
Abstract
The oral and maxillofacial region is a complex microenvironment continuously exposed to microbial challenge, mechanical stress, and oncogenic stimuli. Accumulating human evidence firmly implicates pyroptosis in the pathogenesis of major oral and maxillofacial diseases, with cleaved gasdermin D (GSDMD)/gasdermin E (GSDME) detected in clinical specimens of periodontitis, peri-implantitis, apical periodontitis, oral lichen planus, and oral submucous fibrosis. The central mechanism involves inflammasome assembly (eg, NLRP3), leading to caspase activation, gasdermin cleavage (eg, GSDMD, GSDME), and consequent pyroptotic cell lysis. Functional in vivo studies establish a direct causal link: genetic ablation of caspase-1 attenuates alveolar bone loss in murine periodontitis models, while pharmacologic inhibition of NLRP3 mitigates fibrosis and slows disease progression in rat models of oral submucous fibrosis by suppressing pyroptosis. In peri-implantitis, NLRP3 suppression inhibits macrophage pyroptosis and promotes osteogenesis. Similarly, the activation of GSDMD and GSDME is shown to trigger pyroptosis and exacerbate pulpal and periapical inflammation in apical periodontitis models. Conversely, in oral squamous cell carcinoma, pyroptosis induction potently stimulates anti-tumor immunity. This context-dependent duality of pyroptosis provides the rationale for precision medicine. Emerging strategies reflecting this approach include cell-based therapies using dental pulp stem cells to alleviate pyroptosis and promote nerve regeneration in macaque models as well as advanced nanomaterials such as angiopoietin-coupled carbon nanotubes that modulate pyroptosis-related pathways to enhance bone regeneration in rabbits. Collectively, these insights position the precise modulation of pyroptosis as a new frontier in the management of oral and maxillofacial diseases.
Introduction
The oral and maxillofacial region is a sophisticated anatomical system of specialized tissues, supporting critical functions such as mastication, speech, and esthetics. Diseases affecting this region cause functional impairment, reduced quality of life, and substantial health care burden. Despite advances in conventional therapies, management remains challenging due to complex pathogenesis and high recurrence, underscoring the need for deeper mechanistic research and advanced diagnostic and therapeutic interventions.
Pyroptosis, an inflammatory programmed cell death, has emerged as a critical mechanism in oral and maxillofacial diseases. It is characterized by caspase-mediated gasdermin cleavage. The N-terminal fragments oligomerize to form membrane pores, causing lytic cell death and proinflammatory cytokine release (Chen et al 2022). Diverse stimuli, including bacterial pathogens, mechanical stress, and chemotherapeutic agents, trigger its activation via canonical, noncanonical, or alternative pathways (Booty and Bryant 2022). Accumulating evidence implicates pyroptosis in multiple oral and maxillofacial conditions, including periodontitis, peri-implantitis, apical periodontitis, and oral squamous cell carcinoma.
However, the role of pyroptosis in oral and maxillofacial pathologies is complex and context dependent. It amplifies inflammatory cascades and tissue destruction in conditions such as periodontitis while exerting antitumor effects through immunogenic cell death in oral cancers. This duality complicates therapeutic targeting and necessitates comprehensive understanding of its regulatory mechanisms, which not only provides promising therapeutic targets but also establishes new conceptual frameworks for clinical management.
The rise in publications on pyroptosis in oral and maxillofacial diseases underscores its growing research importance (Fig 1), yet a comprehensive review remains lacking. This review decodes the effect of pyroptosis by elucidating molecular mechanisms, detailing pathogenic functions, and critically evaluating emerging strategies (Fig 2). We further discuss translational challenges and future directions, highlighting pyroptosis as a promising frontier for disease management.
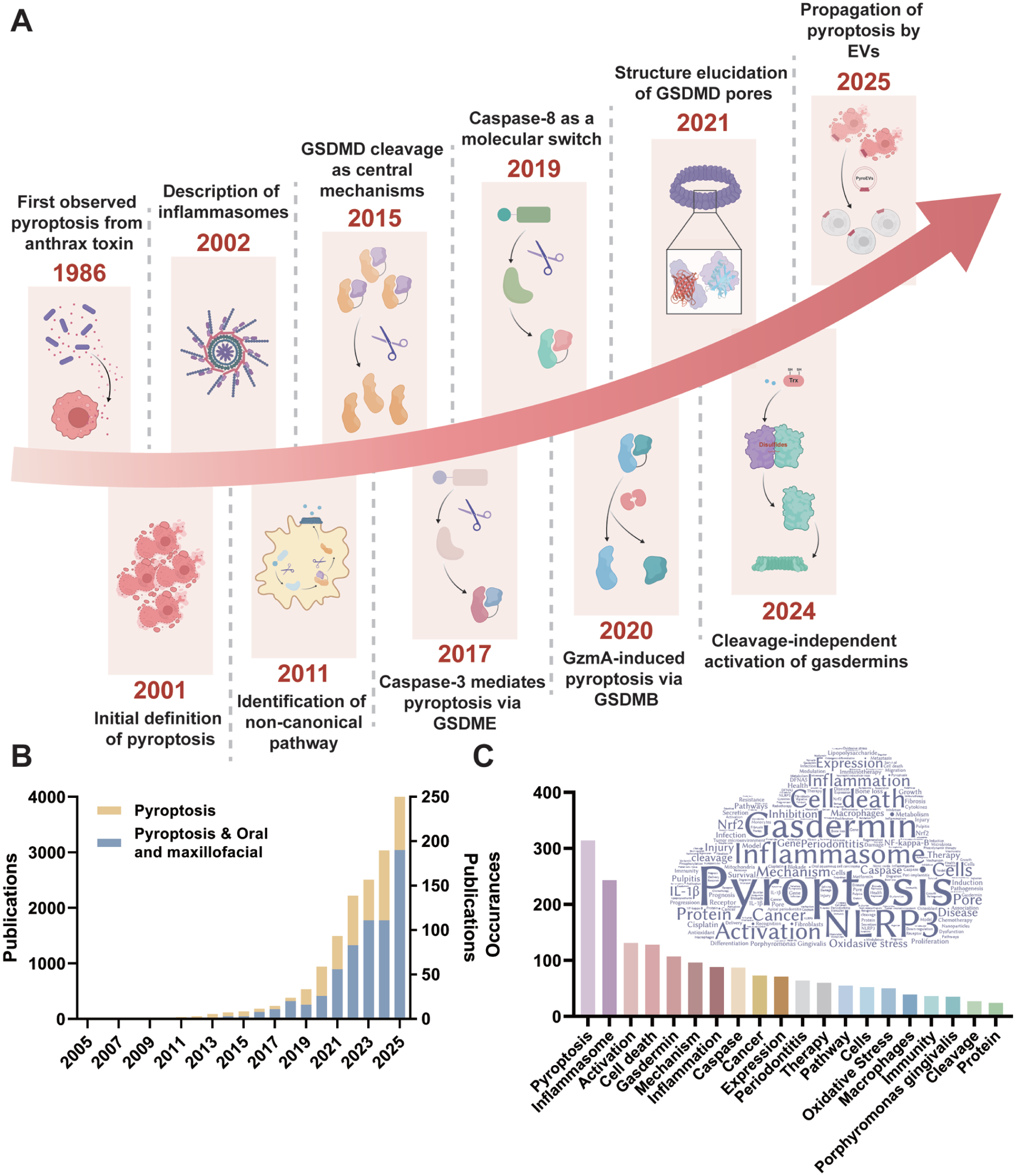
Key discoveries and advances in the development of pyroptosis. (
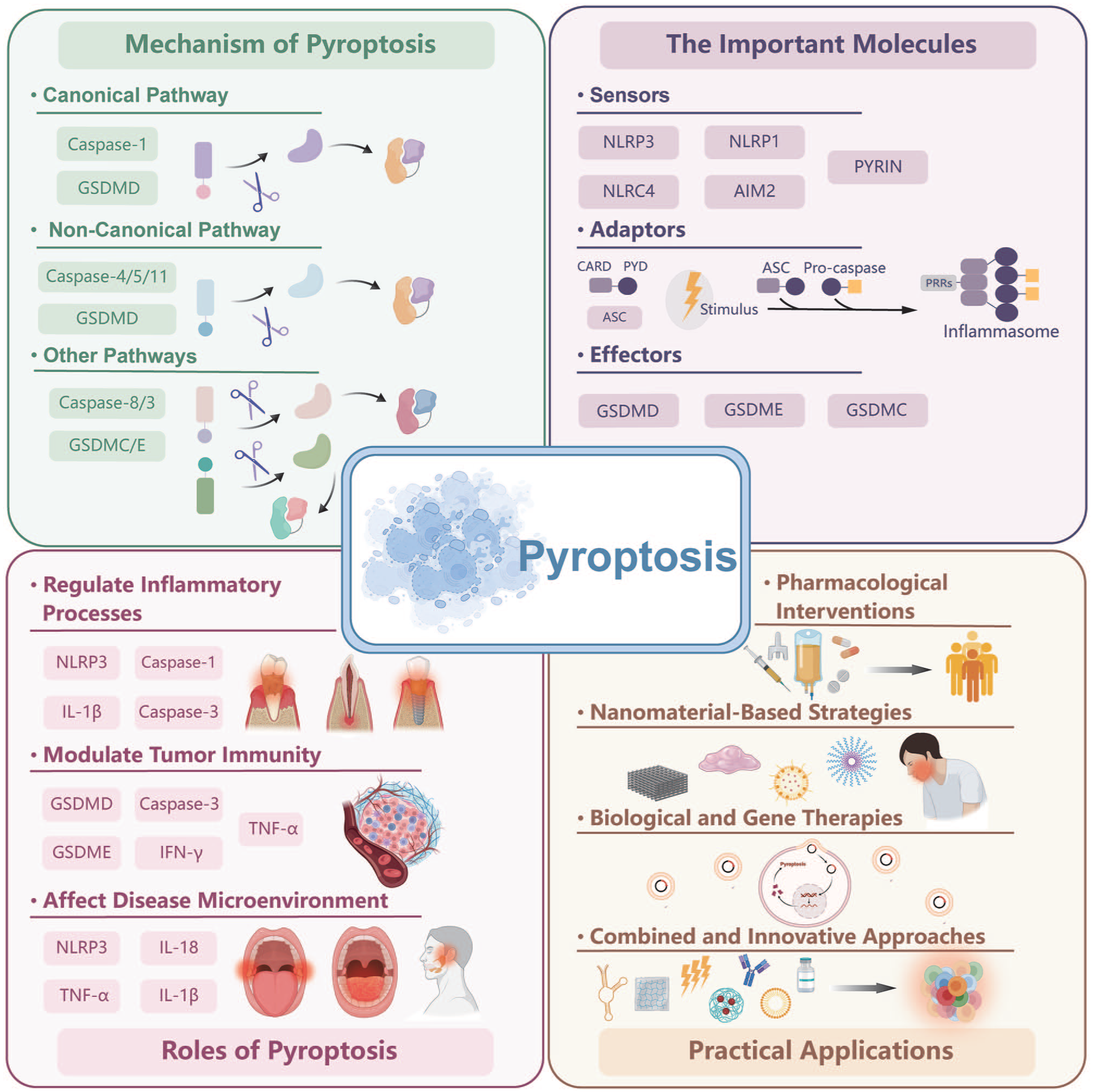
Schematic representation of pyroptosis in oral and maxillofacial diseases.
Mechanism of Pyroptosis
Pyroptosis is a lytic, proinflammatory programmed cell death executed by gasdermins that form membrane pores upon activation. Based on triggers and executors, pyroptosis pathways are classified into canonical, noncanonical, and other caspase-mediated pathways. Their functional significance is supported by growing in vivo evidence, forming the basis for innovative therapeutic strategies (Fig 3).
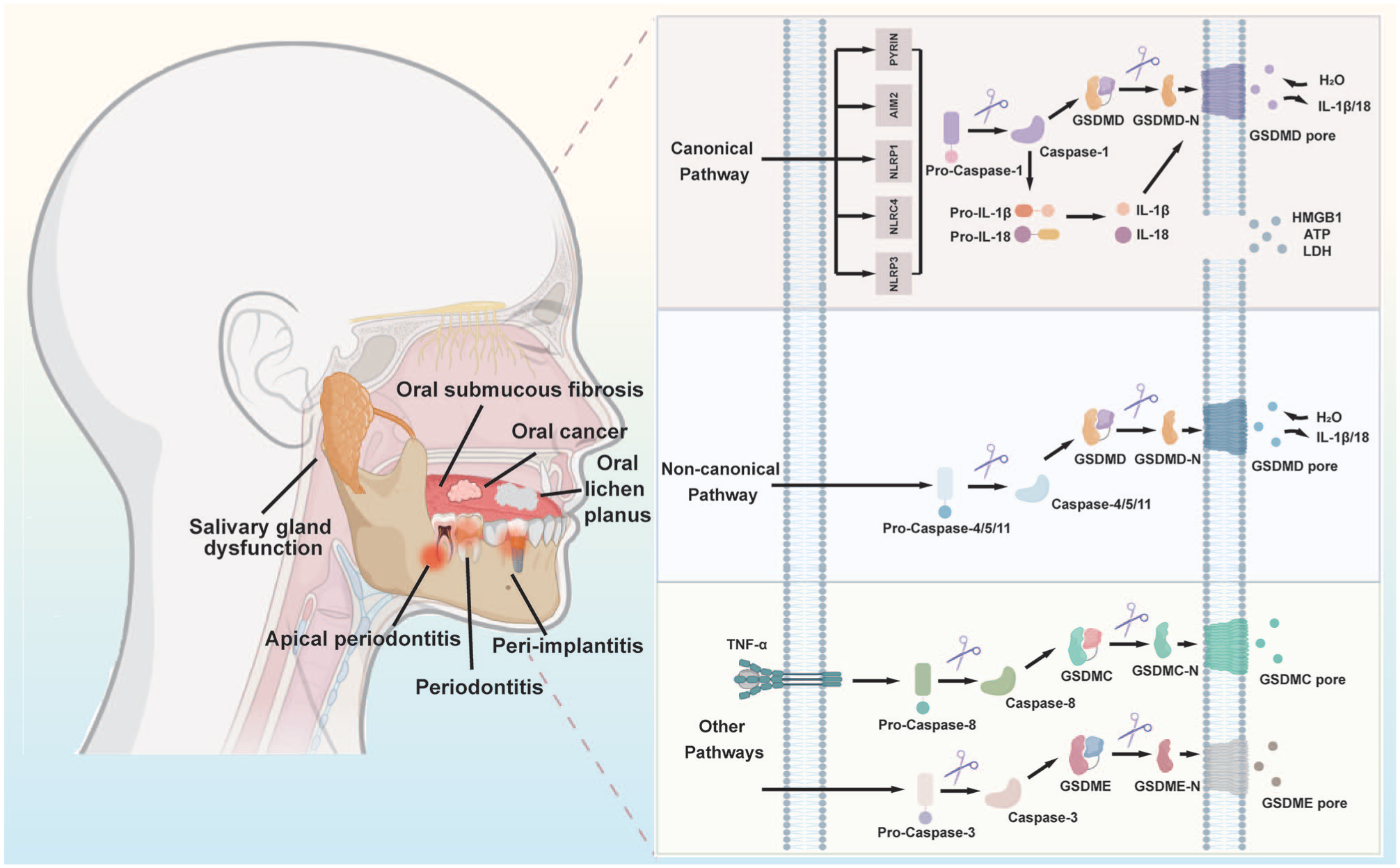
Molecular mechanism diagram of pyroptosis in oral and maxillofacial diseases.
The Canonical Pathway
The canonical pathway is initiated by inflammasome activation and executed by caspase-1. Inflammasomes are multiprotein complexes comprising pattern recognition receptors, such as NLRP3, NLRC4, and AIM2. These receptors sense pathogen-associated molecular patterns or damage-associated molecular patterns. While NLRP3 is the most extensively studied due to its broad responsiveness, other inflammasomes have specialized roles. AIM2 recognizes cytosolic double-stranded DNA, making it relevant for detecting DNA viruses and host DNA. NLRC4 responds to bacterial flagellin and type III secretion apparatus, positioning it as a key sensor for pathogens like Salmonella. Upon ligand binding, these receptors recruit ASC to activate caspase-1, which processes gasdermin D (GSDMD) and cleaves pro-interleukin (IL)–1β/pro-IL-18 into mature forms (Huang et al 2021). The N-terminal fragment of GSDMD (GSDMD-N) oligomerizes and inserts into the cell membrane, forming pores that lead to ion flux, cell swelling, and lytic release of inflammatory mediators.
The crucial role of this pathway is demonstrated by interventional studies in disease models. NLRP3 or caspase-1 ablation suppresses pyroptosis and mitigates bone loss in murine periodontitis (Rocha et al 2020; Zang et al 2020). During orthodontic movement, mechanical stress activates NLRP3 in periodontal ligament cells, inducing caspase-1–dependent pyroptosis to facilitate tooth movement (Chen et al 2024). In diabetes, alpha-2-glycoprotein 1 exacerbates periodontitis by enhancing NLRP3/caspase-1–mediated macrophage pyroptosis and M1 polarization (S. Yang et al 2024).
The Noncanonical Pathway
The noncanonical pathway is often initiated by lipopolysaccharide, which directly activates human caspase-4/5 or murine caspase-11. Activated caspases cleave GSDMD to generate pore-forming GSDMD-N, leading to pyroptotic lysis. The process can be amplified through secondary signals such as pannexin-1 channel activation, adenosine triphosphate (ATP) release, and P2X7 receptor stimulation (Kayagaki et al 2013).
This pathway plays a significant role in oral and maxillofacial infections. Caspase-11 deficiency alters subgingival microbiota and attenuates periodontitis in mice, providing direct genetic proof of its role (Fu et al 2024). Oral pathogens such as Treponema denticola activate caspase-4 in gingival fibroblasts, contributing to periodontitis through the noncanonical pathway. Therapeutically, an ATP-responsive metal-organic framework that suppresses caspase-11/GSDMD activation has demonstrated efficacy in mitigating periodontitis in preclinical models (Q. Yang et al 2024).
Other Caspase-Mediated Pathways
Beyond caspase-1 and caspase-4/5/11, other caspases can also induce pyroptosis by cleaving distinct gasdermins.
Activated caspase-3 executes pyroptosis by cleaving gasdermin E (GSDME). This pathway contributes to tissue destruction in peri‑implantitis, where tumor necrosis factor–α (TNF-α) triggers caspase-3/GSDME-mediated pyroptosis in gingival epithelial and osteoblastic cells, accelerating bone loss (Chen et al 2021). Conversely, in head and neck squamous cell carcinoma (HNSCC), it functions in tumor suppression via MTUS1/ATIP, which activates caspase-3 through mitochondrial dysfunction (Tang et al 2024). More importantly, therapeutic potentials of this pathway are being recognized. Porphyrin‑based photodynamic therapy induces caspase‑3/GSDME-mediated pyroptosis to inhibit oral squamous cell carcinoma (OSCC) progression (X. Liu et al 2023).
Under certain conditions, caspase-8 cleaves gasdermin C (GSDMC), redirecting cell fate from apoptosis to pyroptosis. In the tumor microenvironment, TNF-α stimulation and PD-L1 expression promote caspase-8–dependent GSDMC cleavage, facilitating tumor necrosis (Zhang et al 2024). Infections can also activate caspase-8 via the RIP1/FADD complex, inducing pyroptosis in immune and oral cells (Sarhan et al 2018).
The involvement of multiple caspases and gasdermins highlights the diversity and context specificity of pyroptosis, providing diverse therapeutic targets. However, gasdermin cleavage alone does not confirm lytic pyroptosis. Living cells can secrete IL-1β through nonlytic mechanisms such as exosome release (Ford et al 2025). In addition, cells possess compensatory mechanisms to eliminate gasdermin pores and escape lysis, including membrane budding and ESCRT-dependent repair that shed pore-containing vesicles (Nozaki and Miao 2023). Therefore, complementary approaches are needed to definitively establish lytic pyroptosis.
Pyroptosis is increasingly recognized to interact with other cell death pathways. PANoptosis describes an integrated pathway involving pyroptosis, apoptosis, and necroptosis, mediated by PANoptosomes that engage caspases and RIPKs. For instance, caspase-8 deficiency not only blocks apoptosis but sensitizes cells to RIPK3-dependent necroptosis and modulates GSDMD cleavage (Upmanyu and Upadhyay 2025). Therefore, future studies should consider the integrated death network and compensatory mechanisms.
Pyroptosis in Oral and Maxillofacial Diseases
Accumulating evidence confirms pyroptosis as a key pathogenic mechanism in oral and maxillofacial diseases. Beyond executing inflammation, pyroptosis acts as a critical signaling nexus that integrates microbial challenge, mechanical stress, metabolic dysregulation, and immune responses. This section discusses evidence linking pyroptosis to specific disease processes, providing a mechanistic foundation for therapeutic interventions (Appendix Table 1).
Inflammatory Diseases
Periodontitis
Periodontitis is a chronic inflammatory disease that affects tooth-supporting structures. Pyroptosis has been identified as a central pathogenic mechanism, with genetic, pharmacologic, and clinical evidence demonstrating its role in driving persistent inflammation and alveolar bone loss.
The pathogenesis involves multiple periodontal cells, including gingival fibroblasts, epithelial cells, and periodontal ligament stem cells (PDLSCs). They undergo pyroptosis in response to bacterial challenges such as Porphyromonas gingivalis. Genetic ablation of NLRP3 or caspase-1 attenuates bone loss by inhibiting pyroptosis, underscoring their essential role (Rocha et al 2020; Zang et al 2020). Human data support translational relevance, as clinical samples from periodontitis patients show elevated expression of caspase-1, caspase-4, and NLRP3 (Bostanci et al 2009; Y. Li et al 2021). In addition, mechanical stress and dental calculus also induce pyroptosis, indicating multifactorial activation.
Emerging evidence reveals complex regulatory mechanisms. Circadian disruption through BMAL1 downregulation exacerbates periodontitis by enhancing GSDMD-mediated pyroptosis, suggesting chronotherapeutic interventions might be beneficial (Y. Wang et al 2025). At the epitranscriptomic level, METTL3-mediated m6A modification stabilizes NLRP3 mRNA, amplifying macrophage pyroptosis and accelerating diabetic periodontitis. This positions METTL3 as a promising therapeutic target for metabolic stress–associated periodontal destruction (Yang et al 2025).
Pyroptosis also serves as a central node in intercellular crosstalk and metabolic reprogramming. Pyroptotic macrophages release factors that disrupt glutamate metabolism in PDLSCs, impairing osteogenic differentiation (Sun et al 2024). In diabetes, hepatic exosomes carrying Fasn induce ectopic fatty acid synthesis in periodontal cells, activating GSDMD cleavage and revealing a systemic-paracrine axis of pyroptosis regulation (J. Liu et al 2023). Beyond pyroptosis alone, emerging evidence suggests that periodontitis involves crosstalk among multiple cell death pathways. P. gingivalis can simultaneously activate pyroptosis, apoptosis, and necroptosis in macrophages, exacerbating tissue damage (Wu et al 2025).
Therapeutic translation is progressing beyond conventional approaches. Natural compounds such as caryacrol inhibit pyroptosis through the PVT1/miR-20a-5p axis, indicating potential for periodontitis management (Yu et al 2024). Genetic interventions targeting AZGP1 (S. Yang et al 2024) or METTL3 (Zhou et al 2024) have demonstrated efficacy in preclinical models, suggesting that pathway-specific inhibition may achieve precise pyroptosis modulation.
In summary, pyroptosis in periodontitis involves intricate signaling networks, intercellular crosstalk, and metabolic regulation. Strategies that precisely modulate pyroptosis represent a promising frontier for achieving periodontal homeostasis and regeneration.
Apical periodontitis
Apical periodontitis (AP) arises from pulpal infection and involves periapical bone resorption mediated by immune activation. Pyroptosis contributes significantly to AP pathogenesis, evidenced by elevated caspase-1, -4, and -5 in human periapical lesions. The NLRP3 inflammasome activates caspase-1, cleaving GSDMD to execute pyroptosis. Notably, the AIM2 inflammasome also plays a role, sensing host-derived damaged DNA from necrotic pulp cells and amplifying inflammatory responses. This suggests that multiple inflammasome pathways may converge to drive periapical inflammation. Subsequent cell lysis releases mediators including IL-1β and IL-18, stimulating osteoclastogenesis and bone destruction.
Recent studies indicate that pyroptosis of apical papilla stem cells disrupts local immune homeostasis by impairing regulatory T-cell function, accelerating periapical bone loss (K. Wang et al 2024). Mitochondrial DNA leakage from stressed odontoblasts activates GSDMD-mediated pyroptosis and the cGAS-STING axis, jointly exacerbating pulpal and periapical inflammation (Zhang et al 2021). Bacterial infections such as Fusobacterium nucleatum activate ZBP1-mediated GSDME cleavage, triggering pyroptosis and inflammatory responses (Liu et al 2022). Importantly, pyroptosis exhibits a concentration-dependent dual role in AP. Pyroptosis exerts protective effects at low levels but becomes destructive upon excessive activation, highlighting the need for precisely targeted therapeutic strategies.
Targeting pyroptosis presents a promising strategy for AP management. Ginsenoside Rb1 ameliorates AP by suppressing intracellular calcium overload and subsequent macrophage pyroptosis (Guan et al 2025). Innovative approaches, such as intercellular mitochondrial transfer from mesenchymal stem cells to odontoblasts, alleviate NLRP3-induced pyroptosis and dental pulp damage in rat models, suggesting a novel regenerative strategy (K. Wang et al 2023).
In summary, pyroptosis is a key driver of periapical inflammation and bone loss. Its modulation represents a potential therapeutic strategy, although interventions must be precisely tuned given the context-dependent nature of pyroptosis. Notably, the inflammatory microenvironment in AP may engage multiple death pathways simultaneously, suggesting therapeutic strategies should consider the broader cell death network.
Peri-implantitis
Peri-implantitis is an aggressive inflammatory condition leading to dental implant failure through progressive bone destruction. Although it shares some inflammatory features with periodontitis, peri-implantitis is distinguished by its unique implant–tissue interface, which creates a distinct microenvironment for pyroptosis activation.
Elevated cleaved caspase-3 and GSDME in human peri-implantitis soft tissues confirm the involvement of pyroptosis (Chen et al 2021). Osteoblast pyroptosis at the implant–bone interface directly compromises osseointegration. Macrophage–osteoblast crosstalk amplifies this process, where lncRNA NRIR promotes NLRP3 assembly and macrophage pyroptosis (L. Wang et al 2025). Pyroptosis-associated release of inflammatory mediators inhibits osteogenic differentiation of mesenchymal stem cells attempting to colonize the implant surface, thereby coupling pyroptosis to impaired regeneration. Beyond bacterial components, dysregulated ion signaling via the TRPM2 channel facilitates NLRP3 assembly, promoting pyroptosis while suppressing osteogenesis in models of inflammatory bone loss (Li et al 2023).
While treatments focusing on biofilm control often fail to arrest disease progression, targeting pyroptosis represents a promising complementary strategy. For instance, exosomes from adipose-derived mesenchymal stromal cells inhibit NF-κB/NLRP3/IL-1β–mediated macrophage pyroptosis and enhance mucosal healing (Zheng et al 2024), supporting the potential of biological therapies delivering antipyroptosis agents for peri-implantitis management.
In summary, pyroptosis in peri-implantitis is characterized by direct osteogenesis impairment at the implant–bone interface and the creation of a hostile microenvironment that impedes implant integration. Modulating pyroptosis offers a promising frontier for improving the long-term success of implants. Future studies should validate these mechanisms in disease models and explore gasdermin fragments in peri-implant crevicular fluid as early diagnostic biomarkers.
Cancer
HNSCC, the sixth most common cancer globally, exhibits complex and context-dependent interactions with pyroptosis. Pyroptosis exerts a dual role in tumorigenesis and antitumor immunity, dictating its potential as either a therapeutic target or a therapeutic tool.
The antitumor role of pyroptosis is mediated through its capacity to stimulate immunogenic cell death. GSDME, overexpressed in OSCC, serves as a key mediator. Upon cleavage by chemotherapy-activated caspase-3, GSDME executes pyroptosis and releases damage-associated molecular patterns. This cascade represents an emerging strategy for converting immunologically “cold” tumors into “hot” ones, enhancing sensitivity to T-cell attacks and immune checkpoint inhibitors. Consequently, GSDME expression correlates with improved chemotherapy response and favorable prognosis, positioning it as a potential biomarker to guide treatment decisions. Notably, neutrophil extracellular traps are found to promote HNSCC metastasis by suppressing NLRP3-mediated pyroptosis, revealing a mechanism that undermines the antitumor potential of pyroptosis (Zhai et al 2024). Beyond GSDME, the targeted induction of pyroptosis via nanoparticle delivery of GSDMD-N represents a promising strategy to enhance tumor immunogenicity (Huang et al 2025a).
Conversely, pyroptosis also exhibits a protumorigenic role, wherein chronic inflammation drives aberrant NLRP3 activation in the tumor microenvironment. Persistent NLRP3 signaling fosters an immunosuppressive environment by inducing gasdermin-mediated pyroptosis, which releases cytokines that create a feed-forward loop conducive to tumor growth and metastasis. In addition, AIM2 expression correlates with immune infiltration and influences immunotherapy responses in OSCC, and NLRC4 expression has been linked to an increased risk of tumor progression. Furthermore, bioinformatic analyses show that low pyroptosis scores correlate with enhanced immune activation and improved immunotherapy response (Gao et al 2023). This duality suggests that the effect of pyroptosis is context dependent, determined by timing, magnitude, cellular context, and the specific pathways involved.
Emerging evidence reveals unexpected connections between pyroptosis and therapy resistance. Neoadjuvant chemotherapy with low-dose cisplatin upregulates GSDMD in OSCC, which conversely enhances metastasis through MMP14-mediated epithelial-mesenchymal transition activation (Huang et al 2025b). This indicates that merely upregulating gasdermins without complete pyroptosis regulation might unexpectedly promote tumor progression. In addition, novel therapeutic targets are emerging, as demonstrated by the finding that tumor-intrinsic SIRPA drives pyroptosis evasion in HNSCC by enhancing GSDME ubiquitination and inhibiting succinylation (Song et al 2025).
Targeting the duality of pyroptosis requires precise, context-dependent modulation, a challenge compounded by the intricate crosstalk among different cell death pathways. For instance, oncolytic viruses coordinately induce pyroptosis, apoptosis, and necroptosis in HNSCC, resulting in potent antitumor immunity (Chen et al 2025). Therefore, strategies that locally induce pyroptosis (eg, GSDME reactivation, photodynamic therapy) could synergize with immunotherapy to initiate an antitumor cycle, as seen with CTLA-4 blockade inducing tumor pyroptosis via T-cell–derived cytokines (S. Wang et al 2023). In contrast, in premalignant conditions, suppressing pyroptosis with NLRP3 inhibitors may prevent immune evasion and progression (Jalali et al 2024). Future research should focus on elucidating the integrated cell death network, identifying predictive biomarkers, and addressing compensatory pathway activation to realize the full therapeutic potentials of these strategies.
Other Oral and Maxillofacial Diseases
Pyroptosis contributes to diverse oral and maxillofacial diseases beyond common inflammatory and neoplastic conditions. Understanding these mechanisms provides novel insights into disease progression and reveals potential therapeutic options.
Oral lichen planus (OLP), a chronic inflammatory disorder with malignant potential, features elevated NLRP3 activity and IL-1β/IL-18 levels in affected human tissues (Thi Do et al 2018). Pyroptotic keratinocytes activate NF-κB signaling in T cells, promoting OPA1-mediated mitochondrial fusion and glycolytic reprogramming, which further perpetuate disease progression (Z. Yang et al 2024). This mechanism positions OLP as a disorder driven by an integrated pyroptosis-inflammation-metabolism axis, supported by clinical evidence such as elevated serum IL-18 and disease-associated genetic polymorphisms (El-Howati et al 2023). Targeting the pyroptosis-inflammation-metabolism axis in keratinocytes or interrupting the subsequent T-cell response represents a promising strategy for OLP management.
Oral submucous fibrosis (OSF) is a potentially malignant disorder associated with the consumption of areca nuts. Arecoline, a major alkaloid in areca nuts, induces reactive oxygen species (ROS)–mediated NLRP3 activation in oral myofibroblasts, promoting migration and collagen deposition. Upregulated NLRP3 expression and related signaling components are observed in human OSF tissues, indicating the contribution of pyroptosis to disease pathogenesis (You et al 2019). In addition, emerging evidence suggests that areca nut may induce pyroptosis in epithelial or fibroblastic cells, further amplifying the inflammatory response that drives fibrosis (Yu et al 2024).
Pyroptosis also contributes to salivary gland dysfunction in conditions such as Sjögren’s syndrome and radiation-induced damage. Inflammasome activation and gasdermin-mediated pyroptosis impair acinar cell function, reducing saliva secretion and compromising oral health (S. Li et al 2021). Pyroptosis inhibition restores salivary flow in experimental models, suggesting a potential therapeutic strategy for managing xerostomia (Yu et al 2022). Further evidence indicates that blocking pyroptosis effectively prevents radiation injury of salivary glands, which is exacerbated by a proinflammatory milieu involving cytokines such as IL-1β and IL-18 (Qu et al 2024).
In summary, pyroptosis serves as a common pathogenic mechanism across numerous oral and maxillofacial diseases. Targeting specific upstream triggers or core components of pyroptosis holds significant promise for alleviating these challenging conditions.
Therapeutic Strategies Targeting Pyroptosis in Oral and Maxillofacial Diseases
Molecular characterization of pyroptosis has offered novel therapeutic opportunities for oral and maxillofacial diseases (Fig 4). This section reviews recent advances in pharmacologic, nanomaterial-based, biological, and gene therapies targeting pyroptosis. Combined and innovative approaches are also discussed, emphasizing their mechanistic basis and translational potential (Appendix Table 2).
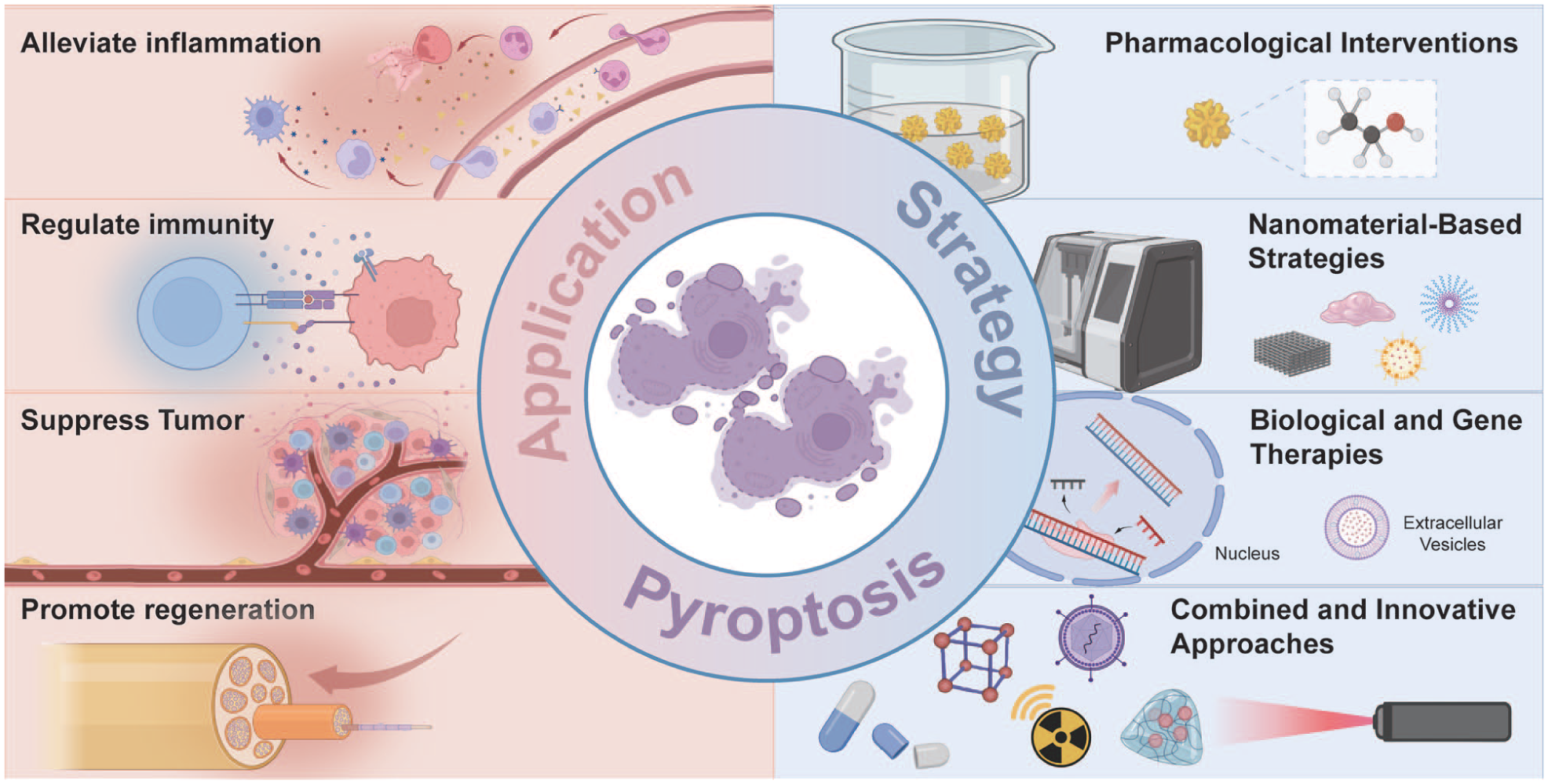
Summary of therapeutic strategies targeting pyroptosis in oral and maxillofacial diseases.
Pharmacologic Interventions
The pharmacologic intervention of pyroptosis is a promising therapeutic strategy for oral and maxillofacial diseases. Several representative compounds have demonstrated efficacy in preclinical models.
Dimethyl fumarate, a Food and Drug Administration–approved agent, attenuates noncanonical pyroptosis in periodontal ligament fibroblasts and dental pulp cells, highlighting potentials for treating pulpitis and AP (Gu et al 2023). Inorganic nitrate mitigates radiation-induced salivary gland dysfunction by suppressing caspase-1–mediated pyroptosis, thereby preserving acinar cell viability and function (Li et al 2021). Okanin induces pyroptosis in oral cancer cells through ROS-mediated activation, promoting immunogenic cell death and converting immunologically “cold” tumors into “hot” microenvironments (Chia et al 2024).
However, most pharmacologic interventions remain at the preclinical stage, facing challenges including off-target effects, limited bioavailability, and insufficient tissue specificity. Repurposing existing drugs, developing novel compounds targeting specific molecules, and optimizing local delivery systems (eg, intrapocket gels or films) represent important future directions.
Nanomaterial-Based Strategies
Nanomaterials have emerged as a prominent research focus in biomedical science, offering innovative solutions to overcome conventional therapy limitations such as poor bioavailability, nonspecific biodistribution, and off-target effects. Several nanomaterial-based strategies have been developed and shown promise in modulating pyroptosis.
Smart nanomaterials enable spatially and temporally controlled modulation of pyroptosis. Glutathione/ROS‑responsive diselenide‑based nanoprodrugs release chemotherapeutic agents and photosensitizers, inducing pyroptosis to promote dendritic cell maturation, enhance T-cell responses, and establish long-term immune memory against distant tumors (Wan et al 2023). Similarly, pH-responsive nanomicelles that co-deliver a Src inhibitor and chemotherapeutic agents induce pyroptosis in acidic tumor microenvironments, potentiating antitumor immunity in HNSCC (Zhu et al 2022). Nucleus-targeted nanoparticles deliver GSDMD‑N plasmid to OSCC cells, triggering pyroptosis and NK cell–mediated antitumor immunity (Huang et al 2025a). In inflammatory diseases, ATP-responsive metal-organic frameworks release Mg2+/Zn2+ to inhibit GSDMD activation, attenuating periodontal inflammation (Q. Yang et al 2024).
These nanomaterials enable spatiotemporal pyroptosis regulation, suggesting their value in precision therapy. Despite promising preclinical efficacy, translational success requires addressing long-term biosafety, potential immune reactivity, and large-scale manufacturing. No clinical trials that directly target pyroptosis have yet been established. Rigorous toxicology studies and large animal model validation are prerequisites for clinical translation.
Biological and Gene Therapies
Biological interventions based on extracellular vesicles, gene editing, and oligonucleotide technologies are emerging as powerful strategies for pyroptosis modulation.
Exosomes from dental follicle stem cells deliver miR-140-3p to suppress NLRP3-mediated pyroptosis in PDLSCs via the DNMT1/SOCS1/NF-κB axis, reducing orthodontic root resorption (Li et al 2025). HSV-1–based oncolytic viruses show promise as a cancer treatment by selectively replicating in tumors and activating ZBP1 to promote antitumor immunity (S. Wang et al 2024). Gene editing reveals that METTL3 knockout alleviates periodontitis by enhancing TNFAIP3 mRNA stability and inhibiting macrophage pyroptosis via suppression of NLRP3 assembly (Zhou et al 2024). RNA interference techniques knocking down Prdx1 elevate ROS levels and trigger pyroptosis in OSCC, suppressing tumor proliferation (Ye et al 2023).
These biologically inspired therapies show promise for specific pyroptosis regulation, but clinical translation is limited by delivery efficiency and immune responses. Safety concerns also remain, including host defense impairment, off-target gene-editing effects, and insertional mutagenesis. Addressing these issues requires stable, targeted delivery and long-term studies to assess therapeutic durability and risks such as malignant transformation.
Combined and Innovative Approaches
The complex pathogenesis of oral and maxillofacial diseases requires multitarget therapeutic strategies. Recent studies increasingly emphasize combined approaches integrating pyroptosis regulation with other treatments to achieve synergistic efficacy.
In cancer therapy, pH-responsive nanoparticles delivering siGSDMD inhibit OSCC growth by suppressing pyroptosis and epithelial-mesenchymal transition (Huang et al 2025b). Multifunctional nanocomposites such as TiO2@Ru@siRNA combine photodynamic therapy with HIF-1α silencing to alleviate hypoxia and induce pyroptosis, thereby enhancing T-cell infiltration and antitumor responses (Zhou et al 2022). For inflammatory diseases, tetrahedral framework nucleic acids improve metformin delivery to target pyroptosis and AMPK signaling, ameliorating diabetic periodontitis (Han et al 2025). In regenerative applications, cell-based therapies involving DPSCs transfer mitochondria to Schwann cells, alleviating pyroptosis and promoting nerve regeneration (Zheng et al 2025). Thus, pyroptosis modulation extends beyond eliminating pathological cells to preserving and enhancing regenerative cell function.
These innovative approaches represent a strategic transition from single-target to multimodal strategies. However, all evidence currently derives from preclinical models, with no established clinical trials targeting pyroptosis in the oral and maxillofacial region. By integrating nanotechnology, immunomodulation, metabolic reprogramming, and cell therapy, they enable precise pyroptosis regulation in complex disease microenvironments. This aligns with precision medicine principles, emphasizing concurrent targeting of the pyroptosis-inflammation-metabolism network to achieve synergistic effects and overcome compensatory mechanisms.
Current Limitations and Knowledge Gaps
Despite evidence linking pyroptosis to oral and maxillofacial diseases, several limitations and knowledge gaps warrant consideration. First, the relative strength of the evidence varies across study types. In vitro studies offer mechanistic insights, while animal models provide functional validation but face species differences (eg, murine caspase-11 and human caspase-4/5) and cannot fully recapitulate human diseases. Human data are most clinically relevant, yet interventional studies directly targeting pyroptosis are lacking. Second, many studies are expression-based, reporting elevated pyroptosis-related molecules in diseased tissues. However, such data do not establish causality, as gasdermin cleavage does not confirm lytic cell death, and bulk tissue analysis cannot identify which specific cell types undergo pyroptosis. Third, gaps persist between mechanistic findings and clinical translation. Most therapeutic strategies are still preclinical, with human safety, bioavailability, and efficacy largely unexplored. The optimal therapeutic window and standardized biomarkers are also lacking. Addressing these limitations through clinical trials and biomarker development is essential before clinical application.
Conclusions and Future Perspectives
Pyroptosis has emerged as a key mechanism in oral and maxillofacial diseases, contributing to tissue destruction, inflammatory responses, and disease progression. Fundamentally, pyroptosis exhibits a dual nature, acting as a double-edged sword in different pathological contexts (Fig 5), which necessitates precise therapeutic strategies. Thus, elucidating pyroptosis mechanisms and disease-specific regulatory networks is critical. Although the integration of nanotechnology breakthroughs with deepening pyroptosis insights has facilitated the development of advanced strategies, clinical translation faces challenges including therapy-induced resistance and compensatory mechanisms.
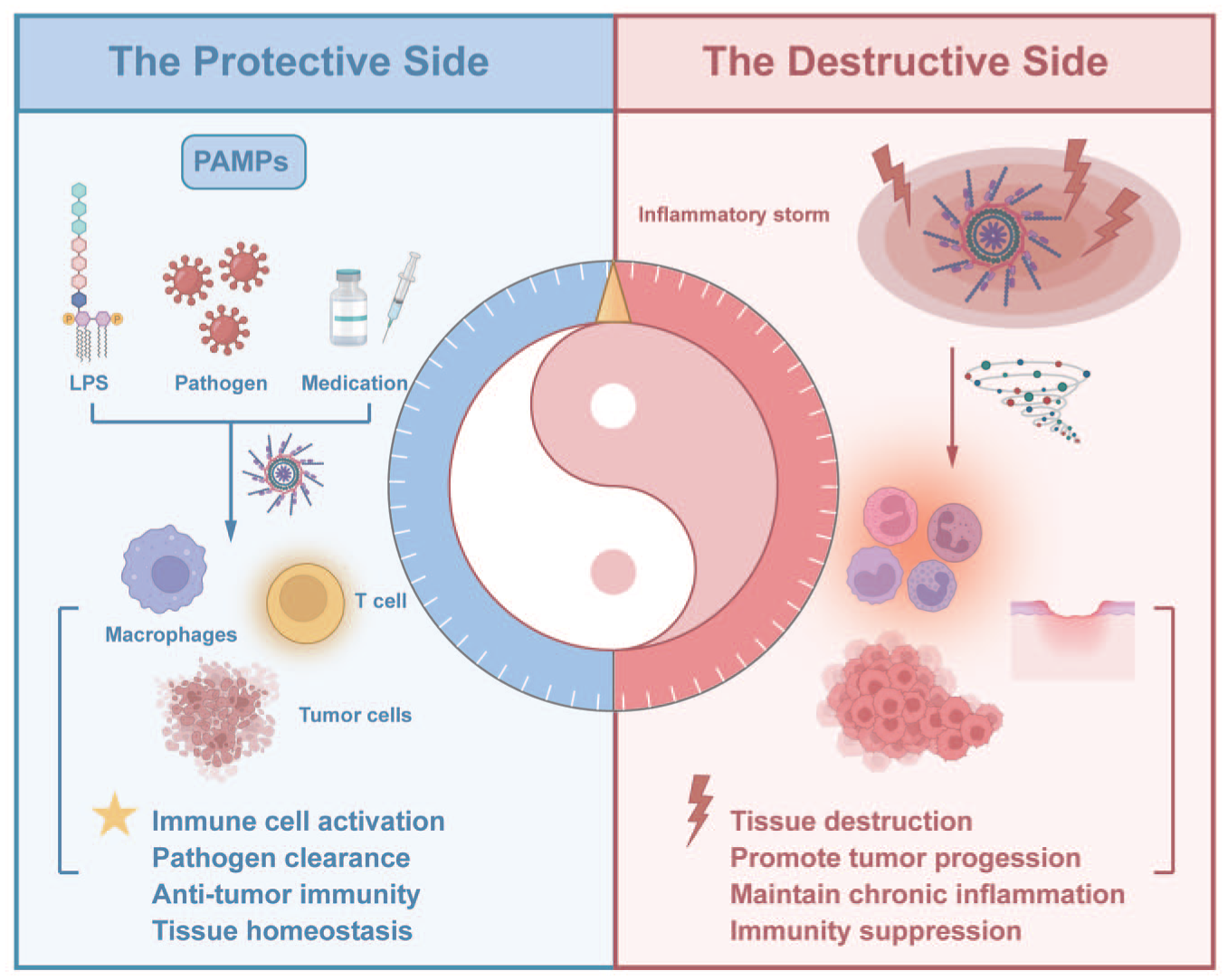
The dual role of pyroptosis in disease development.
From a clinical translation perspective, future research should prioritize several key directions (Fig 6). First, robust biomarkers (eg, circulating gasdermins, inflammatory cytokines, caspase products) need validation for patient stratification and treatment monitoring. Detection in saliva and gingival crevicular fluid enables noninvasive real-time monitoring of disease activity and treatment response. Although preliminary evidence supports the presence of these biomarkers in saliva, assay feasibility requires systematic validation of their stability, concentration, and correlation with disease severity. Regarding specificity, as existing markers such as IL-1β reflect general inflammation, gasdermin fragments may better indicate pyroptosis-driven destruction. But their added clinical value requires careful assessment. Second, the accessibility of the oral cavity allows localized delivery using injectable hydrogels, mucoadhesive patches, or microenvironment‑responsive nanoparticles to maximize bioavailability and minimize side effects. Third, combining pyroptosis modulation with other cell-death pathways may overcome compensatory resistance and enhance therapeutic efficacy. Finally, translation requires validation in clinically relevant models including patient‑derived organoids and 3-dimensional tissue models that recapitulate the oral and maxillofacial microenvironment.
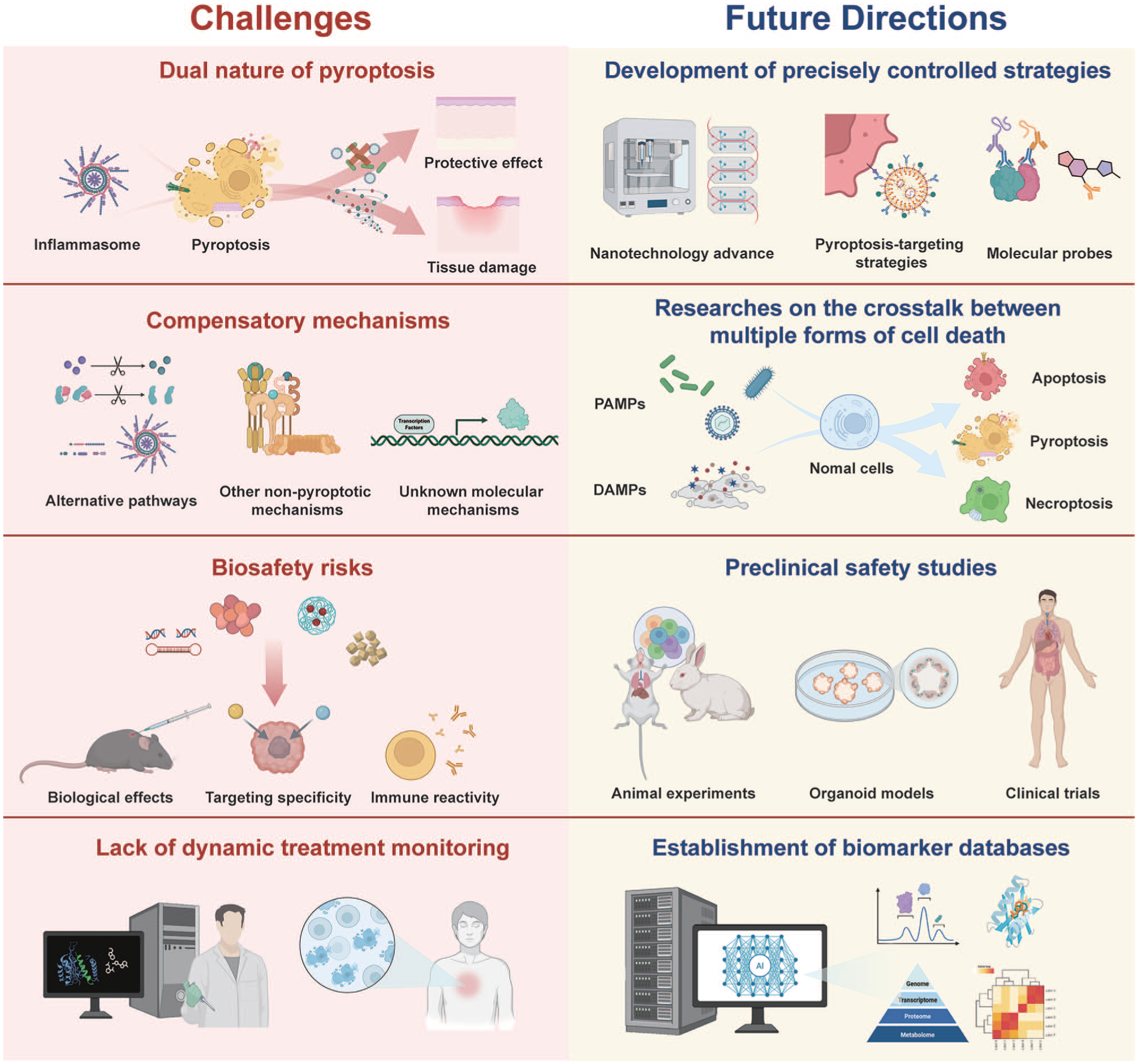
Future directions and challenges for therapeutic strategies targeting pyroptosis in oral and maxillofacial diseases.
In summary, while pyroptosis targeting represents a promising frontier, clinical success requires interdisciplinary collaboration and continued advances in both mechanistic understanding and technology development. We anticipate that pyroptosis-targeted treatments may soon offer novel solutions for managing challenging oral and maxillofacial diseases.
Author Contributions
L. Tang, Y. Wang, contributed to conception and design, data acquisition, analysis, and interpretation, drafted and critically revised manuscript; Z. Yu, Y. Chai, Y. Chen, contributed to analysis, critically revised the manuscript; G. Yang, T. Huang, contributed to conception and design, critically revised the manuscript. All authors gave their final approval and agree to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345261443990 – Supplemental material for Targeting Pyroptosis in Oral and Maxillofacial Diseases
Supplemental material, sj-docx-1-jdr-10.1177_00220345261443990 for Targeting Pyroptosis in Oral and Maxillofacial Diseases by L. Tang, Y. Wang, Z. Yu, Y. Chai, Y. Chen, G. Yang and T. Huang in Journal of Dental Research
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (grant 82471005).
Data Sharing and Data Accessibility
Data availability is not applicable to this article as no new data were created or analyzed in this study. All discussed data and findings are based on previously published studies, which are cited in the reference list.
A supplemental appendix to this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.