
Abstract
Odontoblasts are a tooth-specific cell type responsible for dentin formation. During odontoblast differentiation, dramatic organelle alterations occur with the increase of both mitochondria and endoplasmic reticulum (ER). In multiple cell types, mitochondria–ER contact sites (MERCs) are formed to regulate cellular biological processes via mediating essential interorganelle communications. However, whether MERCs play a role in odontoblast differentiation remains enigmatic. Here, we found a progressive increase of MERCs during the in vitro odontoblastic differentiation using live-cell imaging, transmission electron microscopy (TEM), and in situ proximity ligation assay (PLA). Meanwhile, in situ PLA verified the in vivo elevation of MERCs in the odontoblasts of mouse teeth. Remarkably, the disruption of MERCs via overexpression of FATE1, a protein that increases the distance between mitochondria and ER, suppressed the expression of key odontoblast markers, alkaline phosphatase (ALP) activity, and mineralized nodule formation in the mouse dental papilla cells (mDPCs) after differentiation induction, suggesting a positive role of MERCs for odontoblastic differentiation. Among all MERC-associated genes, Mfn2 expression is significantly increased in the RNA sequencing data of odontoblast-like cells versus undifferentiated mDPCs. Further experiments showed that knockdown of Mfn2 resulted in diminished MERC formation as well as impaired odontoblastic differentiation, as evidenced by the downregulated expression of odontoblast markers, compromised ALP activity, and mineralized nodule formation. The impaired odontoblastic differentiation upon Mfn2 knockdown was rescued by the overexpression of Linker, an enhancer of MERC formation, suggesting that MFN2 enhances MERC formation to promote odontoblastic differentiation. Consistently, in vivo knockdown of Mfn2 also impaired MERC formation as well as odontoblast differentiation and dentinogenesis, which were restored by in vivo Linker overexpression. Taken together, these findings indicate that MERCs are dynamically increased and essential for odontoblast differentiation and dentinogenesis with MFN2 as an essential mediator, which provides an important mechanism of organelle interaction orchestrating odontoblast differentiation.
Keywords
Introduction
Dentin constitutes the primary mineralized tissue within teeth. It is produced by odontoblasts, a specialized cell type differentiating from dental mesenchymal cells (Chai et al 2000; Hall and Gillis 2013). Odontoblasts synthesize and discharge collagenous and noncollagenous proteins to form predentin, which subsequently undergoes mineralization to form mature dentin (Butler 1995; Goldberg et al 2011; Fu et al 2021; Niu et al 2025).
During the process of odontoblast differentiation, the cells undergo remarkable morphological and functional transformations. They evolve from a spindle shape to a tall columnar morphology with increased organelles including mitochondria and endoplasmic reticulum (ER), which facilitate the synthesis and secretion of dentin matrix ingredients (Couve 1986; Couve and Schmachtenberg 2011). Mitochondria, acting as the cellular powerhouses, are involved in regulating metabolic pathways and dynamic remodeling processes. Their functional integrity is critical for the high bioenergetic and biosynthetic demands of dentin matrix production (Matsuishi et al 2018; Zhang et al 2018). For ER, they serve as the central hub for protein synthesis, folding, and modification. ER-mediated regulatory mechanisms, including stress response pathways, have been reported to participate in odontoblast differentiation during dentin formation (Chen et al 2018; Aryal et al 2019; Chen et al 2021).
Recently, emerging evidence indicates that organelles form dynamic interaction networks to cooperatively regulate cellular functions (Arruda et al 2014; Wu et al 2018; Jang et al 2022). Notably, mitochondria and ER physically and functionally interact through specialized membrane contact sites known as mitochondria–ER contacts (MERCs) (de Brito and Scorrano 2010). These structures, mediated by tethering complexes such as VDAC1-IP3R1, facilitate the bidirectional regulation of organelle functions by enabling lipid transfer, calcium signaling, and metabolic coordination. Aberrations in MERCs can lead to the emergence of various diseases, such as neurodegenerative diseases, cardiovascular diseases, and metabolic diseases (Voeltz et al 2006; Magalhães Rebelo et al 2020; Ziegler et al 2021). Nevertheless, it remains unclear whether organelle interactions, particularly MERCs, exist in the odontoblasts. If present, the key involved proteins and their functional roles are still unknown.
In this study, we found the dynamic presence of MERCs during odontoblast differentiation and identified MFN2 as a key regulatory protein. Increased MFN2 expression during odontoblastic differentiation promoted the formation of MERCs, the disruption of which led to abnormalities in odontoblast differentiation and dentin formation.
Materials and Methods
For complete Materials and Methods, please refer to the Appendix.
Plasmid and Small Interfering RNA Transfection
The small interfering RNAs (siRNAs) of Mfn2 were purchased from Genepharma (Appendix Table 1). The plasmids pcDNA3.1-mRFP-Linker and pcDNA3.1-EGFP-FATE1 were purchased from Omega Bio-tek, and their sequences have been previously described (Csordás et al 2006; Doghman-Bouguerra et al 2016). Cell transfection was conducted with Lipofectamine 3000 (Invitrogen) in accordance with the manufacturer’s protocol.
Animals and Cell Culture
All experimental procedures were approved by the Animal Welfare and Ethics Committee of School and Hospital of Stomatology at Wuhan University (approval No. S07924120F), complying with the ARRIVE 2.0 checklist (Animal Research: Reporting of In Vivo Experiments). Mice were obtained from the Hubei Provincial Center for Disease Control and Prevention (Appendix Table 2).
Primary mouse dental papilla cells (mDPCs) harvested from the first molars of mice on embryonic day 16.5 (E16.5) were routinely maintained in DMEM (Hyclone) containing 10% fetal bovine serum (Gibco) and 1% penicillin/streptomycin (Hyclone) at 37 °C under 5% CO2. For odontoblastic differentiation, mDPCs were incubated in differentiation medium supplemented with 50 µg/mL ascorbic acid (Sigma-Aldrich), 10 mmol/L β-glycerophosphate (Sigma-Aldrich), and 10 nmol/L dexamethasone (Sigma-Aldrich).
For in vivo experiments, adeno-associated virus serotype 6 (AAV6) was procured from BrainVTA Biotechnology, and chemically modified in vivo siRNAs designed for enhanced stability and delivery efficiency in living animals were purchased from Genepharma. Injections were performed as previously described (Fu et al 2023). Tissues were harvested at 2 and 3 wk for analysis (Zheng et al 2022).
In Situ Proximity Ligation Assay
MERCs were evaluated using in situ proximity ligation assay (PLA) (Sigma-Aldrich) targeting VDAC1 on the outer mitochondrial membrane and IP3R1 on the ER membrane with anti-VDAC1 (1:200, Proteintech) and anti-IP3R1 (1:100, GeneTex) antibodies. PLA signals representing VDAC1-IP3R1 proximity were imaged with a Leica TCS SP8 confocal microscope (Leica Microsystems) and subsequently analyzed with ImageJ (National Institutes of Health).
Statistical Analysis
Data are presented as mean ± standard deviation (SD). For 2-group comparisons, 2-tailed Student’s t tests were used, and for comparisons among multiple groups, we used analysis of variance (ANOVA; 1 way or 2 way) to assess statistical significances. We identified P < 0.05 as statistically significant (*P < 0.05, **P < 0.01).
Results
MERCs Increase during Odontoblastic Differentiation
To observe whether MERCs play a role during the differentiation of odontoblasts, mitochondria, ER, and MERCs during in vitro odontoblastic differentiation of mDPCs were quantified using Mito-Tracker and ER-Tracker live-cell imaging. Interestingly, the results showed elevated mitochondria, ER, as well as mitochondria–ER colocalization during this process (Fig 1A–A′″). Transmission electron microscopy (TEM) also demonstrated elevated mitochondrial–ER juxtaposition (≦50 nm) (Fig 1B, B′). In addition, in situ PLA was performed using antibodies against VDAC1 and IP3R1, which are a ubiquitous tether of MERCs (Ziegler et al 2021). The results showed increased VDAC1-IP3R1 proximity with the odontoblastic differentiation of mDPCs (Fig 1C, C′). No positive signal was found in the negative control of in situ PLA (Appendix Fig 1). Consistently, in situ PLA using sections of postnatal day 4 (PN4) molars also revealed increased in vivo VDAC1-IP3R1 puncta as cells differentiated from preodontoblasts to odontoblasts (Fig 1D). Overall, our data indicate that the formation of MERCs increases during odontoblast differentiation.
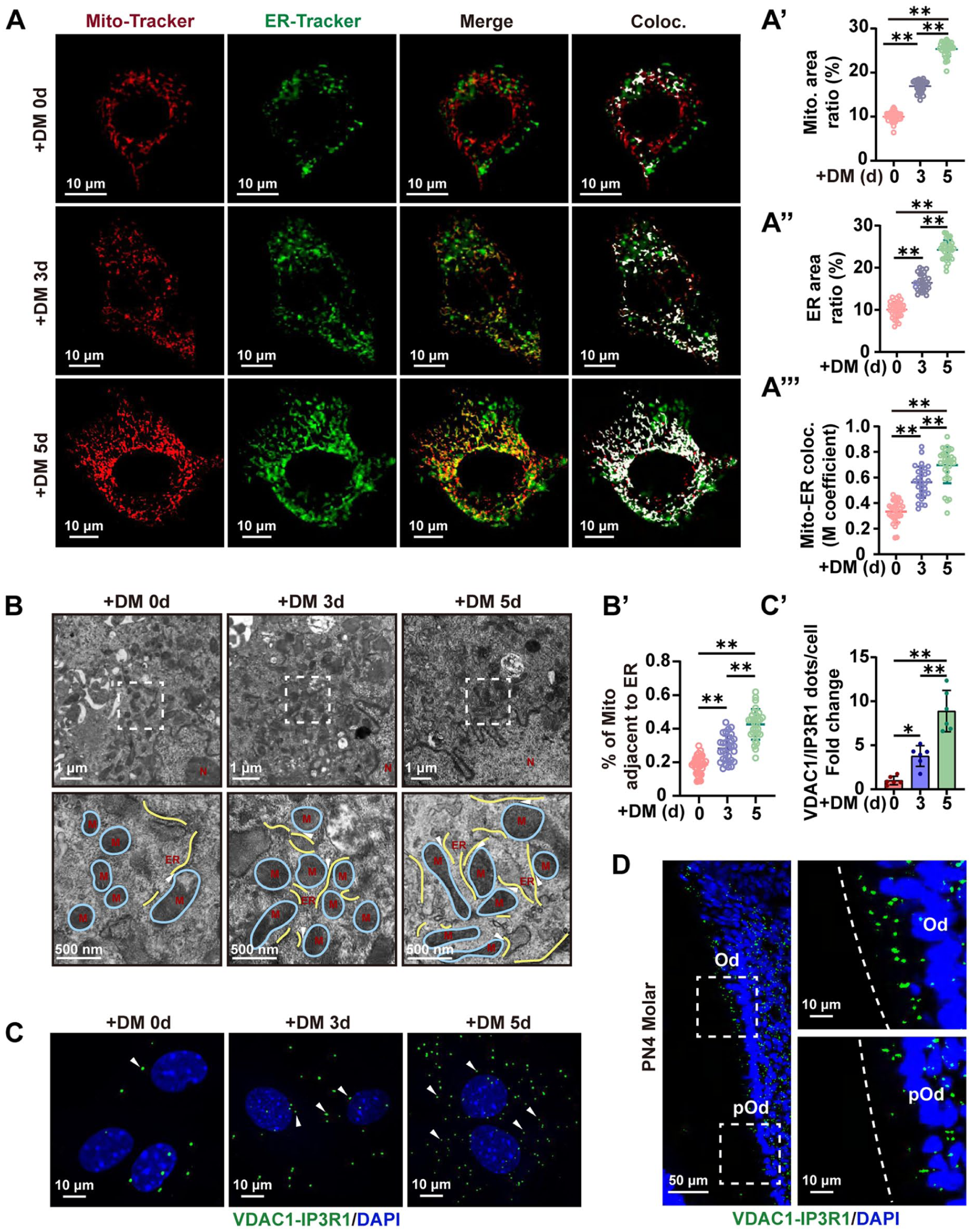
MERCs increase during odontoblastic differentiation. (
Disruption of MERCs Inhibits Odontoblastic Differentiation
To further reveal the function of MERCs during odontoblastic differentiation, a pcDNA3.1-EGFP-FATE1 plasmid was obtained for FATE1 overexpression. Overexpressed FATE1 localized at the interface between ER and mitochondria can increase the distance between mitochondria and ER (Doghman-Bouguerra et al 2016), resulting in the uncoupling of MERCs (Fig 2A). Endogenous FATE1 was absent in mDPCs, and the presence of EGFP and immunoblotting of FATE1 confirmed the ectopic expression of FATE1 in mDPCs after transfection of pcDNA3.1-EGFP-FATE1 plasmid (Fig 2B and C, Appendix Figs 2, 3). TEM analysis verified that FATE1 overexpression attenuated the close apposition between mitochondria and ER in mDPCs after differentiation induction for 3 d (Fig 2D, D′). Reverse transcription quantitative polymerase chain reaction (RT-qPCR) results showed that the mRNA levels of the odontoblast marker genes Col1a1, Dspp, and Dmp1 were obviously decreased in the odontoblastic cells with FATE1 overexpression (Fig 2E). Meanwhile, Western blot analysis confirmed marked decreased protein abundance of COL I, DSPP, and DMP1 in FATE1-overexpressing cells (Fig 2F, F′). In addition, alkaline phosphatase (ALP) staining and Alizarin Red S (ARS) staining demonstrated that FATE1 overexpression brought about a reduction in both ALP activity and mineralized nodule formation capacity (Fig 2G–H′). Thus, MERCs have a positive function in the odontoblastic differentiation of mDPCs.
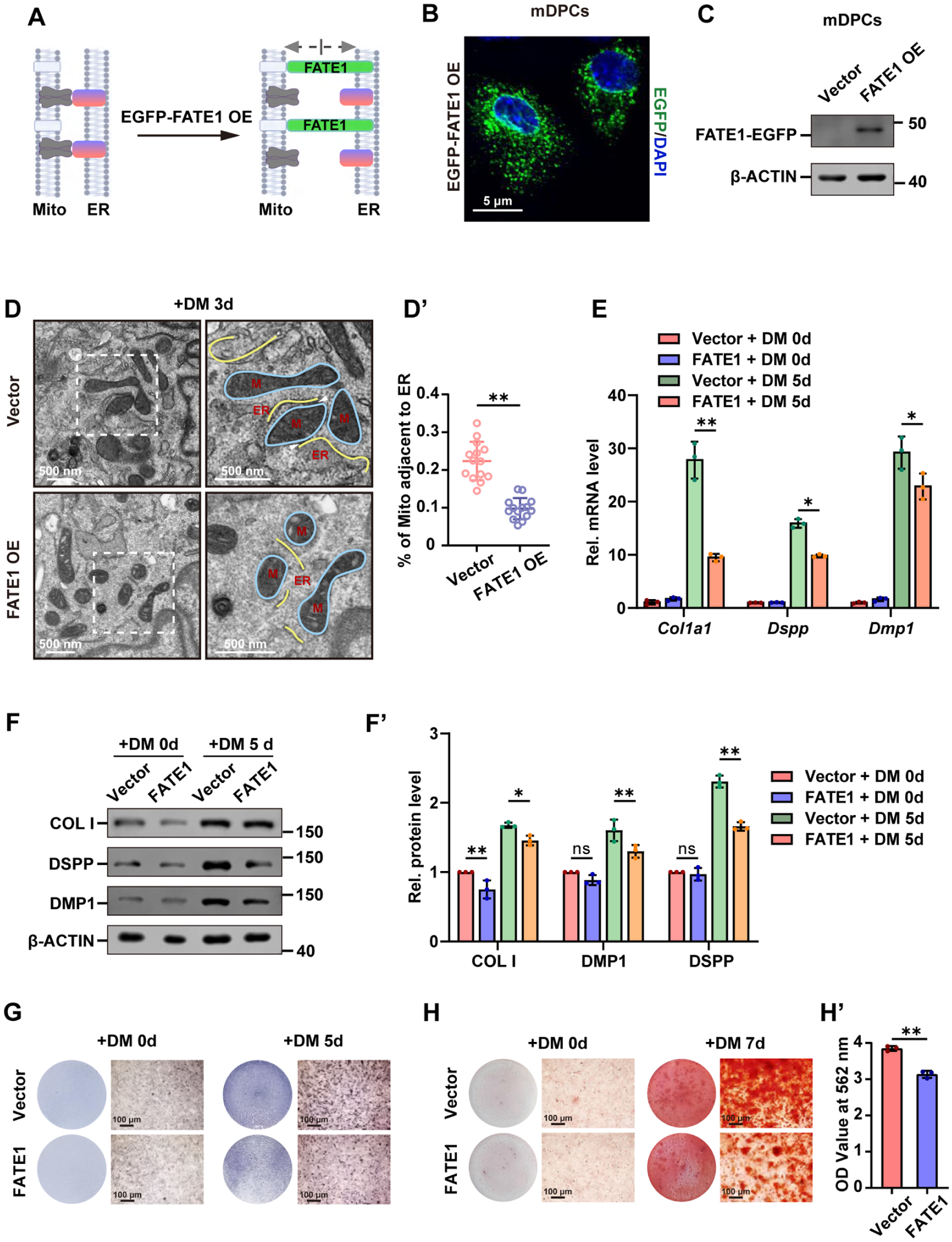
Disruption of MERCs inhibits odontoblastic differentiation. (
Mitofusin2 (MFN2) Expression Exhibits an Upward Trend during Odontoblast Differentiation
To elucidate the underlying mechanisms of MERC formation during odontoblastic differentiation, the known MERC-associated genes (Thoudam et al 2023) were overlapped with the differentially expressed genes (DEGs) from the RNA sequencing data (CRA003233) of mDPCs after differentiation induction for 11 d versus those without differentiation induction (Lin et al 2021). Eighteen DEGs were identified, with Mfn2 being one of the most obviously upregulated genes (Fig 3A–B, Appendix Table 3). To investigate whether MFN2 was involved in the formation of MERCs and thereby affected the differentiation of odontoblasts, we characterized its expression pattern during the odontoblastic differentiation of mDPCs using RT-qPCR, Western blot analysis, and immunofluorescence (IF). The results showed that both the mRNA and protein levels of MFN2 increased progressively during this process (Fig 3C–E′, Appendix Fig 4A). To further investigate the in vivo expression pattern of MFN2, IF was conducted on sections of PN2 incisors, which allows for the visualization of odontoblasts across various stages of differentiation (Zheng et al 2023). The results showed that MFN2 expression was very weak in the undifferentiated dental mesenchymal cells but was moderate in the preodontoblasts and high in the odontoblasts (Fig 3F–I, Appendix Fig 4B). Meanwhile, IF of mouse molars at developmental stages from PN2 to PN10 verified strong MFN2 signals in the differentiated odontoblasts (Fig 3J–L, Appendix Fig 4C–E). Thus, the expression of MFN2 displays an increased tendency during odontoblast differentiation.
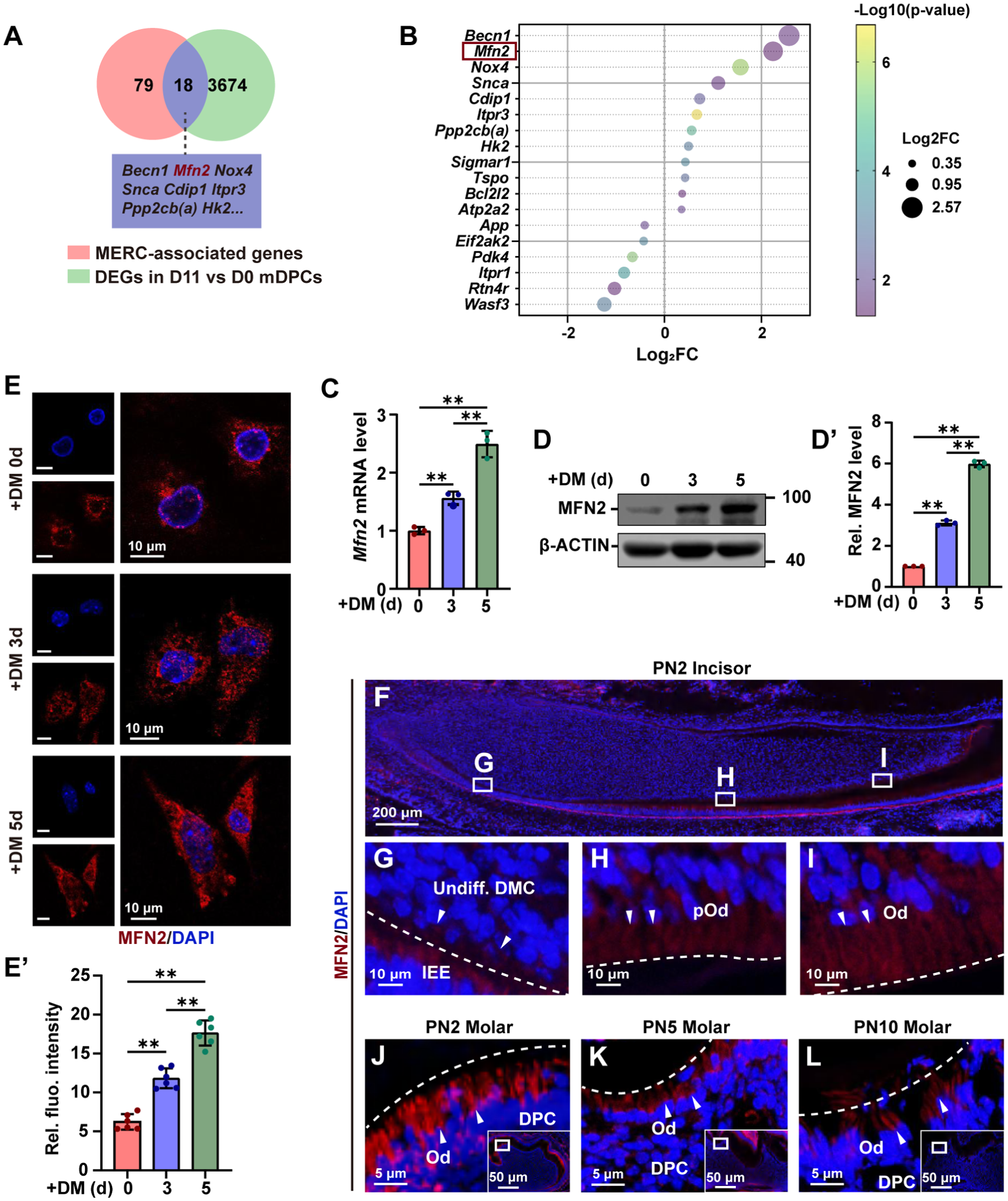
MFN2 expression exhibits an upward trend during odontoblastic differentiation. (
MFN2 Promotes MERC Formation and Odontoblastic Differentiation In Vitro
To verify if MFN2 modifies the formation of MERCs, Mfn2 was knocked down in mDPCs with siRNAs, causing an approximately 60% reduction of the mRNA and protein levels of MFN2 as evaluated by RT-qPCR and Western blot analysis (Fig 4A, Appendix Fig 5A, A′). In situ PLA revealed a significant reduction of VDAC1-IP3R1 proximity puncta in the Mfn2-knockdown group (Fig 4B, B′). Consistently, colocalization of mitochondria and ER was decreased, as shown by Mito-Tracker and ER-Tracker live-cell imaging (Fig 4C, C′). These results indicate that Mfn2 knockdown hampered MERC formation. In contrast to Mfn2 knockdown, overexpression of Linker via transfection of pcDNA3.1-Linker plasmid effectively enhanced MERC formation as shown by TEM (Appendix Fig 5B–C′). This plasmid expresses a fusion protein (mAKAP1[34–63]-mRFP-yUBC6[233–250]) (Csordás et al 2006) designed to enhance MERCs by tethering mitochondria to ER (Fig 4D).
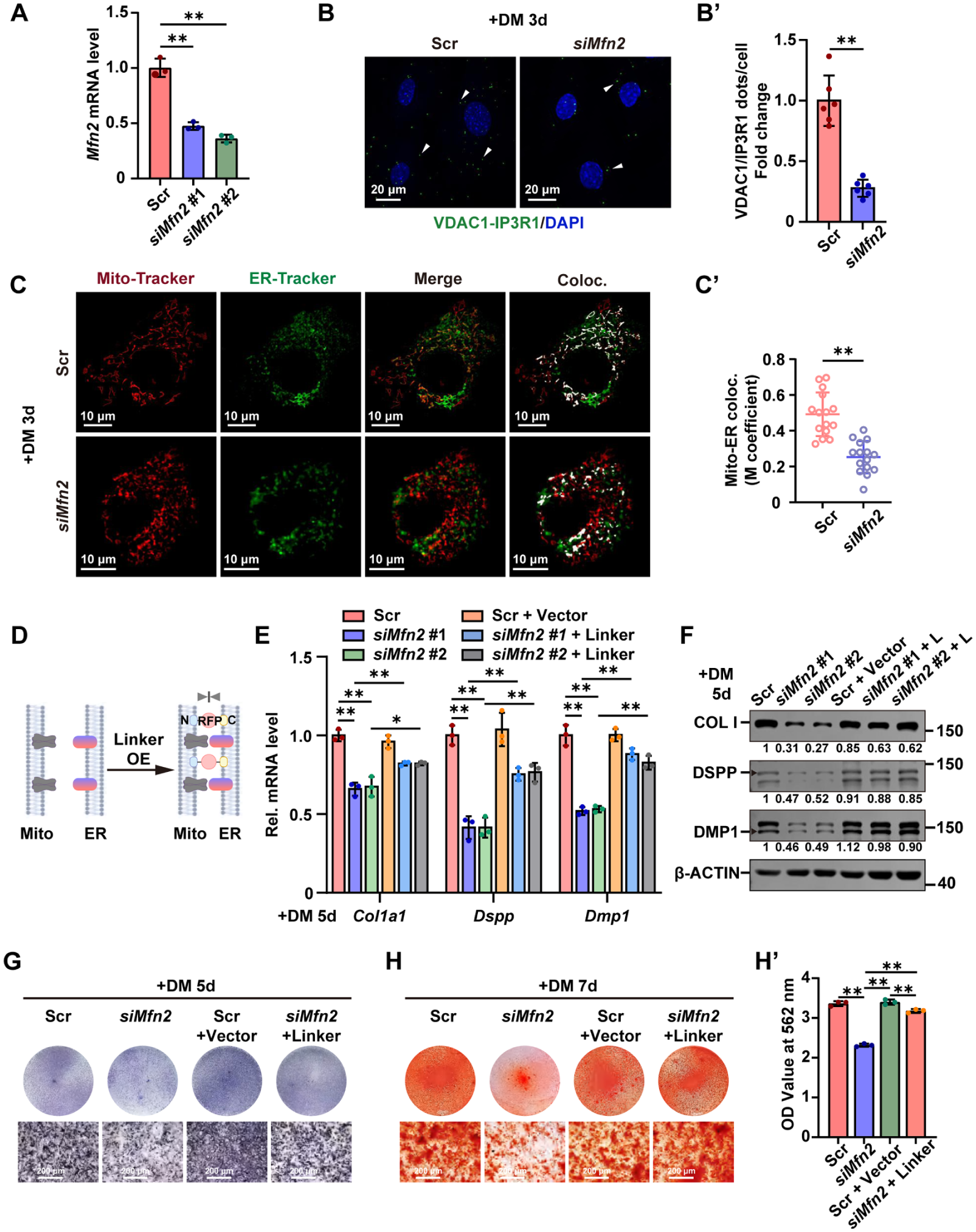
MFN2 promotes MERC formation and odontoblastic differentiation in vitro. (
RT-qPCR and Western blot analysis showed that knockdown of Mfn2 considerably inhibited the mRNA and protein levels of key odontoblast differentiation markers compared with the scramble control group (Fig 4E–F, Appendix Fig 5D). ALP staining and ARS staining revealed a corresponding decrease in both ALP activity and mineralization capacity following Mfn2 knockdown (Fig 4G–H′). Therefore, knockdown of Mfn2 inhibits odontoblastic differentiation of mDPCs.
To investigate whether reduced MERC formation was responsible for the suppressed odontoblastic differentiation due to Mfn2 knockdown, Linker was overexpressed in the Mfn2-knockdown cells. Western blot analysis showed that overexpression of Linker did not influence MFN2 expression (Appendix Fig 5E). Importantly, overexpression of Linker partially rescued the impaired mRNA and protein levels of the odontoblast markers attributed to Mfn2 knockdown as quantified by RT-qPCR and Western blot analysis (Fig 4E–F, Appendix Fig 5D). In addition, ALP staining and ARS staining showed that Linker overexpression obviously reversed both the impaired ALP activity and reduced mineralized nodule formation caused by Mfn2 knockdown (Fig 4G–H′). These findings indicate that the inhibitory effects of Mfn2 knockdown on odontoblastic differentiation can be partially rescued by enhancing MERCs using Linker overexpression. Thus, MFN2 promotes the odontoblastic differentiation of mDPCs through enhancing MERC formation. In addition, MFN2 is well characterized as being involved in mitochondrial fusion (Muñoz et al 2013). Its knockdown resulted in the morphological transformation of mitochondria from elongated tubular shapes to short rodlike forms, with a significant reduction in the mitochondrial length. However, overexpression of Linker failed to rescue the mitochondrial morphological alterations induced by Mfn2 knockdown (Appendix Fig 6A, A′).
MFN2 Facilitates MERC Formation to Enhance Dentin Formation In Vivo
To further assess the impact of MFN2 on odontoblast differentiation in vivo, we injected Mfn2 in vivo siRNA into the first molars of mice weekly from PN0.5 (Wang et al 2020; Fu et al 2023), with tissues harvested at 2 and 3 wk (Fig 5A). Successful knockdown of Mfn2 in odontoblasts was confirmed by IF, which revealed an approximately 60% reduction in MFN2 signals in PN4 molars (Fig 5B [a–d], B′). In situ PLA showed a nearly 70% decrease of VDAC1-IP3R1 puncta in Mfn2-knockdown odontoblasts, suggesting reduced MERC formation following knockdown of Mfn2 in vivo (Fig 5B [e–h], B″). Micro–computed tomography (micro-CT) analysis demonstrated that the volume of mineralized dentin was decreased in the molars of Mfn2-knockdown mice (Fig 5C, C′, Appendix Fig 7A, A′). Hematoxylin and eosin (H&E) staining further showed a decrease in dentin thickness and an increase in predentin thickness in Mfn2-knockdown mice compared with controls (Fig 5D–D″, Appendix Fig 7B–B″). In addition, IF analysis revealed that Mfn2 knockdown markedly reduced the expression of the odontoblast marker proteins including COL I, DSPP, and DMP1 (Fig 5E–E′″). Thus, Mfn2 knockdown inhibits MERC formation, odontoblast differentiation, and dentinogenesis in murine molars.
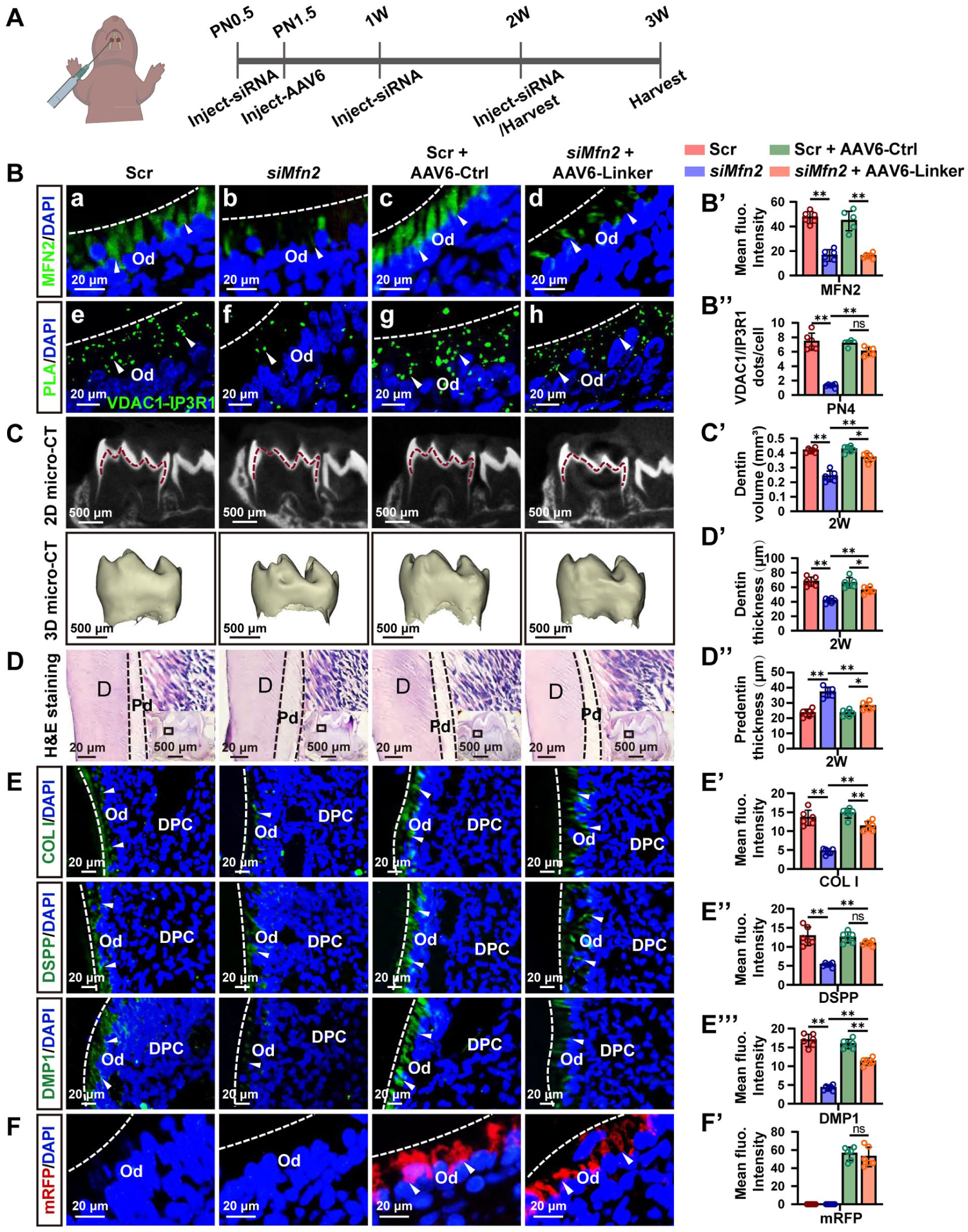
MFN2 facilitates MERC formation to enhance dentin formation in vivo. (
To confirm whether MFN2 regulates dentin formation through MERCs, we engineered an AAV6 vector expressing Linker under the control of an EF1α promoter (named AAV6-Linker) to test whether enhancing MERC formation could rescue the dentin hypoplasia phenotype in Mfn2-knockdown murine molars. This vector also encodes an mRFP reporter for tracking the transduction efficiency. The mRFP expression was clearly observed in the odontoblasts of PN4 molars, suggesting successful overexpression of Linker in vivo (Fig 5F, F′). In situ PLA using anti-VDAC1 and anti-IP3R1 antibodies further demonstrated that AAV6-Linker delivery significantly increased MERC formation in the odontoblasts (Fig 5B [g–h], B″). Micro-CT showed that the volume of mineralized dentin was regained by AAV6-Linker injection compared with the molars from Mfn2-knockdown mice (Fig 5C, C′, Appendix Fig 7A, A′). H&E staining revealed a significant reduction in predentin thickness and increase in dentin thickness in the molars injected with both AAV6-Linker and Mfn2 in vivo siRNA compared with those injected with Mfn2 in vivo siRNA alone (Fig 5D–D″, Appendix Fig 7B–B″). In addition, IF analysis showed that the hampered levels of COL I, DSPP, and DMP1 in Mfn2-knockdown mice were ameliorated following additional AAV6-Linker administration (Fig 5E–E′″). Taken together, MFN2 enhances MERC formation to promote odontoblast differentiation and dentinogenesis in vivo.
Discussion
MERCs are structural domains formed by close apposition of mitochondria and ER that enable interorganelle communication, yet their potential roles in the process of odontoblast differentiation and dentin formation remain largely unexplored (Csordás et al 2006; Arruda et al 2014). Here, we revealed that the formation of MERCs increases and plays a positive role in odontoblast differentiation and dentinogenesis, which is mainly mediated by MFN2. By shifting focus onto the intracellular membrane system and characterizing the dynamic changes of MERCs during differentiation, our work establishes an organelle-centric framework that may provide new etiological insights into dental developmental pathologies.
It has been reported that during the differentiation of mesenchymal cells into odontoblasts, there is an increase in organelles including mitochondria and ER (Sasaki and Garant 1996; Meng et al 2024). In this study, we first detected the changes of mitochondria and ER during the in vitro odontoblastic differentiation of mDPCs by labeling mitochondria and ER with Mito-Tracker and ER-Tracker, respectively (Li et al 2023). The results confirmed a progressive increase of mitochondria and ER during this process, which reflects an adaptation to the heightened physiological demands of energy and protein synthesis in differentiating odontoblasts. Interestingly, mitochondria–ER colocalization was also increased during this process, which was further verified by TEM and in situ PLA. The dynamic changes in MERCs suggest their potential role during odontoblast differentiation. Further experiments demonstrated that overexpression of FATE1 (Doghman-Bouguerra et al 2016), which disrupts MERCs, inhibited odontoblastic differentiation, as evidenced by reduced expression of odontoblast marker genes, diminished ALP activity, and impaired mineralized nodule formation. These data suggest that MERCs positively regulate the odontoblastic differentiation of mDPCs. A similar mechanism has been reported in osteoclasts, in which enhanced formation of MERCs triggers more active osteoclast differentiation (Gou et al 2025).
Several genes have been found to regulate MERC formation and function in hepatocytes and CD8+ T cells (Bassot et al 2021; Yang et al 2023). To identify key regulators of MERC formation during odontoblast differentiation, we intersected the MERC-associated gene set (Thoudam et al 2023) with the DEGs of RNA sequencing data of the mDPCs treated with differentiation medium for 11 d versus those treated for 0 d (Lin et al 2021). Mfn2 was one of the genes with significant upregulation. As a dynamin-related guanosine triphosphatase (GTPase), MFN2 is primarily localized at the outer mitochondrial membrane and participates in multiple fundamental cellular processes, including mitochondrial fusion, cellular metabolic homeostasis, and mitochondria–ER tethering (Muñoz et al 2013; McLelland et al 2018; Bassot et al 2021; Hu et al 2024). Previous studies have demonstrated that MFN2 regulates the differentiation of oocytes and bone marrow mesenchymal stem cells by modulating MERCs (Liao et al 2023; Fu et al 2025). Therefore, Mfn2 was considered a candidate gene essential for MERC formation and odontoblast differentiation. We then systematically characterized the MFN2 expression pattern and observed a rising trend in both the in vitro odontoblastic differentiation process and the in vivo odontoblast layer of murine incisors and molars. Furthermore, the formation of MERCs was inhibited by Mfn2 knockdown, confirming the positive regulatory role of MFN2. Notably, in vitro experiments demonstrated that Mfn2 knockdown inhibited the differentiation of odontoblasts, which were rescued by transfection of the pcDNA3.1-Linker plasmid. Overexpression of Linker did not affect MFN2 protein levels in odontoblastic cells but partially restored the Mfn2 knockdown-induced reduction in MERC formation and the impaired mRNA and protein levels of the odontoblast markers, indicating that MFN2 regulates odontoblast differentiation through MERCs. These results suggested that MFN2 upregulation during odontoblast differentiation enhances the formation of MERCs, thereby promoting the differentiation process in vitro.
To further evaluate the role of MFN2 in MERC formation and odontoblast differentiation under physiological conditions, we employed an in vivo animal model. Knockdown experiments were performed by local microinjection of Mfn2-targeting in vivo siRNAs into the first mandibular molars weekly from PN0.5 (Wang et al 2020; Fu et al 2023). Mandibles were harvested at 2 and 3 wk. We found that dentin mineralization in mouse molars was impaired by Mfn2 knockdown. AAV6 shows high specificity and transduction efficiency for odontoblasts (Fu et al 2023). Here, we found that AAV6-Linker administration promoted MERC formation in the odontoblasts and restored the dentin phenotype in Mfn2-knockdown molars, consistent with the findings of in vitro experiments. Therefore, dentin formation impairment by Mfn2 knockdown is primarily mediated through the inhibition of MERC formation, underscoring the essential role of MERCs in dentin formation.
MERCs serve as crucial bidirectional regulators of interorganelle communication in cells (Arruda et al 2014; Jang et al 2022; Gou et al 2025). A fundamental question that remains to be addressed is how MFN2-mediated MERCs orchestrate odontoblast differentiation and dentinogenesis. Previous studies have shown that MFN2 influences various physiological processes such as mitochondrial turnover, autophagy, and calcium flux, all of which are critical for odontoblast differentiation and dentin formation (McLelland et al 2018; Ding et al 2022; Wang et al 2022). Whether MFN2 influences odontoblast differentiation through these pathways remains unclear. Given that odontoblast differentiation also relies on proper mitochondrial homeostasis (Zhang et al 2023), the unrecovered mitochondrial morphology may explain why the defects caused by Mfn2 knockdown were only partially restored after Linker overexpression. Since MERCs are formed by tethering complexes residing on mitochondrial and ER, the specific interacting partners of MFN2 on the ER of odontoblasts also warrant further investigations in the future.
In conclusion, our findings demonstrate that MERCs mediated by MFN2 drive odontoblast differentiation and dentin formation. This study establishes a previously unrecognized role of MFN2 in regulating MERCs formation during odontoblast differentiation. The MFN2–MERCs axis represents a fundamental regulatory mechanism for dentinogenesis, offering new possibilities for developing regenerative dental therapies targeting organelle interactions.
Author Contributions
J. Sheng, contributed to design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; L. Yan, contributed to acquisition, analysis, and interpretation, drafted the manuscript; Z. Chen, G. Yang, contributed to conception and design, critically revised the manuscript; G. Yuan, contributed to conception and design, data interpretation, critically revised the manuscript. All authors gave their final approval and agreed to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345261458702 – Supplemental material for MFN2-Mediated MERCs Promote Odontoblast Differentiation and Dentinogenesis
Supplemental material, sj-docx-1-jdr-10.1177_00220345261458702 for MFN2-Mediated MERCs Promote Odontoblast Differentiation and Dentinogenesis by J. Sheng, L. Yan, Z. Chen, G. Yang and G. Yuan in Journal of Dental Research
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the National Natural Science Foundation of China (82370913 to G. Yuan and 82270947 to G. Yang), the Fundamental Research Funds for the Central Universities (2042026kf0003, 2042022dx0003), the National Natural Science Foundation of China Key Program (82230029 to Z. Chen).
Data Availability
The RNA-seq data of CRA003233 are available through Search - GSA - CNCB-NGDC. The datasets analyzed in this study can be found in the Supplementary Appendix (![]() ). All data will be made available on request.
). All data will be made available on request.
A supplemental appendix to this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.