
Abstract
Since increased cerebral oxygenation reflects cerebral activation, this study investigated the effect of mastication frequency on prefrontal cortex oxygenation. Eleven young volunteers (nine women, two men; M age = 20.9 years, SD = 0.9) carried out three trials in which they were asked to chew a tasteless gum for 3 min at varying (rates of mastication frequency: 30, 70, and 110). Breaks of 2 min each were interleaved between trials. The oxygenation of the left prefrontal cortex was monitored by near-infrared spectroscopy. We found a significant increase in cortical oxygenation during gum chewing in all three conditions (p < .05), compared with a resting level; we also found a significant difference between the Fast and Slow chewing conditions, and between the Fast and Normal (70 rpm) conditions, both findings seemingly related to activation of a motor command in frontal brain regions. To our knowledge, this is the first report on the effect of mastication frequency on cerebral oxygenation. Possible implications of this finding are discussed.
Keywords
Introduction
Cognitive impairment and dementia resulting from age-related decline are substantial public concerns, because they are common in older people and major causes of morbidity and mortality (Daviglus et al., 2010; Dewey & Saz, 2001). Miura, Yamasaki, Kariyasu, Miura, and Sumi (2003) showed that patients over 65 years with dementia have fewer teeth and smaller occlusal area than similar aged persons without dementia. A possible implication of this association is that masticatory and cognitive functioning in elderly people may be related, perhaps from an association between cerebral hypo-oxygenation, less frequent chewing, and diminished cognitive functioning (Poels et al., 2008).
Mastication consists of the activation of jaw and masticatory muscles concerned with rhythmic and voluntary movement. The motor commands for sequential rhythmic movement are generated by a neural population in the central pattern governor of the brainstem (Nakamura & Katakura, 1995; Nakamura, Katakura, Nakajima, & Liu, 2004). Recent neuroimaging studies suggest that mastication is a complicated movement generated from a neural network in several regions of the brain including the primary motor cortex and prefrontal cortex (PFC; Hasegawa et al., 2013; Momose et al., 1997; Onozuka et al., 2003; Takada & Miyamoto, 2004; Takahashi, Miyamto, Terao, & Yokoyama, 2007; Tamura, Kanayama, Yoshida, & Kawasaki, 2003). However, it remains unclear how such systems are involved in eating behavior.
Previous studies have reported several effects of mastication and gum chewing on functions such as arousal (Otomaru et al., 2003), energy expenditure (Suzuki, Shibata, & Sato, 1992), choice reaction time (Chu, 1994), and working memory (Moss & Scholey, 1996; Wilkinson, Scholey, & Wesnes, 2002). Using event-related potentials, Sakamoto, Nakata, and Kakigi (2009) discovered that gum chewing may improve central nervous system function.
Since mastication is a complicated movement, brain oxygenation is expected to increase with increased mastication frequency, just like it does in other rhythmic movements. Because high-frequency movements require minimal learning, and because the subjects have to attend to their jaw and to the auditory cues to perform the gum-chewing task, it is expected that higher frequency gum chewing associates with greater brain oxygenation.
The near-infrared spectroscopy (NIRS) technique has been found to be useful for investigating cortical oxygenation during movement generation (Derosière et al., 2014; Shibuya & Kuboyama, 2007; Shibuya, Sadamoto, Sato, Moriyama, & Iwadate, 2008). NIRS allows the measurement of changes in concentrations of oxy- (OxyHb) and deoxyhemoglobin (DeoxyHb) using different wavelengths in the near-infrared range. NIRS signals indicate when the blood flow increase related to cortical oxygenation is larger than the increase in oxygen uptake. Thus, cerebral oxygenation increases in a temporally and spatially coordinated manner, tightly linked to changes in neural oxygenation and cerebral blood flow through a complex sequence of coordinated neuronal events (Girouard & Iadecola, 2006). NIRS also infers changes in neural activation that is mirrored by changes in blood oxygenation in the region of the activated cortical area (i.e., increase in OxyHb or decrease in DeoxyHb). This is similar to the blood oxygenation level-dependent contrast in functional magnetic resonance imaging (Villringer & Chance, 1997). The oxygenation response measured by NIRS represents an increase in OxyHb and a slight decrease in DeoxyHb over the activated region (Obrig & Villringer, 2003).
Although the previous studies suggested that the gum chewing increases cerebral oxygenation, it remains unclear how the frequency of gum chewing influences PFC oxygenation. We therefore hypothesized that an increase in gum chewing frequency will encourage an increase of PFC oxygenation.
Methods
Participants
Eleven young, health subjects (nine women and two men; age: 20.0 ± 0.9 years; mean ± SD) were enrolled in this study. The study protocol was approved by the Ethics Committee of Niigata University of Health and Welfare (No. 17619). Each subject provided a written informed consent after the experimenter’s explanation of the study’s aim and methodology.
Procedure
The gum used had previously been tested to ensure no detectable taste and odor (Meiji Co., Ltd., Tokyo, Japan). It consisted of 94.5% Gum Base, 1.5% Lecithin, and 4% Aspartame/Acesulfam K. Subjects were instructed to sit on a chair while wearing an eye mask. They were required to sequentially chew three pieces of gum for 3 min in each trial (Figure 1). The three trials differed in their chewing frequency. The chewing rhythm was regulated by a metronome at the rate of 70 repetitions per minute (rpm; Normal), 110 rpm (Fast), and 30 rpm (Slow). Chewing side was not restricted. Chewing at 70 rpm requires the least conscious effort (Hasegawa et al., 2013). Chewing frequency order was randomized for each subject. Subjects inserted a piece of gum (0.9 g) into their oral cavity immediately before the trial. The gum was removed after each test. Breaks between tests lasted for 2 min.
Protocol for the gum chewing test. Subjects underwent three trials, where they were asked to chew an odorless and tasteless gum for 3 min, with an interval of 2 min between each trial.
Measures
Subjective evaluation of gum sweetness
Because the taste sensation is processed through the following pathway, taste bud; solitary nucleus; parabrachial nucleus; thalamus; and gustatory cortex (Gilbertson, Damak, & Margolskee, 2000), we measured the subjective evaluation of gum sweetness. After completing the three trials, each subject was required to rate the sweetness for each gum using a Visual Analogue Scale (VAS). Scores ranged between 0 (not sweet at all) and 15 (extremely sweet).
Cerebral oxygenation
NIRS (NIRO-200, Hamamatsu Photonics, Hamamatsu, Japan) was used to monitor the changes in hemoglobin concentration in the left frontal lobe, because the left side frontal lobe mainly expresses cerebral oxygenation in right hand subjects. The NIRS optodes consisted of a semiconductor laser light source and a light detector, 4 cm apart from each other. This device can measure the tissue absorbance of light at three wavelengths (780, 805, and 830 nm) to detect concentration changes in OxyHb (ΔOxyHb) and DeoxyHb (ΔDeoxyHb) (Elwell et al., 1994). Relative concentration changes were measured from the resting baseline of OxyHb and DeoxyHb respectively. DeoxyHb is known to be a reliable estimator of changes in tissue oxygenation status, while OxyHb seems to be a more sensitive indicator of regional cerebral blood flow modifications (Hoshi, Kobayashi, & Tamura, 2001). Then, to evaluate cerebral blood volume changes during chewing, we calculated TotalHb (=OxyHb + DeoxyHb). To avoid inclusion of the effects of preparation and attention to the mastication on oxygenation changes in the analyses, we determined the baseline values of OxyHb, DeoxyHb, and TotalHb as the mean value over 30 s before the onset of the first trial. Since the baselines were stable, ΔOxyHb, ΔDeoxyHb, and ΔTotalHb are presented as the magnitude of change from their respective baseline. The optodes were attached to the skin over the frontal skull 2 cm from the midline and 1.5 cm over the eyebrow. The sampling rate of the NIRS was 2 Hz.
Data analysis
All data are expressed as mean ± one standard deviation (SD). The VAS data for the subjective values of sweetness were compared between the three frequencies by a repeated measures one-way analysis of variance (ANOVA). To test the time course of ΔOxyHb and ΔDeoxyHb, we calculated the average of the individual data for each of the 3 min and made statistical comparisons with a repeated measured one-way ANOVA with Dunnett’s Post hoc test. To evaluate ΔOxyHb, ΔDeoxyHb, and ΔTotalHb changes during gum chewing, peak values of ΔOxyHb, ΔDeoxyHb, and ΔTotalHb were compared with the reference values (values at the average during baseline period before the trials) using a repeated measures one-way ANOVA with Bonferroni multiple comparison post hoc test. For all analyses, p < .05 was considered statistically significant.
Results
Subjective estimates of the gum sweetness are shown in Figure 2. Sweetness ratings did not differ significantly between frequencies, F(2,10) = 1.706, p = .207, η2 = 0.86.
Visual Analogue Scales of subjective ratings of sweetness, for each of the three conditions of mastication frequency (Slow, 30 rpm; Normal, 70 rpm; Fast, 110 rpm). Values are mean ± SD. There was no significant difference among three conditions.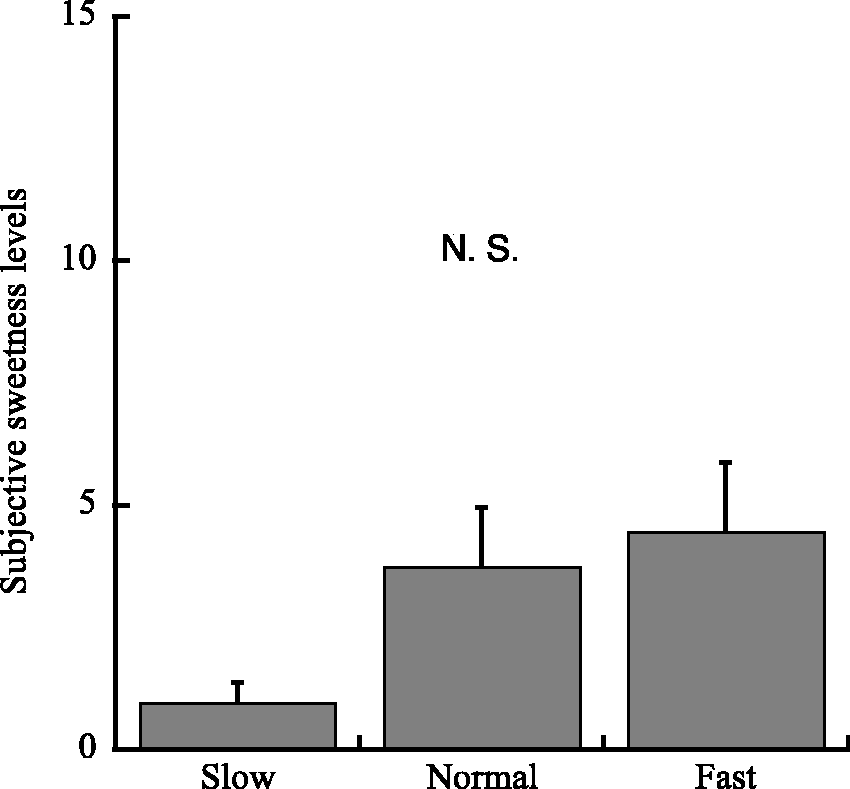
Peak Value Changes in Oxygenation
Figure 3 shows the summaries of oxygenation change in the left PFC for each chewing frequency.
Oxygenation in the prefrontal cortex. Changes from the baseline values in (1) oxyhemoglobin concentration (ΔOxyHb) and (2) deoxyhemoglobin (ΔDeoxyHb) in the prefrontal cortex during gum chewing. Values are mean ± SD. Letters a and b indicate significant differences among three conditions, p < .05.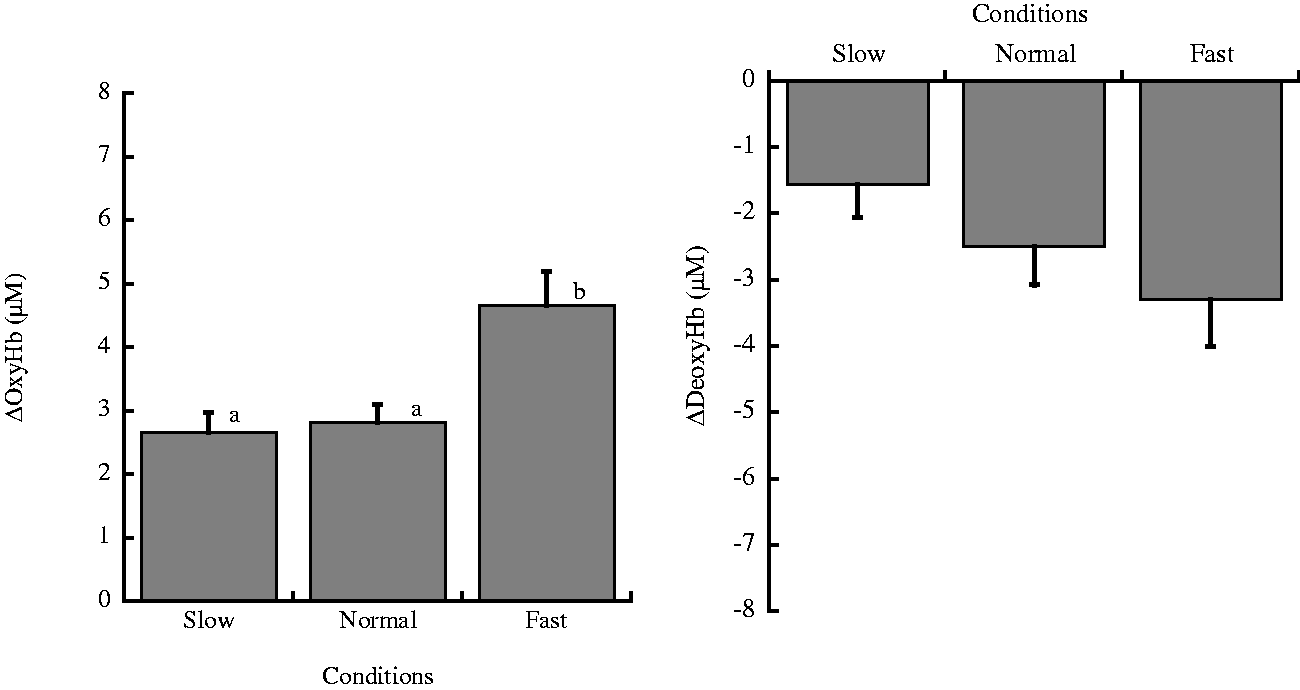
The peak value of ΔOxyHb values from the baseline levels was different between frequencies, F(2,20) = 13.87, p = .0017, η2 = 0.29. Compared with the Slow chewing condition, ΔOxyHb change was higher in all subjects and significantly higher in the Fast chewing condition, t(10) = 4.80, p = .0003, r = 0.84. ΔOxyHb change in the Fast condition was higher in all subjects and significantly higher than in the Normal condition, t(10) = 4.27, p = .0011, r = 0.80. There was no significant difference between the Slow and Normal conditions, t(10) = 0.53, p = 1, r = 0.17).
On the other hand, ΔDeoxyHb showed small but significant decreases. Peak changes in ΔDeoxyHb values from values at resting levels were different between the three different frequencies, F(2,10) = 4.14, p = .03, η2 = 0.11. Compared with the Slow condition, ΔDeoxyHb was lower in all subjects and significantly lower in the Fast condition, t(10) = 2.88, p = .0279, r = 0.67. ΔDeoxyHb change in the Fast condition was not significantly higher than in the Normal condition, t(10) = 1.41, p = .5194, r = 0.41. There was no significant difference between the Slow and Normal conditions, t(10) = 1.47, p = .4752, r = 0.42.
ΔTotalHb changes during three conditions (Slow, Normal, Fast conditions).

In addition to chewing speed, the number of chewing strokes was different among three conditions (Slow, Normal, and Fast conditions), and it remains possible that this factor mostly affected changes in peak oxygenation among three conditions. Oxygenation differences merely reflect differences in temporal summation of NIRS signals that correspond to each single chewing stroke. To evaluate the cerebral activations that depend on chewing strokes, we calculated oxygenation signals per chewing frequency.
Peak changes per frequency of chewing from the baseline values in ΔOxyHb, ΔDeoxyHb, and ΔTotalHb.
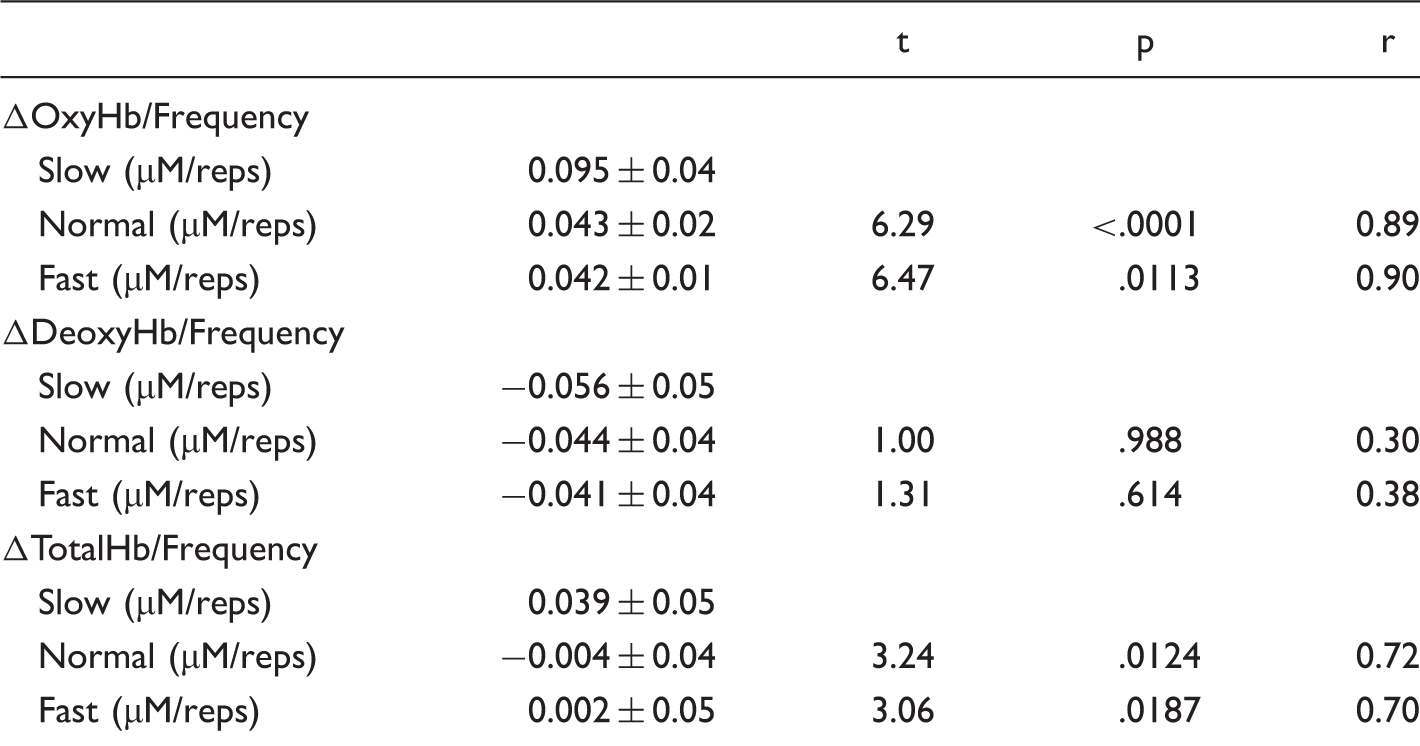
Note. t, p, and r values are given for the differences compared with the Slow condition.
On the other hand, peak changes in ΔDeoxyHb or frequency values from values at resting levels were not different between the three different frequencies, F(2,10) = 0.94, p = .4079, η2 = 0.02. ΔTotalHb or frequency values from values at resting levels were different between the three different frequencies, F(2,10) = 6.62, p = .006, η2 = 0.14. Compared with the Slow condition, ΔTotalHb or frequency change was significantly lower in the Fast condition, t(10) = 3.06, p = .0187, r = 0.70. ΔOxyHb or frequency change in the Normal condition was lower in all subjects and significantly higher than in the Slow condition, t(10) = 3.24, p = .0124, r = 0.72. There was no significant difference between the Normal and Fast conditions, t(10) = 0.18, p = 1, r = 0.06.
Time Course Changes in Oxygenation
Changes from the baseline values in ΔOxyHb and ΔDeoxyHb.
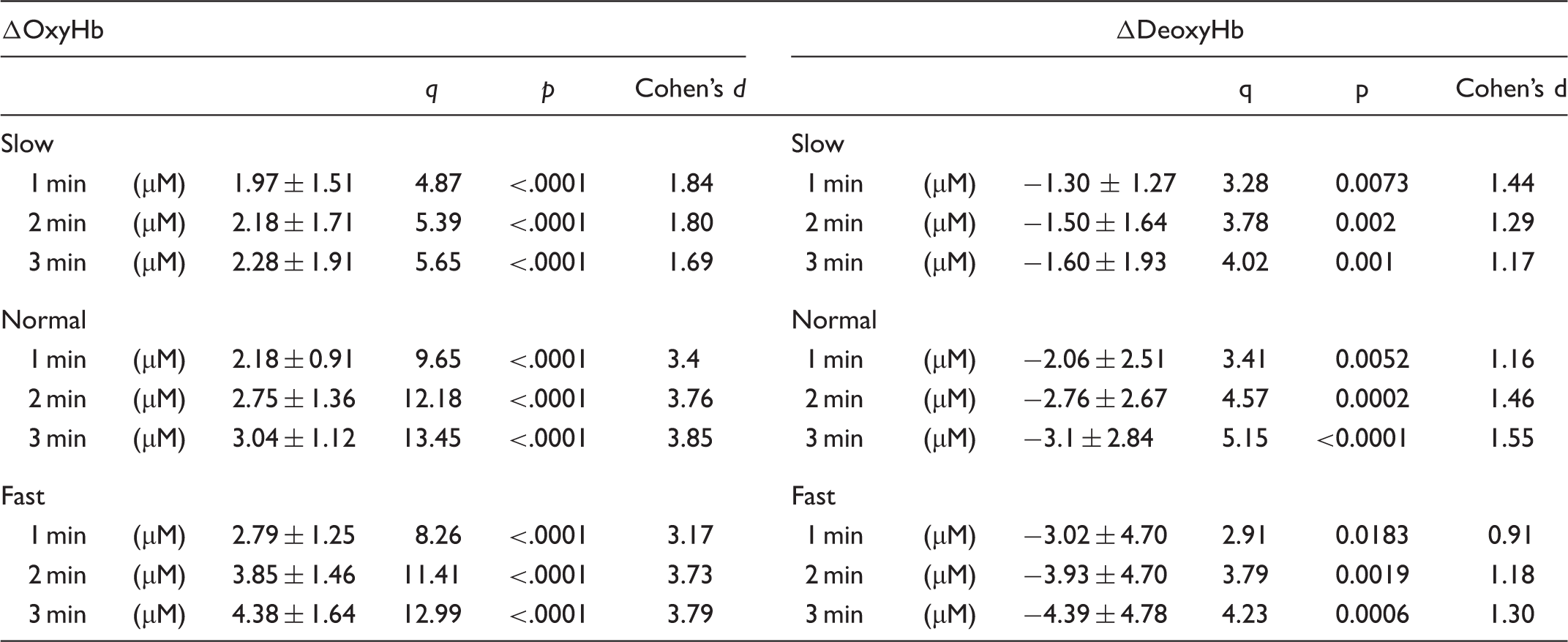
On the other hand, ΔDeoxyHb showed small but significant decreases. ΔDeoxyHb decreased significantly during gum chewing (F(3,10) = 7.01, p = .00104, η2 = 0.19 in Slow; F(3,10) = 10.62, p < .0001, η2 = 0.23 in Normal; F(3,10) = 7.21, p = .00086, η2 = 0.16 in Fast). ΔDeoxyHb significantly decreased 1 min after the onset of gum chewing in all three conditions (p < .01), and sustained oxygenation of ΔDeoxyHb was observed until the end of chewing (p < .001 respectively; Table 3).
Discussion
In the present study, we examined whether the frequency of gum chewing can change cerebral oxygenation. The main finding of the present study is that slow gum chewing (30 rpm) did not decrease the oxygenation in the PFC compared with normal-frequency gum chewing (70 rpm). However, fast gum chewing (110 rpm) significantly increased PFC oxygenation compared with normal-frequency. To our knowledge, this is the first report on the effect of mastication frequency on cerebral oxygenation. We measured ΔOxyHb and ΔDeoxyHb and found statistical effect sizes that were larger in ΔOxyHb than in ΔDeoxyHb. Effect sizes were larger in ΔOxyHb than in ΔDeoxyHb.
Generally, possible explanations for elevated PFC oxygenation during gum chewing include the effect of mastication with an object in the mouth, the effect of taste and hardness perceptions, and the effect of motor-related activities elicited by mastication. In the present study, we used the same gum composition for each of the three conditions. The VAS analysis indicated that the subjects experienced a similar sweetness level in the three conditions. Thus, the cognitive and motivational contexts were identical over these experimental conditions. The taste sensation is processed through the following pathway: taste bud, solitary nucleus, parabrachial nucleus, thalamus, and gustatory cortex (Gilbertson et al., 2000). A previous study (Hasegawa et al., 2013) suggested that PFC oxygenation also increases with taste perception, but, in the present study, there was no difference in taste between the three conditions. Nevertheless, there were significant differences in the oxygenation level of the PFC between the Slow and Fast, and between the Normal and Fast conditions, while there was no significant difference between the Slow and Normal conditions. The differences in oxygenation level between the Fast and other conditions are unlikely to be caused by the sensation of taste and hardness, as the same pieces of gum were used between conditions. The effect size in VAS analysis was high value, however, the previous studies (Gilbertson et al., 2000; Hasegawa et al., 2013) did not report the effect size. Therefore, we could not compare the results between the previous results and our results.
The increase of PFC oxygenation in the Fast condition compared with the other two conditions might be an effect of motor-related activities elicited by mastication. As mentioned earlier, Jenkins, Brooks, Nixon, Frackowiak, and Passingham (1994) reported midbrain activation during the learning of a new finger tapping sequence, whereas no significant activation was detected during performance of a prelearned sequence. Gum chewing consists of the oxygenation of jaw and masticatory muscles concerned with rhythmic and voluntary movement. In that sense, gum chewing belongs in the same category as finger tapping. The PFC oxygenation represented the activation of cerebellorubral pathways during the rhythmic task, although the effect of increased attention in learning a new sequence was another possibility. The frequency of gum chewing might affect brain oxygenation because high-frequency gum chewing has a minimal learning component and because the subjects have to attend to their jaw and auditory cues to perform the gum chewing task. On the other hand, in the Slow and Normal conditions, the movements for gum-chewing would have a maximal learning component because the subjects does not have to attend to their jaw and to auditory cues to perform the rhythmical gum chewing.
A previous study reported that jaw movements for gum chewing did not have a main effect on brain functions as measured by reaction time and the P300 event-related potential (Sakamoto et al., 2009). However, oxygenation effects in the present study raise new questions about possible changes in cognition from jaw movements for high-frequency gum chewing. Further studies are needed to be clear about any relationship between increased cerebral oxygenation during mastication and changed cognitive functioning, especially as might relate to elderly, rather than young participants. In addition, further research is needed to show the precise relationship between brain oxygenation and high-frequency mastication.
A limitation of the present study was that we did not measure the cerebral blood flow using laser-Doppler flowmeter to determine cutaneous blood flow responses. If we had measured the cerebral blood flow using laser-Doppler flowmeter, we could focus on not only the cerebral oxygenation but also on cerebral activation. We calculated TotalHb to evaluate the cerebral blood volume changes during chewing, however, there were no significant changes in TotalHb during chewing. This would imply that there were no effects of cerebral blood flow changes on cerebral oxygenation signals. In addition, the number of chewing strokes was different among the three chewing conditions (Slow, Normal, and Fast conditions), possibly accounting for changes in the peak oxygenation among three conditions since oxygenation differences merely reflect the difference in temporal summation of NIRS signals that correspond to each single chewing stroke. To evaluate the cerebral activations depending on chewing strokes, we calculated the oxygenation signals per chewing frequency. The results of the calculation of the oxygenation signals per chewing frequency indicates that the difference in the cerebral oxygenation among three conditions might be due to the different number of chewing strokes. There is also a debate about whether a dental prosthesis could reorganize masticatory-induced cortical activity pattern including the prefrontal area (Yan, Ye, Zhen, Ke, & Gand, 2008) since the chewing-induced PFC activity may improve cognitive function (Ono, Yamamoto, Kubo, & Onozuka, 2010). In this study, we found more activity in higher chewing frequency condition. If any dental prosthesis improves the masticatory-functions, dental prosthesis would reorganize masticatory-induced cortical activity pattern. Further research would be needed to clarify these relationships concerning gum chewing.
In conclusion, we examined the PFC oxygenation response during several frequencies of gum chewing. We found a significant increase of PFC oxygenation during faster mastication condition, compared with normal and slower mastication conditions. These findings suggest that the frequency of gum chewing affects brain oxygenation.
Footnotes
Authors Contributions
T. Y. carried out the experiments and participated in the sequence alignment and drafted the manuscript. M. S. and S. N. carried out the experiments. N. K., K. Suzuki, and H. I. participated in the design of the study and the sequence alignment. K. Shibuya participated in the sequence arrangement and drafted the manuscript.