
Abstract
Pitch discrimination accuracy has been found to be affected by many factors, including handedness, musical training, interfering stimuli, and spatial location of the auditory stimulus. Separating the stimulus input location of interference tones from initial (reference) and final (comparison) tones leads to more accurate pitch discrimination, but the effects of spatial location relationships between the reference, interference, and comparison tones have not been fully explored. This study examined the impact of stimulus spatial location in 24 young, nonmusician females. Participants determined whether the pitch of reference and comparison tones were the same or different in 20 pitch discrimination conditions with varied interference, spatial relationships, and frequencies. Findings revealed that pitch discrimination accuracy was significantly better when (a) there was no interference, (b) the comparison tone was presented to the contralateral brain hemisphere from reference and interference tones, and (c) the comparison tone was presented to the left ear. We discussed the implications of these findings for therapy programs to strengthen pitch discrimination abilities.
Introduction
Pitch processing is essential to understanding nearly every form of auditory communication. Regardless of whether the auditory signal is instrumental music, vocal music, or speech, pitch perception is a key element for message comprehension. Short-term memory is also vital for both language and musical processing (Cowan, 1999). For an individual to discriminate any difference between two pitches, the initial pitch must be held, at least momentarily, in short-term memory. Pitch memory and processing are important to language processing to understand meaning conveyed by intonation contours; they are also fundamental to music perception, as they are vital to defining melody and conveying musical emotion.
Pitch discrimination ability (PDA) is commonly assessed by asking an individual to listen to both an initial reference and a final comparison tone and make a judgment as to whether the two tones are the same or different. PDA can be affected by many factors, including the presence of any interference tones between the initial reference and final comparison tones (Deutsch, 1970a). Interfering stimuli cause a significant increase in error rate, which is thought to be due to the interference in pitch memory between a reference and comparison tone (Deutsch, 1970a). Tonal stimuli are associated with higher error rates in pitch discrimination than speech stimuli (Deutsch, 1970b), and the overall pitch of interfering stimuli has a significant effect on PDA, with research showing that interfering stimuli that are closer in pitch to the reference and comparison stimuli have a more detrimental effect on pitch discrimination accuracy (Gockel, Hafter, & Moore, 2009; Semal, Demany, Ueda, & Halle, 1996). These findings led to the theory that there is a specialized perceptual system for processing pitch, and this theory has been substantiated by work in the field of amusia, involving patients who can process language but cannot discriminate fine differences in pitch (Ayotte, Peretz, & Hyde, 2002; Peretz & Hyde, 2003).
Behavioral and neurophysiologic research has shown that PDA is also affected by both stimulus-related factors, such as time delays between reference and comparison tones (Estis, Coblentz, & Moore, 2009; Moore, Keaton, & Watts, 2007), and participant-related factors, such as musical training, gender, age, and intelligence (Smith, Bartholomew, Burnham, Tillmann, & Cirulli, 2017). While there is neurophysiologic evidence that pitch is processed in a different neuroanatomical location from language (Imm et al., 2008; Zatorre, Evans, Meyer, & Gjedde, 1992), additional studies suggest a potential for some degree of shared resources and neural overlap in auditory short-term memory of speech and musical processing (Schulze, Zysset, Mueller, Friederici, & Koelsch, 2011; Williamson, Baddeley, & Hitch, 2010).
While these neurophysiologic findings inform our understanding of pitch processing, questions remain regarding the underlying mechanics of how these physiologic processes affect behavioral performance. While prior behavioral studies have documented the differential effects of ipsilateral and contralateral presentations of interference (Deutsch, 1978a; Gockel et al., 2009), we know of no study of the effects of a randomized presentation of spatial location reference, interference, and comparison tones and their interrelationships. This is an important distinction, as environmental stimuli entering the auditory system are not generally presented in a predictable fashion. Therefore, the present study was undertaken to examine the effects of randomized spatial location of input on PDA.
Historically, pitch processing has been thought to occur primarily in the right cortical hemisphere (Bianchi et al., 2017; Coffey, Herholz, Chepesiuk, Baillet, & Zatorre, 2016; Gelfand, 2004; Hyde, Peretz, & Zatorre, 2008; Zatorre, 1988), although some studies have found evidence to dispute this (Allen, Burton, Olman, & Oxenham, 2017; Penagos, Melcher, & Oxenham, 2004). Because of this presumed right hemisphere processing, tonal stimuli presented to the left ear are thought to be processed more efficiently than tones presented to the right ear. Accordingly, we hypothesized that reference and comparison tone stimuli presented to the left ear or binaurally would yield a higher number correct score than stimuli presented to the right ear only. Further, we predicted that same-sided presentations of reference and comparison tones would yield a higher number correct score than different-sided presentations of these tones because fewer cognitive resources are involved when the central auditory system perceives comparative tones to be coming spatially from the same sides. A binaural presentation of reference and comparison tones was predicted to yield higher number correct scores than contralateral presentation because the spatial location of these presentations permits more efficient same-sided brain processing for both tones. Our expectations based on previous research findings (Deutsch, 1978a; Kallman, Cameron, Beckstead, & Joyce, 1987; Micheyl, Delhommeau, Perrot, & Oxenham, 2006) were that monaural, left ear presentations of both reference and comparison tones with monaural right-ear presentations of interference tones would yield the most accurate scores, suggesting that pitch memory works most efficiently when there is a spatial separation of interfering tones from initial and final comparison tones.
Method
Participants
We recruited 24 normal-hearing female participants, aged between 22 years, 4 months and 29 years, 6 months (M = 24.98, SD = 1.74) from among students at the University of South Alabama and the surrounding community. Only female participants were recruited because prior research using similar pitch discrimination methodology in nonmusicians showed no significant gender differences in performance (Estis, Dean-Claytor, Moore, & Rowell, 2011). All participants signed written informed consent forms.
Our sample size was comparable to that of prior work examining pitch discrimination abilities (Deutsch, 1978a; Moore et al., 2007; Pechmann & Mohr, 1992; Semal et al., 1996), and a posteriori power analysis (Statistical Package for Social Sciences) revealed the sample size to have sufficient power for planned statistical analyses. Due to previous findings documenting that left-handed individuals have a performance advantage on pitch memory tasks (Deutsch 1978b), we used only right-handed participants in this study, as determined by participant performances on the Annett Handedness Questionnaire (Annett, 2004). In addition, because musical training has been found to enhance performance on pitch processing tasks (Estis et al., 2009, 2011; Hutka, Bidelman, & Moreno, 2015; Parbery-Clark, Skoe, Lam, & Kraus, 2009; Weaver, DiGiovanni, & Ries, 2017), we used participants who reported no significant musical training, defined as three or more years of individual vocal or instrumental training and no participation in any music theory classes. Our participants also reported no significant history of communication, hearing, or neurological disorders. With regard to hearing status, our participants had thresholds of 25 dB HL or better bilaterally for all frequencies tested (125–4000 Hz) and exhibited 10 dB HL or less asymmetry between the two ears at all frequencies.
Stimuli
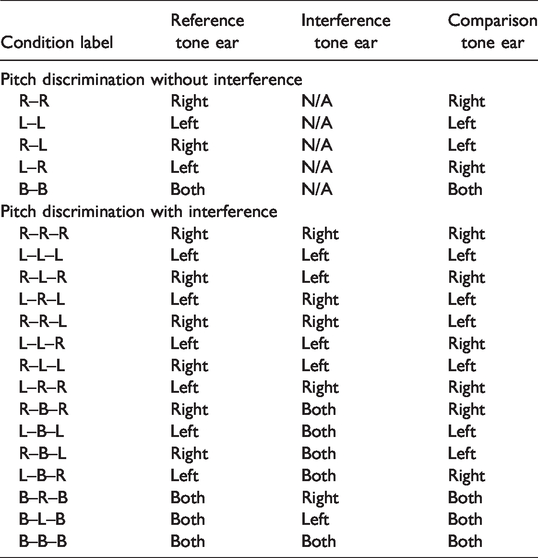
We created 20 pitch conditions for auditory stimuli (see Table 1). Five conditions were pitch discrimination trials with no interference present, while the remaining 15 conditions were pitch discrimination trials with tonal interference present. Using Adobe Audition sound editing software (Version 1.5), all tones were generated with three equal amplitude harmonics, gated on and off, with a 10-ms linear amplitude ramp, each 350 ms in duration.
Condition Labels and Ear of Stimulation for Pitch Discrimination With and Without Interference.
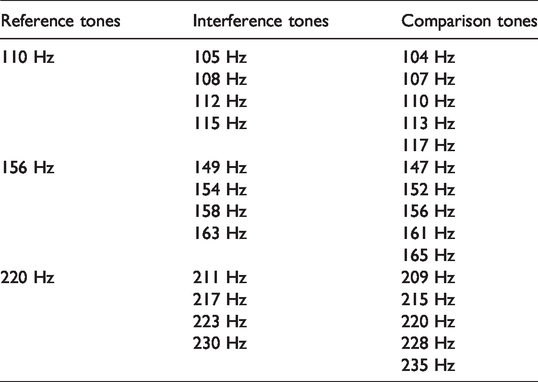
Three reference complex tones were generated with fundamental frequencies of 110 Hz, 156 Hz, or 220 Hz. Fifteen comparison complex tones were generated, differing from each reference tone by –100, –50, 0, +50, or +100 cents (100 cents = one half step in an equal tempered scale; see Table 2). Low frequencies were used as stimuli due to findings that frequency discrimination ability is best in lower frequencies (Wier, Jesteadt, & Green, 1977) and based on prior research utilizing low-frequency stimuli to examine pitch discrimination and memory ability when interfering tones are present (Moore et al., 2007; Semal et al., 1996).
Frequency Relationship Among Reference, Interference, and Comparison Tones.
For no-interference conditions, we used the following presentation sequence: silence (250 ms), reference tone (350 ms), silence (4,000 ms), comparison tone (350 ms), and silence (250 ms). Of these five pitch discrimination conditions, two were same-sided conditions, with both the reference and comparison tones presented to either the right ear (R–R) or to the left ear (L–L). Two were opposite-sided conditions in which the reference tone was presented to either the right ear with a contralateral left ear comparison tone (R–L) or the reference tone was presented to the left ear with a contralateral right ear comparison tone (L–R). Finally, in the remaining condition, we presented both the reference and comparison tone binaurally (B–B).
In the remaining 15 conditions (6–20), we tested pitch discrimination abilities with tonal interference present. In four interference conditions, we used complex reference tones with frequency differences of –75, –25, +25, and +75 cents (see Table 2). These more complex tones were 250 ms in duration and were presented randomly between the reference and comparison tones. Because there was no period of silence separating interference tones, they had a total duration of 1,000 ms for each trial. The presentation sequence for these interference pitch discrimination tasks was as follows: silence (250 ms), reference tone, silence (1,500 ms), four interference tones (250 ms each, totaling 1,000 ms), silence (1,500 ms), comparison tone, silence (250 ms). We presented the reference and comparison tones to the left side, to the right side, and to both sides bilaterally; we presented the interference tones both monaurally and binaurally. Thus, the reference, comparison, and interference tones were presented with the reference and comparison tones to the right ear and the interference tone to the right ear (R–R–R) or left ear (L–L–L), or with reference and comparison tones to either the right or left ear with the interference tones presented to the other side (R–L–R or L–R–L), for a total of 75 trials. Pitch discrimination trials with and without interference were grouped together randomly in four blocks. Each of the 75 trials was presented twice per block, for a total of 150 trials per block, equaling 600 trials for each participant.
Procedures and Equipment
We administered the tasks across two separate sessions, each lasting approximately 75 minutes. The duration between the two sessions was no more than three weeks. The 20 conditions were randomly assigned into four blocks of five, with two blocks presented per session (i.e., a pre-experimental session of 10 blocks and an experimental session of 10 blocks). Within each block, we used a randomization software program to present the trials in random order, having counterbalanced the block order across participants.
We completed all preliminary audiometric procedures and experimental tasks in a double-walled, sound-treated room that met specifications for maximum permissible ambient noise levels (American National Standards Institute, 2003). During the first session, participants completed the Annett Hand Preference Questionnaire (Annett, 2004), read and signed a statement of informed consent, and completed a bilateral pure tone threshold test. The examiner also administered a case history questionnaire with a primary focus on previous communication or neurological disorders, as well as musical ability and background.
The pitch discrimination tasks with and without interference conditions were carried out using a Tucker-Davis Technologies system and a Hewlett-Packard ×1100 personal computer. For these tasks, participants were seated at a table inside a double-walled sound-treated booth, with a 17-inch color monitor and wireless mouse. The experimenter monitored participant progress on a networked computer from outside the sound booth. We presented stimuli using the ECos/Win software program (AVAAZ Innovations, Inc., Ontario, Canada), and we routed them through the RP2.1 enhanced real-time processor, PA5 attenuator, and HB7 headphone driver, before presenting them to participants through Sennheiser HD 265 linear supra-aural headphones. We presented the tonal stimuli at 75 dB SPL.
We asked participants to judge whether the final, or comparison, tone was the same as or different from the initial or reference tone in pitch by using a mouse to select either a same or different icon on the computer monitor. We used a short practice block at the beginning of each session, consisting of four trials of each task (pitch discrimination without interference, pitch discrimination with interference). There were both same and different tone pairs for each set presented. To make the practice task easier, the stimuli were generated separately and had larger differences in pitch between the reference and comparison tones on the different trials than was the case for experimental stimuli. For all tasks, data from each trial were recorded by the Ecos/Win software and exported to a spreadsheet file for further analysis.
Data Analysis
We saved responses from each block in an Excel spreadsheet file and first assessed data for intraparticipant reliability. We analyzed each trial to determine a total number correct score for each condition, and we conducted a Pearson product-moment correlation between the two experimental runs for every condition. We found all conditions to be at least moderately and significantly correlated at the p < .05 level, with the lowest correlation coefficient (r) equaling .454 (p = .026), indicating good intraparticipant reliability. Thus, we were able to average the results of the two experimental runs and use them in further data analysis.
To assess differences in overall accuracy among the 20 conditions, we conducted a one-way repeated measures analysis of variance (ANOVA) using Greenhouse–Geisser corrections for sphericity. We made post hoc pairwise comparisons using Bonferroni adjustments. For further analysis, we collapsed data across the 20 conditions into five groups of conditions: (a) all same, meaning that all stimuli types (reference, interference, and comparison stimuli) were presented to the same ear; (b) reference different, meaning that reference stimuli were presented differently from interference and comparison stimuli; (c) comparison different, meaning that comparison stimuli were presented differently than the reference and interference stimuli; (d) interference different, meaning that interference stimuli were presented differently from reference and comparison stimuli; and (e) all different, meaning that the three types of stimuli were presented differently (e.g., right reference-binaural interference-left comparison). These means were analyzed using an additional one-way repeated measures ANOVA with five levels. For all statistical analysis, an alpha level of .05 was used to determine significance. It should be noted that the frequency of the reference tone was found to be nonsignificant, and, therefore, only a total number correct score per condition was analyzed in further statistical analyses.
Results
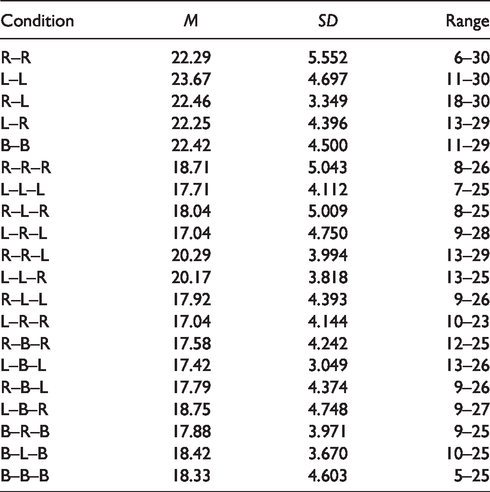
Table 3 shows the mean number correct scores and standard deviations and the range of scores (out of a possible 30 for each condition) for each of the 20 conditions across all participants. As expected, scores for the no-interference pitch discrimination task were more accurate than pitch discrimination scores interference present. We considered the binaural (B–B) condition of both reference and comparison tones without interference to be the baseline task. In this condition, most number correct scores fell between 20 and 25 (66%–83%); there was no perfect score of 30 (100%).
Means, Standard Deviations, and Range (Out of Possible 30) for Total Correct Scores of All Pitch Discrimination Conditions Without Interference (1–5) and With Interference (6–20).
To evaluate differences among the 20 conditions, we conducted a one-way repeated measures ANOVA. As the Mauchly’s test of sphericity was significant (p = .003), we utilized Greenhouse–Geisser corrections for further analysis. There was a significant effect for conditions, F(9, 211) = 17.138, p < .001, ηp2 = .427. To further investigate these differences, we divided the 20 conditions into five pitch discrimination conditions without interference and 15 pitch discriminations with interference. ANOVAs of pitch discrimination in conditions without interference revealed no significant differences, indicating that the ear that received auditory input had no significant effect on pitch discrimination performance when there was no interference. Analysis of pitch discrimination in conditions with interference (one-way repeated measures ANOVA with 15 levels) revealed a significant difference between conditions, F(14, 322) = 3.748, p < .001, ηp2 = .14, with post hoc pairwise comparisons indicating that participants were significantly more accurate in conditions in which the reference and interference tones were presented to the same ear, with the comparison tone presented to the other ear (R–R–L, M = 20.29; L–L–R, M = 20.17) than in conditions in which the interference tones only were presented to the other ear or presented bilaterally (L–R–L, M = 17.04; L–B–L, M = 17.42). In addition, in condition L–L–R (reference and interference tones presented to the left, M = 20.17), participants performed significantly better compared with condition L–R–R (reference tone presented to the left, and interference and comparison tones presented to the right, M = 17.04).
To further examine these data, we analyzed total correct scores across five groups of conditions: (a) all same, meaning that all stimuli types (reference, interference, and comparison stimuli) were presented to the same ear; (b) reference different, meaning that reference stimuli were presented differently from interference and comparison stimuli; (c) comparison different, meaning that comparison stimuli were presented differently than the reference and interference stimuli; (d) interference different, meaning that interference stimuli were presented differently from reference and comparison stimuli; and (e) all different, meaning that the three types of stimuli were presented differently (e.g., right reference-binaural interference-left comparison). Statistical analysis (ANOVA) indicated significant group differences, F(4, 92) = 8.06, p < .001, ηp2 = .259, with participants performing significantly better in conditions in which the comparison tone was presented to a different ear than the reference and interference tones (Figure 1). This difference was significant in comparison to all other groups, except when presentation of stimuli was different for each tone type (all different). Additional ANOVA analysis examining ear of input (left vs. right vs. binaural) revealed that, while the ear of input for the reference and interference tones had no significant impact on performance, accuracy was significantly improved when the comparison tone was presented to the left ear (p = .045). It should be noted that this significant left ear advantage for comparisons tones was evident only when compared with right ear comparison tone inputs, not when compared with binaural comparison tone input.
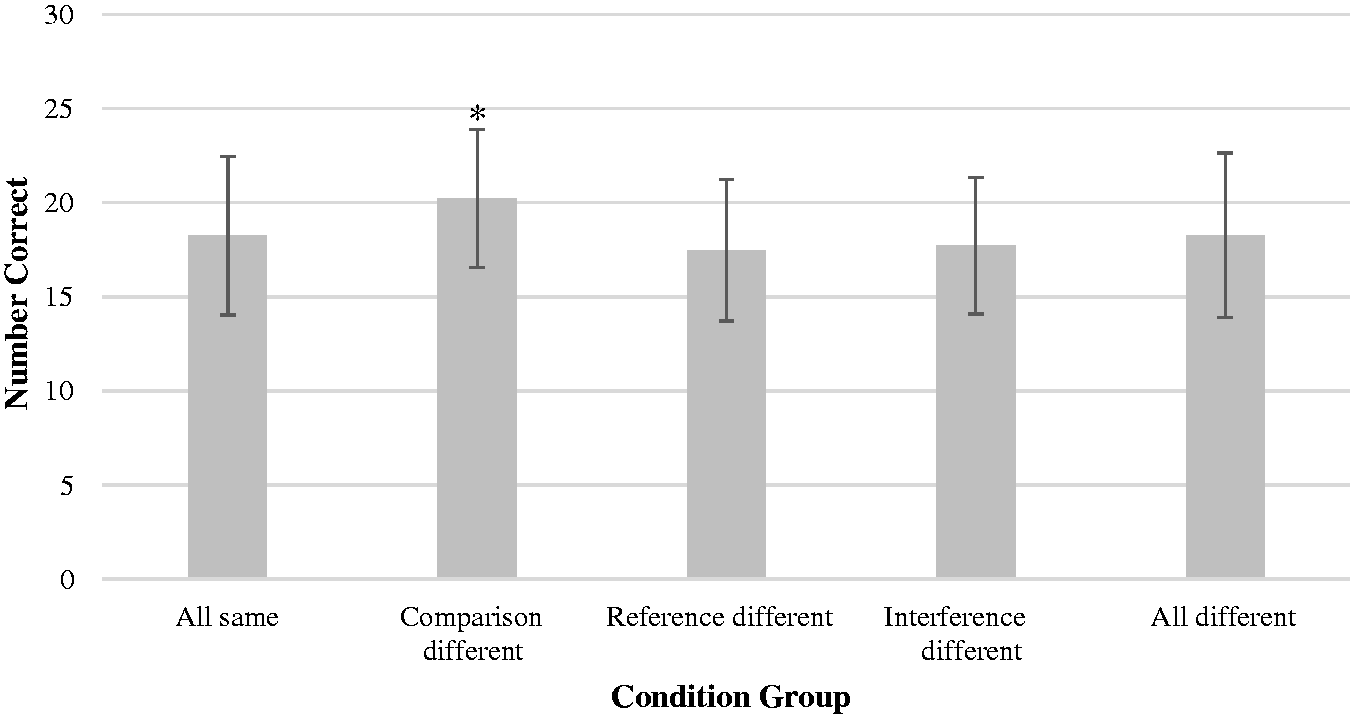
Means and standard deviations for condition by ear of input (* indicates significance, p < .05 in pairwise comparisons with three other conditions: all same, reference different, and interference different).
Discussion
In this study, we sought to better understand the impact of spatial (i.e., right, left, and binaural) sound locations and their interrelationship for reference, interference, and comparison tones in pitch perception within a sample of 24 young female participants of normal-hearing ability without prior musical experience or training. We studied these participants’ performance in conditions in which interference was and was not present, and we varied the side or stimuli presentation for all three types of tones (i.e., reference, interference, and comparison tones). Overall, our findings revealed that (a) pitch discrimination accuracy was significantly better for trials without than with interference; (b) spatial location of input only significantly impacted on pitch discrimination accuracy in the presence of interference; and (c) pitch discrimination accuracy was significantly better for trials in which the comparison tone was presented to a different ear than the reference and interference tones.
Comparing Tonal Pitch Perception With and Without Interference Tones
The results of this study were consistent with our hypothesis and with previous findings of improved pitch discrimination accuracy in no interference versus interference conditions (Deutsch, 1970a; Moore et al., 2007). All five conditions without interference present showed higher mean number correct scores than any of the 15 conditions with interference present (see Table 3). However, only ipsilateral presentation of reference and comparison tones to the left ear (i.e., the L–L condition, M = 23.67) showed significantly better performance compared with all conditions in which interference was present.
Spatial Location of Tonal Input
Regardless of spatial location of input of reference or comparison tones, pitch discrimination accuracy was stable when no interfering tones were present. However, with the addition of interference tones, there was a spatial relationships impact on pitch discrimination accuracy (Figure 1). Unlike prior research (Deutsch, 1978a) indicating improved pitch discrimination with spatial separation of the input location for interference tones, we found that pitch discrimination accuracy was highest when the reference and interference tones were presented to the same ear while the comparison tone was presented to the other ear (R–R–L, M = 20.29; L–L–R, M = 201.7). In fact, among monaural trials, in direct contrast to previous findings, we found pitch discrimination accuracy to be lowest when the reference and comparison tones were presented to the left ear while the interference tone was presented to the right ear (L–R–L, M = 17.04). Participants performed significantly better when the comparison tone was presented both to the left ear and to the opposite ear, relative to the other tones. In the context of varying the ear for the comparison tone, ear of input for the reference and interference tones did not significantly impact pitch discrimination. These findings indicate that the spatial location of the comparison tone is particularly important. The important factor seems to be the contralateral presentation of the comparison tone when both the reference and interference tones are presented to the same ear.
Our findings seem to contradict Deutsch (1978a) and Gockel et al. (2009) who found that spatial separation of input location for interference tones, relative to same-sided presentation of reference and comparison tones, improved pitch discrimination accuracy. However, neither of these two studies examined conditions in which the comparison tone was presented to a different ear than the reference tones. In addition, both studies utilized a blocked presentation style, in which the reference, interference, and comparison tones were each consistently presented to the same ear, even when their combination differed. It was later found that this predictability between side of presentation and type of tone aided the participants’ pitch discrimination (Kallman et al, 1987). Accordingly, we randomized the type of tone presentation to each ear, increasing the task difficulty and thus decreasing our participants’ performance accuracy. Also of note, Deutsch (1978a) utilized only proficient participants who scored at least 80% correct on a preliminary pitch discrimination task.
We suggest that short-term memory may be a factor in our findings, as spatially separating the comparison tone from the interference tone (through their contralateral presentation) may have assisted short-term memory and thereby allowed for more accurate perceptual discrimination between the comparison and interference tones than when these two tones were presented to the same side. The conditions in which the interference and comparison tones were presented ipsilaterally to a different ear than the reference tone (R–L–L and L–R–R) showed lower number correct scores than many other interference conditions (see Table 3). Theoretically, this pattern should also be seen, perhaps even more so, in conditions in which the interference tone was spatially separated from both reference and comparison tones, and in fact, this pattern has been found in previous research (Deutsch, 1978a; Gockel et al., 2009; Kallman et al., 1987) and was suggested in this study by relatively poor accuracy scores under these conditions (R–L–R, M = 18.04; L–R–L, M = 17.04). Again, however, findings of the previous studies were obtained only when tone presentations were blocked rather than randomized. When using random presentation, better performance for contralateral presentation of the interference tones in relation to reference or comparison tones would not necessarily be expected, and, in fact, we found no such advantage. Presentation of the reference and comparison tones to the left ear with interference tones to the right ear (L–R–L) yielded one of the lowest mean number correct scores. It is possible that attentional interaction between reference and interference tones presented ipsilaterally reduces the disruptive effect of the interference tones on pitch memory. Selective attention is known to affect subcortical activity at the level of the brainstem in the processing of spatial and spectral components of sound (Lehmann & Schonwiesner, 2014), but further research is needed to explore the full spectrum of behavioral ramifications of these findings.
Alternatively, interference tones may have a more significant effect on the perception of the comparison tone than the reference tone. This idea is contrary to most research that is currently available on pitch memory, as most current theories focus on interference of short-term memory traces (Baddeley & Hitch, 1974; Cudahy & Leshowitz, 1974; Salame & Baddeley, 1989). Perhaps participants without musical experience or training utilize different memory strategies than musically experienced participants in other studies. Music experienced participants may rely more on the final comparison tone than the initial reference tone to determine a same/different response. Gaab and Schlaug (2003) found that nonmusicians rely more on the brain’s lower order processing centers than musicians. Nonmusicians may rely on different processing or memory strategies in pitch discrimination tasks. If nonmusicians depend more on the final comparison tone to determine a same/different response, then interference tones may have a more detrimental effect on processing the comparison than the reference tone, making the spatial separation of interference and comparison tones more important to them for accurate pitch discrimination. Further research comparing musical and nonmusical participants in their processing of leading and trailing interference tones is warranted.
Finally, there is some evidence of a trade-off between attention and memory in auditory processing (Heinrich, Schneider, & Craik, 2008). As the attentional demands of a task increase, cognitive resources allotted to memory must decrease. If so, then the input location transition from reference to comparison tones may require more attention, disrupting the memory trace. In this study, the conditions with the lowest mean PDA scores were those in which interference tones were presented contralaterally to the reference tone (L–R–L, L–R–R). Research suggests a dynamic interplay in the brain’s processing of auditory information, between higher order cognitive tasks, such as attention, and lower order tasks involving frequency and spatial information (Lehmann & Schonwiesner, 2014), but further research is needed to determine the implications of this study’s findings on perceptual discrimination.
Participant Characteristics
While all participants in this study were considered nonmusicians, there was considerable variability among participants with regard to musical experience. Some participants had absolutely no musical experience, while many others exhibited some degree of musical background, ranging from a few private piano, guitar, or voice lessons to 10 years of ensemble playing or singing (i.e., band, chorus, and church choir). It should be noted that several participants with more considerable musical experience were heard humming during the experiment, and these participants generally had higher PDA scores than participants who did not hum. This information suggests that individuals who have little to no formal musical training can use assistive strategies, such as humming, to perform better.
Previous research in pitch discrimination has sometimes encountered a ceiling effect, in that some participants showed 100% accuracy (Deutsch, 1970a; Moore et al., 2007). In the present study, however, participant scores ranged from 37.5% to 85.33%, averaging 64.36%, illustrating no ceiling effect. In comparison to the Moore et al. (2007) study, our stimuli were similar in frequency and identical in duration, and our participants were similar in age, handedness, and hearing ability. Thus, it is most likely that our participants’ decrease in PDA scores was a result of our participants’ lack of musical training. Moore et al. (2007) did not screen participants for level of musical training. Numerous other studies have found that especially early musical training, versus no training, significantly enhances pitch discrimination (Amir, Amir, & Kishon-Rabin, 2003; Estis et al., 2011; Pechmann & Mohr, 1992). Also, lower scores seen in this study may have been due to participant fatigue. Our participants engaged in two sessions of about 105 minutes each, and several participants reported minor difficulties attending to the task by the end of each session.
Limitations and Directions for Future Research
A limitation of the current study was an imbalance in the number of trials of same and different reference and comparison tones. Future studies should ensure equal numbers of same and different trials to more accurately examine perceptual sensitivity and confirm this study’s findings. In addition, future researchers should examine how participant age affects pitch discrimination by utilizing children, including those with auditory processing or language deficits who have been shown to often exhibit poor pitch processing skills (Bellis, 1999; Bishop, 2000; Clement, Planchou, Beland, Motte, & Samson, 2015; Loucas et al., 2008). Future researchers could examine differences in pitch processing skills between these populations and typically developing peers. The effects of ear of stimulus input on pitch discrimination abilities in these populations may prove to be significant, as children with auditory processing disorders may exhibit problems with interhemispheric transfer of auditory information (Bellis, 1999), a crucial skill in single-sided auditory input that could be a useful diagnostic tool for brain processing inefficiency.
Conclusion
The present study confirmed previous findings showing that PDA was affected by the presence of interfering tones and that discrimination accuracy was better when the reference and comparison tones were the same. We also found that accuracy was improved when the comparison tone was presented to the left ear. Perhaps one of the most interesting and yet puzzling outcomes of the current study was that those conditions in which reference and interference tones were presented to the same ear and the comparison tone was presented to the opposite ear had higher accuracy scores than other conditions. Collectively, these findings suggest that the relative spatial location of the comparison tone is the most important factor in pitch discrimination accuracy.
While replicating research is needed, our findings suggest a hierarchy of difficulty in pitch discrimination tasks that might be utilized in designing pitch training programs for such clinical populations as people with certain subtypes of auditory processing disorder and specific language impairments. Tasks in this hierarchy progress from easiest (pitch discrimination with no interference) to most difficult (discrimination tasks in which the comparison tone is spatially separated from the reference and interference tones). Training individuals along this continuum should help them listen to more finely tuned differences in pitch and might strengthen short-term memory for pitch and sharpen auditory attention skills. For example, initial training should focus on pitch discrimination tasks without interference, and, as discrimination skill improves, interfering tones could be added, but with spatial separation of the comparison tone to aid in discrimination. As pathways are further strengthened and discrimination further improves, the spatial location of the comparison tone could then be presented binaurally and then monaurally to the same side as the reference/interference tones (most difficult). Use of this training hierarchy may strengthen neural pathways used in pitch discrimination to assist suprasegmental speech processing.