
Abstract
Dental caries remains a globally prevalent disease, driven by microbial dysbiosis and competitive ecological shifts within the oral biofilm. A key interaction in this context is the antagonism between Streptococcus mutans and Streptococcus sanguinis, two early colonizers of the dentin surfaces. While bacterial extracellular vesicles (bEVs) have emerged as important mediators of microbial communication, their role in interspecies adhesion and early biofilm dynamics remains poorly understood. This study explored how S. mutans–derived bEVs, isolated from planktonic cultures and biofilms grown on native or glycated type I collagen substrates, modulate S. sanguinis nanoadhesion and initial biofilm formation. Bacterial EVs were characterized using nanoparticle tracking analysis and transmission electron microscopy, and their functional effects were assessed via atomic force microscopy–based single-cell force spectroscopy. This approach enabled direct quantification of bacterial adhesion forces and single-molecule unbinding events under different vesicle-exposure conditions. Our results demonstrated that planktonic-derived bEVs enhanced S. sanguinis adhesion at 5-s contact times, while bEVs from collagen-bound S. mutans biofilms, particularly those grown on glycated matrices, consistently reduced adhesion forces, rupture lengths, and unbinding events. Optical coherence tomography imaging confirmed that these nanoscale effects translated into altered early biofilm architecture, with planktonic bEVs promoting thicker, denser biofilms and biofilm-derived bEVs leading to sparser colonization. Our findings suggest that S. mutans bEVs exhibit a context-dependent modulatory effect on S. sanguinis, enhancing adhesion under planktonic conditions and suppressing it upon biofilm establishment. This biphasic behavior may represent a strategic mechanism for niche domination during caries initiation. Moreover, collagen glycation—mimicking aged or hyperglycemic dentin—further influenced bEV function, underscoring the importance of the host matrix state in microbial interactions. Overall, this study highlights a previously unrecognized role for bEVs in shaping early oral dysbiosis at the single-cell level.
Keywords
Introduction
Dental caries remains one of the most prevalent human diseases, despite recent advances in prevention and treatment strategies (Wen et al 2021). It is a chronic, multifactorial condition primarily driven by dysbiosis within the local microbial community on the tooth (Pitts et al 2017; Du et al 2021). Among the ecological interactions that shape this community, one of the most extensively studied is the competitive relationship between the cariogenic Streptococcus mutans and commensal streptococci such as Streptococcus sanguinis (Giacaman et al 2015; Guo et al 2019). The overgrowth of S. mutans is strongly associated with surface demineralization and the progression of caries lesions, due to continued local pH reduction as a result of carbohydrate metabolism (Sheiham and James 2015; Kim et al 2020). Therefore, host, microbial, and environmental factors that promote the predominance of S. mutans over S. sanguinis and other competing species play a crucial role in the initiation and progression of dental caries.
The colonization of dentin—the most abundant mineralized dental tissue—is particularly noteworthy due to its complex microarchitecture and its type I collagen-rich organic matrix (Goldberg 2011; Álvarez et al 2024). Within this environment, oral microbes can utilize collagen as a key substrate for biofilm formation facilitated by a variety of collagen-binding proteins expressed on the bacterial surface (Avilés-Reyes et al 2017; Álvarez et al 2021). Notably, the collagen matrix of both teeth and the periodontal ligament is susceptible to glycation, a process in which sugars and their derivatives, such as methylglyoxal (MGO), irreversibly bind to proteins altering their biological and mechanical properties over time (Bansode et al 2020; Haluszka et al 2022). Glycation is particularly relevant in elderly individuals and those with chronic hyperglycemia, raising concerns about its broader impact on oral health in aging populations. Importantly, glycation has been shown to enhance the adhesion of key pathogenic bacteria including S. mutans, Porphyromonas gingivalis, and Staphylococcus aureus; thus, it is believed to modulate oral and systemic host–pathogen interactions (Xie et al 2020; Śmiga et al 2021; Leiva-Sabadini et al 2023). In this ecological context, the initial colonization of aged dentin—both its collagen matrix and inorganic mineral phase—depends on the contrasting dynamics of S. mutans and S. sanguinis, each responding differently to the structural and mechanical cues presented by the tooth surface (Leiva-Sabadini et al 2023). This early adhesion may subsequently shape the microbial community development toward either health or disease in the context of root carries, including subsequent interactions with other key microbial strains such as the relevant fungi Candida albicans (Du et al 2021; Zhang et al 2025), underscoring the importance of exploring oral streptococci–surface interactions during the initial process of dentin attachment and root caries.
As part of the biofilm, oral bacteria—including S. mutans—have evolved and optimized a variety of strategies to enhance survival and gain a competitive advantage within the microbiome (Kim et al 2020). Among these strategies, one key discovery is the production of bacterial extracellular vesicles (bEVs): nanoscale vesicles ranging from 20 to 200 nm in diameter, secreted by both gram-negative and gram-positive bacteria (De Langhe et al 2024). Oral bEVs have emerged as key mediators of intercellular communication, influencing microbial quorum-sensing, host–pathogen interactions, and biofilm formation (Liao et al 2014; Leiva-Sabadini et al 2024). However, the specific role of S. mutans–derived bEVs in interspecies communication among oral streptococci remains poorly understood. Particularly, the potential of S. mutans bEVs to modulate the initial attachment of S. sanguinis to dentinal substrates, such as the organic type I collagen matrix, and to influence early biofilm formation has not been explored. Furthermore, it remains unknown whether the transition of S. mutans planktonic to biofilm states, or the glycation of collagen, induces functional changes in the activity of its bEVs with respect to interspecies communication and bacterial adhesion.
Advanced experimental techniques such as atomic force microscopy (AFM)–based single-cell force spectroscopy (SCFS) enable real-time characterization of bacterial adhesion and subcellular levels using living cells. In this context, AFM-based SCFS has been used to investigate interactions between oral bacteria and various surfaces of interest with nano- to pico-newton sensitivity (Olivares et al 2025). Despite its promise, SCFS has not yet been applied to access the influence of bEVs on the nanoscale adhesion dynamics of living bacteria in the context of cariogenic biofilm development and early oral dysbiosis. To address this gap, this present work aimed to determine the impact of S. mutans bEVs on the surface adhesion of S. sanguinis, considering both biofilm formation and collagen glycation. This was achieved through a combination of noninvasive techniques, AFM-based SCFS and optical coherence tomography (OCT).
Methodology
Bacterial Strains and Culture Conditions
For all experiments, S. mutans UA159 and S. sanguinis SK36 strains were kept at −80 °C and cultured on brain heart infusion (BHI) agar plates or culture medium at 37 °C under aerobic conditions for S. mutans and 37 °C with 5% CO2 for S. sanguinis.
Collagen Coating and Glycation of Substrates
Twelve-well plates were coated with a thin collagen layer, using 250 µL of rat tail type I collagen (First Link) at a final concentration of 0.5 mg/mL. Substrates were incubated at 37 °C for 1 h, after which the supernatant was discarded. For glycated collagen, 10 mM of MGO (Sigma-Aldrich, M0252) was added and incubated for 3 d under the same conditions and copiously washed with sterile 1× phosphate-buffered saline (PBS).
Isolation of Planktonic and Biofilm bEVs
To obtain planktonic bEVs, a culture of S. mutans (1 × 107 CFU in 100 mL BHI) was allowed to grow for 24 h in a 250-mL flask at 37 °C (PLK_bEVs). The same concentration and BHI medium was used to initiate biofilm growth on control collagen (Col_bEVs) and glycated collagen (MGO_bEVs). For this, biofilms were grown on collagen-coated wells for 24 h at 37 °C, after which the supernatant was discarded to remove unbound cells, and bound cells were harvested with a cell scraper. From the collected bacterial cells, a series of centrifugation, filtration, and ultracentrifugation at 122.000 × g (Beckman L-70 ultracentrifuge, SW28 rotor) at 4 °C was used to isolate bEVs from all groups, using a previously described protocol (Leiva-Sabadini et al 2025). Samples were stored at −80 °C until used.
Characterization of S. mutans bEVs
To assess size and concentration, nanoparticle tracking analysis was used (NS300 NTA, Malvern Panalytical). Sample stocks were diluted 1:80 in filtered 1× PBS with a camera level of 14 and a capture time of 20 s and measured in triplicate for 12 to 130 particles per frame. The bEV concentration and particle size were analyzed using NanoSight 3.1 software. For transmission electron microscopy (TEM), bEVs were incubated on Formvar/carbon-coated copper meshes (Ted Pella), counterstained with 2% uranyl acetate, and visualized with TEM (Talos F200C G2 system).
AFM SCFS of S. sanguinis Adhesion
To study the changes in the adhesion of S. sanguinis after exposure to S. mutans EVs, following growth in BHI, 7.5 × 106 CFU/mL of S. sanguinis was incubated for 30 min at 37 °C + 5% CO2 with 5 × 108 particles/mL of either PLK_bEVs, Col_bEVs, MGO_bEVs, or 1× PBS (control).
Following bEV incubation, single-cell force nanoscopy was carried out by fabricating functionalized living-cell AFM probes. After Nanowizard IV AFM (JPK Instrument) calibration using MSNL-10 (cantilever D, K = 0.03 N/m, Bruker) obtaining spring constant values between 0.038 and 0.056 (N/m), cantilevers were immersed in a 20-µL droplet of 0.1% w/v poly-L-lysine solution (PLL; Sigma) for 2 min, rinsed with nuclease-free H2O, and air-dried for 10 min. Subsequently, a 20-µL droplet of S. sanguinis was added to a sterile petri dish, in which single cells were identified and softly approached with the PLL-coated AFM for 30 s. Bacterial immobilization was confirmed when the AFM tip was retracted from the petri dish.
Once the functionalized cell probes were created, SCFS measurements were carried out in PBS buffer against collagen substrates fabricated with a previously published protocol (Vaez et al 2023). Bacterial probes were approached to the substrate with a soft setpoint of 0.5 nN, 1-µm Z-length, and a 2-µm/s velocity. For each probe, 400 force curves were generated for 0-s and 5-s contact times. Three biological replicates were made for each condition using independent MSNL-10D living-cell cantilevers and independent collagen gels. The maximal adhesion force and single minor-unbinding rupture lengths and adhesion forces were extracted from the resulting force curves (Fig 1). Contour lengths for single-unbinding events were predicted using the wormlike chain model. All data were analyzed using the proprietary JPK Data Processing 6.4.27 software.
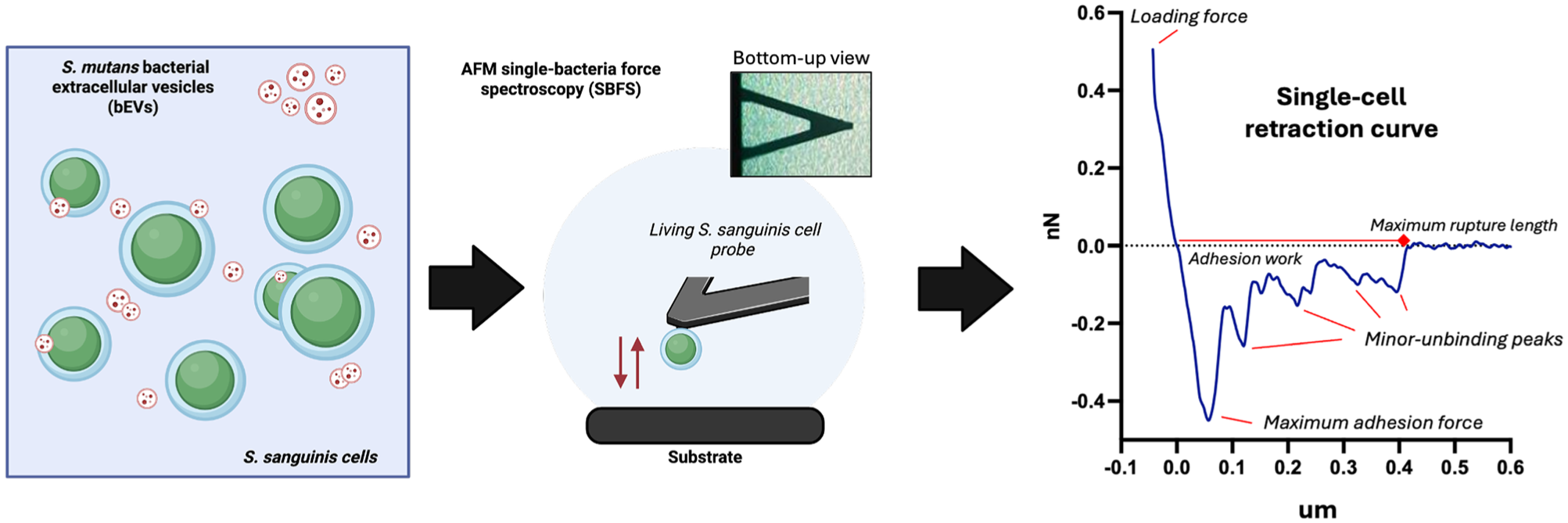
Experimental setup for Streptococcus sanguinis living-cell single-bacteria force spectroscopy (SBFS). Diagrammatic representation of the co-incubation of living S. sanguinis cells with S. mutans–derived bacterial extracellular vesicles for SBFS experiments and an example retraction force–distance curve from the experimental setup. The force curve diagram highlights nanomechanical parameters such as the maximum adhesion force, maximum rupture length, and minor unbinding peaks (representing single unbinding events between cell probe and substrate). Each one of these minor unbinding peaks consists of an adhesion force value and a rupture length, which is extracted and tabulated during data analysis.
OCT Exploration of Early S. sanguinis Biofilms
Following single-cell experiments, OCT was used to assess the impact of bEVs on the early formation of S. sanguinis on the inorganic component of dentin. For this, opaque autoclaved hydroxyapatite discs with a 9.5-mm diameter (Clarkson Chromatography Products Inc.) were placed into 24-well plates with BHI media supplemented with 1% sucrose to promote biofilm formation. In each well, 1.25 × 106 CFU/mL of S. sanguinis and 5 × 108 bEVs/mL were inoculated and incubated at 37 °C and 5% CO2 for 72 h, replenishing the bEVs every 24 h.
Following incubation, a VivoSight OCT scanner (Michelson Diagnostic Ltd.) was used to measure the cross-sectional thickness of the biofilms. Each disc was carefully placed into a 60-mm petri dish and covered with approximately 4 mL of 1× PBS. We obtained 6 × 6 mm2 scan areas with 120 frames for each condition and a pixel size of 4.65 µm, and the images were analyzed using ImageJ v1.54 software. Each condition was analyzed 3 times to acquire 3 representative images per condition, and 5 distance measurements of both the high- and low-density areas of each biofilm were extracted (Pattem et al 2018).
Statistical Analysis
All data were tabulated and analyzed using GraphPad Prism 8 software. After normality and variance homogeneity testing, Kruskal–Wallis with Dunn’s multiple comparisons post hoc tests (nonparametric) or 1-way analysis of variance with Tukey’s multiple comparisons test (parametric) were used to determine significant differences among bEV groups, considering significance at P < 0.05.
Results
Biofilm-Derived S. mutans bEVs Modulate the Early Attachment of S. sanguinis to Substrates
Using our established protocol, bEVs were successfully isolated from S. mutans grown under 3 conditions: planktonic (PLK_bEVs) cultures, biofilms on control type I collagen (Col_bEVs), and biofilms on glycated type I collagen (MGO_bEVs) (Appendix Fig 1). Notably, biofilm-derived EVs were smaller in size compared with PLK_bEVs. In addition, Col_bEVs exhibited a lower particle concentration (particles/mL) than those from the other 2 experimental groups. For all experimental conditions, isolated bEVs were co-incubated with S. sanguinis prior to their immobilization onto functionalized AFM cantilevers to assess real-time bacterial nanoscale adhesion (Fig 1).
SCFS experiments revealed that S. sanguinis nanoadhesion to collagen substrates was differentially modulated by bEV treatment across the different experimental groups. At 0-s contact times, PLK_bEVs and MGO_bEVs significantly reduced S. sanguinis adhesion (Fig 2A and B; Appendix Fig 2). In contrast, the opposite effect was seen for Col_bEVs (Fig 2A and B). However, at extended cell-surface contact times (5 s), the trend shifted: S. sanguinis treated with biofilm-derived S. mutans bEVs exhibited reduced nanoadhesion, while treatment with PLK_bEVs led to increased adhesion. In all cases, increasing the contact time resulted in higher adhesion forces and a greater number of single-unbinding events, as illustrated in the representative force-distance curves (Fig 2C).
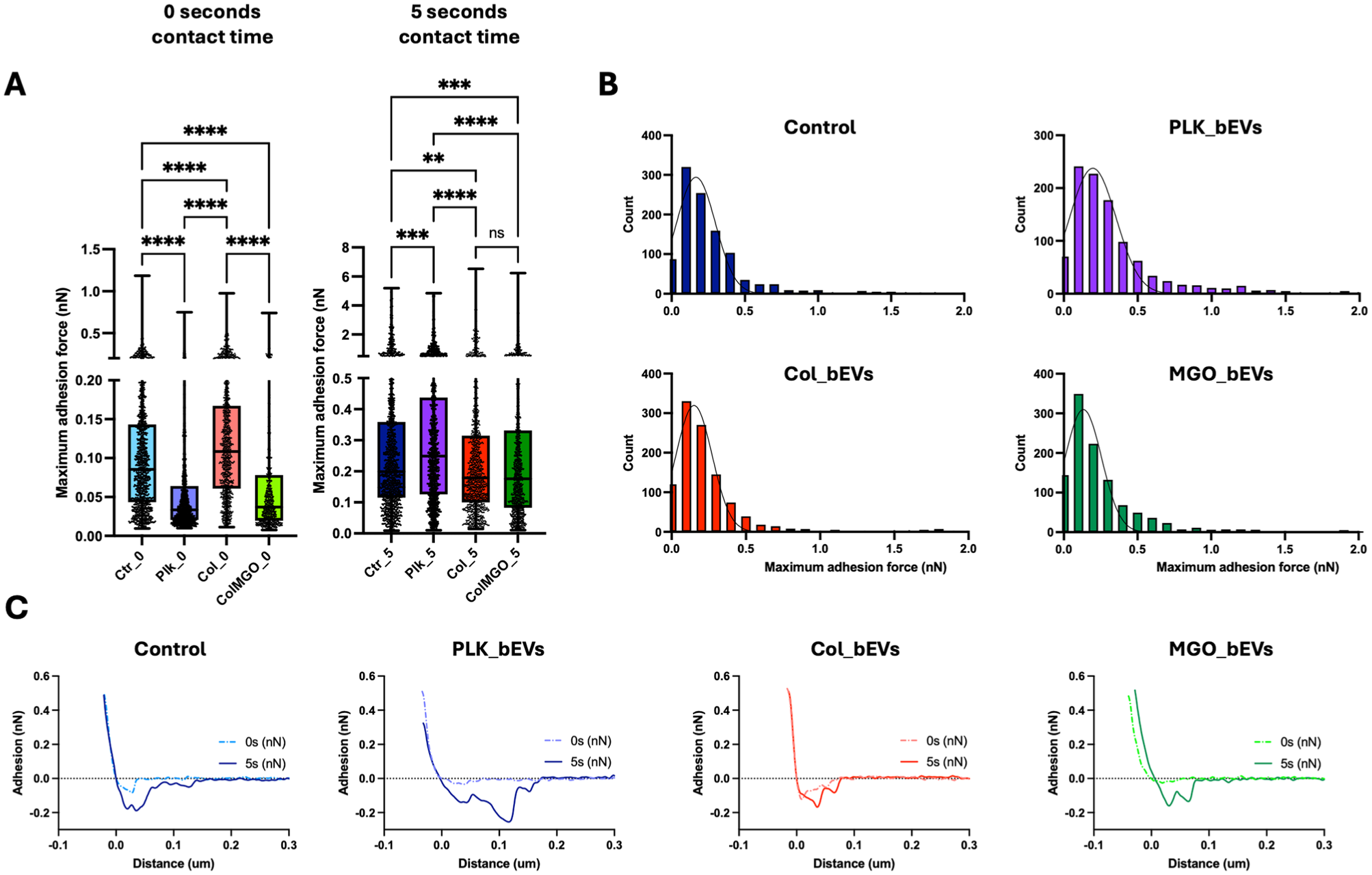
Streptococcus mutans–derived bacterial extracellular vesicles (bEVs) modulate the early nanoadhesion of Streptococcus sanguinis to biological surfaces. (
In addition to adhesion forces, the rupture length of the S. sanguinis–collagen single-molecule interactions following bEV exposure was also assessed (Fig 3). Across all experimental groups, increasing the contact time to 5 s resulted in longer rupture lengths. The shortest rupture lengths were observed in the MGO_bEVs group, with a median of 97 nm at 5 s, compared with 115 nm, 122 nm, and 107 nm for the control, PLK_bEV, and COL_bEV groups, respectively (Fig 3). When observing the rupture force of each unbinding unit, higher forces were observed for contour lengths in the ~400- to 800-nm range across all groups, with peak values reaching ~6 nN (Fig 4A). Notably, the minor unbinding peaks—associated with adhesin–collagen interactions—were significantly lower in the MGO_bEVs group at 5-s contact times compared with all other experimental groups (Fig 4B). A comprehensive summary of all SCFS-derived adhesion parameters is presented in Figure 4B, highlighting that biofilm-derived S. mutans bEVs, particularly those isolated from biofilms on glycated substrates, led to reduced S. sanguinis nanoadhesion, rupture forces, and contour lengths.
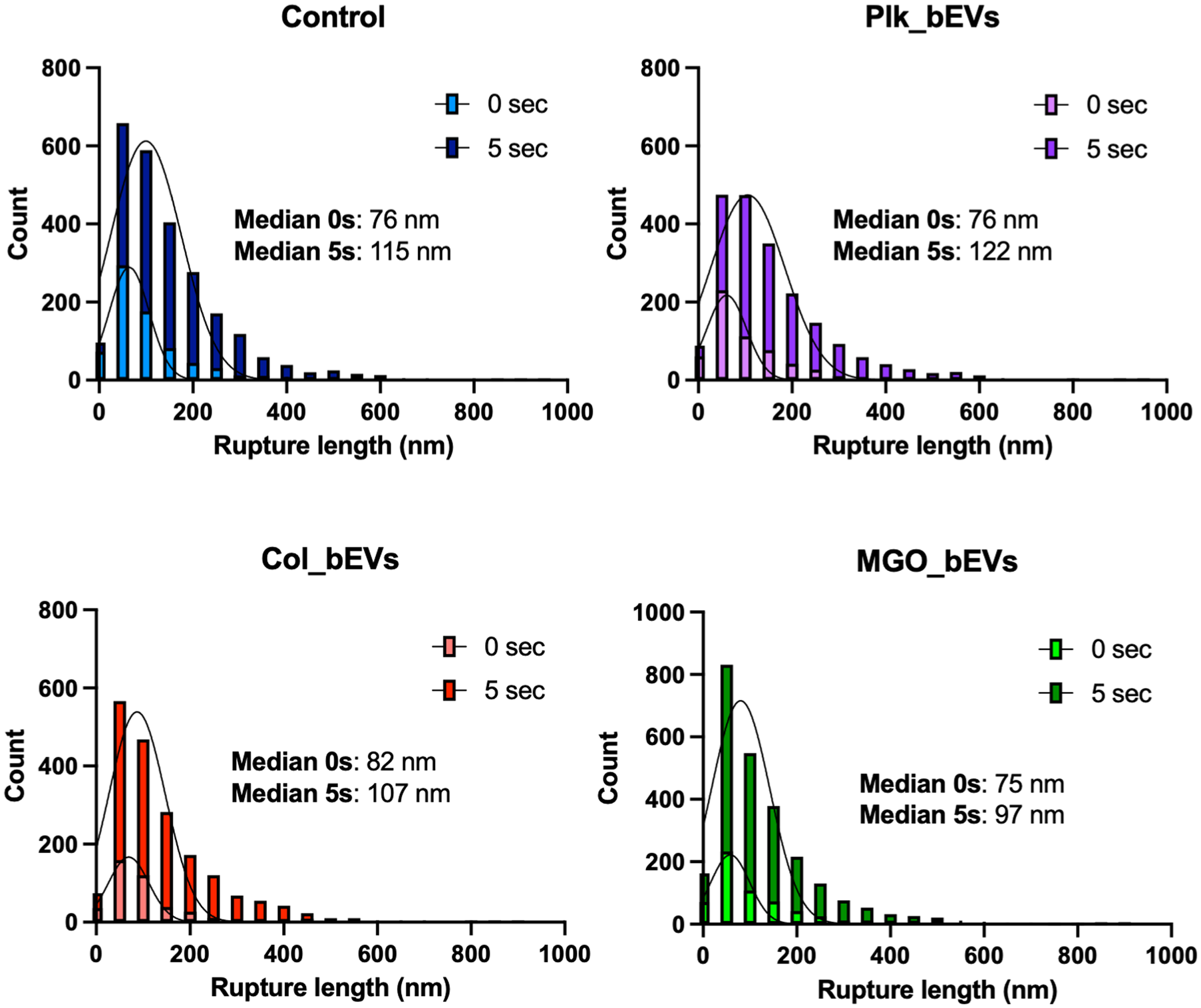
Rupture lengths for living Streptococcus sanguinis single-cell unbinding events from type I collagen surfaces after exposure to Streptococcus mutans bacterial extracellular vesicles (bEVs). Histograms for single-unbinding rupture lengths, for all force curves obtained with 0-s and 5-s dwelling times.
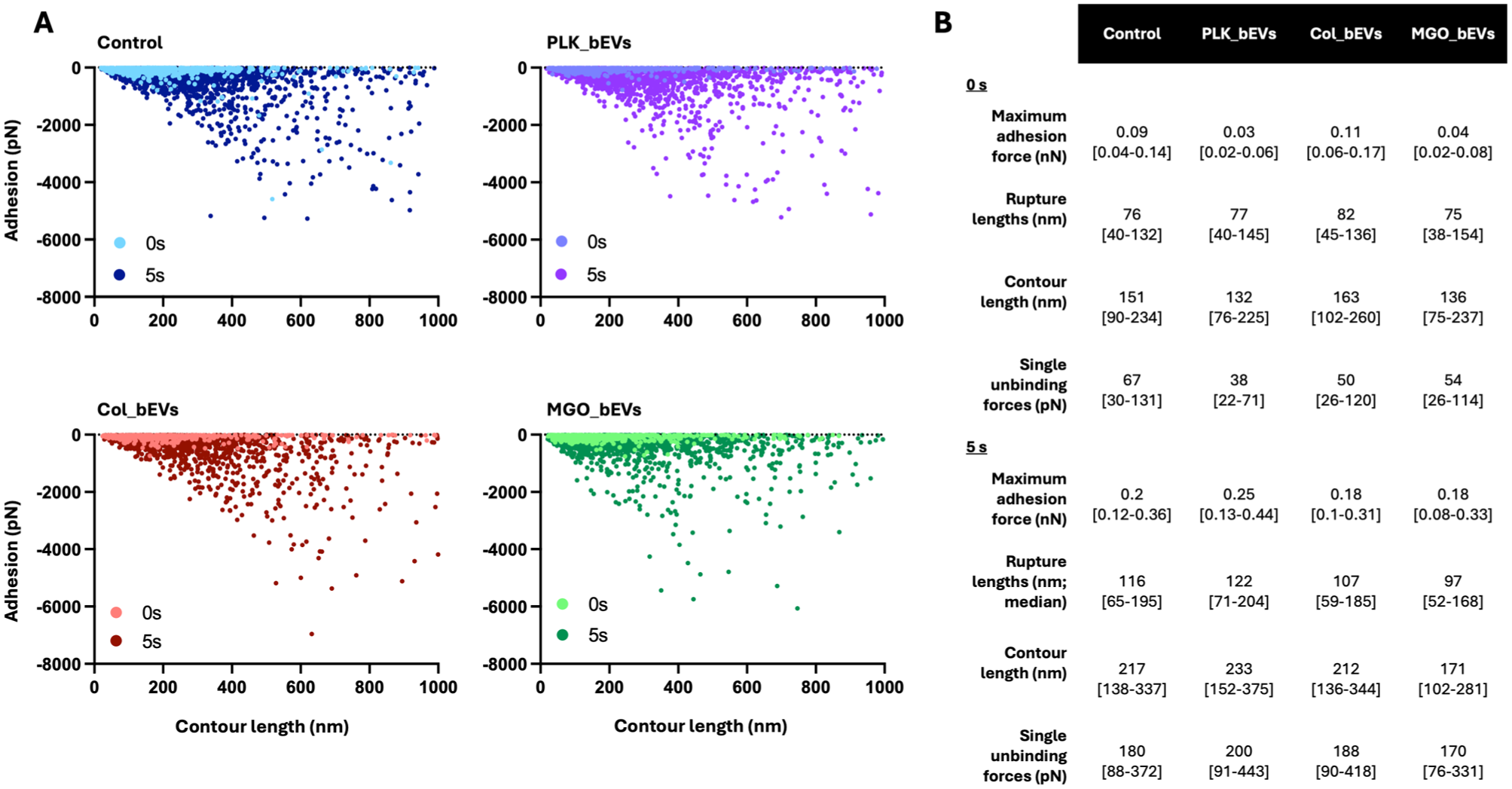
Streptococcus mutans–derived bacterial extracellular vesicles (bEVs) affect early molecular interactions between Streptococcus sanguinis and biological surfaces. (
S. mutans bEVs from Biofilms Reduce S. sanguinis Initial Biofilm Formation
Following SCFS analysis, the impact of S. mutans–derived bEVs on early S. sanguinis biofilm was evaluated using OCT (Fig 5). Live biofilms grown on hydroxyapatite (HAP) substrates were imaged, revealing 2 distinct structural regions: a high-density biofilm region adjacent to the substrate and a low-density region toward the biofilm surface (Fig 5A). The thickness of the high-density biofilm region was increased after incubation with PLK_bEVs from S. mutans. In contrast, the opposite effect was seen for bEVs from the biofilm conditions (Fig 5B and C). No significant differences were observed in the thickness of the low-density biofilm region across groups. However, exposure to MGO_bEVs resulted in an overall reduction in biofilm thickness compared with the PLK_bEVs group (Fig 5C).
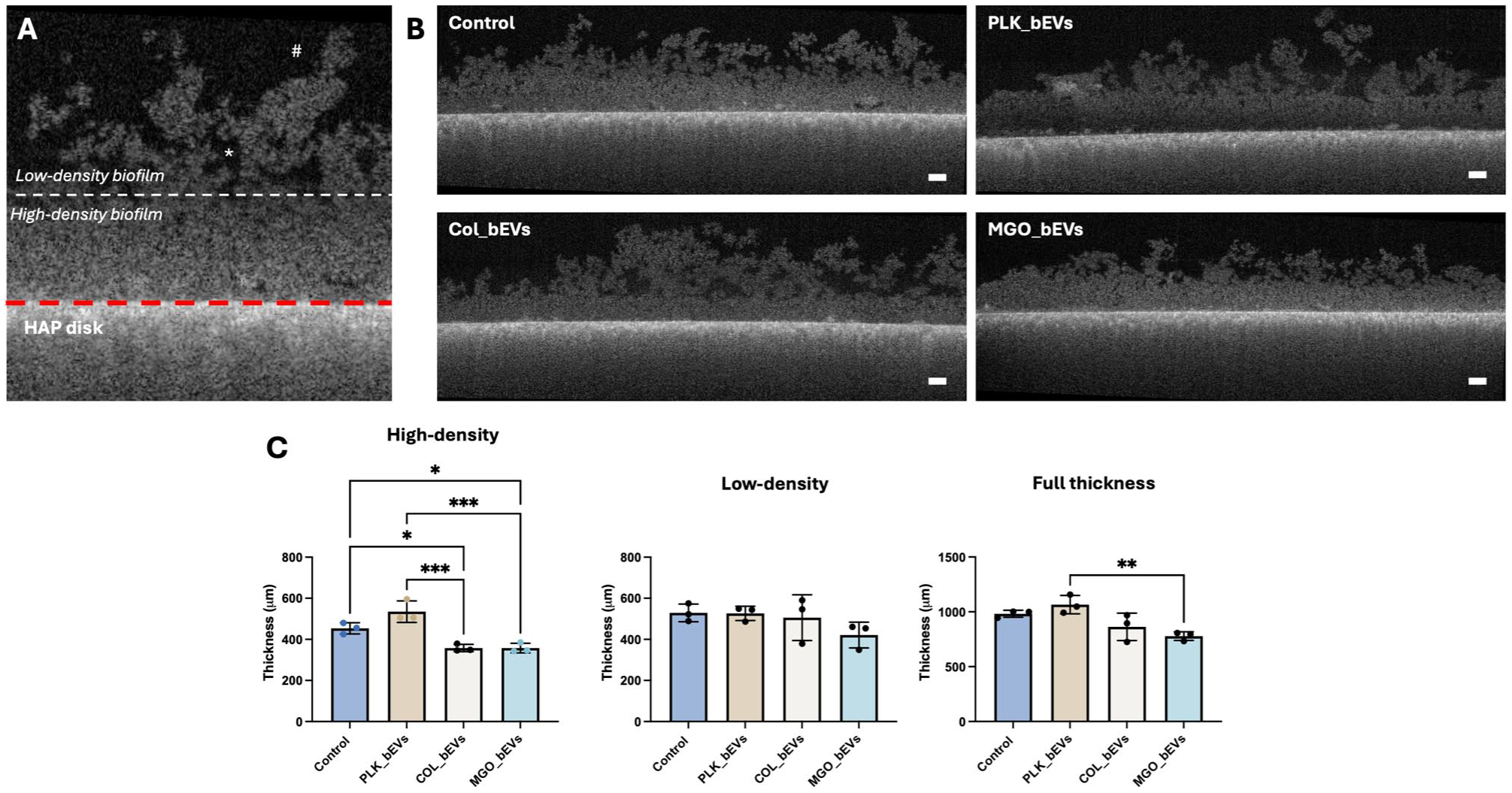
Streptococcus mutans–derived bacterial extracellular vesicles (bEVs) modulate Streptococcus sanguinis biofilm formation. (
Discussion
During dental biofilm formation, early microbial colonizers such as oral streptococci actively compete for substrate attachment within the local ecological niche (Hibbing et al 2010; Rostami et al 2022). In this context, several well-characterized dysbiotic factors have been attributed to S. mutans, including glycosyltransferase (Gtf) activity, modulation of extracellular polymeric substance (EPS), expression of collagen-binding proteins, and acidification of the surrounding environment (Kim et al 2020; Álvarez et al 2021; Jakubovics et al 2021). Our findings suggest that, in addition to these mechanisms, S. mutans–derived bEVs also contribute to early-colonizer dysbiosis by modulating the single-cell attachment and early biofilm formation of S. sanguinis. The influence of bEVs spans multiple biological scales from nanoscale modulation of individual adhesin–surface interactions to microscale effects on cellular attachment and early biofilm growth and establishment onto HAP substrates (Figs 2 and 5). Therefore, these results underscore the critical role of interspecies communication in the initial stages of biofilm development on dentin (Muras et al 2022) and highlight how bEVs may shape early streptococcal dysbiosis between S. mutans and S. sanguinis (Schuh et al 2019; Huang et al 2023).
Our SCFS findings reinforce that increased contact time between S. sanguinis and the surface enhances adhesion forces via bond strengthening, aligning with previous reports in the literature (Boks et al 2008; Huang et al 2015; Olivares et al 2025). Specifically, a 5-s contact time facilitated a greater number of adhesin–surface interactions and strengthened these bonds through time-dependent mechanisms (Carniello et al 2018). This enhanced binding was reflected in the longer rupture lengths observed at 5 s, indicating more stable and robust cell–surface interactions compared with the 0-s contact time (Fig 3).
Most notably, our results indicate that the effect of S. mutans–derived bEVs on S. sanguinis adhesion and biofilm formation onto collagen and HAP is highly dependent on the physiological state of the vesicle-producing cells (Leiva-Sabadini et al 2025). Particularly, PLK_bEVs promoted the attachment of S. sanguinis, whereas vesicles harvested from collagen-bound biofilms inhibited both S. sanguinis adhesion and early biofilm formation (Figs 2 and 4). This effect was mostly noticeable at 5-s contact times, although MGO_bEVs also showed significant adhesion inhibition at 0-s contact times. Overall, this biphasic behavior suggests a dynamic regulatory mechanism: during planktonic phases, S. mutans bEVs may promote the establishment of an initial streptococcal biofilm to facilitate attachment and survival. However, once S. mutans has successfully colonized the surface, its bEVs appear to switch toward suppressing the adhesion and biofilm formation of competing species such as S. sanguinis. These findings are similar to earlier reports showing that planktonic S. mutans bEVs are enriched in biofilm-promoting Gtfs, while biofilm-derived bEVs carry cargo associated with S. mutans–specific survival factors such as thiol peroxidase, an enzyme capable of neutralizing S. sanguinis–secreted hydrogen peroxide, a key mediator of their antagonistic interaction (Kreth et al 2005; Ruxin et al 2021; Leiva-Sabadini et al 2025). These cargo-based variations also correlate with microscopical and size changes (Leiva-Sabadini et al 2025) and may hint toward mechanistic differences across the different S. mutans bEV profiles, such as altering the way they interact with surfaces and modulate their effect on target cells.
Our results are also consistent with earlier findings that microbial environmental changes can modulate bEV production, composition, and cargo. For example, planktonic S. mutans bEVs—harvested under similar parameters—can promote biofilm formation in other competitors, including Actinomyces spp. (Senpuku et al 2019). A similar effect has been recently reported for Pseudomonas aeruginosa bEVs, which exhibit either biofilm-promoting or biofilm-inhibiting properties depending on the bacterial growth phase at the time of vesicle harvest (Saad et al 2024). Although collagen glycation modulated bEV sizes and some of the SCFS early single-bacterium adhesion parameters (Fig 2), no significant differences in OCT biofilm thickness were found compared with Col_bEVs (Fig 5). Therefore, it remains possible that glycation also plays a role as a modulating factor for bEV-based communication by S. mutans, similar to what has been observed with other environmental factors such as the shift from planktonic to attached or extracellular DNA secretion, as demonstrated in previous reports (Xie et al 2020; Leiva-Sabadini et al 2023).
Within the biofilm environment, the modulation of S. sanguinis early attachment by S. mutans–derived bEVs carries several important biological and clinical implications. First, it is well established that tooth demineralization occurs primarily at the biofilm–tooth interface, particularly in regions densely colonized by S. mutans microcolonies (Kim et al 2020). In such regions, bEV production may make the recolonization by S. sanguinis difficult, sustaining local acidification and promoting the progression of caries lesions. Second, AFM experiments confirm that the adhesion-inhibiting effect of S. mutans bEVs affects the initial cellular and subcellular S. mutans attachment mechanisms (Schuh et al 2021). However, the potential influence of S. mutans bEVs on other key virulence factors such as EPS production, biofilm nanomechanical stability, and bacterial viability remains to be elucidated (Cugini et al 2019; Gloag et al 2020).
Despite significant differences observed between PLK_bEVs and biofilm-derived bEVs, fewer distinctions were noted between bEVs isolated from biofilms grown on glycated collagen versus control collagen substrates (Figs 2 and 5). Previous work has suggested that collagen glycation can influence bacterial attachment and biofilm formation of oral bacteria to collagen substrates (Leiva-Sabadini et al 2023). Future studies should be carried out to unravel the potential mechanisms behind these observations. Similar modulatory effects have been described for bEVs derived from P. gingivalis and various gut microbes (Zhang et al 2021; Díaz-Ubilla et al 2025). These findings further support the emerging view that bEVs are critical mediators of microbial communication and pathogenesis across different ecosystems and should be further explored in the context of oral disease progression.
Finally, this work demonstrates that AFM-based SCFS is a powerful technique for quantifying the impact of bEVs on cross-species adhesion in the context of microbial dysbiosis (El-Kirat-Chatel and Beaussart 2018). This experimental pipeline can potentially allow for the study of bEV effects on intraspecies, interspecies, and interkingdom communication, particularly in relation to changes in bacterial adhesion and nanomechanical properties following bEV exposure, under controlled biological and biochemical conditions (Cho et al 2023). This work also highlights that the functional impact of bEVs is modulated by the physiological state of the donor bacteria—whether planktonic or surface attached—underscoring the need for further mechanistic studies to elucidate this change.
While this SCFS experimental approach provides novel insights, certain methodological limitations remain, including the use of limited bacterial strains and difficulties replicating the complex, in vivo oral environment (i.e., salivary components, multispecies interactions) in an in vitro setting. Also, bEVs from S. mutans were isolated from aerobic conditions, and further investigations should confirm if these findings also hold true for microaerophilic and anaerobic environments. These challenges highlight important directions for future research, including the integration of AFM with microfluidic flow systems, real-time multiscale microscopical imaging, and the use of broader bacterial communities to deepen our understanding of bacterial interactions during the initial phases of oral biofilm formation. Moreover, studies should continue to explore how bEVs modulate the crosstalk among key members of the oral biofilm beyond S. mutans–S. sanguinis interactions, particularly within mature, established multispecies biofilms associated with dental caries and periodontal disease.
Conclusion
The effect of S. mutans bEVs on the early adhesion of S. sanguinis to substrates varies as a function of their isolation either from planktonic or biofilm states on nonglycated and glycated collagen. Specifically, bEVs derived from planktonic S. mutans promote the adhesion of S. sanguinis to collagen substrates and support early biofilm formation, whereas bEVs isolated from collagen-attached biofilms exert the opposite effect, inhibiting adhesion and biofilm initiation. To our knowledge, this is the first study to provide direct evidence that bEVs are critical mediators of interspecies communication between S. mutans and S. sanguinis and may play a pivotal role in modulating commensal-pathogen dynamics in the context of dental caries.
Author Contributions
C. Leiva-Sabadini, contributed to data acquisition, analysis, and interpretation, drafted the manuscript; C.M. Lévesque, L. Bozec, C. Schuh, contributed to conception and design, data analysis and interpretation, critically revised the manuscript; S. Aguayo, contributed to conception and design, data analysis and interpretation, drafted the manuscript. All authors gave their final approval and agree to be accountable for all aspects of the work
Supplemental Material
sj-docx-1-jdr-10.1177_00220345261417246 – Supplemental material for Streptococcus mutans Extracellular Vesicles Regulate Early Streptococcus sanguinis Nanoadhesion
Supplemental material, sj-docx-1-jdr-10.1177_00220345261417246 for Streptococcus mutans Extracellular Vesicles Regulate Early Streptococcus sanguinis Nanoadhesion by C. Leiva-Sabadini, C.M. Lévesque, L. Bozec, C. Schuh and S. Aguayo in Journal of Dental Research
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the ANID FONDECYT grants (Nos. 1220804 and 1220803). In addition, C.L.-S. is supported by the Beca ANID PhD scholarship (No. 21220799). This work was also supported by the Natural Sciences and Engineering Research Council of Canada (NSERC) grants RGPIN-2021-02694 (to L.B.) and RGPIN-2019-06454 (to C.M.L.); the Ontario Research Fund (ORF) and Canadian Fund for Innovation project No. 42809; and finally, the Faculty of Dentistry at the University of Toronto for supporting the research. Acknowledgment is also extended to the CAMiLoD Imaging Facility at the Faculty of Dentistry for providing access to their imaging facilities and to Mehrnoosh Neshatian for her support with the contact angle measurements. ![]() was partially created in Biorender.com.
was partially created in Biorender.com.
A supplemental appendix to this article is available online.