
Abstract
Precise control of orthodontic tooth movement (OTM) requires a deep understanding of the biological mechanisms underlying alveolar bone remodeling. Although the immune system is known to influence osteoclastogenesis, the functional significance of neutrophils in the noninfectious, sterile inflammatory environment of OTM remains largely unexplored. Here, we aimed to elucidate the mechanisms by which neutrophils contribute to bone resorption during OTM. OTM was induced in mice using a 10-g mesial force. Single-cell RNA sequencing (scRNA-seq) was performed on periodontal ligament tissues to characterize cellular heterogeneity and intercellular communication. In addition, neutrophil subpopulations and developmental trajectories were analyzed using pseudotime analysis. Finally, the functional role of neutrophils was validated in vivo by systemic depletion using an anti-Ly6G antibody. scRNA-seq identified 11 cell clusters, revealing that neutrophils are a primary source of proinflammatory cytokines, including tumor necrosis factor (TNF), interleukin (IL)–1β, and Oncostatin M (OSM). Immunofluorescence analysis confirmed that neutrophils preferentially accumulated on the compression side. CellChat analysis and transcriptomic profiling identified a TNF-TNF receptor 2 (TNFR2) signaling axis directed from neutrophils to macrophages. Subclustering revealed an “Inflammatory-Neu” subset that expands during OTM and expresses CC ligand chemokine family members to recruit macrophages. In vivo, neutrophil depletion significantly attenuated macrophage accumulation on the compression side, resulting in diminished tooth movement distance and reduced osteoclastogenesis. Our findings demonstrate that neutrophils are indispensable upstream regulators of OTM. By maturing into a proinflammatory phenotype, neutrophils coordinate macrophage recruitment and activation via the TNF-TNFR2 axis, thereby driving osteoclastogenesis. This study provides a novel biological framework for understanding the osteoimmunological microenvironment during orthodontic loading, and it also identifies potential targets for controlling tooth movement.
Introduction
For orthodontists, achieving precise control over tooth movement is the ultimate objective of clinical practice. As the force delivered by orthodontic appliances is fundamentally a mechanical phenomenon, biomechanical analyses using methods such as the finite element method have enabled researchers to predict tooth displacement with some degree of accuracy (Kawamura et al 2021; Guo et al 2024). However, clinical outcomes often diverge from this theoretical prediction, resulting in inadequate tooth movement and prolonged treatment periods. Orthodontic tooth movement (OTM), which induces a complex biological response through the application of force, results in alveolar bone resorption mediated by the formation and activation of osteoclasts on the compression side (Kitaura et al 2025). To precisely predict and control tooth movement during this process, it is essential to have a deep understanding of the biological mechanisms involved. Yet several aspects of osteoclast formation in OTM remain unclear. Therefore, elucidation of the osteoclast regulatory mechanisms in OTM would be an important step toward achieving a more predictable orthodontic treatment.
Advances in osteoimmunology have revealed the pivotal role of the immune system in bone metabolism (Tsukasaki and Takayanagi 2019). The interplay between osteoimmunological inflammatory responses and bone remodeling during OTM has garnered significant attention recently (Chaushu et al 2022; Alghamdi et al 2023). Studies using bulk RNA sequencing to analyze gene expression changes in mouse OTM models have suggested that neutrophil signaling is activated during this process (Klein et al 2020). Furthermore, evidence suggests that neutrophils may contribute to osteoclastogenesis in OTM, as the depletion of γδT cells substantially reduces both neutrophil infiltration within the periodontal ligament (PDL) and the rate of tooth movement (Wald et al 2021). A recently published single-cell atlas of mouse OTM models also showed high expression of cytokine-related genes in specific neutrophil subtypes (Wang et al 2025). Although neutrophils have traditionally been regarded as rapid responders to invading pathogens, recent studies have demonstrated that they exhibit functional diversity (Zhang et al 2024). As alveolar bone resorption in periodontal disease is driven mainly by infectious inflammation, the role of neutrophils has been extensively reported in that context (Hiyoshi et al 2022; Kim et al 2023; Ando et al 2024). Conversely, OTM is a noninfectious, sterile inflammatory process. The specific contribution of neutrophils to tooth movement during OTM remains largely unexplored.
In this study, we demonstrated that neutrophils significantly influence osteoclastogenesis and tooth movement in a mouse OTM model. Single-cell RNA sequencing (scRNA-seq) analysis revealed that neutrophils highly express tumor necrosis factor (TNF) during the inflammatory phase and that TNF signaling is critical for osteoclastogenesis. These findings provide novel insights into the role of neutrophils in OTM-associated osteoclastogenesis and represent a major step toward the precise control of tooth movement.
Materials and Methods
Animals and OTM Model
Male C57BL/6J mice (8 wk old) were used, with institutional approval (No. 2024DnA-003-03). OTM was induced by applying a 10-g mesial force to the maxillary left first molar using a nickel-titanium closed-coil spring for 6 or 12 d (Appendix Fig 1A) (Kitaura et al 2008).
scRNA-seq and Bioinformatic Analysis
On day 6, PDL cells were harvested from pooled maxillary first molars (n = 5 per group) (Appendix Fig 1B) and processed using Chromium Next GEM Single Cell 3′ Reagent Kits v3.1 (10x Genomics). Data were analyzed using the Seurat R package for clustering and cell-type annotation (Appendix Fig 2). Pathway enrichment and cell–cell communication were evaluated using the clusterProfiler and CellChat packages, respectively. Neutrophil heterogeneity and developmental trajectories were analyzed via subclustering and Monocle3.
Neutrophil Depletion and In Vivo Evaluation
For systemic neutrophil depletion, mice received intraperitoneal injections of an anti-Ly6G antibody (clone 1A8; 0.4 mg) or an isotype control starting 1 d before OTM and every 3 d thereafter (Daley et al 2008; Ando et al 2024). On day 12, the tooth movement distance was measured using dental impressions. To validate the efficiency of neutrophil depletion and evaluate its effect on macrophage recruitment, immunofluorescence staining for Ly6G and CD68 was performed. Osteoclastogenesis was evaluated using tartrate-resistant acid phosphatase (TRAP) staining.
Statistical Analysis
Data are presented as mean ± standard deviation (SD). Statistical significance was determined using Student’s t test or 1-way analysis of variance, followed by Tukey’s post hoc test (P < 0.05).
Detailed experimental procedures, including bioinformatics pipelines and antibody information, are provided in the Supplementary Appendix.
Results
Single-Cell Atlas Identifies Neutrophils as a Major Source of Inflammatory Cytokines during OTM
To elucidate the cellular heterogeneity of the PDL under orthodontic force, we performed scRNA-seq analysis on the PDL tissues harvested from the control and OTM groups (day 6). After rigorous quality control and filtering, the integrated dataset was visualized using a Uniform Manifold Approximation and Projection (UMAP). Unsupervised clustering identified 11 distinct cell populations that were annotated based on the expression of canonical marker genes (Fig 1A, B). These clusters comprised fibroblasts, neutrophils (Ly6G and S100a8), PDL progenitors, endothelial cells, epithelial cells, macrophages, osteoblasts, B cells, Schwann cells, smooth muscle cells/pericytes, and erythrocytes (Fig 1B). We then compared the cellular composition of the control and OTM groups (Fig 1C, D). Quantitative analysis of the cell proportions revealed dynamic shifts in the PDL microenvironment during OTM. Although fibroblasts were markedly increased in the OTM group, the proportion of immune cells, including neutrophils, remained relatively stable between the control and OTM conditions in the scRNA-seq dataset (Fig 1E). Given that OTM triggers a robust inflammatory response, we sought to identify the specific cellular sources of inflammatory mediators within the PDL. We focused on the expression of Tnf, Il1b, and Osm, which are cytokines critical for neutrophil-mediated inflammatory responses (Zhang et al 2024). Violin plots and UMAP-based feature plots revealed that the expression of these cytokines was predominantly enriched in the neutrophil cluster and in macrophages (Fig 1F, G). Transcriptomic analysis showed that Tnfsf11, which encodes receptor activator of NF-κB ligand (RANKL), was primarily expressed by the fibroblast cluster in the OTM group, while its receptor, Tnfrsf11a (encoding RANK), was mainly detected in the macrophage cluster, which includes osteoclast precursors (Appendix Fig 3). These findings suggest that neutrophils, together with macrophages, represent prominent sources of inflammatory cytokines and thereby contribute to the establishment of a proinflammatory microenvironment during OTM.
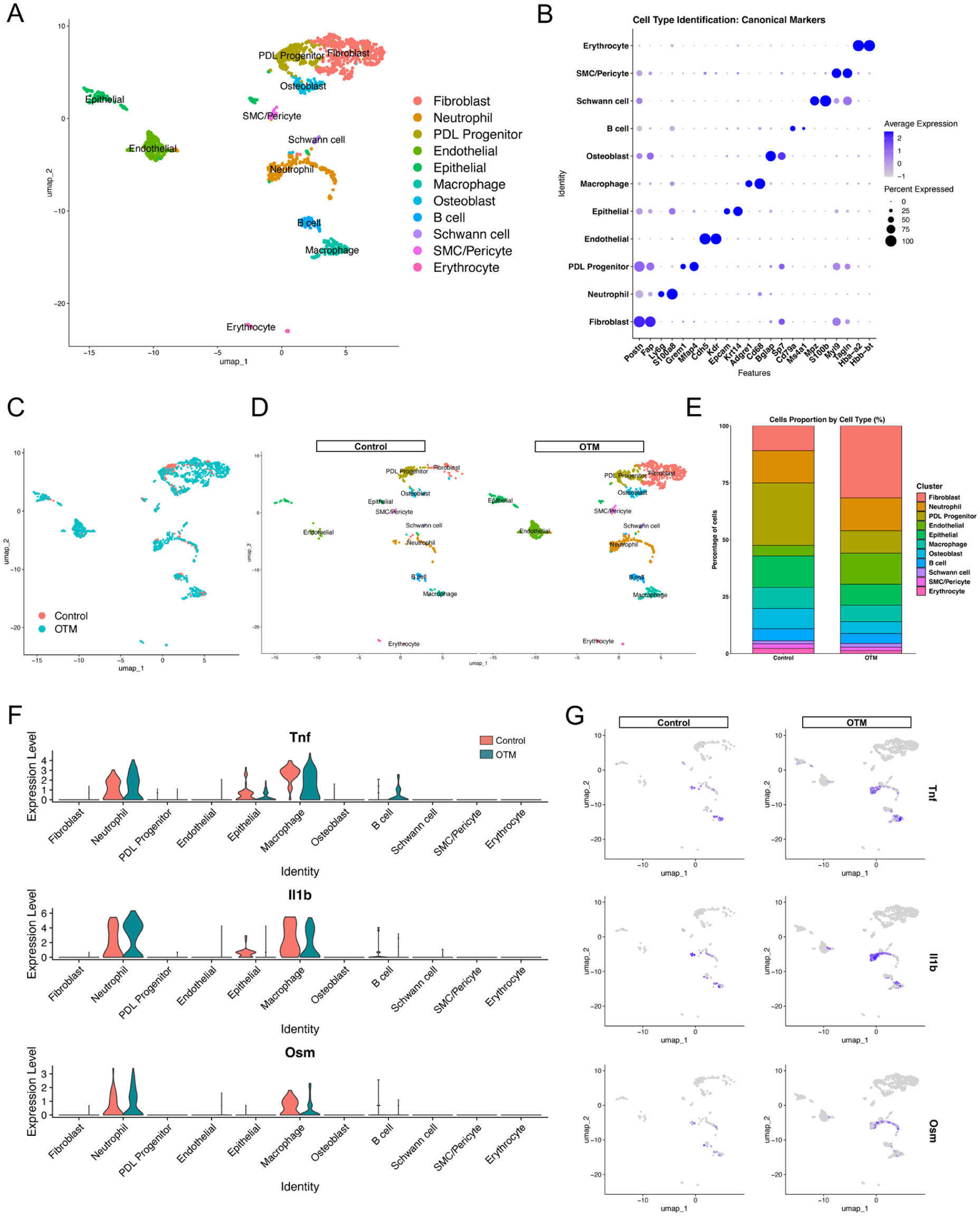
Single-cell transcriptomic landscape of the periodontal ligament (PDL) during orthodontic tooth movement (OTM). (
Neutrophils Preferentially Accumulate at the Compression Side during OTM
We performed immunofluorescence staining for Ly6G (neutrophils) and CD68 (macrophages) in the maxillary PDL to characterize the spatial distribution of neutrophils during OTM. In the control group, Ly6G-positive neutrophils and CD68-positive macrophages were distributed on both the compression and tension sides. Conversely, following OTM induction, both cell types preferentially accumulated on the compression side, coinciding with regions of active bone resorption (Fig 2A, Appendix Fig 4). To assess the functional implications of this recruitment, we performed Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of neutrophil and macrophage marker genes. Notably, the “osteoclast differentiation” pathway was significantly enriched in both populations, suggesting its direct involvement in alveolar bone resorption during OTM (Fig 2B).
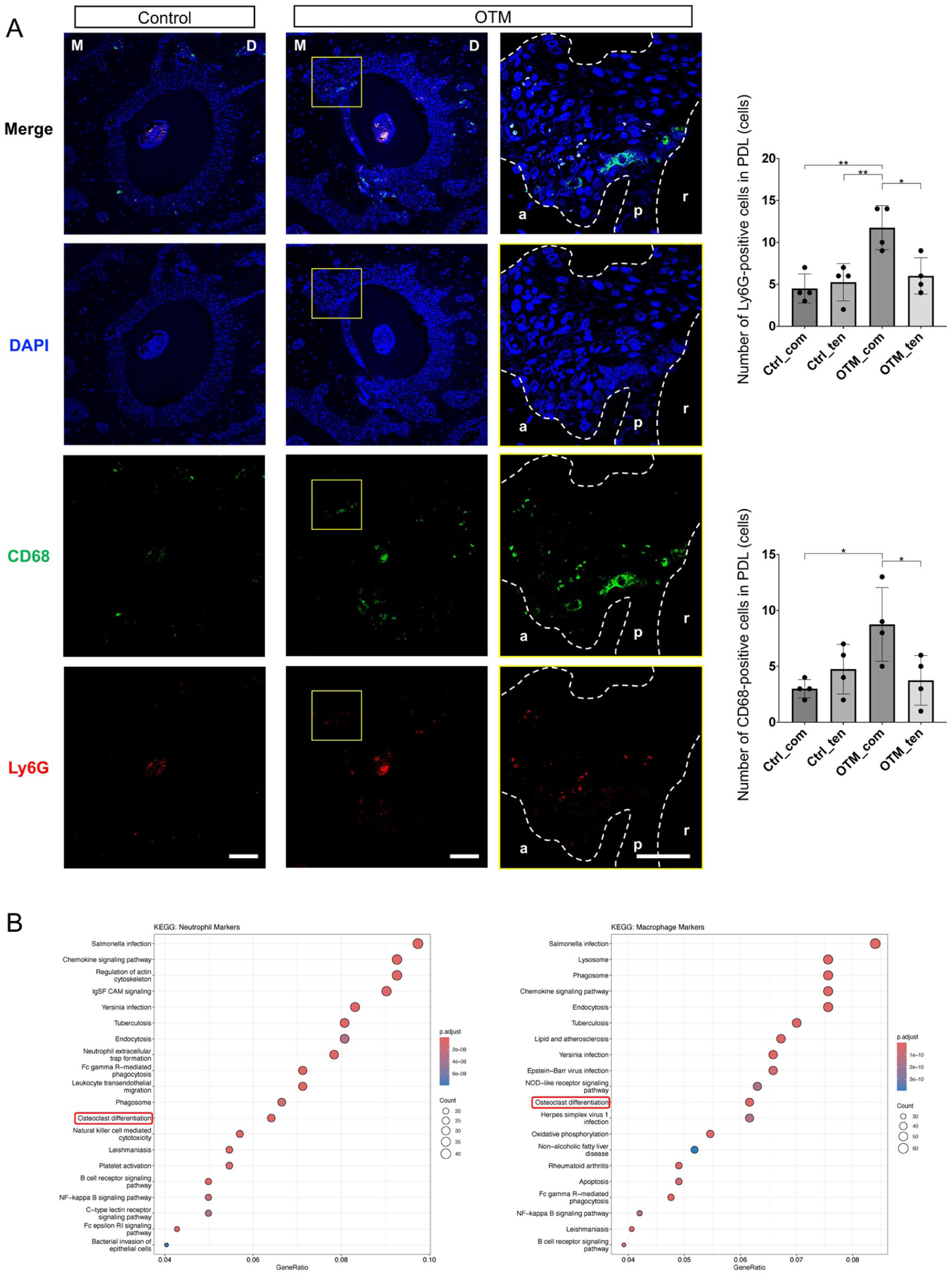
Spatial distribution of neutrophils and functional enrichment of neutrophils and macrophages during orthodontic tooth movement (OTM). (
Neutrophils Coordinate Macrophage Responses via the TNF Signaling Axis
To elucidate the intercellular communication orchestrating this microenvironment, we used CellChat to analyze cell–cell communication. In the OTM group, both the number and strength of signaling interactions increased compared with those in the control (Fig 3A). A comparison of the overall signaling pathways revealed that neutrophils prominently contributed to various proinflammatory networks, including TNF, interleukin (IL)–1, and Oncostatin M (OSM) (Fig 3B). Focusing on these specific pathways, we found that while neutrophils actively produced multiple mediators, their communication with macrophages was highly selective. IL-1 signaling was undetectable, and OSM signaling showed a negligible interaction between these clusters. Conversely, the TNF signaling pathway was prominently directed from neutrophils to macrophages in the OTM group (Fig 3C, Appendix Fig 5). Network centrality analysis identified neutrophils as the predominant “sender” of TNF signals, with macrophages acting as the primary “receiver” (Fig 3D, E). A detailed interaction analysis revealed that this crosstalk was primarily mediated by the TNF–Tnfrsf1b pair in the OTM group (Fig 3F). Consistently, transcriptomic analysis showed that the expression of Tnfrsf1b (encoding TNF receptor 2) was upregulated in macrophages following OTM treatment (Fig 3G). Collectively, these findings indicate that the neutrophil–macrophage TNF signaling axis is a pivotal regulatory component of the OTM-induced inflammatory landscape, warranting a deeper investigation into the functional diversity of the infiltrating neutrophil population.
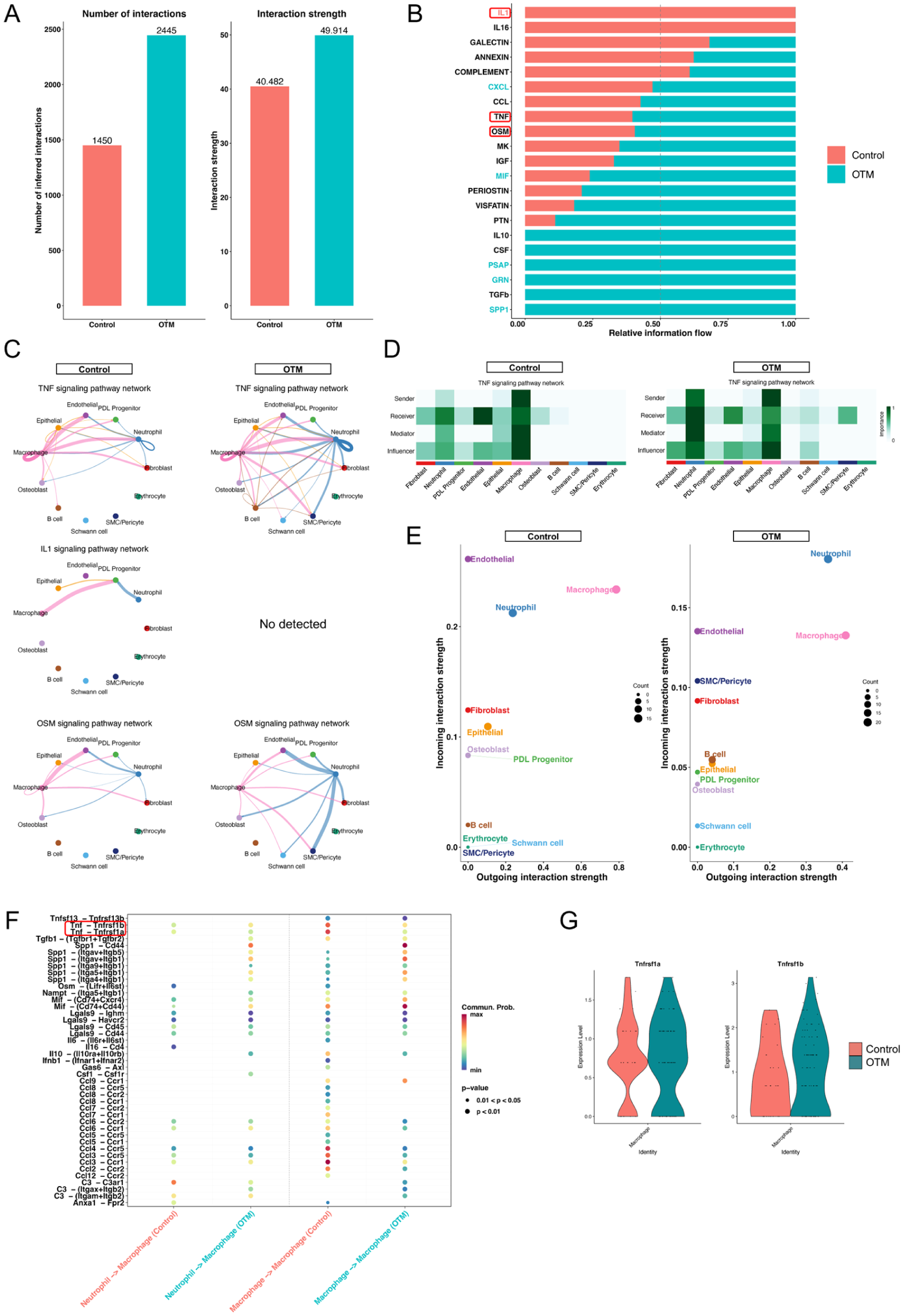
CellChat analysis identifies the neutrophil–macrophage tumor necrosis factor (TNF) signaling axis during orthodontic tooth movement (OTM). (
Identification of a Proinflammatory Neutrophil Subset Driving Macrophage Recruitment
To explore the functional heterogeneity of the infiltrating neutrophils, we performed a subclustering analysis, which identified 3 distinct subsets: Initial-Neu, Inflammatory-Neu, and Adapted-Neu (Fig 4A). Specifically, these subclusters were defined and named based on their characteristic marker gene profiles: Initial-Neu was characterized by high expression of immature markers (Ngp and Ltf), Inflammatory-Neu showed robust expression of proinflammatory factors (S100a8, S100a9, Tnf, and Ccl3), and Adapted-Neu was identified by genes associated with tissue adaptation and functional modulation (Ahnak and Klf4). These subsets were characterized by their unique transcriptomic profiles (Fig 4B, C). Comparative analysis between groups revealed a dynamic shift in neutrophil composition following OTM induction (Fig 4D). Notably, the proportion of the Inflammatory-Neu subset was markedly increased in the OTM group compared with that in the control, while the Initial-Neu population diminished (Fig 4E). To elucidate the transition patterns between these states, we performed a pseudotime analysis using Monocle3. The results demonstrated a clear transition path originating from Initial-Neu, progressing through Inflammatory-Neu, and terminating in the Adapted-Neu state (Fig 4F). Along this trajectory, the expression of Ngp decreased rapidly, while in the later pseudotime period, Tnf peaked and Ahnak increased steadily, reflecting the dynamic activation and functional shift of neutrophils during OTM (Fig 4G). Gene Ontology (GO) enrichment analysis was performed to further characterize the role of the expanded Inflammatory-Neu subset. The marker genes for this subset were significantly enriched in terms related to “leukocyte migration,” “myeloid leukocyte activation,” and “tumor necrosis factor superfamily cytokine production” (Fig 4H). These findings suggest that this specific neutrophil subset not only produces inflammatory signals but also actively coordinates the recruitment of other immune cells. Consistent with the GO analysis, the inflammatory-Neu subset expressed high levels of CC ligand chemokine family members, including Ccl3, Ccl4, and Ccl6 (Fig 4I). Correspondingly, transcriptomic analysis of the macrophage population revealed significant expression of their receptors Ccr1, Ccr2, and Ccr5 (Fig 4J). Altogether, these results suggest that OTM-induced polarization of neutrophils toward an inflammatory phenotype promotes the localized recruitment and activation of macrophages.
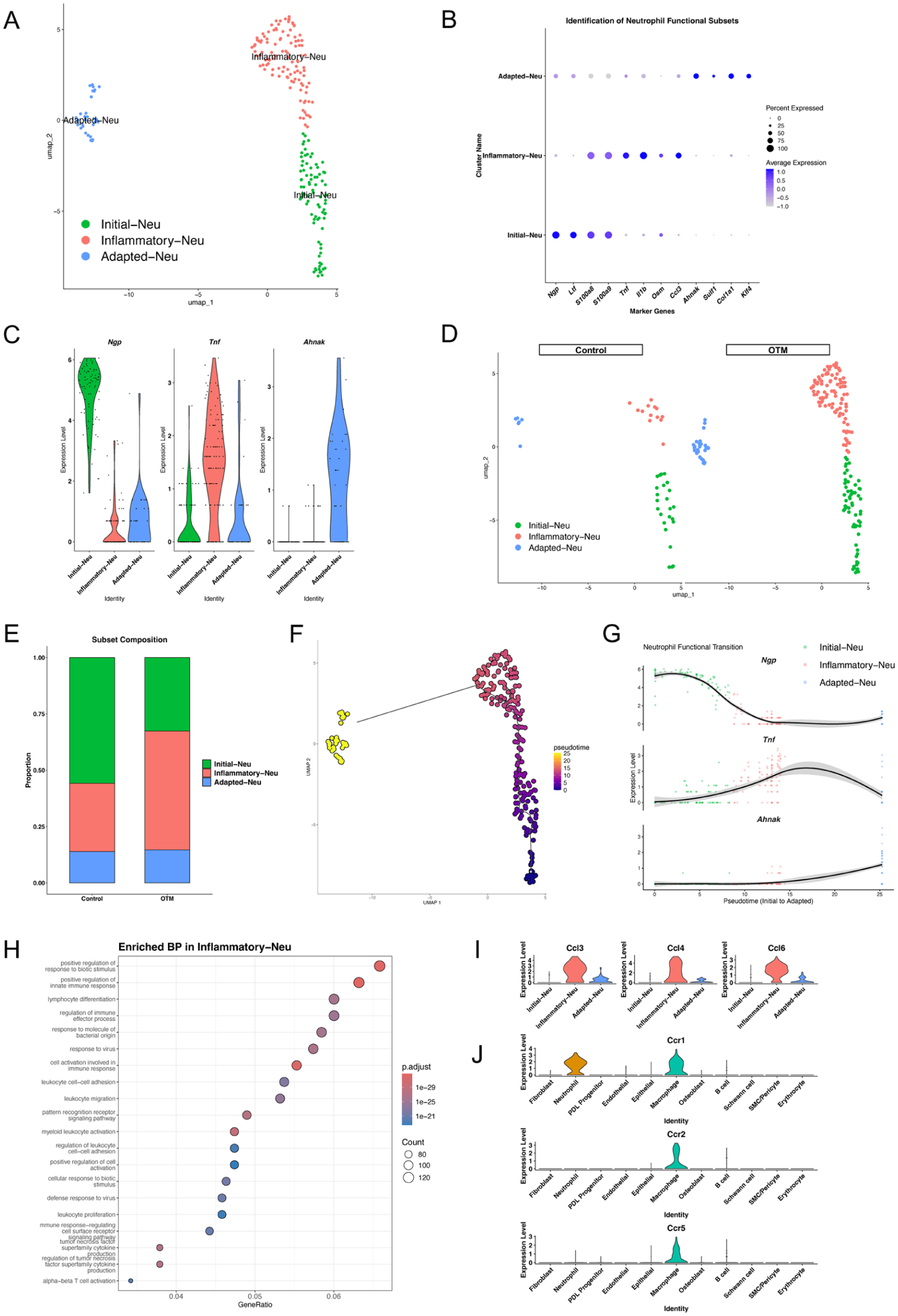
Characterization of neutrophil functional subsets and their role in macrophage recruitment. (
Neutrophil Depletion Attenuates Tooth Movement and Osteoclastogenesis
To definitively establish the functional necessity of neutrophils in OTM, we performed systemic neutrophil depletion using an anti-Ly6G antibody (Fig 5A). The efficacy of the depletion and its impact on macrophage recruitment were validated using immunofluorescence staining. Notably, the absence of neutrophils significantly reduced the accumulation of CD68-positive macrophages on the compressed side (Fig 5B). Consequently, the distance of tooth movement was significantly reduced (Fig 5C, Appendix Fig 6). Histological analysis confirmed that the number of TRAP-positive multinucleated osteoclasts was significantly diminished following neutrophil depletion (Fig 5D). In support of these findings, scRNA-seq analysis confirmed the enrichment of macrophage (Cd68)–, neutrophil (Ly6g)–, and osteoclast (Acp5, Ctsk)–related markers within OTM-induced cell clusters (Appendix Fig 7). Collectively, by integrating single-cell transcriptomic profiling with in vivo neutrophil depletion, our findings established OTM-induced inflammatory neutrophils as critical upstream regulators that orchestrate macrophage recruitment and TNF-dependent osteoclastogenesis within the PDL microenvironment (Fig 5E).
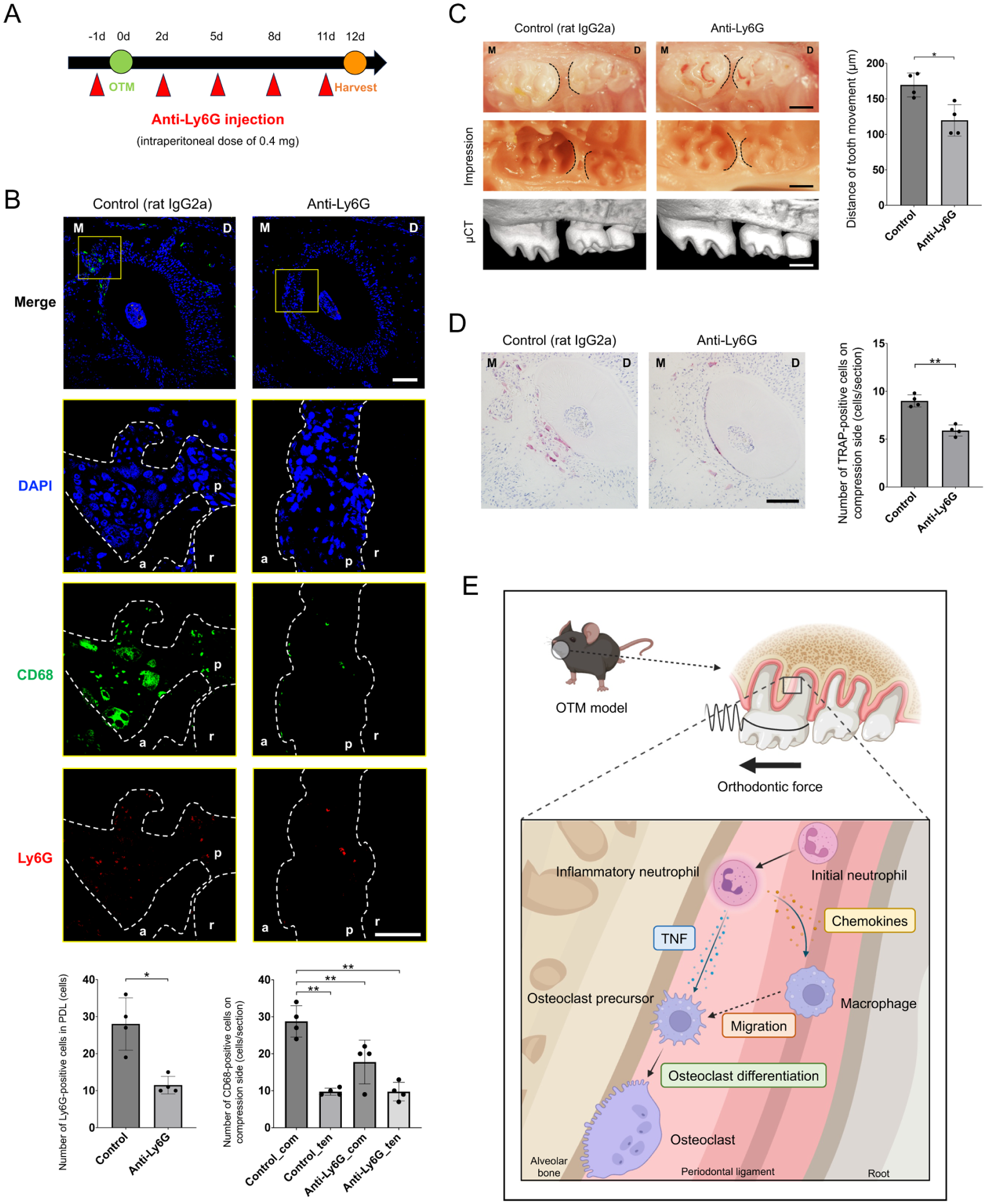
Neutrophil depletion attenuates macrophage recruitment, orthodontic tooth movement (OTM), and osteoclastogenesis. (
Discussion
OTM is a unique biological process characterized by bone resorption on the compression side, a phenomenon that seemingly contradicts Wolff’s Law and Frost’s Mechanostat Theory, both of which associate mechanical loading with bone formation (Wolff 1892; Frost 2003). This discrepancy is attributed to the specialized anatomical role of PDL, which acts as a signal transducer to regulate adjacent alveolar bone remodeling (Wen et al 2025). While recent scRNA-seq studies on OTM mouse models have primarily focused on the alveolar bone (Xu et al 2022; Fan et al 2024; Wang et al 2025), our study specifically targeted the PDL to capture the fundamental triggers of OTM. Our identification of 11 distinct cell populations is largely consistent with previous PDL atlases (Zhao et al 2022; Ando et al 2024; Chida et al 2025), providing a strong foundation for analyzing sterile inflammatory responses during OTM.
A crucial finding of this study is that neutrophils are a primary source of proinflammatory cytokines, particularly TNF, during OTM. While the classical paradigm emphasizes PDL fibroblasts as the initial mechanosensors that trigger early inflammation (within 1–3 d), our temporal focus on day 6 highlights a distinct orchestration phase. During this period, the relative contribution shifts from initial mechanotransduction by fibroblasts to active immune coordination by neutrophils. While fibroblasts provide the essential osteoclastogenic signal (RANKL), neutrophils appear to amplify the local inflammatory environment and recruit macrophages to the compression side. Thus, neutrophils act as a critical bridge between early mechanical sensing and late-stage bone remodeling. It is well-known that TNF induces osteoclastogenesis (Azuma et al 2000; Kitaura et al 2004) and synergizes with RANKL (Fuller et al 2002). Although macrophages were previously considered the main producers of TNF in OTM (He et al 2015), our data demonstrated that neutrophils contribute equally or even more significantly to TNF expression. Our spatial immunofluorescence analysis confirmed that Ly6G-positive neutrophils preferentially accumulated on the compression side, which was likely guided by signals from dead cells during mechanical loading (McDonald et al 2010; Li et al 2016; Ohori et al 2026). KEGG analysis highlighted the enrichment of neutrophil extracellular trap formation, which has recently been linked to alveolar bone loss in periodontitis (Si et al 2026). This suggests that neutrophils in OTM use mechanisms similar to those in infectious inflammation to drive bone resorption despite the absence of pathogens.
TNF signaling is mediated by 2 different receptors: the 55-kDa TNF receptor 1 (TNFR1) and the 75-kDa TNF receptor 2 (TNFR2) (Kitaura et al 2022). CellChat analysis identified the TNF-TNFR2 axis as a key mediator of neutrophil–macrophage communication. Although the relative importance of TNFR1 and TNFR2 in osteoclastogenesis remains debatable (Abu-Amer et al 2000; Kobayashi et al 2000), our findings support the specific significance of TNFR2 in OTM. A previous study has shown that OTM distance and osteoclastogenesis are more severely impaired in TNFR2-deficient mice compared with TNFR1-deficient mice (Yoshimatsu et al 2006). The upregulation of Tnfrsf1b (encoding TNFR2) in macrophages following OTM suggests that neutrophil-derived TNF specifically targets this receptor to regulate the osteoclastogenic niche, a mechanism distinct from that of other inflammatory bone diseases, where TNFR1 might predominate.
Although neutrophils have traditionally been regarded as a homogeneous population of immune cells, they are now recognized to exhibit significant functional heterogeneity (Ng et al 2025). Our subclustering analysis revealed the functional heterogeneity of neutrophils, identifying an Inflammatory-Neu subset that expanded during OTM. This subset appears to serve as a chemotactic hub that expresses CC ligand chemokine family members to recruit macrophages to the compression side. The subsequent transition to the Adapted-Neu state or potential reverse migration (Ji et al 2024) may represent a resolution phase or a shift in the functional state. The essential role of this neutrophil-driven recruitment was confirmed by depletion experiments, in which the loss of neutrophils significantly attenuated macrophage infiltration, osteoclastogenesis, and tooth movement. While our results mirror the findings in periodontitis (Ando et al 2024), it is important to note that neutrophil progenitors in the bone marrow have recently been reported to suppress osteoclast formation (Isojima et al 2025). This functional dichotomy, in which mature inflammatory neutrophils promote bone resorption, while neutrophil progenitors may be bone protective, emphasizes the need for subset-specific therapeutic targeting in orthodontic treatment.
Nevertheless, this study has some limitations. The scRNA-seq data lacked spatial resolution, meaning that cells from the compression and tension sides were analyzed as a pooled population. The minute scale of the mouse PDL precluded the technical separation of the compression and tension sides, which may have masked side-specific molecular signatures within the population. Furthermore, because this study focused on the PDL, interactions with cell populations deep within the alveolar bone remain unexplored. Future studies using spatial transcriptomics will be essential to map these intercellular interactions at high resolution. In addition, while systemic neutrophil depletion using the anti-Ly6G antibody is a well-established method, it does not entirely exclude the potential influence of systemic inflammatory shifts on the OTM. Therefore, attributing the observed effects to local neutrophil–macrophage interactions within the PDL requires cautious interpretation.
In conclusion, we have identified neutrophils as indispensable upstream regulators of OTM. By maturing into a specific inflammatory phenotype, neutrophils recruit macrophages and drive osteoclastogenesis via the TNF-TNFR2 axis. These findings establish a previously unrecognized neutrophil-macrophage axis as a fundamental driver of orthodontic bone remodeling, thereby advancing our understanding of the complex cellular orchestration underlying OTM.
Author Contributions
F. Ohori, H. Kitaura, contributed to conception and design, data acquisition and interpretation, drafted and critically revised the manuscript; K. Narita, A. Marahleh, J. Ma, K. Kanou, Z. Fan, A. Lin, K. Murakami, contributed to acquisition and interpretation, critically revised the manuscript; H. Kanetaka, contributed to conception and design, critically revised the manuscript. All authors provided their final approval and agreed to be accountable for all aspects of this work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345261459909 – Supplemental material for Neutrophils Orchestrate Osteoclastogenesis in Orthodontic Tooth Movement
Supplemental material, sj-docx-1-jdr-10.1177_00220345261459909 for Neutrophils Orchestrate Osteoclastogenesis in Orthodontic Tooth Movement by F. Ohori, H. Kitaura, K. Narita, A. Marahleh, J. Ma, K. Kanou, Z. Fan, A. Lin, K. Murakami and H. Kanetaka in Journal of Dental Research
Footnotes
Acknowledgements
The authors would like to thank the support system for young researchers who used the research equipment, instruments, and devices at Tohoku University. We are grateful to the Center of Common Research Laboratory, Graduate School of Dentistry, Tohoku University, for providing access to the micro–computed tomography imaging. We also acknowledge the Biomedical Research Core of the Tohoku University Graduate School of Medicine for its support and for the facilities used for immunofluorescence imaging.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by JSPS KAKENHI (grant numbers JP24K20049, JP24K23539, and JP25K02830) of the Japan Society for the Promotion of Science.
Data Availability
The single-cell RNA sequencing datasets generated in this study are accessible through the NCBI Gene Expression Omnibus (GEO) under accession number GSE324651.
A supplemental appendix to this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.