
Abstract
Periodontitis, caused by periodontal pathogens such as Porphyromonas gingivalis, is a risk factor for Alzheimer’s disease (AD) progression. Neutrophils, which are abundant in patients with periodontitis, release neutrophil extracellular traps (NETs) to resist microbial infection. We explored the mechanism and effects of P. gingivalis–induced NETs on the relationship between periodontitis and neuroinflammation. A murine periodontitis model was established via oral local application of P. gingivalis with or without tak242 (TLR4 inhibitor). Maxillary bones were evaluated via micro–computed tomography. The proportion of neutrophils was determined by flow cytometry. NET formation and morphology were analyzed via a cell-free DNA kit, a neutrophil elastase enzyme-linked immunosorbent assay (ELISA) and myeloperoxidase ELISA kit, reverse transcription polymerase chain reaction (RT-PCR), western blotting and immunofluorescence. Behavior tests were used to investigate cognitive ability. Neuroinflammation was assayed by immunohistochemistry (IHC) and RT-PCR. Amyloid precursor protein (APP) processing was measured by IHC. In vitro experiments explored the functional mechanism underlying the effects of P. gingivalis–induced NETs on the neuron–glia unit. We observed significant alveolar bone resorption with elevated neutrophil count and increased NET formation in mice with periodontitis. Cognitive abilities were impaired by periodontitis. Neuroinflammation manifested as glia activation and upregulated inflammatory cytokines, and APP processing was altered by the elevated expression of APP and PSEN1. These changes were specifically reversed by tak242. In vitro, P. gingivalis–induced NETs mediated M1 polarization in BV2 cells and changed APP processing in N2a cells, along with the activation of TLR4/Myd88/NF-κB and GSK3β/Akt, which is consistent with the in vivo findings. In conclusion, P. gingivalis–induced NETs play pivotal roles in the relationship between neuroinflammation and cognitive impairment. Furthermore, the effects of P. gingivalis–induced NETs on neuron–glia unit were related to TLR4 activation.
Keywords
Introduction
Periodontitis is a common, chronic oral infectious condition that has a potential bidirectional relationship with the development of Alzheimer’s disease (AD) (Teles et al 2022). Periodontal pathogens have been studied as important actors in recent years (Akiyama et al 2000). Porphyromonas gingivalis, the most significant periodontal pathogen, has been observed in the brains of patients with AD and AD animal models (Sansores-Espana et al 2021; Ciccotosto et al 2024). P. gingivalis lipopolysaccharide (LPS) significantly contributes to the relationship between AD and periodontitis (Hu et al 2020). P. gingivalis DNA and related proteins were detected in the gingiva and brains of mice with periodontitis in our previous studies (Shen et al 2024; Jiang et al 2025). Moreover, its gingipains affect the integrity of the blood–brain barrier (BBB) (Dominy et al 2019). Because inflammation is a key pathological driver of AD development (Dioguardi et al 2020), chronic P. gingivalis infection has increased proinflammatory cytokine levels in peripheral tissues and the circulatory system, decreased integrity of the BBB, and activated inflammation in the brain (Pritchard et al 2022). However, the detailed mechanisms underlying inflammation-associated processes from the periphery to the center between periodontitis and AD require further exploration.
Neutrophils are the first host defensive cells to face bacterial invasion and infection (Bui et al 2019; Sangineto et al 2023). Neutrophil infiltration in peripheral tissues (e.g., the gingiva and blood) is well known to occur during periodontitis. In different pathological conditions, including bacterial infection, trauma, and neurodegeneration, neutrophils infiltrate the central nervous system (CNS) when the BBB is disrupted. Neutrophils can translocate from the blood to the brain for neutrophil infiltration, which reveals the existence of communication between the brain and peripheral immune system (Chou et al 2023). These findings indicate that neutrophils play important roles in the relationship between the periphery and the CNS. Neutrophils employ neutrophil extracellular trap (NET) formation, degranulation, and reactive oxygen species (ROS) to eliminate invading pathogens and maintain homeostasis and defense (Zenaro et al 2015, Pietronigro et al 2023). NETs, composed of a complex of chromatin (DNA) and polymorphonuclear proteins, including histones, myeloperoxidase (MPO), and neutrophil elastase (NE), are early triggers of periodontal inflammation (Kim et al 2023). The activation of Toll-like receptor 4 (TLR4) has been shown to increase hypercitrullination of histones, a posttranslational modification that initiates NET formation ex vivo (Wang et al 2023). NETs drive inflammatory responses in the skin through TLR4 activation (Shao et al 2019). NETs can potentially promote intestinal epithelial necroptosis via the regulation of TLR4-mediated mitophagy (Chu et al 2024). NETs disrupt the balance between osteogenesis and osteoclastogenesis via the TLR4 pathway in patients with arthritis (Li et al 2025). More importantly, TLR4 was identified as a hub gene, elucidating the link between periodontitis and AD in the Disease Gene Network database (Ge et al 2024). However, the potential mechanisms through which P. gingivalis–induced NETs affect the neuron–glia unit via the TLR4 signaling pathway are still unclear.
We proposed to explore the role of NETs as a mediator between periodontitis and neuroinflammation through TLR4 activation. We established a mouse model of periodontitis in which mice were pretreated with a TLR4 inhibitor (tak242). First, the neutrophil percentage and NET formation in peripheral tissues and brains were measured. Second, cognitive function was evaluated via behavioral tests. Neuroinflammation and pathological alterations in the brain were confirmed. Finally, we investigated whether the underlying mechanisms were related to TLR4 activation.
Methods
The details are described in the Appendix Methods. The Ethical Committee of Animal Care and Experimental Committee of the Ninth People’s Hospital affiliated with the Shanghai Jiao Tong University School of Medicine (SH9H-2025-A13-1) gave approval of all experimental protocols. This study conformed to the updated ARRIVE (ARRIVE 2.0 guidelines) 2.0 guidelines.
Results
Assessment of Alveolar Bone Loss and TLR4 Expression
P. gingivalis contributed to alveolar bone destruction on the buccal and palatal sides in the periodontitis (PD) group, as shown in Figure 1A. The root furcation was specifically exposed, and alveolar bone crest–cementoenamel junction (ABC-CEJ) increased in the PD group (232.9 ± 25.29 μm) compared with that in the control (CON) group (149.5 ± 16.69 μm, P = 0.0001) (Fig. 1B). In addition, the bone volume (BV)/total volume (TV) was decreased in the PD group (65.69 ± 2.806%) compared with that in the CON group (74.04 ± 2.058%, P = 0.0006), as shown in Figure 1C. As shown in Figure 1D, the bone surface (BS)/BV ratio increased in the PD group (82.49 ± 6.660/mm) compared with that in the CON group (63.53 ± 9.752/mm, P = 0.003). Experimental periodontitis was successfully induced in mice by P. gingivalis. Moreover, these changes were alleviated in the PD + tak group (ABC-CEJ: 152.2 ± 23.27 μm, BV/TV: 72.99% ± 2.443%, BS/BV: 68.57 ± 3.26/mm), suggesting that tak242 administration reduced alveolar bone resorption caused by P. gingivalis.
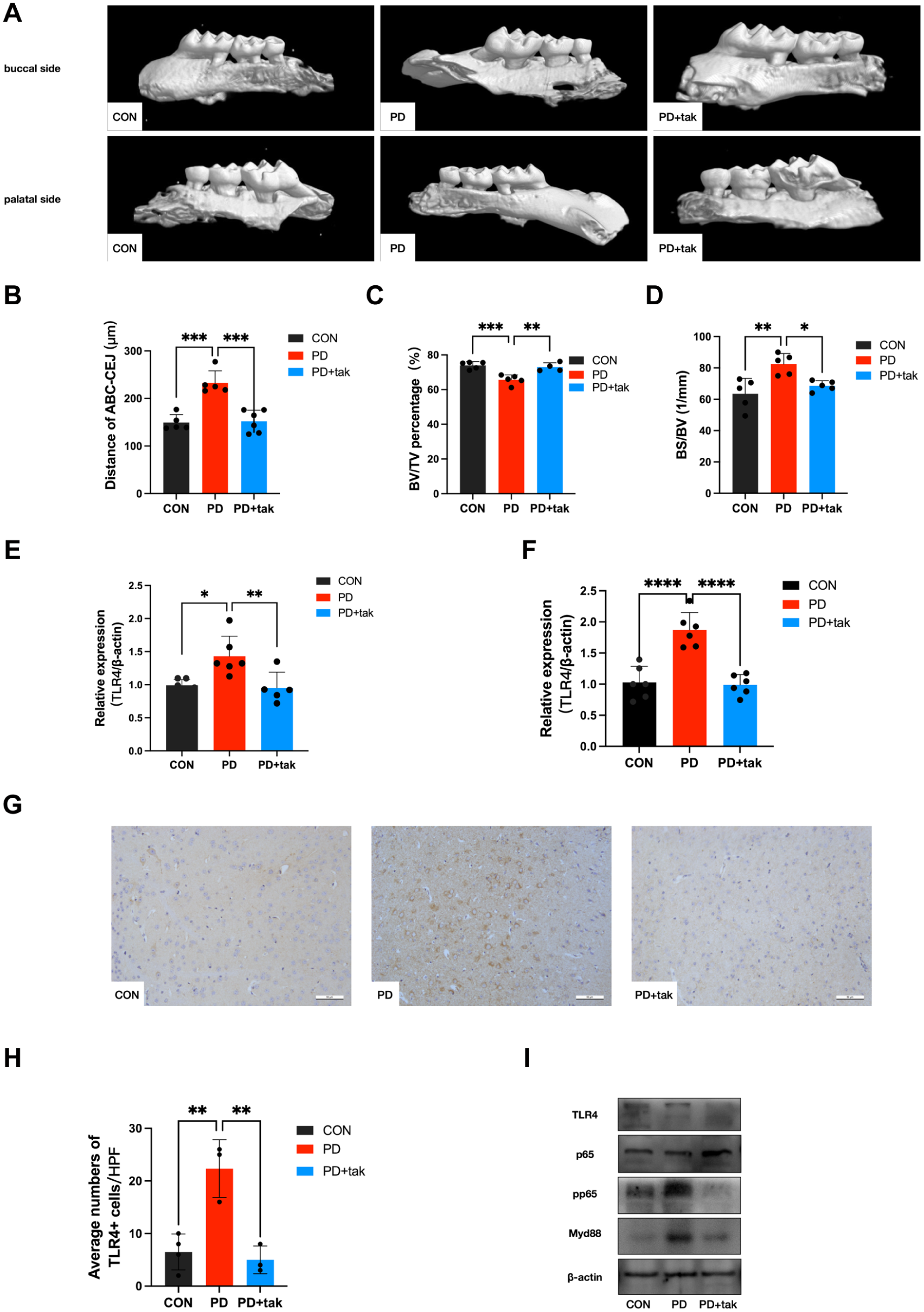
Assessment of alveolar bone loss and Toll-like receptor 4 (TLR4) expression. (
In both peripheral blood mononuclear cells (PBMCs) and the cortex, TLR4 messenger RNA (mRNA) expression was significantly increased in the PD group (PBMCs: P = 0.012; cortex: P < 0.0001) (Fig. 1E, F). There were no significant differences between the CON group and the PD + tak group (PBMCs: P = 0.9538; cortex: P = 0.9589). Cortical immunohistochemistry (IHC) staining revealed an obvious increase in the number of TLR4+ cells in the PD group compared with the CON group (22.33 ± 5.508 vs. 6.5 ± 3.416/high-power field [HPF]) (Fig. 1G, H). In the PD + tak group, the average number of TLR4+ cells significantly decreased. Furthermore, we measured protein expression related to the TLR4/Myd88/NF-κB signaling pathway in the mouse cortex. Increased TLR4, Myd88, and pp65 protein expression was detected in the PD group, and this was reduced by tak242 administration in the PD + tak group (Fig. 1I).
Detection of Neutrophils in the Periphery and Brain
The presence of neutrophils was confirmed by the presence of CD45+CD11b+Ly6G+ cells (Fig. 2A). Compared with that in the CON group (4.56% ± 1.284%), the proportion of neutrophils in the gingiva clearly increased in the PD group (8.512% ± 0.6929%, P = 0.0004), as shown in Figure 2B. Next, we analyzed the presence of neutrophils in the blood and brain to measure neutrophil accumulation. The number of neutrophils in the blood and brain was significantly greater in the PD group than in the CON group (11.97% ± 1.894% vs. 6.457% ± 0.2479%, P = 0.01333; 3.620% ± 0.3492% vs. 1.595% ± 0.2932%, P < 0.0001) (Fig. 2C, D). In addition, compared with the PD group, the increase in the number of neutrophils decreased after the addition of tak242 in the PD + tak group (gingiva: 2.863% ± 0.3086%, P < 0.0001; blood: 5.770% ± 2.495%, P = 0.0073; brain: 1.615% ± 0.5167%, P < 0.0001).
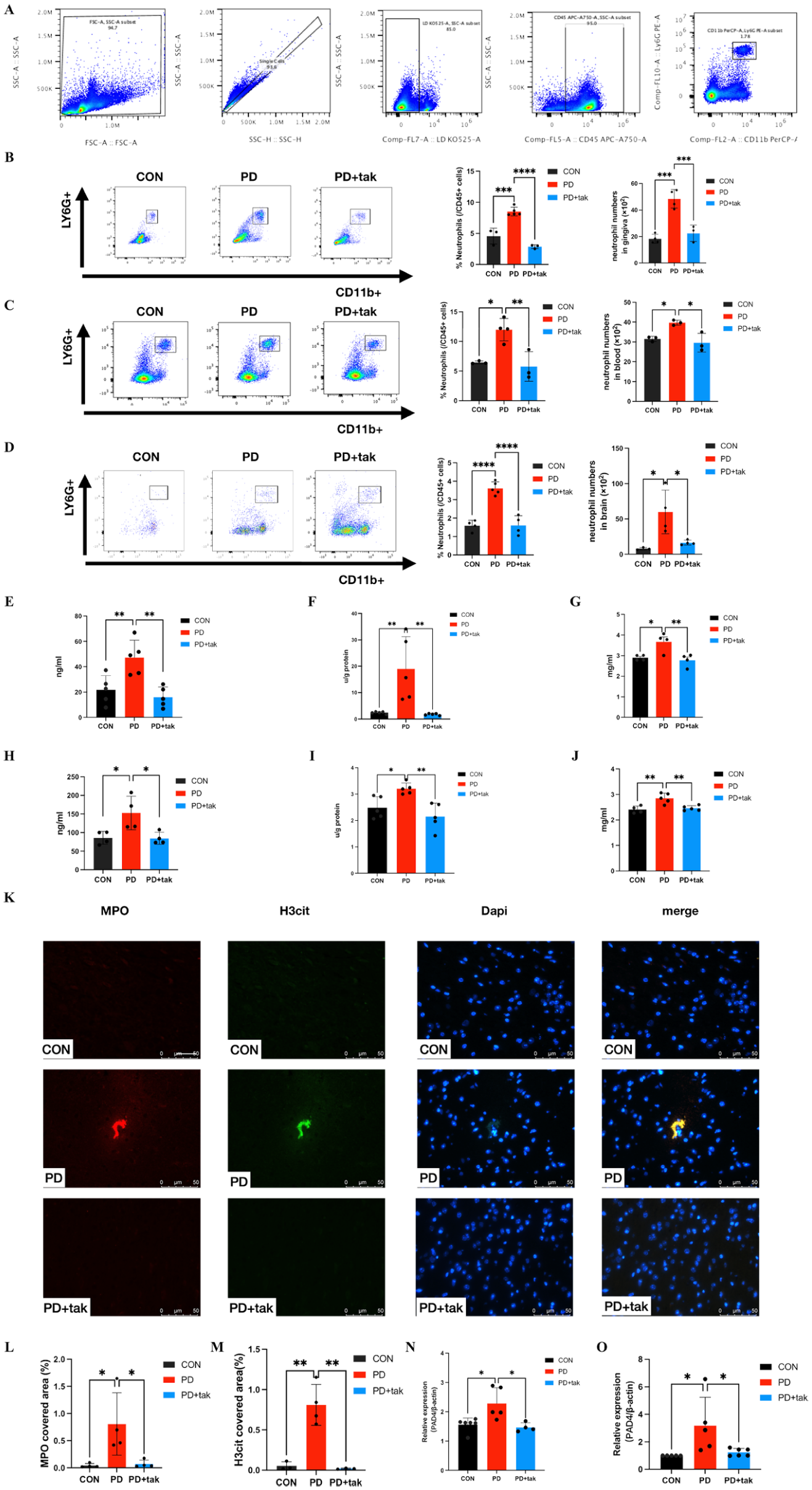
Proportion of neutrophils and neutrophil extracellular trap (NET) formation. (
NET Formation in the Periphery and Brain
NET formation in the periphery and brain was confirmed in mouse plasma and cortical tissues. Compared with the CON group (21.75 ± 11.28 ng/mL), the PD group exhibited higher cell-free DNA (cf-DNA) concentrations (47.28 ± 13.68 ng/mL, P = 0.0095) in plasma (Fig. 2E). MPO expression in plasma was markedly greater in the PD group (19.00 ± 12.16 u/g protein) than in the CON group (2.422 ± 0.3979 u/g protein; P = 0.0075) (Fig. 2F). NE levels in plasma were also elevated in the PD group (3.674 ± 0.4599 mg/mL) compared with those in the CON group (2.901 ± 0.1275 mg/mL, P = 0.0212) (Fig. 2G). After tak242 treatment, the concentrations of cf-DNA, MPO, and NE in plasma decreased to 16.10 ± 7.966 ng/mL, 1.767 ± 0.35 u/g protein, and 2.78 ± 0.3048 mg/mL, respectively, in the PD + tak group. Moreover, compared with those in the CON group, cf-DNA, MPO, and NE levels in the cortex seemed to increase in the PD group (cf-DNA concentration: 152.9 ± 45 ng/mL vs. 86.03 ± 17.24 ng/mL; P = 0.0257; MPO concentration: 3.206 ± 0.2126 u/g protein vs. 2.488 ± 0.4137 u/g protein; P = 0.0355; NE concentration: 2.854 ± 0.1993 mg/mL vs. 2.409 ± 0.1433 mg/mL; P = 0.0031) (Fig. 2H–J). The addition of tak242 led to a significant decrease in cf-DNA, MPO, and NE levels in the brain in the PD + tak group (cf-DNA concentration: 84.75 ± 16.24 ng/mL; MPO concentration: 2.153 ± 0.5055 u/g protein; NE concentration: 2.461 ± 0.09598 mg/mL).
Using fluorescence microscopy, we observed that NET-like structures labeled for MPO and H3cit expression significantly increased in the mouse cortex of the PD group compared with those in the CON group (Fig. 2K). There were clearly more MPO- and H3cit-expressing areas in the PD group than in the CON group (Fig. 2L, M). In the PD + tak group, MPO and H3cit expression clearly decreased than that in the PD group.
Simultaneously, there was significantly higher Pad4 mRNA expression (Fig. 2N) in PD group PBMCs compared to those in the CON group, which decreased following tak242 administration in the PD + tak group. Similar results for the mRNA expression levels of Pad4 (Fig. 2O) were observed in the cortex.
Evaluation of Learning and Memory
The timeline of the experiment is shown in Figure 3A. Open field test (OFT) was used to assess spontaneous activity in the mice in the 3 groups. The total distance covered (Fig. 3B) and average speed (Fig. 3C) did not differ significantly across the 3 groups. These findings indicate that the locomotor activity of the mice was not influenced by P. gingivalis or tak242 administration.
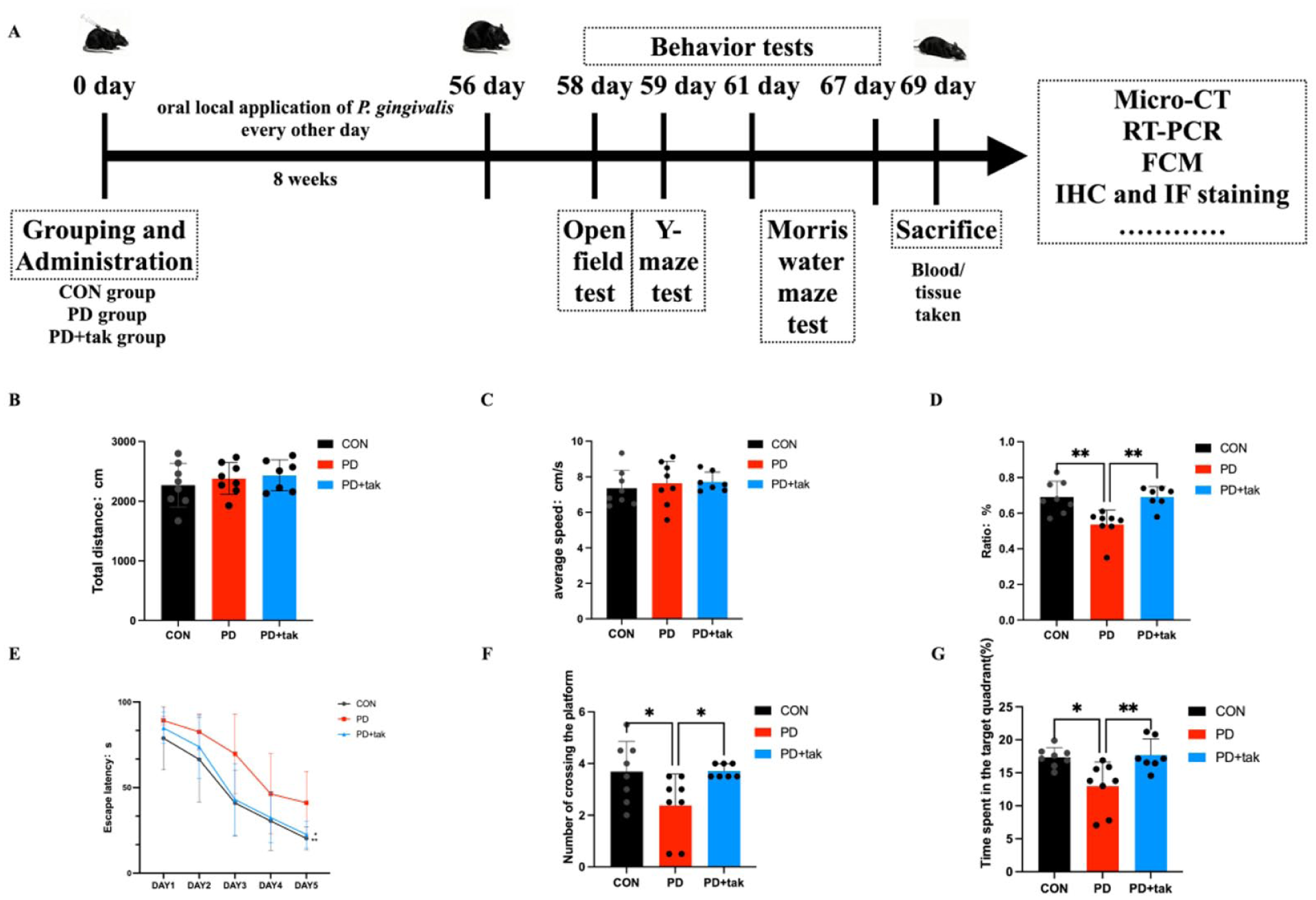
Effects on learning and memory. (
In the Y-maze test, the alternation ratio significantly decreased in the PD group (0.5370 ± 0.08020) compared with that in the CON group (0.6909 ± 0.08914, P = 0.0021), as shown in Figure 3D. In the PD + tak group, the alternation ratio returned to 0.6911 ± 0.05797.
The mice in 3 groups were trained for 5 d, and all groups experienced a decrease in latency during the 5-d training period (Fig. 3E). On day 5 of training, the escape latency of the PD group (39.75 ± 14.07 s) was significantly longer than that of the CON group (20.31 ± 6.748 s, P = 0.0471). There was no significant difference between the CON group and the PD + tak242 group (22.57 ± 7.79 s, P = 0.8631). During the probe test, mice in the PD group performed fewer platform crossings (Fig. 3F) and spent less time in the target quadrant (Fig. 3G) than those in the CON group (2.375 ± 1.217/times vs. 3.688 ± 1.163/times, P = 0.427; 12.97 ± 3.661 s vs. 17.36 ± 1.458 s, P = 0.0102). The changes observed in the behavior tests were alleviated by tak242 administration in the PD + tak group compared with the PD group (number of platform crossings: 3.714 ± 0.2373/times, P = 0.0462; time in the target quadrant: 17.36 ± 2.435 s, P = 0.0075).
Neuroinflammation and Amyloid Precursor Protein Processing
The expression of IBA1 was upregulated in activated microglia. The number of IBA1+ cells was significantly greater in the PD group than in the CON group (22 ± 3.606 vs. 13 ± 2, P = 0.0396) (Fig. 4A). We used GFAP staining to label activated astrocytes. Compared with that in the CON group (36.25 ± 4.992), the number of GFAP+ cells in the hippocampus was greater in the PD group (57.33 ± 2.517, P = 0.0027) (Fig. 4B). In the PD + tak group, the numbers of IBA1+ and GFAP+ cells decreased significantly (10.67 ± 4.163 and 33.75 ± 7.228, respectively).
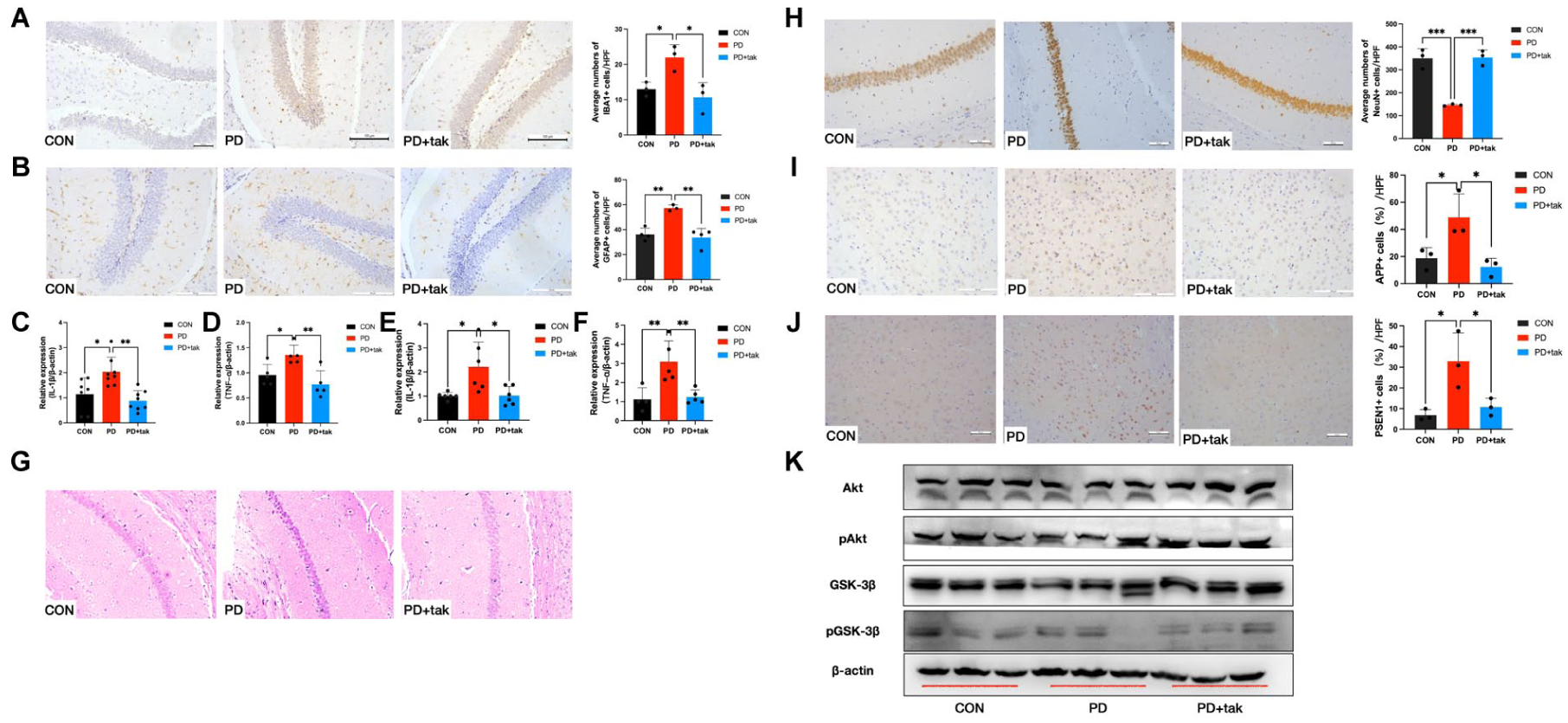
Neuroinflammation and amyloid precursor protein (APP) processing in mice. (
The mRNA expression levels of IL-1β (2-fold increase) (Fig. 4C) and TNF-α (1.3-fold increase) (Fig. 4D) in PBMCs were significantly higher in the PD group than in the CON group, and these levels decreased in the PD + tak group, indicating that P. gingivalis-induced inflammation in the blood was attenuated by tak242 treatment. In the cortex, similar results were observed for IL-1β (2-fold increase) (Fig. 4E) and TNF-α (3-fold increase) (Fig. 4F) mRNA expression levels in the PD group compared with the CON group. Increased IL-1β and TNF-α expression was reversed by tak242 administration in the brains of mice in the PD + tak group.
Hematoxylin and eosin (H&E) staining of the CA1 region revealed shrunken neurons, swollen cells, and a disordered cell arrangement in the PD group (Fig. 4G). Moreover, compared with those in the CON group, the number of surviving neurons in the hippocampus in the PD group markedly reduced (146.3 ± 4.509 vs. 350.7 ± 42.16, P = 0.0005) (Fig. 4H), suggesting exacerbated neuronal degeneration following P. gingivalis administration. In the PD + tak group, the number of neurons increased (354 ± 32.74).
As shown in Figure 4I, J, increased expression of amyloid precursor protein (APP) and presenilin 1 (PSEN1) protein, presented as brown cytoplasmic particles, were observed in the cortex of the PD group compared with the CON group (APP: 48.85 ± 17.26% vs. 18.72 ± 7.824%, P = 0.0428; PSEN1: 32.90 ± 13.72% vs. 6.796 ± 2.75%, P = 0.0213). These changes were reversed by tak242 administration in the PD + tak group (APP: 12.20 ± 6.418%; PSEN1: 10.77 ± 4.22%).
Compared with those in the CON group, the level of pGSK3βSer9 and pAkt in the mouse cortex were significantly lower in the PD group (Fig. 4K). Notably, these reductions were reversed by tak242 treatment in the PD + tak group.
NET Formation in P. gingivalis–Treated Neutrophils
P. gingivalis led to enhanced cf-DNA production. Compared with the con group (274.5 ± 131.1 ng/mL), and the P. gingivalis-stimulated (pg) + tak group (374.8 ± 11.42 ng/mL), cf-DNA concentrations were significantly increased in the P. gingivalis-stimulated (pg) group (623.9 ± 89.59 ng/mL, Fig. 5A) was observed. NE levels were also elevated in the pg group (6.770 ± 1.082 mg/mL) compared with those in the con group (3.310 ± 0.9545 mg/mL) (Fig. 5B) and decreased to 4.043 ± 0.1644 mg/mL in the pg + tak group. MPO expression was markedly greater in the pg group (8.511 ± 1.117 u/g protein) than in the con group (2.092 ± 0.8044 u/g protein) (Fig. 5C). After tak242 was administered, its expression clearly decreased (3.933 ± 1.703 u/g protein) in the pg + tak group. Immunofluorescence (IF) staining of H3cit and MPO, revealed that the increased NET formation in the pg group was attenuated by tak242 in the pg + tak group (Fig. 5D).
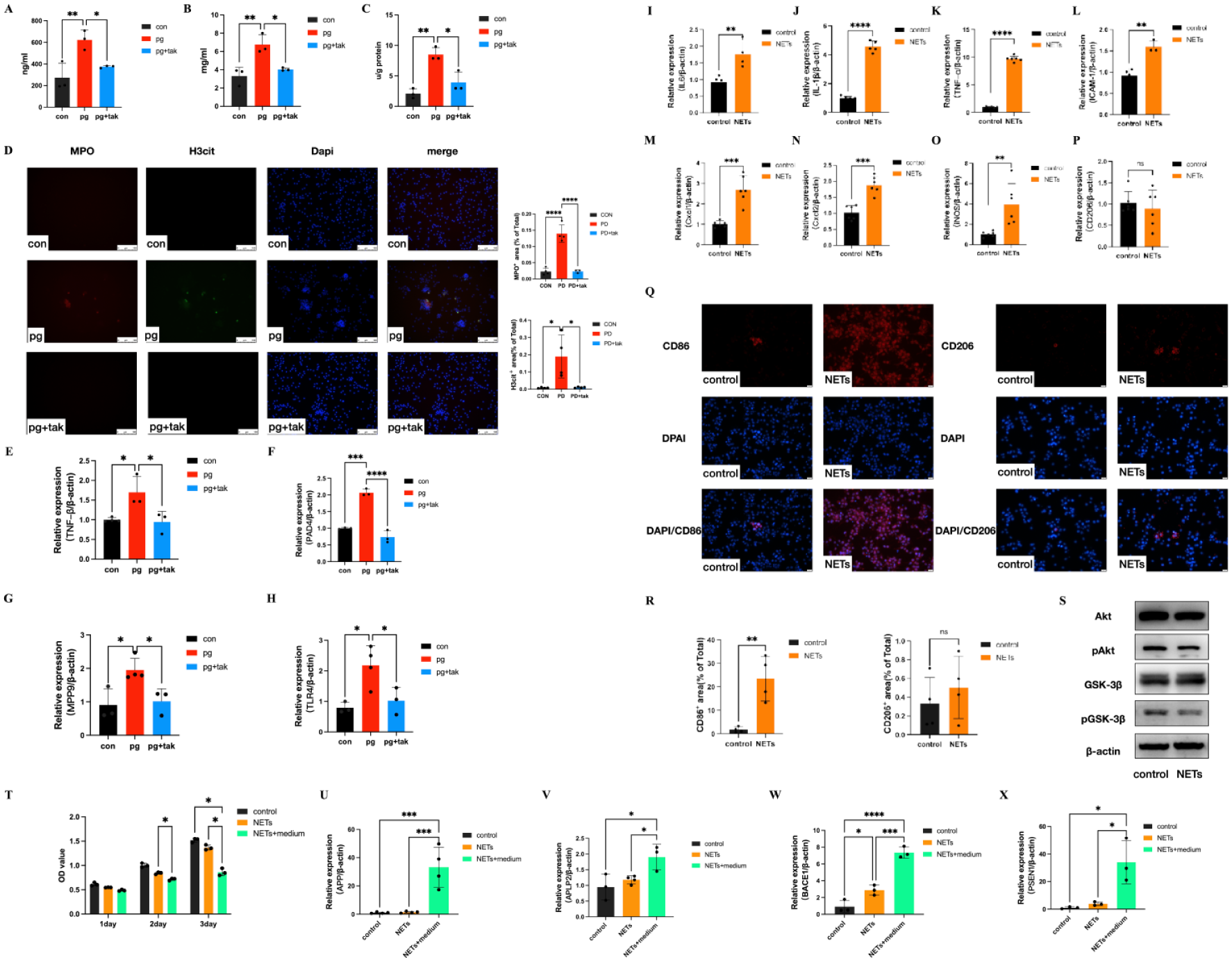
Neutrophil extracellular trap (NET) formation by neutrophils after Porphyromonas gingivalis stimulation and the effects of P. gingivalis–induced NETs on neuron–glia unit cells. (
Similar to the results of the in vivo studies, the pg group showed 1.5-fold, 2-fold, 2-fold, and 2-fold increases in the mRNA expression of TNF-α (Fig. 5E), PAD4 (Fig. 5F), MMP9 (Fig. 5G), and TLR4 (Fig. 5H), respectively, which were markedly restored by tak242 administration in the pg + tak group.
Effects of P. gingivalis–Induced NETs on the Neuron–Glia Unit
The mRNA expression levels of IL-6 (Fig. 5I), IL-1β (Fig. 5J), TNF-α (Fig. 5K), ICAM-1 (Fig. 5L), Cxcl2 (Fig. 5M), and Cxcl2 (Fig. 5N) increased 1.5-fold, 4-fold, 8-fold, 1.5-fold, 2-fold, and 1.5-fold, respectively, in BV2 cells with NETs treatment. Furthermore, the results revealed that NETs promoted M1 microglial polarization, which increased iNOS mRNA expression 4-fold after NETs treatment (Fig. 5O) and CD86 expression (Fig. 5Q, R). There was no significant difference in CD206 mRNA (Fig. 5P) or protein expression (Fig. 5Q, R) related to M2 polarization in BV2 microglia with NETs treatment. As shown in Figure 5S, NETs noticeably decreased pAkt and pGSK3βSer9 protein phosphorylation, which suggests that BV2 microglia were activated by NETs treatment via the GSK3β/Akt signaling axis.
In the N2a neuron cell assays, compared with that in the NETs group, the proliferation of N2a neurons in the NETs + medium group was significantly lower (2 day: P = 0.0439, 3 day: P = 0.0281) (Fig. 5T). Furthermore, compared with those in the NETs group, the mRNA expression of APP (Fig. 5U), APLP2 (Fig. 5V), BACE1 (Fig. 5W), and PSEN1 (Fig. 5X) in the NETs + medium group increased 15-fold (P = 0.001), 2-fold (P = 0.0489), 3-fold (P = 0.0005), and 10-fold (P = 0.0158), respectively.
Discussion
A widespread disease in elderly individuals, periodontitis, the major pathogen underlying which is P. gingivalis, is recognized as a high-risk factor for AD (Kwon et al 2021; Vanova et al 2023). Notably, this pathogen is thought to be a key mediator linking the 2 diseases (de Buhr and von Kockritz-Blickwede 2020). Once P. gingivalis infects the oral cavity, it may spread and access the brain, where it damages the BBB and recruits immune cells to defend against infection (Keller et al 2020). Among these recruited immune cells, neutrophils serve as the first line of core defenders, migrating to inflamed tissues to combat bacterial invasion (Gour et al 2024; Rosenzweig et al 2024). However, the mechanisms underlying the function of neutrophils in the context of periodontitis and AD remain poorly defined. Here, we demonstrate a causal link between NETs and neuroinflammation in a well-established model of periodontitis in vivo and in vitro via the TLR4 signaling pathway.
In this study, we successfully established periodontitis by oral administration of P. gingivalis for 8 wk. In preliminary experiments, we compared 4-wk and 8-wk periodontitis models. The Y-maze test showed no significant difference in cognitive function between the 4-wk periodontitis group and the control group, along with no difference in inflammatory cytokines in mice brains between the 4-wk periodontitis group and the control group (data not shown). The 8-wk periodontitis model induced cognitive impairment and neuroinflammation, which has been published in our previous study and other published studies (Aravindraja et al 2022; Lei et al 2023; Shen et al 2024).
These results indicate that P. gingivalis increased the frequency of neutrophil infiltration and NET formation both in the periphery and in the brain. Typically, neutrophils are scarce in the CNS under healthy conditions. However, P. gingivalis and its virulence factors can induce chronic inflammation and damage BBB function to improve the infiltration of the CNS by neutrophils. Neutrophils employ mechanisms involving phagocytosis, degranulation, ROS production, and NET formation (Li et al 2022). NET formation can damage the BBB and injure surrounding neurons. We hypothesized that NET formation in the periphery could migrate via the systemic circulation and thereby disrupt the BBB. Furthermore, it should be emphasized that cytotoxic proteins (including histones, NE, and defensins) and autoantigens (such as DNA, MPO, and proteinase 3) in NETs may have negative consequences for the host (Collins et al 2023). In our study, the level of cf-DNA was greater in the brains of mice with periodontitis than NE and MPO expression. These findings suggest that the decondensed DNA released from neutrophils in the brains of mice with periodontitis plays a key role in neuroinflammation.
Furthermore, we explored the effects of NETs on the neuron-glia unit. Several studies have shown that NET formation contributes to the pathogenesis of neurodegenerative and neuroinflammatory conditions (Shi et al 2023). NET formation can increase the secretion of proinflammatory cytokines to disrupt brain homeostasis and worsen brain injury (Li et al 2023). We report that NETs promoted M1 polarization in microglia but had no effect on M2 polarization. Microglia have 2 polarization phenotypes: M1-type microglia promote the neuroinflammatory response, and M2-type microglia inhibit the neuroinflammatory response. The polarized phenotypes of microglia determine the function of neuroinflammation (Li et al 2020). Coincidentally, the expression of inflammatory cytokines and adhesion molecules increased significantly in microglia after exposure to NETs, indicating M1 microglial activation. Although β-secretase (BACE1) and γ-secretase (PSEN1/PSEN2) activity is increased in neurons exposed to NETs, activated microglia can amplify the NETs-induced increase in aberrant APP processing.
The TLR4 activation is among the main triggers of the inflammatory response and regulates not only peripheral inflammation but also neuroinflammation (Hu et al 2020). NET formation seems to be induced from neutrophils after LPS stimulation through the binding of TLR4 (Shao et al 2019). As a result, in mice with periodontitis, we demonstrated that the expression of TLR4, Myd88, and pp65 increased, suggesting that activation of the TLR4 signaling pathway could be prevented by tak242. It has been reported that the GSK3β/Akt pathway is involved in regulating TLR4 expression (Guo et al 2023). Furthermore, western blot analysis revealed that NETs induced microglial M1 polarization and neuronal APP processing by modulating the activity of the GSK3β/Akt pathway. Taken together, the results of the in vitro and in vivo experiments highlight that P. gingivalis–induced NET mediated neuroinflammation through the TLR4/Myd88/NF-κB and GSK3β/Akt signaling pathways.
While periodontitis is linked to AD, this study is the first to demonstrate that P. gingivalis–induced NETs directly activate the TLR4/Myd88/NF-κB and GSK3β/Akt axes in the neuron–glia unit. However, chronic AD animal models and primary neurons/microglia cells should be obtained through further research. Our future experimental studies will also define the underlying molecular mechanisms using TLR4 gene knockout and PAD4 inhibitor animal models to determine the level of NET formation in periodontitis and address these important translational questions.
Author Contributions
L. Xu, contributed to conception and design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; Y. Zhou, contributed to conception, data analysis, drafted the manuscript; H. Chen, T. Li, contributed to conception, data acquisition, drafted the manuscript; J. Zhuang, Y. Jiang, contributed to conception, data interpretation, drafted the manuscript; Z. He, contributed to conception, data acquisition and interpretation, drafted the manuscript; Z. Song, W. Zhou, contributed to conception and design, data acquisition, analysis, interpretation, critically revised manuscript, and funding acquisition. All the authors gave their final approval and agree to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345261425925 – Supplemental material for Porphyromonas gingivalis–Induced NETs Mediate Neuroinflammation via TLR4 Activation
Supplemental material, sj-docx-1-jdr-10.1177_00220345261425925 for Porphyromonas gingivalis–Induced NETs Mediate Neuroinflammation via TLR4 Activation by L. Xu, Y. Zhou, H. Chen, J. Zhuang, Y. Jiang, T. Li, Z. He, Z. Song and W. Zhou in Journal of Dental Research
Supplemental Material
sj-docx-2-jdr-10.1177_00220345261425925 – Supplemental material for Porphyromonas gingivalis–Induced NETs Mediate Neuroinflammation via TLR4 Activation
Supplemental material, sj-docx-2-jdr-10.1177_00220345261425925 for Porphyromonas gingivalis–Induced NETs Mediate Neuroinflammation via TLR4 Activation by L. Xu, Y. Zhou, H. Chen, J. Zhuang, Y. Jiang, T. Li, Z. He, Z. Song and W. Zhou in Journal of Dental Research
Footnotes
Acknowledgements
The authors thank the Shanghai Key Laboratory of Translational Medicine on Ear and Nose Diseases (14DZ2260300) for support with the study.
A supplemental appendix to this article is available online.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the National Natural Science Foundation of China (82270974 and 82071112), the Science and Technology Commission of Shanghai Municipality (STCSM, No. 22ZR1437500), and the Cross Disciplinary Research Fund of Shanghai Ninth People’s Hospital, Shanghai Jiao Tong University School of Medicine (JYJC202225, JYJC202308).
Data Availability
All data used during the current study are available in the main text and appendix materials.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.