
Abstract
Pre-immunization with inactivated antigens has been developed as an alternative to the use of ‘dirty’ mice, which in contrast to specific pathogen free (SPF) mice, harbour a range of pathogens. Within certain research areas, such mice are considered better models for humans than SPF mice, as they have an immune system that better mirrors human immunity. We inactivated murine adenovirus type 1 (FL), minute virus of mice, mouse hepatitis virus (A59), respirovirus muris (Sendai), Theiler’s encephalomyelitis virus (GD7) and Mycoplasma pulmonis by ultraviolet irradiation. We show that pre-immunization with these inactivated pathogens combined with adjuvant prior to the dietary induction of obesity in C57BL/6NTac mice substantially reduced the group sizes needed for showing an effect of the GLP-1 receptor analogue, liraglutide. Nesting, open field and novel object behaviours of the mice were unaffected. We conclude that pre-immunization with inactivated pathogens may be a simple tool to increase power in this type of intervention study on the DIO mouse model.
Introduction
The specific pathogen free (SPF) concept based upon (a) caesarean section or embryo transfer of initial breeders, (b) protective barrier housing behind and (c) screening of samples for infections was introduced in 1958, 1 because infections in laboratory rodents caused animal mortality and staff zoonoses. 2 The Federation of European Laboratory Animal Science Associations (FELASA) guidelines set high standards for how to screen for which agents, 3 which has eliminated many of these. 2 However, the immune memory of SPF mice in contrast to adult humans, wild mice and pet-shop mice, resembles the immune system of a new-born baby. 4 This may limit the translatability of animal studies. 5 Therefore, some research groups have made so-called ‘dirty’ mice by co-housing of pet-shop mice with laboratory mice, 4 ‘feralization’ by housing mice in an enriched environment continuously supplied with materials from farm animal stables and nature 6 and embryo transfer of C57BL/6 embryos to wild mice, so-called ‘wildlings’. 7 These are exposed to previously eradicated pathogens,4,6,7 and eventually also carry a microbiota more comparable to that of wild mice. 7
Compared with SPF mice, ‘dirty’ mice infected with influenza virus as humans survive far better. 8 Like humans they exhibit a declining antibody response and more human-like transcriptional signatures after influenza vaccination, while SPF mice exhibit a long lasting antibody response. 9 Feralized laboratory mice are better protected against colorectal carcinogenesis. 6 The anti-CD28 immune blocker caused a nearly fatal cytokine storm in humans subjected to a clinical trial, 10 which in contrast to SPF mice was also observed when retested in wildlings. 7 As wildlings did not respond to the blocker with an increased number of regulatory T cells as observed in the SPF mice, 7 this drug candidate would probably not have passed the preclinical phase had ‘dirty’ mice been applied.
Reintroduction of active pathogens to modern laboratory animal facilities may render the animals in different phases of infection. Also the animal diseases, the zoonoses, and increased mortality may reappear. We, therefore, previously developed and validated a more controllable regime. 11 We selected and inactivated by ultraviolet irradiation five common murine viruses from different viral orders and one respiratory bacterial pathogen (Figure 1(a)). 11 Injecting this mixture twice subcutaneously with an adjuvant increased the number of CD8+ effector memory T cells to a level comparable to that of wild mice, and induced high serum antibody titres to the pathogens. 11 Co-housed control mice remained sero-negative and screening sentinels for two years after did not reveal any animals positive to any of these six agents. 11 Comparing the effect with phosphate buffered saline (PBS) or with one of the two adjuvants Addavax or Alhydrogel (alum) showed that the immune impact was solely due to the inactivated pathogens. 11
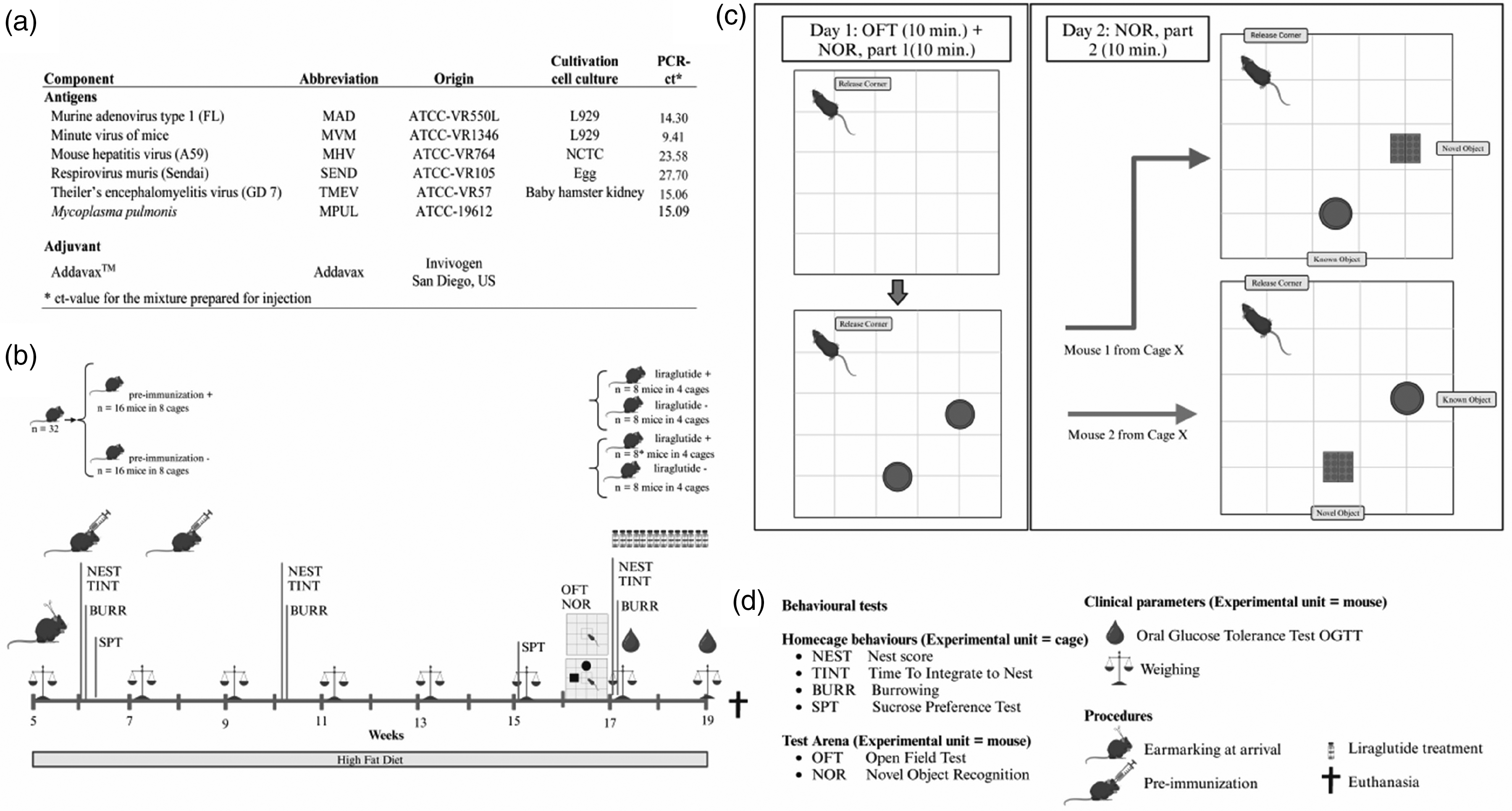
Study design for testing the impact of pre-immunization on treatment with liraglutide and behaviour in diet-induced obese C57BL/6NTac mice. (a) Inactivated pathogens. (b) Time line. (c) Open field test (OFT) and Novel Object Recognition test (NOR) and (d) Text explanations. *One mouse euthanized due to obstipation three days after start of intervention. Created with Biorender.com. PCR: polymerase chain reaction-cycle threshold.
This pre-immunization significantly increased the response in a mouse skin inflammation model. 11 Therefore, we wanted to test it in an even more commonly applied model with a higher potential for reduction in global animal use. In 2022 the European Union used 195,776 mice for studies in metabolism and endocrinology. 12 One of the most common models is the diet-induced obese (DIO) mouse. 13 When fed a high fat diet male C57BL/6 mice become obese, while the females seem to be resistant. 14 Adipocyte-produced TNF-α block of the insulin receptor 15 drives glucose intolerance in both humans 16 and DIO mice, 17 which is, therefore, highly linked to low grade inflammation. 18 Weight, glucose tolerance and long term blood glucose correlate significantly to the levels of TNF-α, interleukin (IL)-12 and IL-10. 18 Hypothetically, pre-immunization may, therefore, influence the power of DIO model studies. If it promotes the development of glucose intolerance as observed in conventional facilities compared with SPF facilities, 19 the therapeutic window would increase. However, pre-immunization may also increase variation. 11
Globally high selling glucagon-like peptide-1 (GLP-1) receptor agonists, such as liraglutide and semaglutide, 20 activate the GLP-1 receptor. 21 This slows gastric emptying, inhibits the release of glucagon and stimulates insulin production. 21 As this leads to increased glucose tolerance, and reduced hyperglycaemia, food intake and body weight, 21 these drugs have become favourites against human type 2 diabetes and obesity. 22 Their effect in DIO mice is highly comparable to that in humans. 23
Systemic immune activation may affect mouse behaviour. Intraperitoneal lipopolysaccharide (LPS) injections may alter behavioural responses, 24 the preference for sucrose, 25 and locomotor activity in mice. 26 A neonatal hepatitis B vaccination impaired the behaviour and neurogenesis of mice. 27 Hypothetically, pre-immunization in mice may, therefore, influence behaviour. The DIO mouse model with longer periods of low experimental activity offers a possibility of testing behaviour without involving further mice.
We aimed to study whether a previously validated pre-immunization regime with inactivated pathogens would improve the power of intervention studies in the DIO mouse model. Therefore, we induced the DIO model in male pre-immunized or control C57BL/6NTac mice subsequently treated with liraglutide. We hypothesized that pre-immunization prior to the induction of obesity would increase power, when testing the liraglutide effect on weight loss as primary read-out. Secondarily, we hypothesized that open field behaviour, sucrose preference, burrowing and nest building behaviours would not be affected by pre-immunization.
Animals
The studies were approved by the Animal Experimentation Council under the Animal Experimentation Inspectorate, Ministry of Food, Fisheries, and Agriculture in Denmark (licence number 2017-15-0201-01262), in accordance with the Danish Animal Experimentation Act (Act no. 63 19/01/2024) and the European Union Directive 2010/63/EU on the protection of animals used in scientific research. Health monitoring, following international guidelines, 3 revealed no indications of listed infections. The study was planned in compliance with the PREPARE guidelines, 28 and it is reported in compliance with the ARRIVE guidelines. 29 Power analysis, taking into account effect and variation experienced in previous studies on murine pre-immunization 11 and liraglutide intervention in DIO mice, 30 determined group sizes.
In total, 32 five-week-old male C57BL/6NTac mice (Taconic Biosciences; Lille Skensved, Denmark) were randomly assigned to pairwise housing in open-type 1284L Eurostandard II L cages (Techniplast, Varese, Italy) at the University of Copenhagen’s AAALAC-accredited barrier-protected experimental rodent facility. The facility controlled temperature at 22°C ± 2°C, a relative humidity of 45–55% and a 06:00–18:00 h light–dark cycle with lights on at 06:00 h. Cages contained aspen bedding (Tapvei, Harjumaa, Estonia), a handling tube (Datesand, Stockport, UK), cotton nestlets (Ancare, Bellmore, NY, USA), an aspen mouse-sized chew block (Tapvei, Harjumaa, Estonia), a play tunnel and a cardboard house (Brogaarden, Lynge, Denmark). Cages were cleaned and provided with new bedding and nest material weekly. Enrichment items were replaced only if shredded by the mice. The mice had unrestricted access to high fat diet (Research Diets D12492, New Brunswick, NJ, USA) and tap water throughout the entire housing period. Daily clinical observation did not reveal any symptoms, except for one less active liraglutide treated non-immunized mouse euthanized three days after intervention start, and at necropsy diagnosed with obstipation.
Materials and methods
Study setup
Pre-immunization
The infectious agents were inoculated on media (Figure 1(a)) at QM diagnostics and were harvested and suspended in PBS buffer (concentrations in Supplementary material Table S1 online) except for murine respirovirus, for which allantois fluid supernatant was used. Suspensions were frozen at −80°C. The vials were transported on dry ice to Copenhagen, where each of the suspensions were suspended in PBS to obtain 10 µg per 50 µl of each pathogen (pathogen-mix) and transferred aseptically to sterile 9 cm petri dishes and subjected to 120 s ultraviolet irradiation at an intensity of 100 J/m2 (CL-1000 Ultraviolet crosslinker UVD; Fisher Scientific, Roskilde, Denmark) and stored at −80°C. Prior to administration, the pathogen-mix was mixed in a 1:1 ratio with adjuvant (Figure 1). The mixture was sent back to QM Diagnostics to estimate the Ct-value for each pathogen (full PCR protocol in the repository). Sixteen mice were pre-immunized subcutaneously at the ages of six and eight weeks with 10 µg of each pathogen as 50 µl of pathogen-mix + 50 µl of adjuvant (Figures 1(a) and (b)). Control mice were injected subcutaneously with a similar amount of PBS.
Liraglutide treatment
Liraglutide diluted 1:49 in sterile filtered dilution buffer (50 mM Na2HPO4 · 2H2O, 70 mM NaCl and 0.05% polysorbate 80, pH adjusted to 7.5) were injected subcutaneously (0.4 mg/kg) in eight mice in each of the two groups once daily for two weeks at 15:00–17:00 h in volumes of 0.1, 0.125 or 0.15 ml depending on their body weight: 20–30 g, 31–40 g or 41–50 g, respectively (Figure 1(b)). To avoid anorexia in the first two days of intervention, the animals received only half a dose of liraglutide (1:99 dilution). The remaining eight mice in each group received the corresponding amount of dilution buffer.
Characterization of obesity and type 2 diabetes
All mice were weighed at arrival and every second week until termination. Oral glucose tolerance was tested before and after liraglutide intervention (Figure 1(b)). Fasting glucose (t = 0) was measured (Freestyle Mini Glucometer, Hermedico, Copenhagen, Denmark) in mice fasted for 6 h (from 07:00 h) after tail vein puncture followed by gavage with 2 mg/g BW glucose (500 mg/ml glucose solution, Amgro I/S, Copenhagen, Denmark) and subsequent blood glucose monitoring after 15, 30, 60, 90 and 120 min.
Behavioural tests and observations
Home cage behaviours
Home cage behaviour was assessed as nesting behaviour and burrowing behaviour at 6, 10 and 17 weeks of age and sucrose preference at 6 and 15 weeks of age (Figure 1(b)).
Nest score
The nests were scored prior to the weekly cage change, where the nests were well-established. The nests were scored from 0 to 5 using a previous scoring protocol, 31 in which the fundamental basic shape of the ultimate, optimal nest is regarded as a sphere with a mouse-sized opening (Table 3).
Pre-immunization with inactivated pathogens in C57BL/6NTac mice was hypothesized not to influence nest scoring, burrowing and sucrose preference as expression of welfare-compromising, pain and depression causing conditions. Differences are non-significant between the behaviours of pre-immunized and control mice.
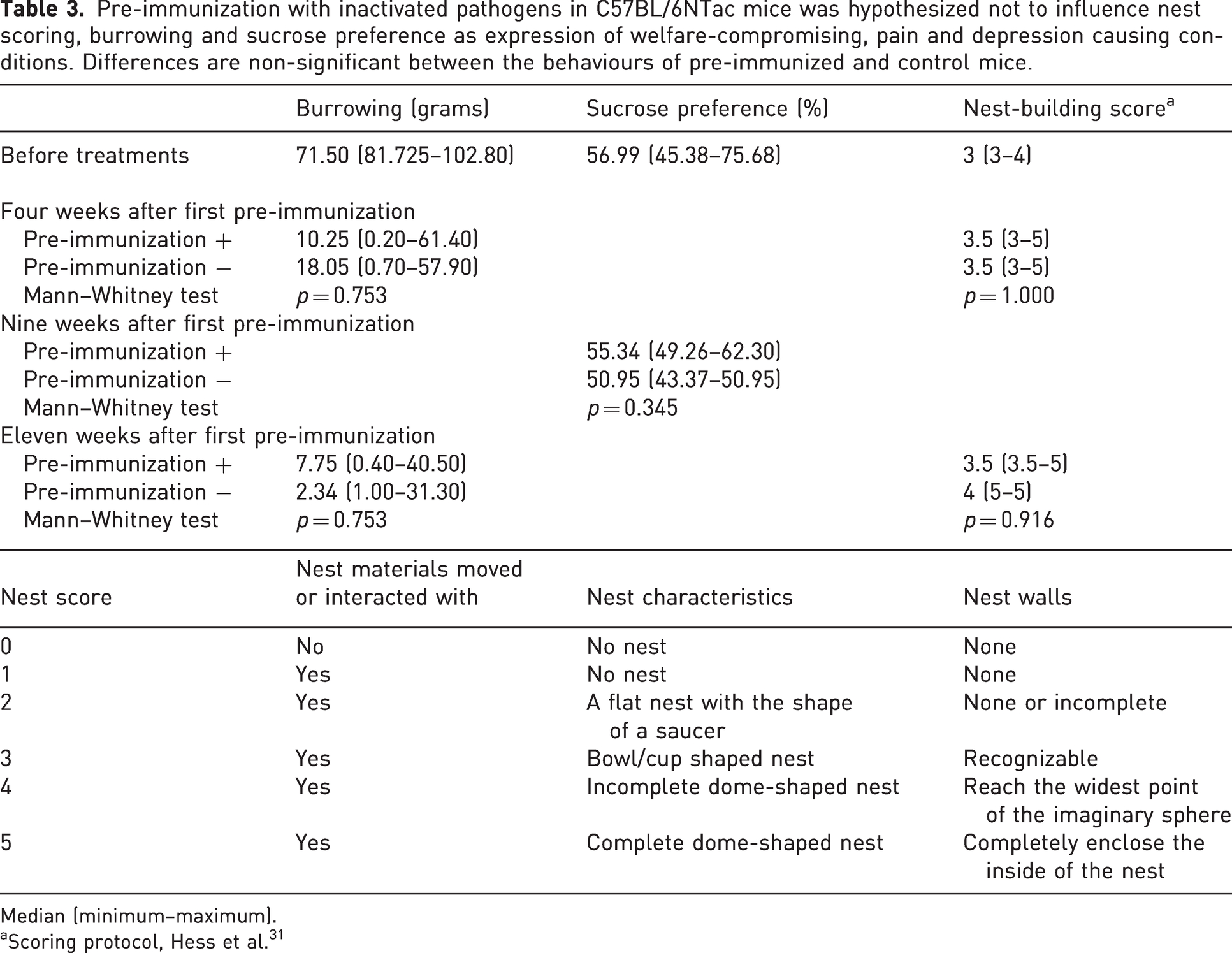
Median (minimum–maximum).
Scoring protocol, Hess et al. 31
Time-to-Integrate-into-Nest Test (TINT)
A one-quarter nestlet cotton-fibre square (5 cm × 5 cm) was, within the first 4 h of the light phase, placed in each cage in the corner opposite the existing nest and visible from the outside. The cages were left unattended for 10 min whereafter the observer noted whether the nestlet had been manipulated/moved. When the square had been moved from the start position, the test was scored as positive, and if not the test was scored as negative.
Burrowing behaviour
The mice were introduced to burrowing aspen bedding filled tubes for 24 h prior to testing. On the test day, the mice were given a non-transparent burrowing tube (length, 20 cm; diameter, 6.8 cm; closed at one end and raised 3 cm at the other end) filled with 100 g aspen bedding in the back left corner of the cage. The cardboard house, handling tube and all nesting material were removed from the home cage and given back after the test. After 2 h, the remaining bedding in the tube was weighed and the amount burrowed/removed was calculated.
Sucrose preference test
A fresh 1.0% (w/vol) sucrose solution (Sigma-Aldrich, cat. no. S8501, Søborg, Denmark) was prepared daily. It was kept at 4°C and at test start warmed to 22°C ± 2°C. Food or water deprivation was not applied at any time. After one week of habituation with two bottles of tap water, the mice were habituated to two bottles of 1% sucrose solution. Hereafter, they were subjected to a 48-h choice test with continuous access to two bottles (tap water and 1% sucrose solution) randomly placed and switched after 24 h to avoid side preference in drinking behaviour. The bottles were weighed before and after the test and sucrose preference calculated as a percentage of the consumed sucrose solution from the total amount of liquid consumed, that is: preference = (sucrose intake/total intake) × 100%. 32
Open field test and Novel Object Recognition (NOR) test
Fear level and cognitive abilities were assessed on video recordings using an open field test and a NOR test at 16 weeks of age (Figure 1(b) and (c)). Prior to test start, mice in their home cages were acclimatized to the test room for 30 min. Over two consecutive days an open field test was followed by introduction to two identical objects on test day 1 and the NOR test was performed on test day 2. The open field test additionally functioned as a 10-min habituation to the test arena prior to the NOR test (Figure 1(c)). On day 1 the mouse was transferred by a handling tube to the centre of the open field arena (50 cm × 50 cm). The mouse was allowed free exploration for 10 min, removed from the arena using the handling tube while two identical green glass tealight holders (diameter 7.5 cm and height 5 cm) (Rosendahl, Hørsholm, Denmark) were placed in the arena. The mouse was returned to the opposite corner of the arena (the release corner), facing the corner and allowed to explore the two objects for 10 min. On day 2 all mice in their home cages were allowed 30 min of habituation to the test room. One clean glass tealight holder and four assembled clean Lego blocks (Lego Duplo®, Billund, Denmark; 6.3 cm × 3.1 cm × 2.3 cm, forming a 6.3 cm × 3.6 cm × 4.3 cm square object) to function as the novel object were placed in the arena. From each pair/cage one mouse had the novel object placed randomly to the right, while the other mouse had it to the left. The mouse was placed in the release corner using tunnel handling. After being allowed to explore the objects for 10 min, the mouse was returned to his cage using the tunnel. The arena was cleaned with 70% ethanol and allowed to dry before each mouse introduction.
Euthanasia
At 19 weeks of age all mice were euthanized by cervical dislocation after 0.1 ml/10 g body weight of a mixture of 25% Midazolam (5 mg/ml midazolam, B. Braun, Melsungen, Germany) and 25% Hypnorm (0.315 mg/ml of fentanyl citrate and 10 mg/ml of fluanisone, Skanderborg Apotek, Denmark) subcutaneously.
Exclusion, humane endpoints and blinding
No exclusion criteria were set prior to the study. Humane endpoints were reduction in food/water intake and activity, more than 20% weight loss compared with expectable growth, and reduced well-being and illness. The observers/test operators were blinded to the treatment, and animals were randomly assigned to the test order. The facility manager, not taking part, was aware of treatments.
Statistics
Statistical tests are summarized in Table 1. Quantitative data were tested for normality and equality of variances. Weight loss in the control mice as well as burrowing, sucrose, nest building and memory data did not follow a normal distribution and, therefore, these parameters were tested with a Mann–Whitney test. Power analysis for weight loss was done on ranked data.
Statistical tests applied for testing the impact of pre-immunization with inactivated pathogens diet-induction and subsequent liraglutide treatment of male C57BL/6NTac mice.

GraphPad Prism, Boston, MA, USA.
Minitab, Coventry, UK.
Two-tailed test.
One-tailed test.
Power = 0.90, p < 0.05.
OGTT: oral glucose tolerance test; TINT: Time-to-Integrate-into-Nest Test; DIO: diet-induced obese
Results
Studying the effect of liraglutide had more power in pre-immunized mice
Pre-immunized mice were weighed every second week both during induction of obesity and after treatment with liraglutide, and oral glucose tolerance was tested before and after liraglutide treatment (Figure 1). Pre-immunization did not influence the induction of the model, as there was no difference in effect or variation in weight development (p = 0.6088) (Figure 2(a)), fasting blood glucose (p = 0.6803) (Figure 2(b)) or glucose tolerance (p = 0.6954) (Figure 2(c)) between pre-immunized and control mice during diet induction. Liraglutide only in the pre-immunized mice caused a significant weight loss (p = 0.0009) and reduction of fasting blood glucose (p = 0.0106), while these were non-significant for the non-immunized mice (p = 0.1598; p = 0.0732; Figure 2(d) to (f)). Liraglutide compared with placebo significantly improved the glucose tolerance for both the pre-immunized (p = 0.0007) (Figure 2(g)) and the control mice (p = 0.0389) (Figure 2(h)). Therefore, it was expectable that pre-immunization may reduce the number of mice used in such intervention studies. A power analysis for a target power of 0.90 and p < 0.05 showed that a significant effect of liraglutide on weight could have been shown with only five pre-immunized mice in each group, while 95 non-pre-immunized control mice would be needed to obtain a significant p-value (Table 2). Effect on glucose tolerance and fasting blood glucose could have been shown with four and five pre-immunized instead of 24 and 15 non-immunized control mice, respectively (Table 2).
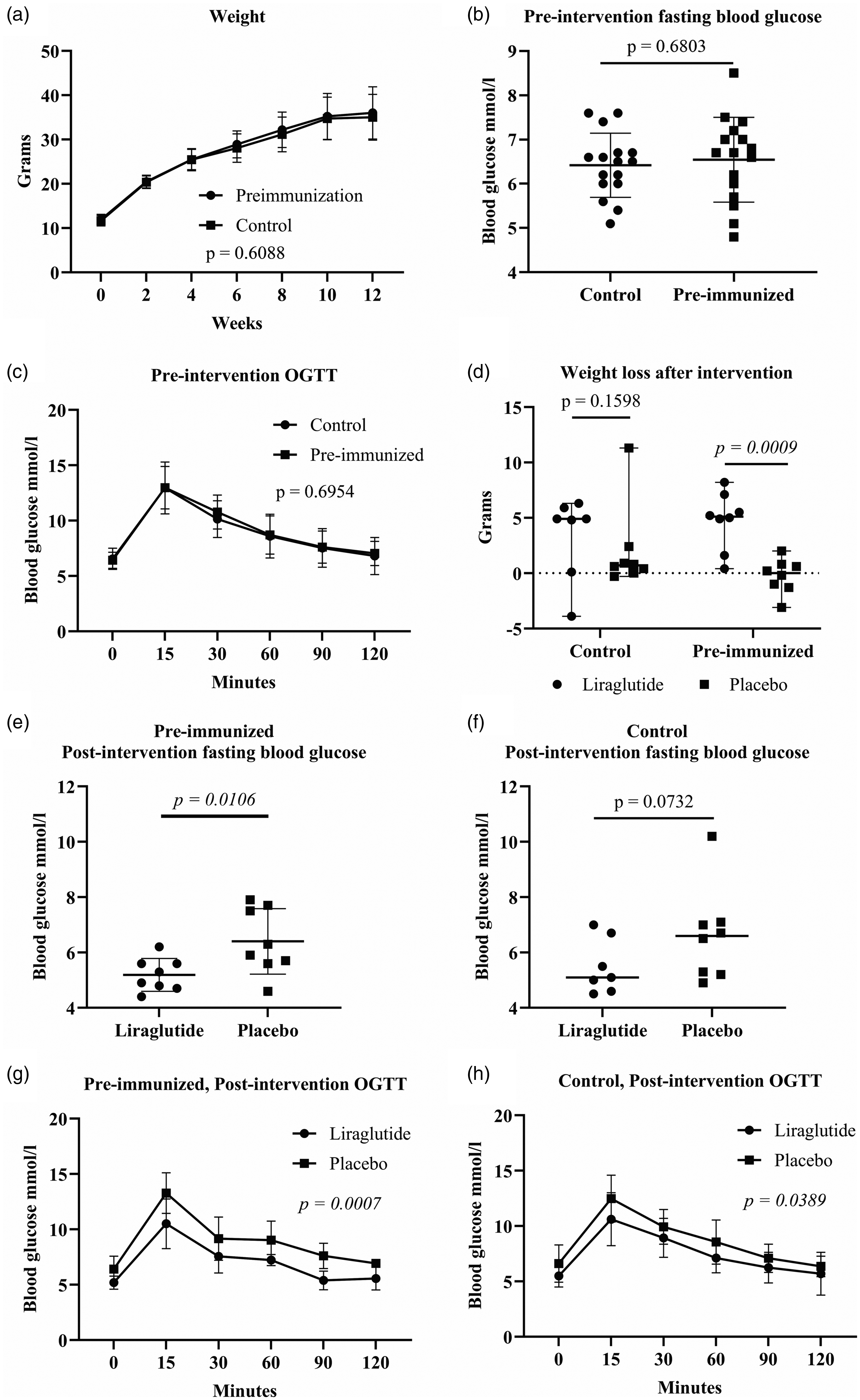
Pre-immunization with inactivated pathogens prior to the induction of obesity was hypothesized to increase power when liraglutide was tested in male diet-induced obese C57BL/6NTac mice. Weight development (a), fasting blood glucose (b) and oral glucose tolerance test (OGTT) (c) did not differ between pre-immunized and control mice during diet induction of obesity. Liraglutide treatment induced significant weight loss (d), lower fasting blood glucose (e and f), and improved glucose tolerance (g and h) only in the pre-immunized mice and not in the control mice. (a) to (c) and (e) to (h), mean ± SD; (d), median ± range.
Actual power obtained by a group size n = 8, and group sizes needed to show an effect (differences between before and after treatment) in a Student’s t-test of liraglutide on weight, the area under curve of an oral glucose tolerance test (OGTT-AUC), and fasting blood glucose in male diet-induced obese C57BL/6NTac mice either pre-immunized with inactivated pathogens or acting as controls.
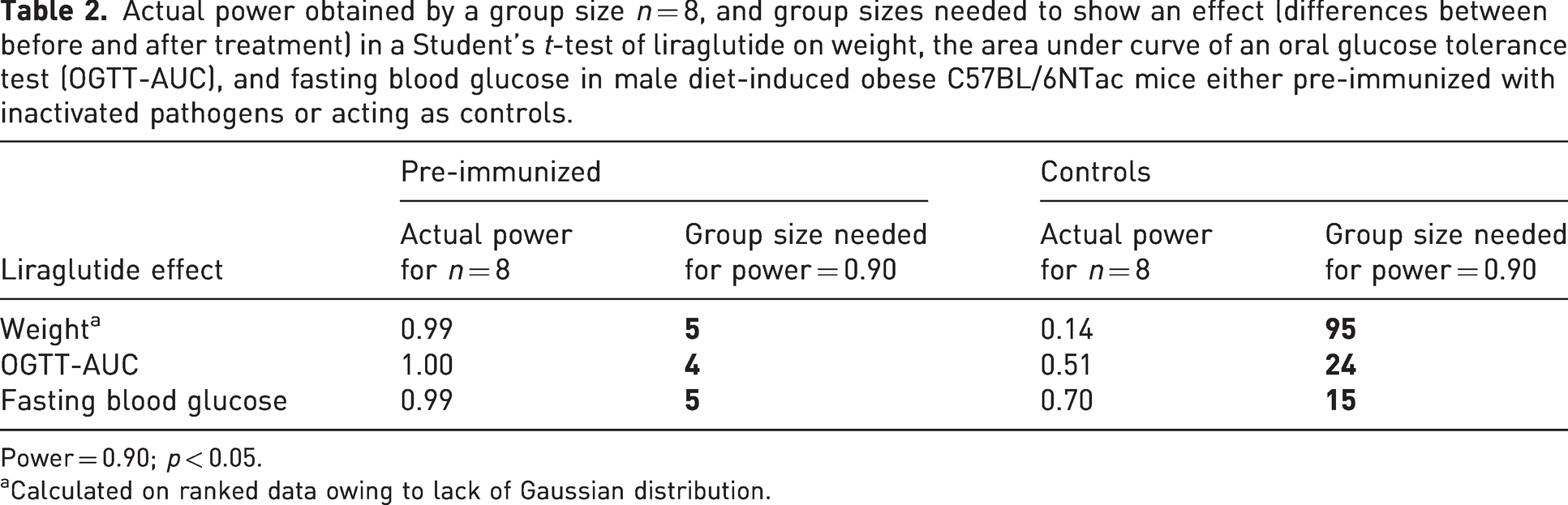
Power = 0.90; p < 0.05.
Calculated on ranked data owing to lack of Gaussian distribution.
The behaviour of the mice was unaffected by the pre-immunization
To assess depression-like changes and signs of anhedonia (lack of motivation for stimuli previously considered pleasurable), 33 sucrose preference test was applied one week after pre-immunization (Figure 1(b)). To monitor changes in cognitive behaviours a NOR test was done adjacent to the open field test 11 weeks after pre-immunization (Figure 1(b) and (c)). One, five and 12 weeks after pre-immunization, nest building behaviour was assessed (Figure 1(b)) to assess first the quality of the nest using a nest score, 34 followed by TINT, that is, a measure of the time it takes the mice to integrate new nest material into an existing nest. 34 Nests scoring and a TINT have been shown to be negatively affected by welfare-compromising conditions such as, for example, postoperative pain and experimental colitis. 34 Burrowing was assessed as a spontaneous, highly motivated and species-specific behaviour affected by various pain and stress factors (Figure 1(b)). 35
There were no differences in burrowing, sucrose preferences or nest building behaviour between pre-immunized and control mice (Table 3), indicating that the pre-immunization did not induce pain, distress or depression in the mice. Pre-immunized did not differ from control mice in their latency to move or touch the two unknown objects in the open field arena (Figure 3(a) to (c)) or in their duration and number of crawls on the objects (Figure 3(a) to (e)). However, there was a significantly higher variation in pre-immunized compared with control mice for latencies to touch objects (Figure 3(b)) (p = 0.0057), the total duration of crawling behaviour (Figure 3(d)) (p = 0.0004) and the total number of crawls (Figure 3(e)) (p = 0.0065). Memory coefficients did not differ between pre-immunized and control mice (p = 0.4062), that is, pre-immunization did not cause any memory deficiencies in the mice (Figure 3(f)). As expected, the number of touches on novel and known objects differed significantly (p = 0.0164), that is, the mice had a normal memory. There were no differences in TINT (Supplementary Table S2).
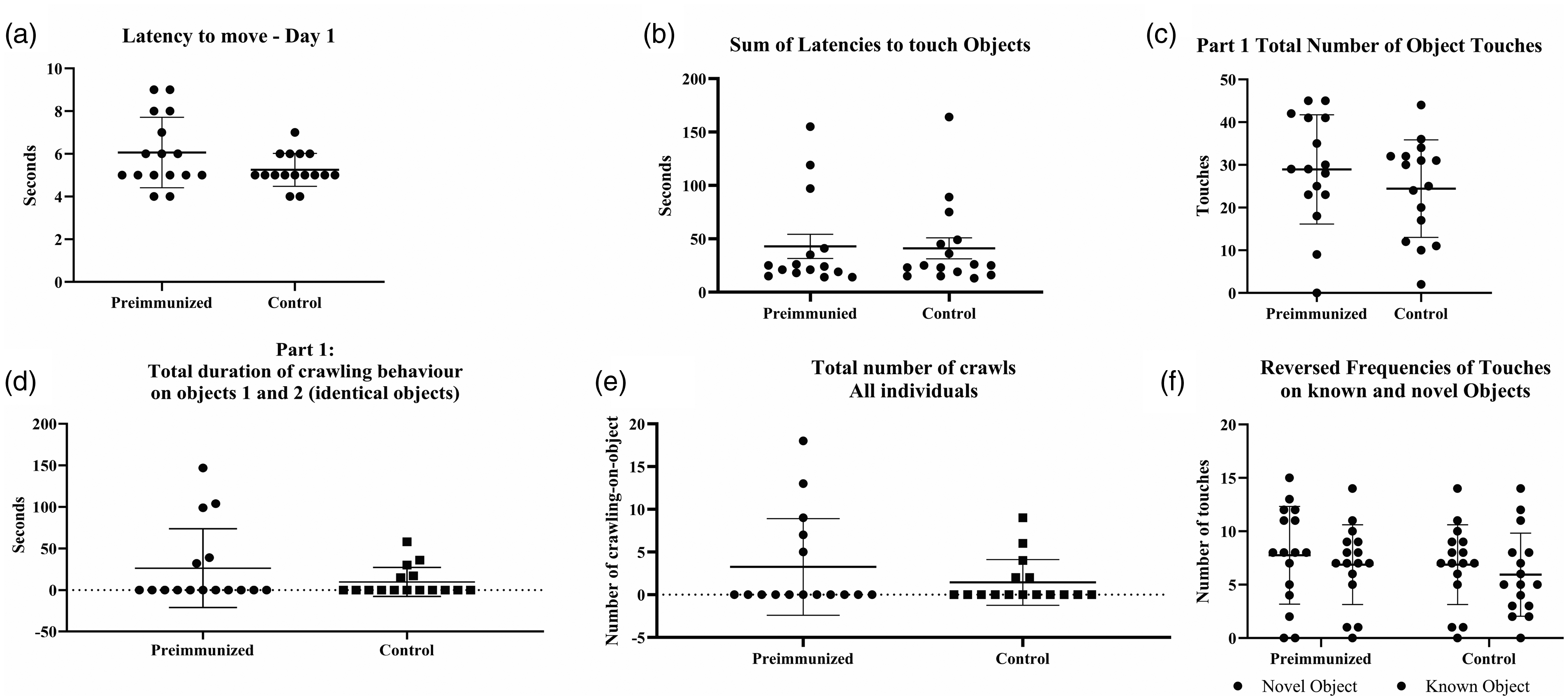
Pre-immunization with inactivated pathogens in C57BL/6NTac mice was hypothesized not to influence exploratory behaviours in a Novel Object Recognition test 11 weeks later. There were no differences in latency to move in the arena (a), latency to touch the objects (b), number of touches on the objects (c), duration of crawling on the objects (d) and number of crawls on the objects (e), as well as reversed frequencies of touches on known and one novel object on day 2 (f). However, pre-immunized compared with control mice had a significantly higher variation for latencies to touch objects (b) (p = 0.0057), the total duration of crawling behaviour (d) (p = 0.0004) and the total number of crawls (e) (p = 0.0065).
Discussion
Pre-immunization increased the power of intervention with liraglutide in the DIO mouse model for the key parameters of liraglutide treatment, that is, reduced weight, improved glucose tolerance and reduced fasting blood glucose, compared with the non-immunized control mice. For the pre-immunized mice, the power for all three parameters was close to 100%, and fewer mice than eight could have been used to obtain the target power of 90%, while for the non-immunized control mice more mice would be needed.
‘Dirty’ mice in some research areas may lead to more translational results.4,6,7 So far, ‘dirty’ mouse creation implies that the animals are infected with active pathogens,4,6,7 which, in addition to the risk of spreading these to clean SPF animals, also means that the infection state will vary between the different pathogens and the individual animals. In contrast, pre-immunization with inactivated pathogens is done at well-defined time points in relation to the experiment and the age of the animal. The present pre-immunization regime has previously been validated. 11 This showed its capability of inducing a CD8+ effector memory T cell response, 11 as is observed in feral and laboratory mice co-housed with pet-shop mice. 4 In a skin inflammation mouse model this pre-immunization regime significantly increased primary read-out cytokine responses. 11 Interestingly but not surprisingly, these five-fold higher responses also came with an increase in the inter-individual variation. 11 Inter-individual variation did not differ in our primary DIO readouts, that is, weight, glucose tolerance and fasting blood glucose, but the differences in effect due to pre-immunization, although enough to increase the power, were also more discrete than in the skin inflammation model. 11
The DIO mouse model is under strong influence of the host microbes, which includes bacteria, phages and viruses. 36 Most of these are opportunists or symbionts. Some specific bacteria are important for expressions in other animal models, for example, as known in non-obese diabetic mice for the anti-inflammatory mucin-degrader Akkermansia muciniphila, 37 which is also promoted by liraglutide, 38 and reduced by DIO induction. 18 Liraglutide greatly reduces the Firmicutes/Bacteroidetes ratio. 38 Incorporation of microbiota profiles in the data evaluation in liraglutide intervention studies in C57BL/6NTac mice may reduce the group sizes by up to 50% for showing the effect of liraglutide on weight. 30 In contrast to C57BL/6NTac mice, C57BL/6NRjj mice do not lower their long term blood glucose as a response to liraglutide treatment, but the microbiota is also much different between mice from Taconic and Janvier. 30 For instance, C57BL/6NRjj mice carry the anti-inflammatory Bifidobacterium spp., which C57BL/6NTac mice do not. 30 DIO induction changes the microbiota, but to a much lower degree than the basic differences in microbiota of mice of the same strain from different breeders. 36 The microbiota of wild mice is much higher compared with commercially available laboratory mice, the microbiota of which is dominated by Firmicutes and Bacteroidetes, while wild mice, at the cost of Firmicutes, have a large fraction of Proteobacteria. 8 This more complex nature of the microbiota in ‘dirty’ mice is probably also important for some of the responses observed, and, if so, this is not addressed by our pre-immunization regime in contrast to wildlings, 7 pet-shop mice co-housed 4 and feralized mice. 6
The EU statistics report 12 does not list the number of DIO models among the 195,776 mice used for metabolism and endocrinology research, but it is a highly common model. 13 Therefore, the application of such simple tools as pre-immunization with inactivated pathogens and incorporation of microbiota composition in the data evaluation 30 might reduce animal use substantially. Compared with the maintenance of ‘dirty’ mice with active pathogens, pre-immunization with inactivated pathogens is a far easier approach to handle in modern facilities. From an animal welfare point of view, pre-immunization in contrast to co-housing with pet-shop mice 4 does not increase mortality.
It is a limitation that we do not know the exact mechanism by which pre-immunization influences the liraglutide effect and exactly which pathogens and host responses are responsible for the effect. The present and a previous study 11 showed effect of the pre-immunization regime on C57BL/6N mice and BALB/c mice from Taconic and BALB/c mice from Charles River, but as the microbiota differs between the same sub-strain from different breeders, 36 further studies are needed to show the general applicability.
The pre-immunization did not seem to influence the behaviour of the mice, but there was a significant increase in the variation of their exploratory behaviour, which might reduce the power of some behavioural studies.
In conclusion, pre-immunization with inactivated pathogens may be a simple tool to increase power in intervention studies on the DIO mouse model.
Supplemental Material
sj-pdf-1-lan-10.1177_00236772241279058 - Supplemental material for Pre-immunization of diet-induced obese male mice with inactivated pathogens increases power in a liraglutide intervention study
Supplemental material, sj-pdf-1-lan-10.1177_00236772241279058 for Pre-immunization of diet-induced obese male mice with inactivated pathogens increases power in a liraglutide intervention study by Caroline Falkenberg, Dorte B Sørensen, Camilla HF Hansen, Martin F Toft and Axel K Hansen in Laboratory Animals
Supplemental Material
sj-pdf-2-lan-10.1177_00236772241279058 - Supplemental material for Pre-immunization of diet-induced obese male mice with inactivated pathogens increases power in a liraglutide intervention study
Supplemental material, sj-pdf-2-lan-10.1177_00236772241279058 for Pre-immunization of diet-induced obese male mice with inactivated pathogens increases power in a liraglutide intervention study by Caroline Falkenberg, Dorte B Sørensen, Camilla HF Hansen, Martin F Toft and Axel K Hansen in Laboratory Animals
Footnotes
Data availability statement
Declaration of conflicting interests
CF declares that she for previous projects has been funded by commercial enterprises. CHFH declares that she has collaborated with the pharmaceutical industry and received funding from this source as described on https://ivh.ku.dk/english/employees/?pure=en/persons/306048. DBS declares that she has collaborated with the pharmaceutical industry and received funding from this source as described on https://ivh.ku.dk/english/employees/?pure=en/persons/114880. MFT is an employee at QM Diagnostics. AKH declares that he has collaborated with pharmaceutical industry and received funding from this source, as well as he is the owner of a diabetes related patent as described on https://ivh.ku.dk/english/employees/?pure=en/persons/107126.
Funding
This study was partly funded by the Centre for Applied Laboratory Animal Research (www.calar.dk) which is a collaboration between Novo Nordisk A/S, Lundbeck A/S and University of Copenhagen.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.