
Abstract
The thyroid gland’s proper function is essential for controlling the metabolism, which varies to some extent between the sexes, thus maintaining the homeostasis. This research aimed to investigate differences in the histological structure and number of mast cells in the thyroid gland of male and female rats, and to establish whether there is a correlation between the levels of thyroid-stimulating hormone (TSH) and the activation index (Ia), which quantifies the functional activity of the gland based on histological parameters. Thyroid glands of 20 Wistar rats were analyzed morphometrically and stereologically to determine follicle size, distribution, and volume density of the epithelium, colloid, and connective tissue. Male thyroid glands had more small and medium follicles and a higher volume density of epithelium, while female thyroid glands had more large follicles and a higher volume density of connective tissue. The volume density of colloid was not significantly different between the sexes. The histological structure of the thyroid glands in both groups was in accordance with the measured TSH levels. The correlation between TSH serum levels and Ia were established in both sexes. Both the TSH serum levels and Ia were lower in females compared to males. A higher number of mast cells was noted in the connective tissue of female thyroid glands, compared to those of males, but this difference was not significant. The results of this study indicate that there are differences in the histological structure of male and female rat thyroid glands and that Ia can be used as a reliable complementary parameter for assessment of thyroid gland function.
Introduction
In the field of clinical practice, understanding the differences in baseline physiology between the sexes and the mechanisms underlying these disparities is crucial for elucidating the sex-related aspects of disease pathogenesis, treatment responses, and outcomes. 1 These differences are equally significant in preclinical research, where they should be considered as an experimental factor to enhance the reproducibility and translational relevance of results, and to avoid possible biases. 2 Given the central role of the thyroid gland in maintaining homeostasis, the higher prevalence of thyroid dysfunction in women, 3 and the continued use of rats as the most common small animal model in preclinical studies after mice, investigating sex differences in the histological architecture of the rat thyroid gland under physiological conditions holds significant scientific and translational importance.4,5
Thyroid hormones, primarily thyronine (T3), thyroxine (T4), and calcitonin, regulate the activity of enzymes and proteins, which control cellular metabolism. 6 Additionally, thyroid gland hormones’ vital role in the fetal development of various organ systems, primarily nervous, but also other systems including the skin, bones, and urogenital system has been demonstrated.7–11 The levels of T3 and T4 are under the control of thyroid-stimulating hormone (TSH) from the pituitary gland, which is itself regulated by a thyrotropin-releasing hormone (TRH) from the hypothalamus. 12 Thyroid hormones influence the hypothalamic-pituitary-thyroid axis through a negative feedback mechanism, whereby increased circulating T3 and T4 suppress TRH and TSH secretion, maintaining hormonal balance. Conversely, decreased levels stimulate increased TSH secretion, leading to thyroid gland stimulation and growth. 13
In order to quantify the functional activity of the thyroid gland based on histological parameters, the thyroid gland activation index (Ia) is utilized. This index is calculated as the ratio of the volume or number of follicular epithelial cells relative to the colloid content within the gland, providing an indicator of the gland’s secretory activity and functional state. 14 A higher Ia generally reflects increased gland activity, often associated with elevated TSH levels and a stimulated histological appearance, such as hypertrophy of follicular epithelium and reduced colloid storage.
Disruptions in thyroid function can be classified as primary (affecting the gland itself), secondary (at the pituitary level), or tertiary (at the hypothalamic level). 12 These functional abnormalities are frequently related to environmental and lifestyle factors, including exposure to chemicals and pollutants that act as endocrine disruptors. Interestingly, studies involving animal models, such as those conducted by Marić et al., indicate that female rats are more sensitive to the thyrotoxic effects of such endocrine disruptors, particularly in response to metal mixtures, suggesting sex-related differences in susceptibility. 15
The morphology of the thyroid gland, including follicle size, distribution, and the volume density (Vv) of its structural components—epithelium, colloid, and connective tissue—provides valuable insights into its functional state. 16 These parameters are directly linked to hormone synthesis, storage, and secretion. For instance, enlarged follicular epithelium may indicate increased activity, whereas increased colloid volume density suggests a decreased secretory demand. 16 Evaluating these parameters allows a comprehensive assessment of the gland’s physiological status and its adaptive responses to various stimuli.
Furthermore, immune system components, particularly the mast cells present within the thyroid stroma, are increasingly recognized as modulators of glandular activity. Mast cells are involved in immune responses and have been proposed to influence TSH responsiveness and glandular remodeling, especially under pathological conditions.17–19 The distribution and function of mast cells exhibit sex differences, attributed in part to hormonal influences such as androgens and estrogens, which may contribute to the higher incidence of autoimmune thyroid diseases in females.20,21 Understanding these differences is thus of high translational relevance, as sex-specific mechanisms can inform targeted therapeutic strategies.
This study compares the histological structure of the thyroid gland in male and female rats by analyzing follicle size, the presence of follicles of different sizes (small, medium, large), and the volume density (Vv) of thyroid gland structural components (epithelium, colloid, connective tissue). The study also calculates the thyroid gland Ia as a measure of gland activity, compares these values with serum TSH levels, and explores correlations between histological and hormonal parameters. The prevalence of mast cells within the connective tissue is also assessed in both sexes, contributing to a better understanding of the sex-specific structural and functional characteristics of the thyroid gland, in order to enhance understanding of the importance of sexual differences in the preclinical research of thyroid gland function and disorders.
Animals, materials, and methods
Experimental animals
The thyroid glands studied were sampled from twenty 6-month old Wistar rats (10 males and 10 females). These animals were part of the control groups in an experiment examining the impact of endocrine disruptors on the neuroendocrine system. 15 All experimental procedures were approved by the Ethics Committee for Experimental Animal Research of the Faculty of Pharmacy, University of Belgrade; and the Ministry of Agriculture, Forestry, and Water Management–Veterinary Directorate (ethical permit number: 323-07-11822/2018-05). Animal handling/housing was conducted in strict accordance with the ARRIVE guidelines 2.0 (https://arriveguidelines.org/arrive-guidelines) and with the institutional Guidelines for Care and Use of Laboratory Animals at the Faculty of Pharmacy, University of Belgrade.
Housing and maintenance
After introducing the rats to the laboratory animal facility, the animals were left to acclimate to the new environment for 7 days. The rats were housed in same sex groups of three to four animals in a standard cage system, divided according to the standard for socially compatible individuals. The laboratory animals were housed in a climate-controlled facility (21°C ± 0.5°C) with a 12-h light-dark cycle, and the rats had ad libitum access to food and water. We used pelleted rat food produced by FSH DOO Gebi, Serbia, with a minimum iodine E-2 content of 0.15 mg/kg. Quality control of the rat food was conducted and is guaranteed by the Institute for Animal Husbandry’s laboratory for chemistry and microbiology, Belgrade, Serbia. The male and female control groups did not undergo any treatment. Euthanasia for the experimental animals was conducted in accordance with the AVMA (American Veterinary Medical Association) guidelines for the euthanasia of animals, via intravenous pentobarbital (100 mg/kg body weight; Euthasol Euthanasia Solution, Produlab Pharma Production B.V., Raamsdonksveer, Netherlands), during which blood was collected in heparinized polypropylene tubes (32 U/tube). The animals were always euthanized at 12 p.m. due to the significance of circadian fluctuations in the activity of mast cells and histamine, and serotonin content in the rat thyroid gland. 22 In line with the 3R principle (Replacement, Reduction, Refinement), various organs from both sexes in all experimental groups were sampled and preserved for future research on endocrine disruptors.
Sampling and preparation of the thyroid gland for light microscopy
After sampling, the thyroid gland tissue was fixed in 10% neutral-buffered formalin (pH = 6.8) for 24 h at room temperature. The tissue was then dehydrated in a series of ethanol solutions with increasing concentrations (70%, 96%, and 100%), cleared in xylol, embedded in paraffin, and serially sectioned at a 5 µm thickness with a rotary microtome (Slee Cut 5062, SLEE medical GmbH Nieder-Olm, Germany). The thyroid gland sections were stained following the standard haematoxylin and eosin protocol (Merck Millipore, Darmstadt, Germany) and mounted with DPX medium (Sigma Chemical Co., St. Louis, MO, USA). Additionally, thyroid samples were stained using the Van Gieson method and Azan staining to better visualize the connective tissue, primarily collagen fibers, and to more easily distinguish the connective tissue from muscle fibers; toluidine blue was used for highlighting the mast cells in thyroid gland tissues. The staining procedures for the Van Gieson method, Azan, and toluidine blue staining were performed according to the protocol recommended by Švob. 23
Histological, stereological, and morphometric analyses
Histological analyses were performed using an Olympus CX31 microscope, a digital camera (UC50 Soft Imaging Solutions), and SensEntry1.13 software (Münster, Germany).
The size of follicles was determined by measuring the two longest perpendicular lengths, and then calculating the follicle diameter (fd) as an average of these two lengths. Based on the fd and descriptive statistic parameters, namely the first and third quartiles, the follicles were divided into three categories: small (fd ⩽ 73.27 µm), medium (73.27 µm <fd < 123.73 µm), and large (fd ⩾ 123.73 µm). The measurements were performed on 30 randomly selected follicles for each thyroid gland sample.
Stereological analysis of the thyroid gland was conducted on the serial sections using an M42 multipurpose stereological grid, consisting of 42 test points and 21 test lines over an area of 36.36 mm2. 24 The grid was used to determine the relative stereological parameter, volume density (Vv), which indicates the proportion of the thyroid gland’s total volume occupied by one of its main structures: epithelium (Vve), colloid (Vvc), and connective tissue (Vvct). By knowing the total number of test points in the M42 grid, and the number of those points that fall on each of the structures, we can calculate their Vv using the following formulas: Vve = Pe/Pt, Vvc = Pc/Pt, and Vvct = Pct/Pt, where Pe, Pc, and Pct are number of test points within each structure and Pt is the total number of points (42) in the test system. The number of fields per sample used for assessing the Vv was determined based on the formula devised by Kališnik. 25
The thyroid gland Ia was determined using the formula: Ia = Vve/Vvc.
Mast cells were counted on toluidine blue-stained sections using a low-power (×10) objective across eight fields of view per animal, reported as the number of mast cells per 1 mm2.
Measurement of TSH levels
To determine TSH levels, blood samples were collected via cardiac puncture. The samples were centrifuged at 3000 × g for 30 min to separate the serum, which was then frozen at −20°C and stored until analysis. TSH levels in the rats’ blood were measured using the Rat TSH ELISA Kit (Fine Biotech Co., Ltd.), following the manufacturer’s instructions.
Statistical analysis
Statistical analysis of the results was conducted using GraphPad Prism version 6 (GraphPad, San Diego, CA, USA). The descriptive analysis was used to determine the first and third percentiles. The normality of the data distribution was tested using the Shapiro–Wilk test. The fd of male and female thyroid glands were compared with the Mann–Whitney U test. The chi-square test of independence was used to assess whether there was a relation between animal sex and the presence of different sized categories of follicles in the thyroid gland. The differences in Vv, Ia, TSH levels, and the number of mast cells between male and female rats were evaluated using the Student’s t-test. Pearson’s correlation was used to assess the correlation between TSH levels and thyroid gland Ia in both sexes. Statistical significance was set at p < 0.05 and the results are presented as mean values ± standard deviation.
Results
Histological and morphometric analysis of thyroid gland structure
Microscopic examination of male (n = 10) and female (n = 10) rat thyroid gland tissue sections revealed two laterally positioned lobes on both sides of the trachea. Thin connections, consisting of follicles and connective tissue, were observed between the gland’s lobes in tissue sections cut mid-series. A prominent capsule of dense connective tissue was noted on the thyroid gland’s surface, with trabeculae of both dense and loose connective tissue extending into and dividing the thyroid parenchyma. These structures were more distinct in the sections stained with the Van Gieson and Azan methods. Within the larger-diameter trabeculae, blood vessels and peripheral nerves, featuring both myelinated and non-myelinated fibers, were visible. The male thyroid parenchyma predominantly consisted of small and medium follicles (Figure 1(a)), with epithelium consisting of columnar or high-columnar thyrocytes (Figure 1(a) and (b)). The colloid appeared to be acidophilic and smooth, with small reabsorption vacuoles near the apical surface of the thyrocytes noticeable in some follicles (Figure 1(a) and (b)). In contrast, the female thyroid parenchyma mainly contained large and medium follicles, with epithelium made up of columnar or low-columnar thyrocytes (Figure 1(c) and (d)). The colloid was less acidophilic and largely smooth, with more prominent and larger reabsorption vacuoles compared to males (Figure 1(c) and (d)). The measurements of the follicular diameter confirmed that males had more small and medium follicles, considering that the average diameter was 89.45 ± 25.79 µm (mean ± standard deviation). In contrast, the average diameter of the female thyroid gland follicle was 113.12 ± 44.37 µm (mean ± standard deviation), confirming the histological observation that the parenchyma consists mainly of large and medium follicles. Comparing the follicular diameters of male and female thyroid glands using the Mann–Whitney U test showed that this parameter was significantly greater in female thyroid glands (p < 0.0001) (Figure 1(e)). The chi-square test of independence showed that there was a significant relation (χ2 = 63.08, p < 0.0001) between the sex of rats and presence of small, medium, and large follicles in the thyroid glands (Figure 1(f)).
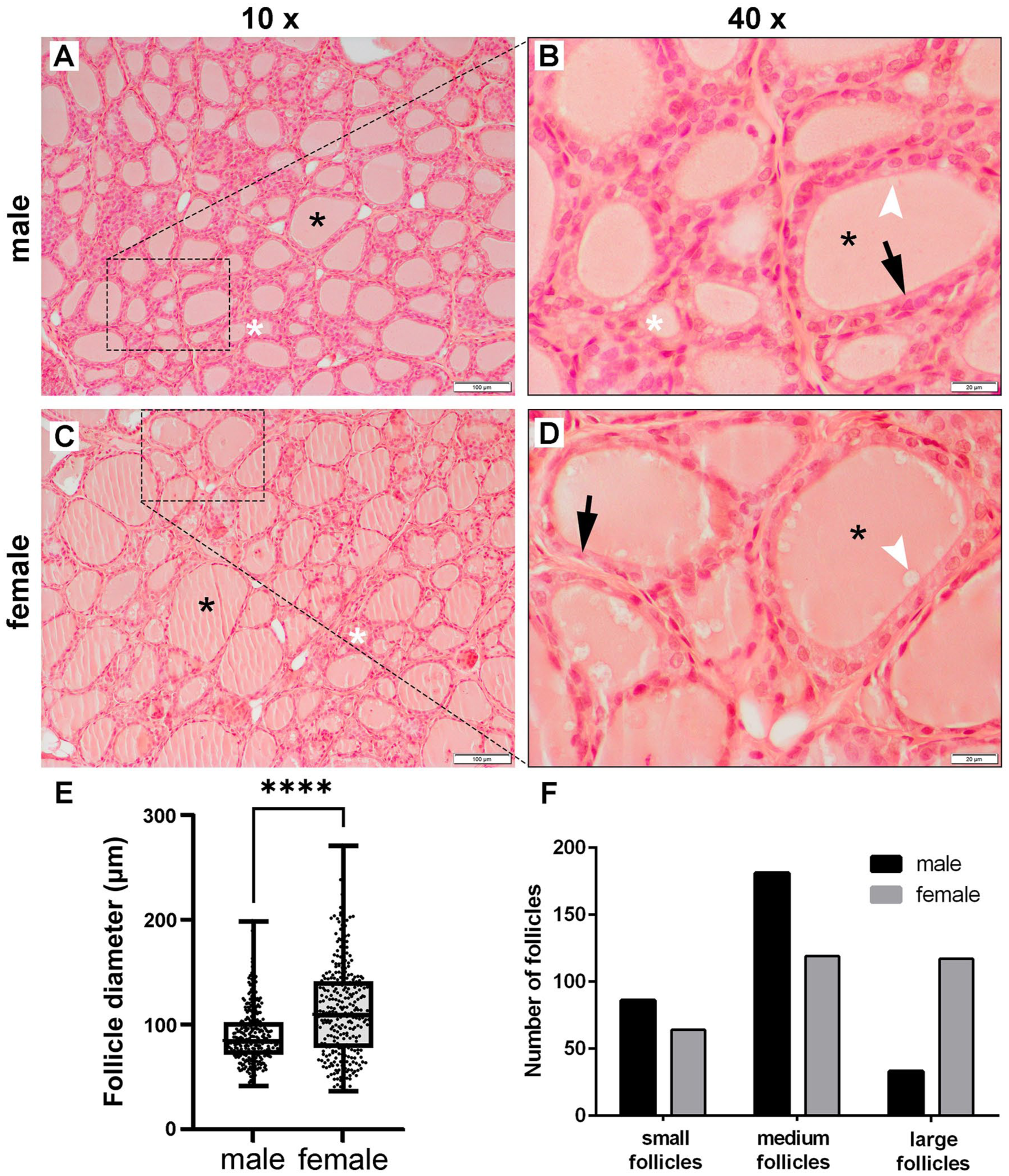
Representative photomicrographs of thyroid glands: (a) and (b) male (n = 10) Wistar rats, and (c) and (d) female (n = 10) Wistar rats. Tissue sections stained with haematoxylin/eosin viewed in (a) and (c) using a low-power (×10) objective lens (bar: 100 μm), and in (b) and (d) with a high-power (×40) objective lens (bar: 20 μm). (e) Graph representing the follicle diameter of male (mean ± standard deviation = 89.45 ± 25.79 μm) and female (mean ± standard deviation = 113.12 ± 44.37 μm) thyroid glands: (graph represents mean values ± standard deviation). (f) Graph representing the distribution of different sized follicles in male and female rat thyroid glands.
Stereological analysis of the thyroid gland
Stereological analysis of male (n = 10) and female (n = 10) thyroid glands revealed a significant reduction in Vve in females (p < 0.05), while Vvc was about 12% smaller in males but not statistically significant (Figure 2(a), Table 1). Males exhibited a significantly smaller Vvct compared to females (p < 0.01) (Figure 2(a), Table 1). Tissue sections stained with Van Gieson and Azan methods showed distinct connective tissue septa and fine connective tissue surrounding the follicles (Figure 2(b) to (g)). Van Gieson staining colored the connective tissue light red, while colloid and thyrocyte cytoplasm appeared yellow (Figure 2(b), (c), (e), and (f)); Azan stained the connective tissue blue, and thyrocyte cytoplasm red (Figure 2(d) and (g)). The presence of larger areas of connective tissue in female thyroid glands (Figure 2(e) to (g)) compared to males (Figure 2(b) to (d)) was clearly noticeable. Females’ thyroid glands had significantly lower Ia compared to males (p < 0.05) (Figure 2(h), Table 1). Additionally, blood TSH levels were significantly lower in female rats (p < 0.05) (Figure 2(i), Table 1). Pearson correlation analysis indicated a moderate positive correlation between serum TSH levels and Ia in both males (r = 0.7937, p < 0.01) and females (r = 0.7215, p < 0.05) (Table 2).
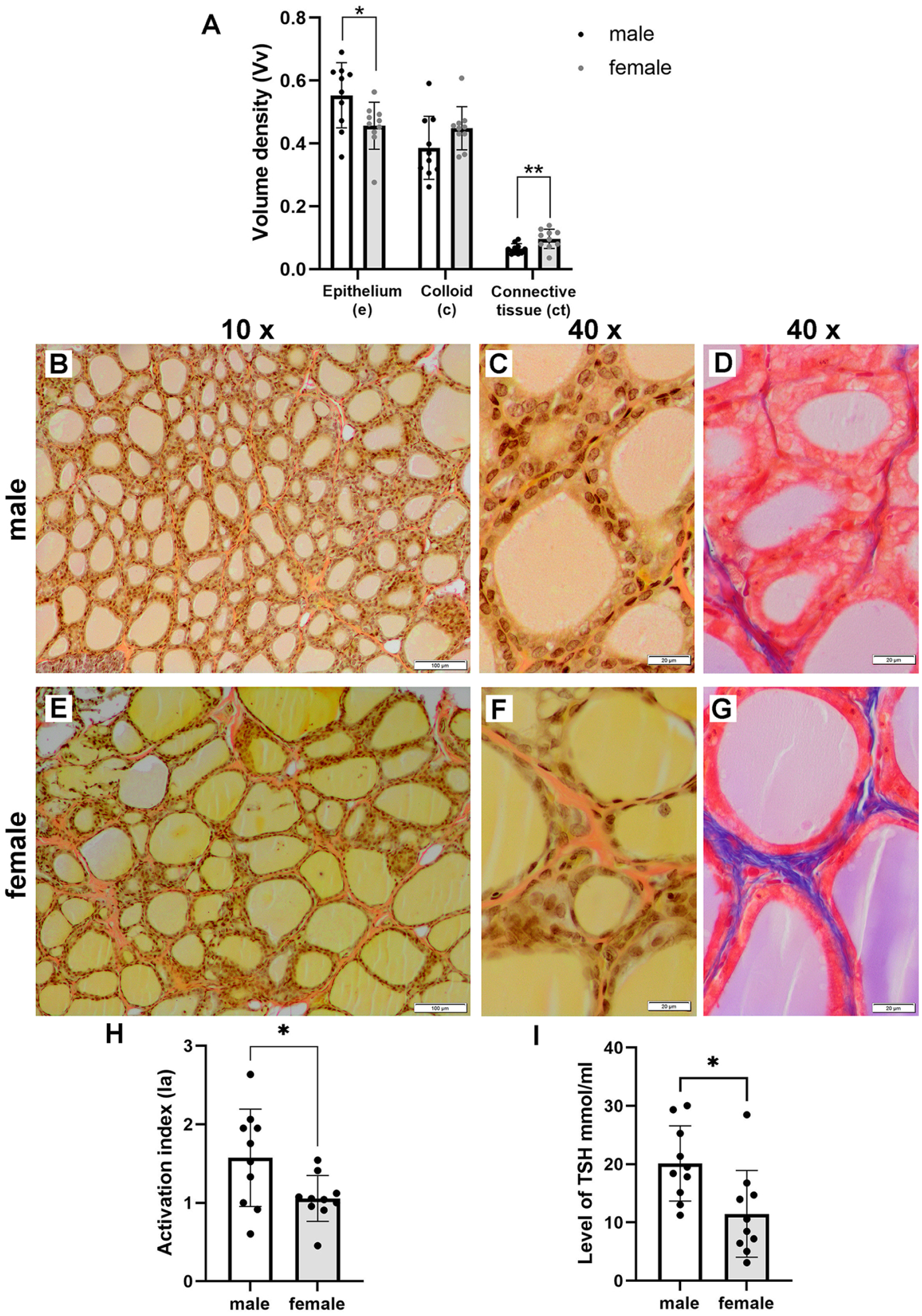
Graph representing (a) mean values ± standard deviation of the volume density of the epithelium (Vve: mean ± standard deviation = 0.55 ± 0.10 in males and 0.45 ± 0.07 in females), colloid (Vvc: mean ± standard deviation = 0.39 ± 0.10 in males and 0.44 ± 0.07 in females), and connective tissue (Vvct: mean ± standard deviation = 0.06 ± 0.09 in males and 0.09 ± 0.03 in females) (*p < 0.05; **p < 0.01). Representative photomicrographs of thyroid glands of (b) to (d) male (n = 10) Wistar rats, and (e) to (g) female (n = 10) Wistar rats. Tissue sections stained using the Van Gieson method viewed in (b) and (e) with a low-power (×10) objective lens (bar: 100 μm), and in (c) and (f) with a high-power (×40) objective lens (bar: 20 μm). Tissue sections stained using the Azan method viewed in (d) and (g) with a high-power (×40) objective lens (bar: 20 μm). (h) Graph representing the mean values ± standard deviation of the activation index (Ia) of thyroid glands (mean ± standard deviation = 1.57 ± 0.62 in males and 1.06 ± 0.29 in females); and (i) TSH levels (mean ± standard deviation = 20.12 ± 6.45 in males and 11.49 ± 7.44 in females).
Results of the stereological analysis and TSH serum level measurements.
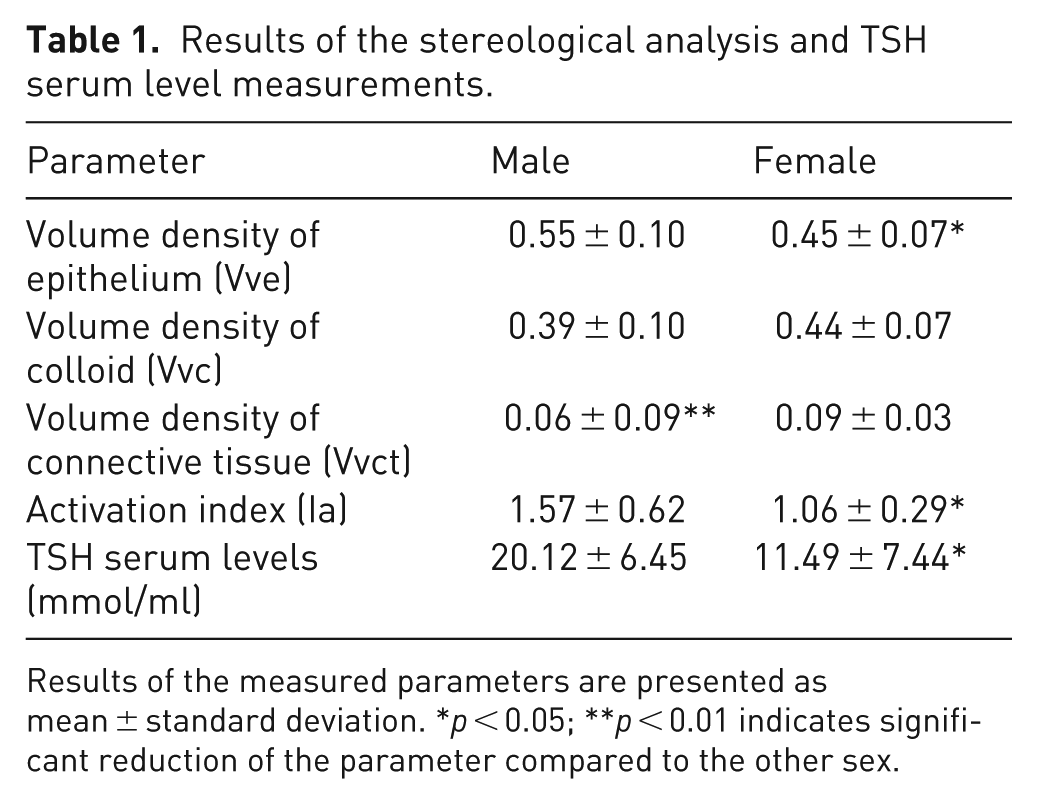
Results of the measured parameters are presented as mean ± standard deviation. *p < 0.05; **p < 0.01 indicates significant reduction of the parameter compared to the other sex.
Pearson correlation analysis between thyroid gland activation index and TSH levels, N = 20.
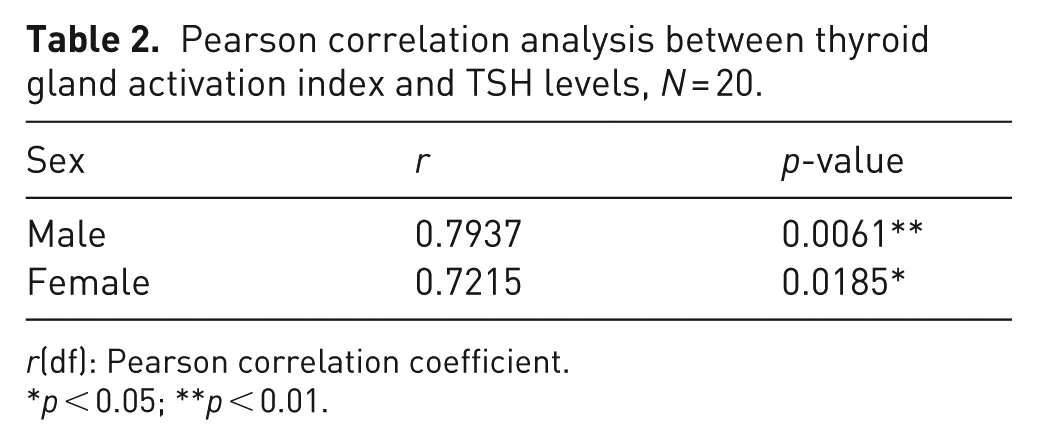
r(df): Pearson correlation coefficient.
p < 0.05; **p < 0.01.
Morphometric analysis of mast cell numbers
In the blue-stained toluidine sections, epithelial cells appeared dark blue, while connective tissue and colloid were stained in lighter shades. Due to the high content of sulfated glycosaminoglycans in the granules occupying the mast cells’ cytoplasm and the metachromatic properties of the toluidine blue dye, mast cells were clearly visible in the connective tissue as red-purple stained cells. The average number of mast cells per 1 mm² in the female thyroid glands (n = 10) was 42.5 ± 9.56 (mean ± standard deviation), while in the male thyroid glands (n = 10) the average number of mast cells per 1 mm² was 41.0 ± 11.49 (mean ± standard deviation). It is important to note that this difference was not statistically significant (p = 0.11) (Figure 3(a)–(e)).
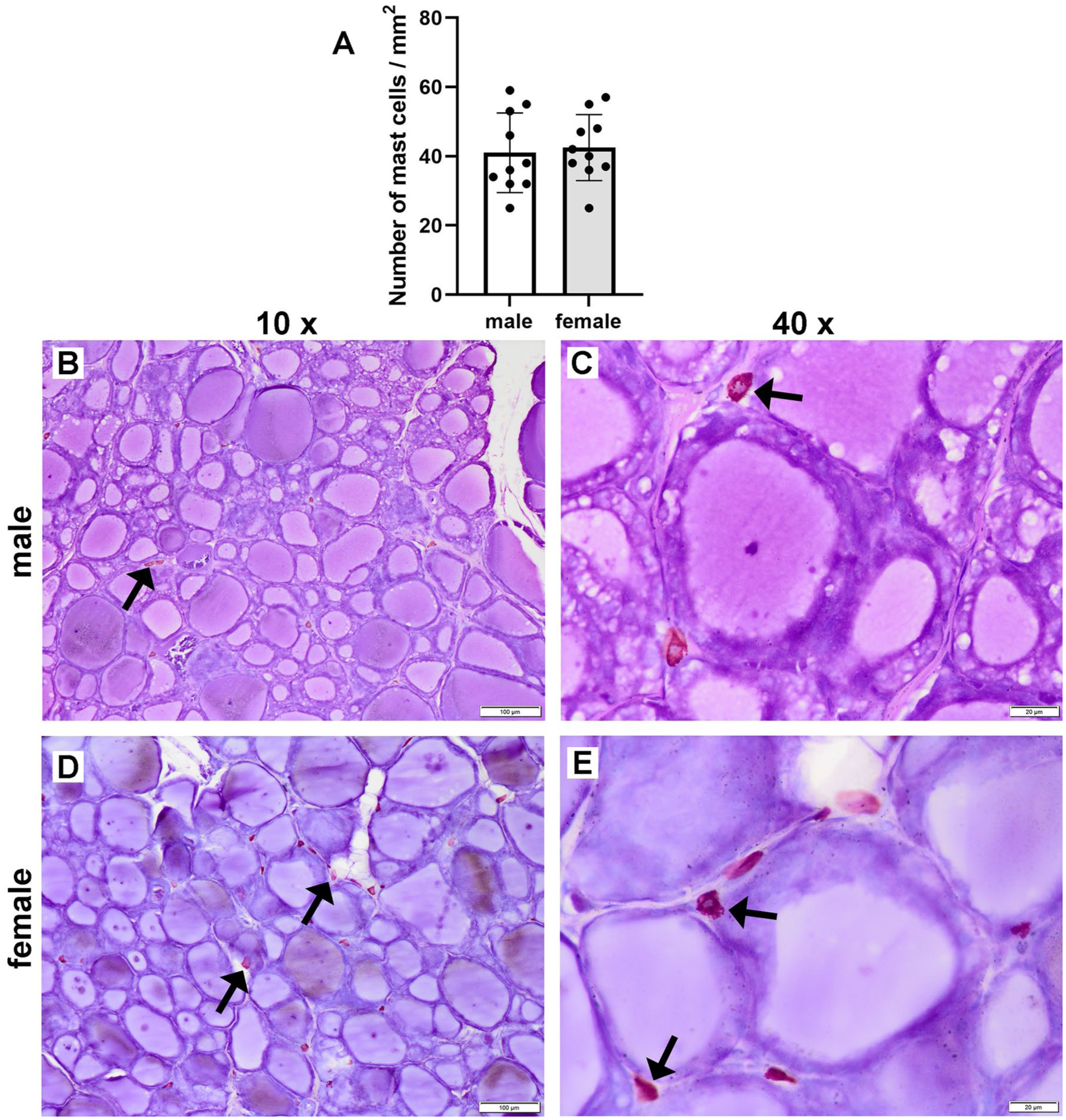
Graph presenting (a) the mean values ± standard deviation of the number of mast cells per 1 mm2 in the thyroid gland tissue of male (mean ± standard deviation = 41 ± 11.49) and female Wistar rats (mean ± standard deviation = 42.5 ± 9.56). Representative photomicrographs of thyroid glands of (b) and (c) male (n = 10), and (d) and (e) female (n = 10) Wistar rats. Tissue sections stained with toluidine blue viewed in (b) and (d) with a low-power (×10) objective lens (bar: 100 μm), and in (c) and (e) with a high-power (×40) objective lens (bar: 20 μm).
Discussion
The significant sexual dimorphism of rat thyroid glands found in our study is corroborated by the research of Mahmood et al. 26 The significantly higher follicular diameter values found in female rats’ thyroid glands, as well as the chi-square test of independence showing a significant relation between female sex and the presence of large follicles in the thyroid glands confirmed Mahmood et al.’s conclusions 26 that the greater relative gland weight in females is due to a predominance of large follicles. In male rat thyroid glands Mahmood et al. 26 observed a larger mean size of follicular cells, which aligns with the significantly higher Vve values in male thyroid glands found in our study. It is interesting to note that studies of hypothyroidism in rats describe the thyroid glands of affected animals having smaller thyroid follicles lined with epithelium made of higher-columnar thyrocytes.8–10 In this regard, our histological findings of small follicles being predominant in male rat thyroid glands, as well as Vve being significantly higher in males suggest that the thyroid glands in males are less active than those in females.
Stereological analysis revealed differences in the volume densities of other structural components besides the aforementioned epithelium (Vve). Female thyroid glands had significantly higher Vvct; the values of Vvc were also higher in females but this increase was not significant. The absence of significance in the comparison of this parameter could be explained by the results of the chi-square test of independence, which demonstrated the existing relation between sex and the presence of differently sized follicles in male and female rats’ thyroid glands. It is possible that both groups have a similar quantity of colloid in the thyroid glands, but that it is redistributed among follicles of different sizes. Interestingly, stereological studies of thyroid glands in other species demonstrate similar sexual differences as those observed in the rat thyroid glands here. Research of thyroid glands from male and female rabbits revealed that males had significantly higher Vve, while Vvct was increased in females. Although a larger Vvc was observed in females, the difference was not significant. 27 Parchami and Dehkordi conducted a stereological analysis of duck thyroid glands, finding higher Vve and Vvc values in males and an increased Vvct in females. 28 Although the differences were not significant, the study concluded that the morphological properties of duck thyroid glands varied between the sexes. The similarities of sexual differences observed in the thyroid structure of animals belonging to different orders, and even different classes, could be explained by the fact that the thyroid gland is one of the phylogenetically oldest endocrine glands of vertebrate species. 29 Although there are some differences in the gross morphology between species, 30 the thyroid gland is structurally conserved in all vertebrate species, and shows a similar structure of follicles as well as function. 29 It is possible that conservation of this structure led to the conservation of sexual differences in vertebrate species.
TSH is a pituitary hormone that induces morphological and functional changes in the thyroid, depending on its activity. 31 Research comparing cell types in the pars distalis of male and female rat pituitary glands indicates a greater volume density of TSH cells in males. 32 A broader study involving 96 female and 97 male rats showed that the mean TSH value in the plasma of male rats was significantly higher than in the females. 33 Our finding of male rats having significantly higher serum levels of TSH is aligned with the findings of previously mentioned studies. It is important to note that determining TSH levels is considered the best test of thyroid function and, oftentimes, if TSH is within normal range, the use of additional tests is not indicated. 34 However, there are some problems with this practice. Firstly, studies suggest that hormone reference intervals used to assess thyroid function do not take into account several factors that may cause variation in hormone levels: age, sex, stress, immune status, and so forth. 35 Secondly, there are conditions characterized by mildly elevated or decreased levels of TSH that remain within the reference intervals, although the thyroid function is impaired: subclinical hyperthyroidism and hypothyroidism, respectively. 34 For this reason, we wanted to explore the relationship between TSH levels and Ia, considering that Ia is a parameter that assesses thyroid gland function from a histological standpoint. Previous rat studies showed that serum TSH levels are directly proportional to Ia.25,36 However, there is a lack of studies that show a direct correlation between these two variables. Our study found that Ia was higher in males than females, aligning with elevated TSH levels in male serum. By calculating the Pearson correlation coefficient, we also demonstrated that a positive correlation between the TSH levels and Ia existed in both male and female rats. As a purely histological parameter, Ia is less prone to the fluctuations that affect biochemical parameters like hormone levels, and the correlation established in our study demonstrated that Ia could be used as reliable complementary parameter for assessing thyroid function. The use of Ia in thyroid gland research is important because it can serve as a parameter that can confirm whether the morphology of the gland aligns with the biochemical data, and would be particularly useful in assessing thyroid function in cases where normal TSH levels do not necessarily reflect euthyroidism (i.e., presence of heterophile antibodies, mutation of the thyroid hormone receptor gene, TSH-secreting pituitary adenoma). 34
Mast cells are known to play a role in inflammatory and immune responses by storing histamine in cytoplasmic granules. Some studies indicate that mast cells can also synthesize and store TSH and T3.17–19 Bianco et al. found that TSH’s effect on the rat thyroid was nearly completely inhibited by cromoglycate, a mast cell stabilizer, whereas treatment with BW48-80, which destabilizes mast cells, increased T3 and T4 levels. 17 These findings positioned mast cells as mediators of the thyroid gland’s response to TSH. Other studies suggest that mast cells influence thyroid function in conditions when it is impaired, like Hashimoto’s syndrome and Graves’ disease, which mainly affect females. 19 Additionally, Chieffi Baccari et al. reported an increased number of mast cells in hypothyroid rats’ tissues. 18 In our study, we observed no significant difference in the number of mast cells in thyroid tissue between male and female rats, though females had higher mean values, which contradicted the findings of Chieffi Baccari et al. 18 Further research with a larger sample size, or use of molecular methods to determine the activation status of mast cells present in the thyroid gland tissue of male and female rats, would help elucidate the effects of mast cells on thyroid function.
Conclusion
Our histological analysis revealed clear sex-specific differences in the structure of the rat thyroid gland, underscoring the importance of including sex as a biological variable in experimental design. Due to increasing concerns that the hormone reference intervals used to assess thyroid function do not take in account all factors that cause variation in hormone levels, and the existence of thyroid gland disfunctions that are not adequately represented by TSH levels, only measuring TSH may hinder the quality of thyroid function assessments in preclinical research settings. The strong correlation observed between the thyroid Ia and serum TSH levels in our study suggests that Ia may serve as a practical complementary parameter to ensure a more precise and complete estimation of thyroid function.
Footnotes
Author contributions
The authors confirm contribution to the paper as follows: study conception and design: IM, TLB; funding acquisition: ABĐ; project administration: TLB; data collection: IM, AN, EN, BBP, JJ, ĐM; analysis and interpretation of results: AN, ĐM; draft manuscript preparation: IM, JJ, EN; reviewing and editing the final version of the manuscript: AR, BBP, ABĐ, TLB.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Ministry of Education, Science, and Technological Development of the Republic of Serbia (Contract No. 451-03-66/2024-03/200143) and by the Fund for Science of the Republic of Serbia (Promis program, DecodExpo, No. 6066532).
Ethical considerations
The protocol for animal experiments was approved by the Ethics Committee for Experimental Animal Research of the Faculty of Pharmacy, University of Belgrade, and the Ministry of Agriculture, Forestry, and Water Management–Veterinary Directorate (ethical permit number: 323-07-11822/2018-05), in compliance with the National Institutes of Health guidelines for the care and use of laboratory animals.
ORCID iDs
Data availability statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.