched in a boundary surrounding the osteogenic midline extension within palatal shelves of Pax9-/- mouse embryos. Importantly, the loss of Pax9 may lead to impaired maturation of palatal osteogenic mesenchymal cells, further evidence of its role as a critical transcriptional regulator of patterning and morphogenesis. This contribution to our fundamental understanding of the molecular and cellular underpinnings of a cleft palate phenotype strengthens the framework required to assess the safety and efficacy of targeted therapeutic strategies to prevent or correct cleft palate disorders. Coupled with our prior proof-of-concept in vivo correction of cleft palate defects in Pax9-/- mouse embryos, this additional knowledge of Wnt signaling perturbation allows for more precise preclinical testing of Food and Drug Administration–approved Wnt agonist therapies to correct such defects, both in utero and postnatally.
Author Contributions
J.O. Piña, contributed to conception and design, data acquisition and interpretation of data, performed all statistical analyses, drafted and critically revised the manuscript; R. Raju, F.R. Faucz, contributed to conception and design, data acquisition and interpretation of data, critically revised the manuscript; D.M. Roth, C. Padilla, contributed to data acquisition, critically revised the manuscript; E.W. Winchester, J. Iben, contributed to data acquisition and interpretation of data, critically revised the manuscript; J.L. Cotney, contributed to interpretation of data, critically revised the manuscript; R.N. D’Souza, contributed to conception and design, data interpretation of data, critically revised the manuscript. All authors gave their final approval and agree to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345241256600 – Supplemental material for Spatial Multi-omics Reveals the Role of the Wnt Modulator, Dkk2, in Palatogenesis
Supplemental material, sj-docx-1-jdr-10.1177_00220345241256600 for Spatial Multi-omics Reveals the Role of the Wnt Modulator, Dkk2, in Palatogenesis by J.O. Piña, R. Raju, D.M. Roth, E.W. Winchester, C. Padilla, J. Iben, F.R. Faucz, J.L. Cotney and R.N. D’Souza in Journal of Dental Research
Footnotes
Acknowledgements
We thank our lab manager, Parna Chattaraj, for her administrative support throughout manuscript preparation; lab member Dr. Fahad Kidwai for administrative oversight of animal colony logistics; lab member Dr. Aye Chan Myo for her technical assistance during revision of this work; Dr. Sergey Leiken (NIH/NICHD) for his technical and analytical support in establishing our independent next-generation sequencing workflow and Xenium in situ assays; Dr. Blake Warner (NIH/NIDCR) for his support in Xenium instrument access; Dr. Vardit Kram (NIH/NIDCR) for her guidance with µCT scanning and analysis; and Dr. Ricardo D. Coletta (University of Campinas, FOP, Brazil) for his review and feedback during manuscript preparation and to all reviewers and editors at JDR who contributed to the overall composition of this final work.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: ZIA HD009015 “Wnt Signaling Pathway: Genes in Craniofacial Development: Palate and Tooth,” National Institutes of Health, National Institute of Dental and Craniofacial Research (RDS); National Institutes of Health, National Institute of Child Health and Human Development, Intramural Research Training Award (JOP/DMR); 1F30DE031149-01, 5R03DE028588-02, 2R35GM 119465-06, 5R01DE028945-04 (JLC).
A supplemental appendix to this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.