
Abstract
Epithelial rests of Malassez (ERMs) constitute a 3-dimensional epithelial meshwork extending from the tooth apex to the junctional epithelium (JE). This position, coupled with observations from a variety of injury models, suggests that ERMs may be involved in sensing disruptions to the mechanical integrity of the periodontium. To test this hypothesis, finite element modeling was employed to predict strain states in tissues surrounding ERMs (e.g., the periodontal ligament [PDL], supracrestal connective tissues, and the JE), which, by virtue of their attachment to the tooth surface, experience distortional strains during mastication. An in vivo hyper-loading mouse model was employed to map these distortional strains to location-specific and time-dependent morphological changes in ERMs. Quantitative immunohistochemistry was used to interrogate ERMs for changes in the expression of Piezo1, β4 integrin, plectin, YAP, TRPV4, and phosphorylated FAK. Collagen and oxytalan fiber organization around ERMs was compared between normal- and hyper-loading conditions. Axin2CreERT2;R26RmTmG/+ mice were used to lineage-trace Wnt-responsive ERMs under normal and hyper-loading conditions, and Wnt signaling from ERMs and the JE was disrupted using K14CreERT2;Wlsfl/fl mice. Collectively, these analyses predicted that hyper-loading produced high distortional strains in the JE and supracrestal connective tissues, which correlated with sites of ERM enlargement. ERMs expressed mechanosensory proteins, whose levels were significantly elevated by hyper-loading. Load-activated ERMs were Wnt-responsive, and if Wnt secretion was blocked, then ERMs no longer expressed mechanosensory proteins and no longer enlarged in response to hyper-loading. A deterioration in the JE attachment and the PDL followed. Thus, the ERM network is positioned to sense distortional strains produced by mastication; they possess the molecular machinery to sense and respond to masticatory forces and, via a Wnt-dependent mechanism, coordinate tissue-level responses to mechanical loading in the PDL, supracrestal connective tissues, and the JE.
Keywords
Introduction
The periodontium remodels in response to physical forces (Hine 1962). In the jaw skeleton, masticatory forces are transferred to the periodontium via the dentition, and basic mechanics dictate that this masticatory load is balanced by a set of equal and opposite loads that take the form of stresses and strains. When stresses and strains are excessive, they cause tissue damage (Shoham and Gefen 2012); when they are insufficient, the result is tissue atrophy (Pena et al. 2008). Between these extremes, stresses and strains stimulate remodeling of bone, the periodontal ligament (PDL), and the junctional epithelium (JE).
The dynamic nature of these mechanoadaptative responses is essential for maintaining the overall health of the entire periodontium, but how cellular responses are coordinated between the periodontal soft tissues remains an enigma. The JE, the supracrestal fibers, and the PDL are all bound to the tooth and thus experience stresses and strains in response to mastication (Jang et al. 2015; Yuan et al. 2023), but their distinct anatomical locations make it challenging to envision how their cellular responses are synchronized.
We considered one possibility: that these soft tissues possess a “just-in-time” mechanism for dynamically adjusting to the amount of stress/strain applied. A feature of a just-in-time mechanism is that the system exists at a relatively low/inactive state until triggered (e.g., in this case, by increased masticatory loading). In response to high distortional strains produced by hyper-loading, the system could then theoretically switch to an activated state and elicit a highly specific and localized molecular/cellular response at the location of the increased load. This response to the amount of stress/strain would last only as long as needed to compensate for the increased demand. This kind of just-in-time method, if it existed, would provide versatility in terms of cellular dynamics but would also depend on a mechanosensitive assembly (Eckert et al. 2025). Such a mechanosensitive assembly has not been described linking the JE, the supracrestal connective tissue, or the PDL, but we hypothesized that the epithelial rest of Malassez (ERM) network could serve this role.
Since their discovery by Serres (in 1817) and Malassez (1885), there has been a deep interest in the origin, distribution, and possible functions of ERMs (Wesselink and Beertsen 1993; Keinan and Cohen 2013; Xiong et al. 2013). ERMs appear in thin tissue sections as isolated islands of epithelial cells; in reality, the tissue is a 3-dimensional, fenestrated, mesh-like structure (Spouge 1984). This network, along with its extension from the root tip to immediately adjacent to the JE (Grant and Bernick 1969), struck us as ideally suited to sense strains within the tissues and potentially coordinate their responses to changes in mechanical input. The following in silico, in vitro, and in vivo experiments were undertaken to test this hypothesis.
Methods and Materials
All experimental procedures followed ARRIVE (Animal Research: Reporting of In Vivo Experiments) 2.0 guidelines. Please see the Appendix for animal husbandry and care; in vivo hyper- and hypo-loading models; tissue collection, handling, and histological analyses of murine and human tissues; single-cell RNA sequencing; quantitative immunohistochemistry (qIHC); and finite element (FE) modeling.
Results
Mastication produces distortional strains in the PDL and in the supracrestal region, which are elevated by hyper-loading
FE modeling was used to calculate the magnitude of distortional strains in the PDL, the supracrestal region, and the JE under conditions of normal masticatory function and hyper-loading (Xu et al. 2019). The FE model was derived from micro–computed tomography (µCT) data (see Appendix Methods), and material properties were assigned to soft and hard tissues (Nishihira et al. 2003; Fill et al. 2012). Under conditions of normal masticatory load, the PDL experienced a mean distortional strain of 0.55% ± 0.45% (Fig. 1A). Under hyper-loading conditions, the mean distortional strain in the PDL doubled to 1.3% ± 0.45% (Fig. 1B, C).
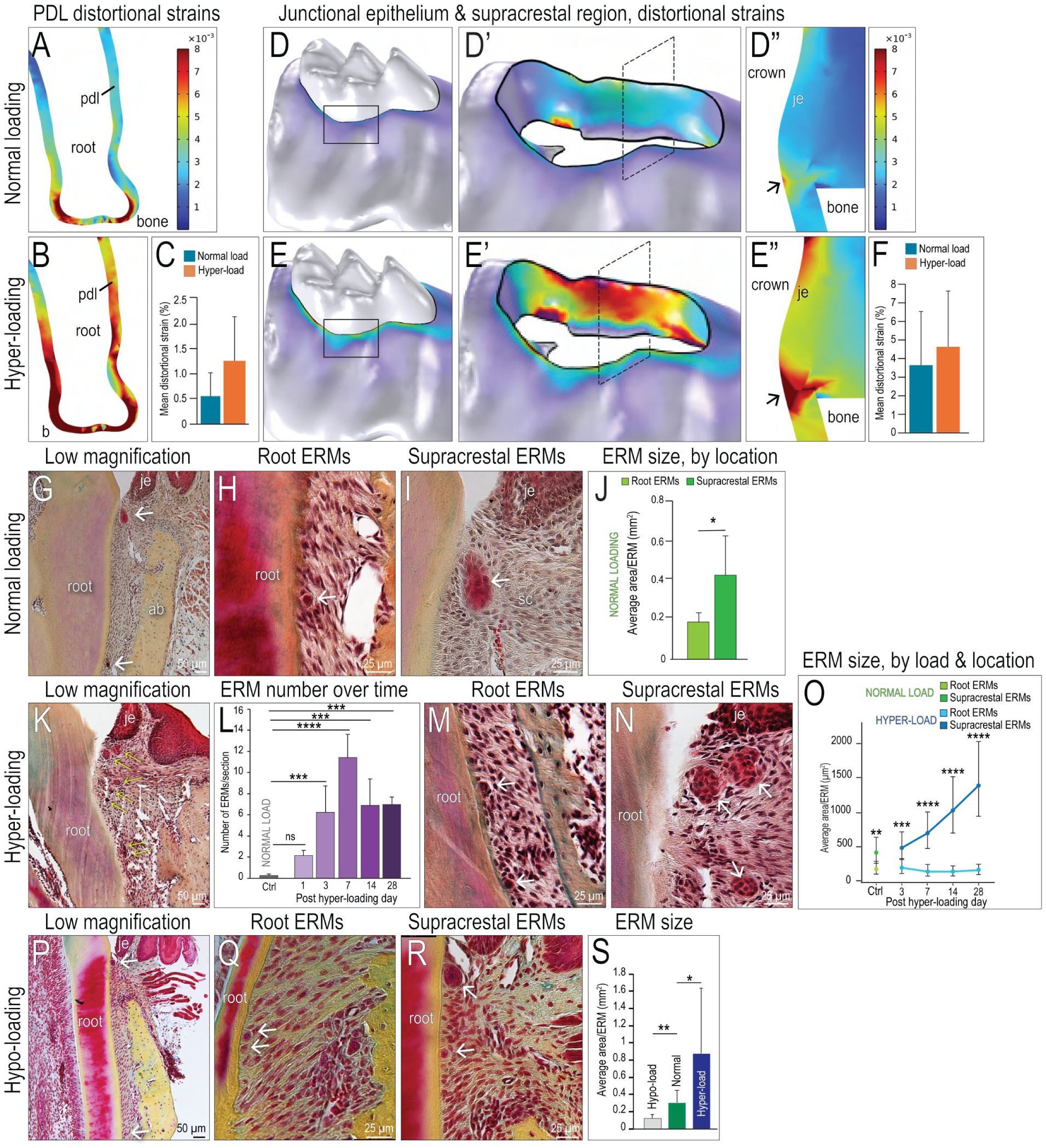
Epithelial rests of Malassez (ERMs) respond to distortional strains in the PDL, the junctional epithelium (JE), and the supracrestal connective tissues. (
By virtue of its hemidesmosomal attachment to the tooth, the JE also experiences distortional strains in response to normal masticatory forces (Yuan et al. 2023). The FE model predicted distortional strains in the JE/supracrestal region that were twice the magnitude experienced by the PDL (Fig. 1D, D′, D′′, F). Hyper-loading led to a mean distortional strain of 4.6% ± 3.0% in the JE/supracrestal region (Fig. 1E, E′, E′′), quadruple that experienced by the hyper-loaded PDL (Fig. 1F). When examined in detail, the highest strain centered on the supracrestal connective tissues (Fig. 1E′, E′′).
ERMs in the Supracrestal Region Are Larger Than ERMs along the Root
The PDL and JE are anatomically distinct structures, but based on their shared attachments to a moving tooth, their responses to hyper-loading would likely be coordinated to provide appropriate tissue-level responses. We hypothesized that the 3-dimensional ERM meshwork was a candidate tissue for this task. ERMs were observed in the PDL space (Fig. 1G, H) and the supracrestal connective tissue, coronal to the alveolar bone crest (Fig. 1G, I), under normal masticatory loading conditions. The size of these anatomically distinct ERMs was quantified (see Methods and Materials), and those in the supracrestal region were consistently, significantly larger than those along the root (green bars, Fig. 1J).
Hyper-loading Triggers the Selective Expansion of Supracrestal ERMs
Larger ERMs were located in the supracrestal region, which experienced higher distortional strains compared to the PDL space (Fig. 1). To test whether the size or number of ERMs was influenced by strain, an in vivo hyper-loading model was employed (see Methods and Materials; Xu et al. 2019). The periodontium of teeth subjected to hyper-loading for 3, 7, 14, and 28 d was compared against baseline data from periodontia subjected to normal masticatory loading (Fig. 1J).
Within 1 d of hyper-loading, more ERMs were detected (yellow arrows, Fig. 1K; quantified in Fig. 1L). The total number of ERMs increased between days 1 and 3, as well as again between days 3 and 7 of hyper-loading (Fig. 1L). After 7 d of hyper-loading, the number of ERMs remained relatively constant and always significantly more than observed around normal loading controls (Fig. 1L).
Changes in ERM size were also observed. Under hyper-loading conditions, supracrestal ERMs were larger than root ERMs (compare Fig. 1M, N), and over the duration of hyper-loading, this difference in size became more pronounced (Fig. 1O). Thus, ERM number and size exhibited a time-dependent responsiveness to masticatory hyper-loading.
Hyper-loading by Itself Does Not Trigger Periodontal Inflammation
Factors other than mechanical force could influence ERM number and/or size. For example, hyper-loading could cause an inflammatory response (Leone et al. 2026), which indirectly affected the number and size of ERMs. To evaluate this possibility, the distribution and number of neutrophils and macrophages in the hyper-loading group were compared to controls subjected to normal masticatory loading. No apparent differences between hyper-loaded and control groups in terms of neutrophil and macrophage number and distribution in either the PDL space or the JE were observed (Appendix Fig. 1).
Hypo-loading Reduces the Size of Supracrestal ERMs
Hyper-loading correlated with an increase in the number and size of ERMs; we next tested whether hypo-loading had the opposite effect. After 4 wk of hypo-loading (see Methods and Materials; Zhang et al. 2019), ERMs along the root surface (Fig. 1P, Q) and ERMs in the supracrestal region (Fig. 1P, R) were significantly smaller compared to both controls and the hyper-loaded group (Fig. 1S). These data further supported the conclusion that the ERM network was responsive to changes in masticatory forces.
ERMs Are Mechanoresponsive
Do ERMs possess the molecular machinery to detect a physical force and convert it into a biochemical signal? Human ERMs expressed the mechanically gated ion channel Piezo1 (Fig. 2A); single-cell RNA sequencing (scRNA-seq) data confirmed expression of Piezo1 in human ERMs and in JEs (Fig. 2B). The mechanosensitive adhesion protein β4 integrin (Fig. 2C, D), the mechanosensitive linker protein plectin (Fig. 2E, F), and the mechanoresponsive ion channel TRPV4 (Fig. 2G, H) were also detected in human ERMs and in the JE.
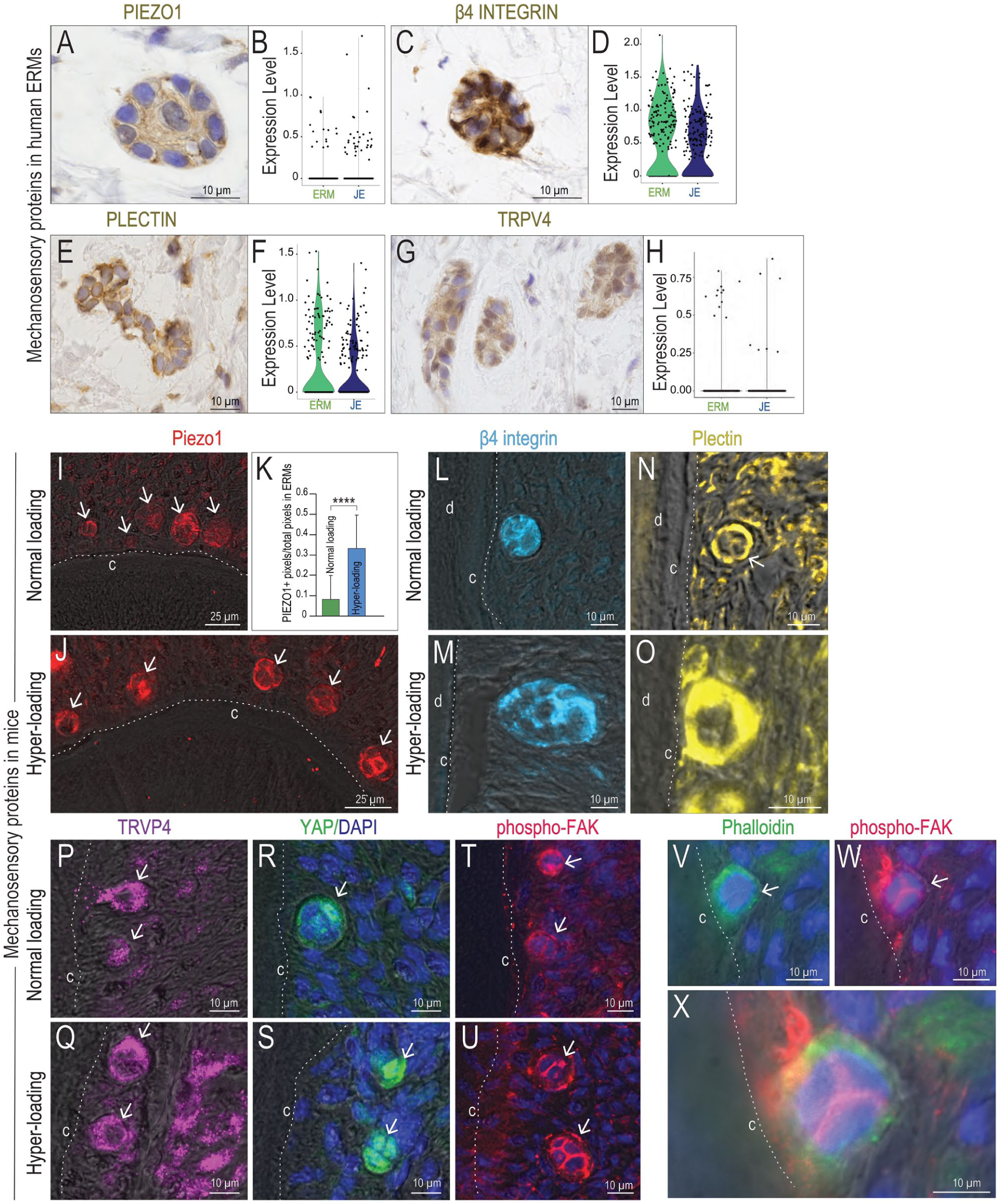
Epithelial rests of Malassez (ERMs) express mechanosensitive and mechanoresponsive proteins. Human ERMs express (
Murine ERMs also expressed Piezo1 (Fig. 2I), and its expression was significantly elevated in load-activated ERMs (Fig. 2J, K). The mechanosensitive proteins β4 integrin (Fig. 2L, M) and plectin (Fig. 2N, O), as well as TRPV4 (Fig. 2P, Q) and YAP (Fig. 2R, S), were all expressed in ERMs under both normal and hyper-loading conditions, with expression being consistently higher in ERMs induced by hyper-loading.
Of particular note was the pattern of phosphorylated focal adhesion kinase (phospho-FAK) expression in ERMs under normal and hyper-loading conditions (Fig. 2T, U). Phalloidin staining identified F-actin+ microfilaments (Fig. 2V), and colocalization with phospho-FAK (Fig. 2V, W, X) provided strong evidence that ERMs were involved in transducing mechanical signals into biochemical signals.
ERMs Initiate Matrix Remodeling in Response to Hyper-loading
Thus far, our data demonstrated that ERMs possess the molecular machinery to sense and respond to mechanical forces; we postulated these forces were transmitted to ERMs via surrounding connective tissue fibers. Transmission electron microcopy illustrated the proximity of human ERMs to collagen fibers (Fig. 3A) and to oxytalan fibers (Fig. 3B). Histological staining for oxytalan fibers illustrated their close association with ERMs (Fig. 3C). βIII tubulin+ neurons encircled ERMs (red arrow, Fig. 3D). Expression of matrix metalloproteinase 13 (MMP13) in ERMs (Fig. 3E) suggested their local extracellular matrix was prone to remodeling.
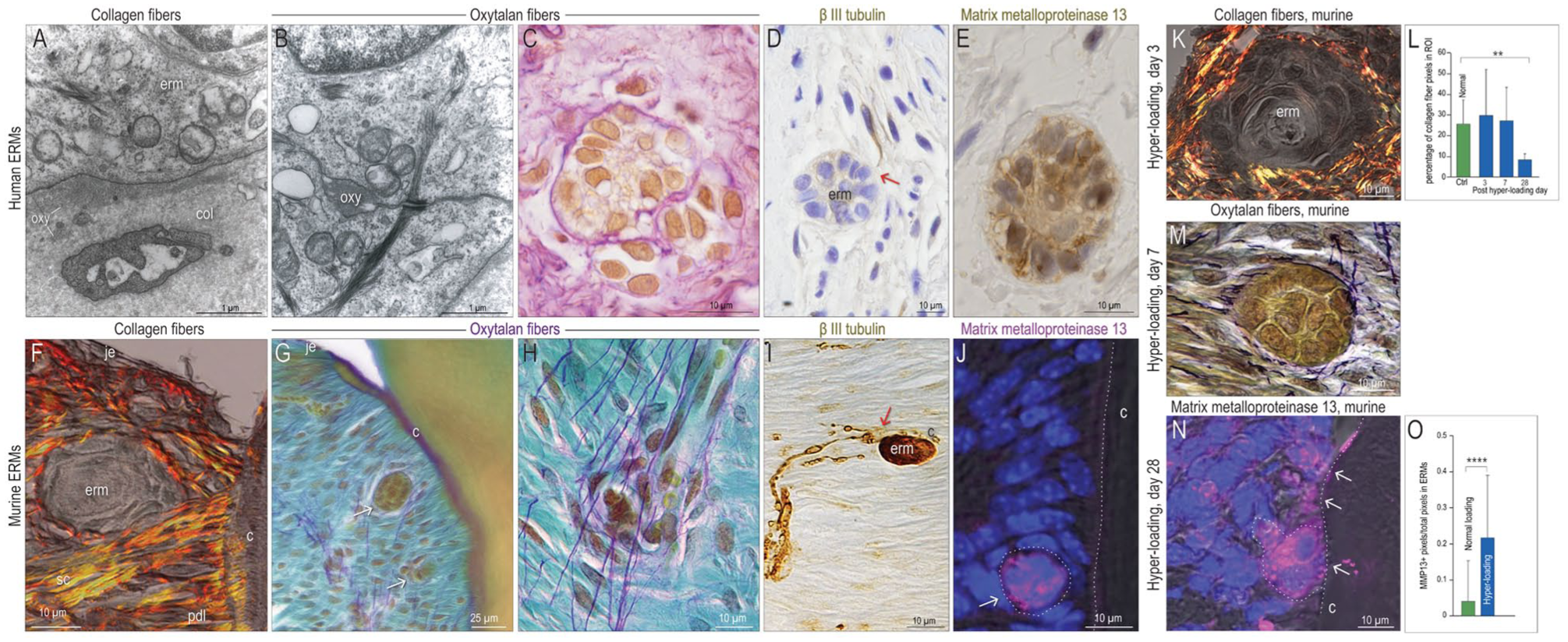
Remodeling of the extracellular matrix around epithelial rests of Malassez (ERMs) is stimulated by hyper-loading. Representative transmission electron microscopic images showing (
Similar relationships were noted between murine ERMs and picrosirius red–stained collagen fibers (Fig. 3F) and purple-stained oxytalan fibers, which are typically oriented perpendicular to collagen fibers (Fig. 3G, H). βIII tubulin+ neurons were in close proximity to murine ERMs (red arrow, Fig. 3I). Murine ERMs were also immunopositive for MMP13 (Fig. 3J).
Hyper-loading in the mouse model resulted in apparent collagen fiber micro-ruptures (Fig. 3K, Appendix Fig. 2); collagen content analyses indicated that continued hyper-loading was detrimental to fiber density (Fig. 3L). Fragmented oxytalan fibers were also evident around load-activated ERMs (Fig. 3M, Appendix Fig. 2). Load-induced damage to the extracellular matrix correlated with significantly increased expression of MMP13 in and around murine ERMs (Fig. 3N, O).
ERMs Release Wnt Signals That Influence the Mechanoresponsiveness of the PDL
To coordinate mechanosensitive responses in the PDL, supracrestal connective tissue, and JE, we speculated that load-activated ERMs released signaling molecules that could locally influence cellular behavior. The following lines of evidence implicated Wnt/β-catenin signaling in this process.
The first line of evidence came from lineage-tracing the fates of Wnt-responsive cells in ERMs. Tamoxifen was delivered to the Wnt lineage tracer strain Axin2CreERT2;R26RmTmG/+. Three days later, small Keratin5+ ERMs, some of which costained with green fluorescent protein (GFP), were detected (Fig. 4A–A′′). Very few PCNA+ cells were detectable in the PDL (Appendix Fig. 3), indicating that under normal loading conditions, Wnt-responsive ERMs were generally quiescent.
The second line of evidence came from fate-mapping Wnt-responsive cells under hyper-loading conditions. Tamoxifen was delivered to Axin2CreERT2;R26RmTmG/+ mice, and immediately thereafter, hyper-loading was initiated. After 3 d, Keratin5+ ERMs were significantly larger (arrows, Fig. 4B; quantified in Fig. 1), and the number of Wnt-responsive cells increased (Fig. 4B′, B′′), which corresponded with an increase in PCNA+ cells in the PDL (Appendix Fig. 3). Most Wnt-responsive cells were positioned adjacent to the cementum, and some colocalized with the Keratin5+ ERMs (Fig. 4B–B′′).
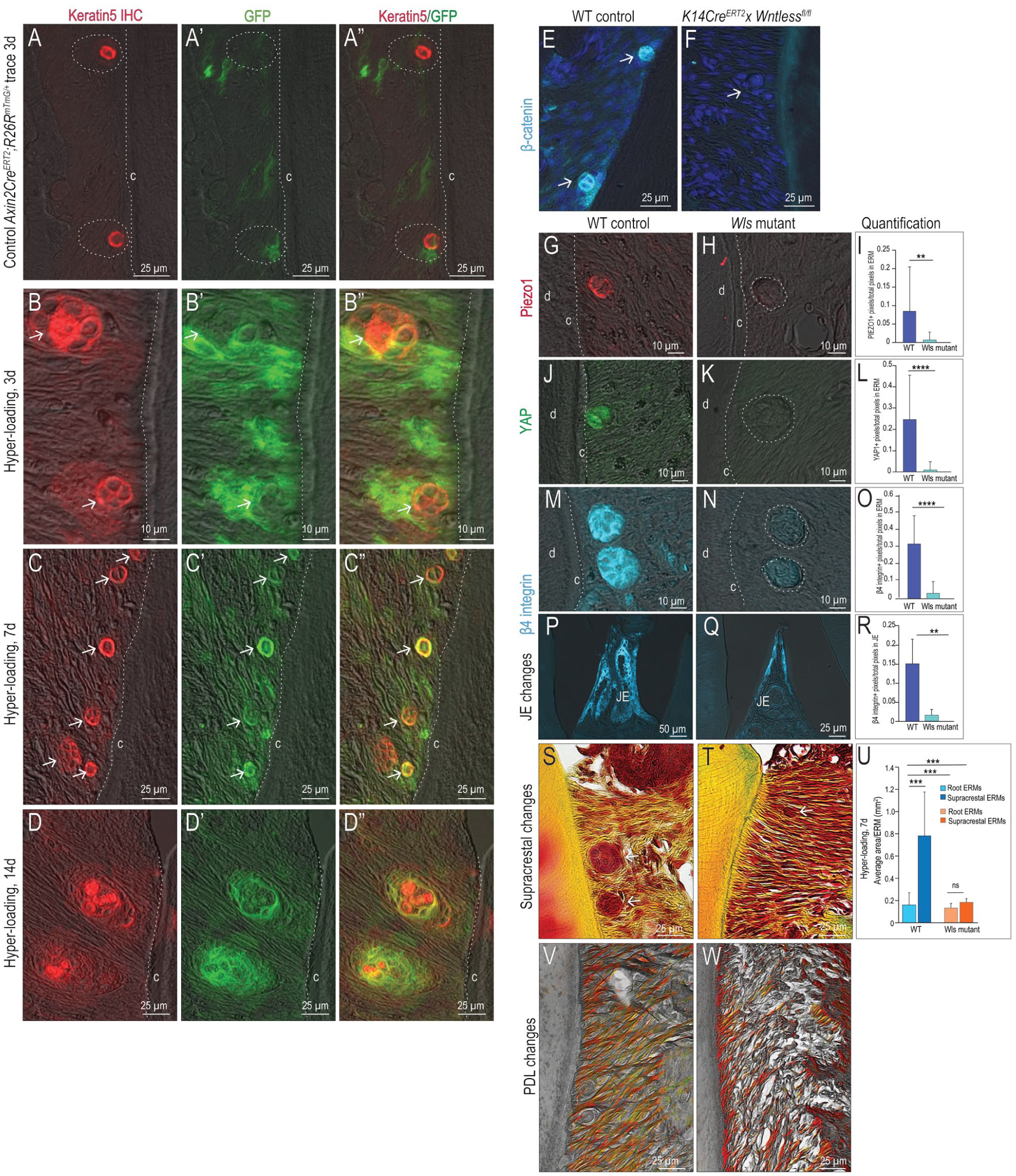
Supracrestal epithelial rests of Malassez (ERMs) adjacent to the junctional epithelium (JE) show the most significant enlargement in response to hyper-loading. Axin2CreERT2;R26RmTmG/+ mice received tamoxifen and 3 days later immunostaining for (
After 7 d of hyper-loading, the number and size of ERMs continued to significantly increase (arrows, Fig. 4C–C′′; quantified in Fig. 1); load-activated ERMs maintained their Wnt-responsive status (Fig. 4C–C′′). GFP+ Wnt-responsive cells remained associated with the enlarged Keratin5+ ERMs throughout longer periods of hyper-loading (Fig. 4D–D′′). Together, these data strongly suggested that Wnt signaling was involved in ERM responses to hyper-loading.
To directly test whether Wnt signals emanated from ERMs, a K14Cre driver was crossed with a Wntlessfl/fl strain. GFP immunostaining in K14CreERT2;R26RmTmG/+ mice confirmed tissue specificity of the Cre to ERMs (Appendix Fig. 4) and the JE, and then K14CreERT2;Wlsfl/fl mice were generated. The chaperone protein Wntless (Wls) is essential for Wnt protein secretion (Banziger et al. 2006), and immunostaining for β-catenin confirmed that canonical Wnt signaling was inhibited in ERMs of K14CreERT2;Wlsfl/fl mice (compare Fig. 4E, F).
A third line of evidence that Wnt/β-catenin was involved in ERM signaling came from molecular analyses. Seven days after tamoxifen delivery, expression of Piezo1 (Fig. 4G–I) and YAP (Fig. 4J–L) was lost in Wls mutant ERMs. β4 integrin expression was absent in Wls mutant ERMs and in Wls mutant JEs (compare Fig. 4M, N and 4O, P; quantified in Fig. 4Q, R). These data demonstrated that Wnt/β-catenin signaling was required for ERM mechanosensitivity, as well as JE attachment.
Our fourth line of evidence supporting an essential role for Wnt/β-catenin signaling in ERM mechanotransduction came from analyses of Wls mutant mice after initiating hyper-loading. Normally, supracrestal ERMs enlarge in response to hyper-loading; in Wls mutants, these supracrestal ERMs did not expand (compare Fig. 4S, T; quantified in Fig. 4U). In addition to defects in the JE and supracrestal region, PDL fibers deteriorated. Picrosirius red–stained collagen fibers in the PDL of control mice exhibited their characteristically dense, linear, elongated morphology (Fig. 4V). In Wls mutants, PDL fibers were sparse, fragmented, and truncated (Fig. 4W).
Discussion
Functionless Relics or an Essential Mechanosensory Element in the Periodontium?
The vestige of (Hertwig’s) epithelial root sheath, ERMs emerge during tooth development and endure for the lifetime of most mammals (Reitan 1961). Embryonic tissues are equipped to remove redundant cells (Abud 2004); consequently, this persistence of an ERM network argues against them being simply “developmental jetsam” (Cohen 1976). We started this study by asking, Why does this ERM network persist? And what are the circumstances that provoke ERM reactivation?
The ERM Network Is Ideally Positioned to Detect and Respond to Mechanical Input
Our hypothesis was that ERMs function as a mechanotransducing network, which is supported by 4 lines of evidence. First, ERMs are embedded in a mechanoresponsive extracellular matrix (ECM). ERMs in supracrestal connective tissues are encircled by collagen and oxytalan fibers, and because the JE and supracrestal fibers are firmly attached to the tooth surface, these tissues, along with supracrestal ERMs, experience stresses and strains created by masticatory loading (Fig. 1). The same relationship holds true in the PDL space: because of its attachment to the tooth surface, the PDL and root ERMs experience stresses and strains in response to masticatory forces (Fig. 1). Thus, the ERM network appears to be ideally positioned to detect and respond to changes in mechanical loading of the dentition.
ERMs Are Responsive to Changes in Masticatory Load
Hyper-loading of the dentition triggers an increase in the size and number of ERMs; conversely, hypo-loading results in a significant reduction in ERM size (Fig. 1). This change in size is most significant in the supracrestal region, where distortional strains are twice as high as in the PDL (Fig. 1). These experiments demonstrate a correlation between mechanical loading and ERM biology.
Hyper-loading is not the sole stimulus that leads to ERM activation. ERM “reactivity,” especially in the supracrestal region, has been observed after severing supracrestal fibers in a gingivectomy procedure (Ramfjord et al. 1966), after orthodontic tooth movement (Gilhuus-Moe and Kvam 1972; Talic et al. 2003), after ligature-induced periodontitis (Manisagian et al. 2018), and in a variety of genetic models of periodontal disease (Sojod et al. 2017; Wang et al. 2018). A shared feature of all these conditions is that they disrupt the mechanical integrity of the periodontium. Thus, our proposed function for the ERM as a mechanosensory network aligns with ERM activation in these other models of injury and/or disease.
ERMs Possess the Molecular Machinery for Mechanotransduction
The third line of evidence supporting our hypothesis that ERMs function as a mechanotransducing network is the fact that ERMs express multiple proteins with well-described mechanosensitive and mechanoresponsive functions (Fig. 2). ERM-specific expression of mechano-sensitive and mechano-responsive proteins significantly increases in response to hyper-loading (Fig. 2). The fact that hyper-loading led to increased expression of the mechanosensory proteins further suggests that ERMs adjust this molecular machinery to adapt to a changing mechanical environment, perhaps via a positive feedback loop. Other epithelial tissues harbor such a positive feedback loop involving Piezo1 and Wnt/β-catenin signaling (He et al. 2024).
ERMs Function as an Essential Wnt-Dependent Mechanotransducive Network
Several studies confirm that Wnt signaling is directly regulated by mechanical cues (Przybyla et al. 2016; Zhou et al. 2020; Li et al. 2021; Oak et al. 2025), and our fourth line of evidence supporting a mechanoresponsive function for ERMs came from studies involving manipulation of Wnt/β-catenin signaling in ERMs. Fate-mapping experiments demonstrated that ERMs and, significantly, load-induced ERMs are Wnt-responsive (Fig. 4).
The essential role of Wnt signaling in ERM mechanotransduction was demonstrated by genetically deleting Wls from K14-expressing cells. This strategy resulted in loss of mechanosensitive protein expression in ERMs (Fig. 4). ERMs typically expand in response to hyper-loading (Fig. 1), but Wnt-deficient supracrestal ERMs failed to enlarge, demonstrating that they were no longer sensitive to mechanical input (Fig. 4). In the Wls mutant, PDL fibers were thin, short, and sparse, and the JE began to lose its attachment (Fig. 4). Together, these data support our conclusion that the ERM network functions as a Wnt-dependent mechanosensor.
Limitations of the Current Study
In this study, we integrated analyses conducted in human tissues with analyses of murine tissues, and one might legitimately ask whether tissues from 2 species should be compared. There are multiple variables to consider: for example, it was not possible to ascertain if patients from which ERMs were collected had normal masticatory function or not, whereas a mouse model permitted control over variables such as masticatory loading. There is also the issue of age: in humans, ERM numbers change with age (Simpson 1965), and in our study, murine age was kept constant. Consequently, we are comparing not only different species but also tissues from different life stages. Further, human samples come from a genetically diverse population, whereas our murine population is far more genetically homogeneous.
Despite these and other differences, there are 2 major justifications for undertaking such comparisons. First, in both species, ERMs share a developmental origin (e.g., Hertwig’s Epithelial Root Sheath [ERS]). This shared origin suggests a conserved biological identity and, potentially, conserved functions. Second, the signaling pathways controlling tooth development and periodontal homeostasis (e.g., Wnt, Notch, Shh, BMP) are highly conserved across mammals; consequently, it is reasonable to presume that the mechanisms controlling ERM quiescence/activation are substantially the same in both species. Finally, human tissues allow us to make observations about ERMs, from which we can make hypotheses that are testable within a mouse model. If our experimental data support a particular hypothesis, we may then be able to formulate therapeutic strategies based on those animal models that have potential value in treating human disease.
Conclusions
ERMs are positioned in the right place and at the right time to sense distortional strains produced by masticatory loading. ERMs possess the molecular machinery to sense and respond to masticatory loading, altering their size and number in response to hyper-loading. ERMs use a Wnt-dependent mechanism to coordinate tissue-level responses to physical forces delivered by mastication. Together, these data support the conclusion that the ERM meshwork functions as a mechanosensory organ and is a critical part of how periodontal tissues cope with functional changes in their mechanical environment.
Author Contributions
B. Liu, J.B. Brunski, F. Hermans, contributed to conception and design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; I. Lambrichts, J.A. Helms, contributed to conception and design, data analysis, and interpretation, drafted and critically revised the manuscript; E. Chang, M.A. Hakeam, M.M. Torabi, F. Aellos, P.C. Ochweri, I. Evens, M.R. James, C. Huang, contributed to data acquisition, analysis, and interpretation, critically revised the manuscript. All authors provided final approval and agreed to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345261455868 – Supplemental material for Epithelial Rests of Malassez Form a Wnt-Dependent Mechanoresponsive Network
Supplemental material, sj-docx-1-jdr-10.1177_00220345261455868 for Epithelial Rests of Malassez Form a Wnt-Dependent Mechanoresponsive Network by B. Liu, I. Lambrichts, E. Chang, M.M. Torabi, F. Aellos, P.C. Ochweri, I. Evens, M.R. James, M.A. Hakeam, C. Huang, J.B. Brunski, F. Hermans and J.A. Helms in Journal of Dental Research
Supplemental Material
sj-docx-2-jdr-10.1177_00220345261455868 – Supplemental material for Epithelial Rests of Malassez Form a Wnt-Dependent Mechanoresponsive Network
Supplemental material, sj-docx-2-jdr-10.1177_00220345261455868 for Epithelial Rests of Malassez Form a Wnt-Dependent Mechanoresponsive Network by B. Liu, I. Lambrichts, E. Chang, M.M. Torabi, F. Aellos, P.C. Ochweri, I. Evens, M.R. James, M.A. Hakeam, C. Huang, J.B. Brunski, F. Hermans and J.A. Helms in Journal of Dental Research
Footnotes
Acknowledgements
The authors thank the anonymous reviewers selected by JDR to evaluate this work. Their expertise, along with their queries and criticisms, reflected deep knowledge and insight into the biology of ERMs. Their questions prompted us to undertake a number of additional experiments that ultimately improved this article, and for that, we are grateful.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: F.H. was supported by the Fund for Scientific Research (FWO)—Flanders (Belgium) (1226325N to F.H.). I.L. and F.H. were supported by the FWO (G0A2E26N). B.L., M.M.T., and J.A.H. were supported by NIH 1R01DE031270-01A1 and NIH 1R25DE03355901A1 to J.A.H. Computational resources for all single-cell transcriptome analyses were provided by the Vlaams Supercomputer Centrum (VSC), managed by the FWO.
Data Availability
The data and materials that support the findings of this study are publicly available. Histological and immunohistochemical images are deposited in the Stanford Digital Repository (DOI: ![]() ). Gene expression analysis in human ERM was performed using publicly available human PDL data (GEO accession number GSE161266).
). Gene expression analysis in human ERM was performed using publicly available human PDL data (GEO accession number GSE161266).
A supplemental appendix to this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.