
Abstract
Performing bilateral saccadic eye-movements has enhanced subsequent verbal recall among consistent handers (people who use their dominant hands for virtually all tasks) but not inconsistent handers (people who use their non-dominant hands for some tasks); but eye-movements prior to encoding have disrupted recall in consistent handers. We sought to better understand how this research might be applied to recalling written text presented in ways that differentially affected eye movements. We manipulated text column width in order to test whether induced eye movements and resultant right hemisphere activations would lead consistent handed (versus inconsistent handed) readers to experience encoding disruptions and poorer recall of the written passage. We presented participants, divided by handedness consistency and gender, with a story about a fictional island in which the lines of text were either ∼28 or ∼120 characters wide. We later asked participants to answer 20 questions about the story and measured their memory performance by both their percentage of correct answers and by their correct answers adjusted for a guessing tendency. We found a handedness by text width interaction. Consistent handers who read the story in a narrower column width showed poorer recall than both inconsistent handers and consistent handers who read the story in the wider text column. We suggest a chain of events such that text width altered eye-movements that, in turn, activated right hemisphere brain processes associated with orienting attention and attentional control, competing with consistent handers’ encoding.
Keywords
Introduction
Bilateral saccadic movements (i.e., moving the eyes in both left and right directions) are important for reading. Studies have shown that patterns of bilateral saccadic eye movements distinguish readers such that skilled readers tend to make larger saccadic jumps and engage in less backtracking than less skilled readers (Rayner, 1998). Bilateral eye movements are also associated with brain lateralized cognitive tasks (Kocel et al., 1972) in that rightward eye movements are associated with better performing left hemisphere controlled tasks and leftward eye movements are associated with better performing tasks controlled by the right hemisphere.
Research outside of the reading context suggests that there are also attentional processes recruited by bilateral eye movements that can enhance long term memory (LTM) retrieval. Christman et al. (2003) conducted two studies in which they found that participants who performed bilateral saccadic eye movements subsequently showed fewer false recollections on oral verbal list learning and also recalled more information from a written journal they kept over six days than did control group participants who performed no eye-movements. In a separate study, Christman et al. (2006) reported that participants who performed bilateral saccadic eye movements also recalled earlier childhood memories than participants in a control group. Christman et al. (2006) explained their findings in terms of the Hemispheric Encoding and Retrieval Asymmetry (HERA) hypothesis (Habib et al., 2003), positing that the left cortical hemisphere becomes relatively more active during information encoding while the right hemisphere becomes relatively more active during episodic retrieval. Thus, Christman et al. (2006) proposed that bilateral saccadic eye movements increase interhemispheric interaction, allowing for more transfer of the memories activated during right hemisphere mediated retrieval.
This interpretation has been challenged recently by Lyle and Martin (2010) who used a tachistoscopic letter matching task to demonstrate that bilateral eye movements do not seem to increase interhemispheric transfer. They proposed that bilateral eye movements enhance top down attentional control that, in turn, enhances LTM retrieval. This attentional theory has gained further support and is consistent with the role bilateral saccadic eye movements have had in increased orienting attention (Lyle & Edlin, 2015).
An interesting interaction has been observed in studies of the effects of bilateral eye movements on episodic memory indicating likely elements of truth to both the explanations of hemispheric asymmetry suggested by Christman et al. (2006) and attentional processing suggested by Lyle and Martin (2010). Lyle et al. (2008) conducted two studies in which participants were presented with a list of 50 words and then given either free recall tests (in one study) or recognition tests (in the other study). Lyle et al. divided their participants into consistent and inconsistent handers, since handedness consistency (strength of preference for one’s dominant right hand) was expected to be negatively associated with heavy reliance on right hemisphere cognitive processing (Prichard et al., 2013). They found that only consistent handers benefited from bilateral eye movements for either enhanced free recall or recognition memory. This suggests that increased right hemisphere activitation from bilateral eye movements selectively enhanced retrieval in consistent handers. More precisely, Christman and Butler (2005) found that when eye movements were performed prior to encoding, consistent handers showed memory detriments, but when consistent handers performed eye movements prior to retrieval, they had higher recall and fewer false alarms. Eye movements prior to retrieval seemed to give consistent handers increased access to beneficial right-hemisphere-based processes, enhancing word retrieval; yet, eye movements prior to encoding were detrimental to retrieval. Brunye et al. (2009) found no detrimental effects of horizontal eye movements on retrieval of visual information.
Attentional control is directly relevant to this discussion. A broad literature has shown that the right hemisphere is particularly involved with attention processes, particularly attention toward the broader environmental context, even to the point of directing the left hemipshere’s focused attention (e.g., McGilchrist, 2010; Ornstein, 1997). Hence, increased right hemisphere activity might increase attentional control, even without necessarily activating inter-hemispheric communication through the major commissures between the two hemispheres. Nobre et al. (1997) showed the importance of the right hemisphere for visual attention processes. They recorded PET-measured changes in brain activity in six right-handed males following two tasks designed to induce attention with bilateral eye movements. In both tasks, these participants focused on a central fixation point with a visual angle width of 0.5 degrees. At either side of the central fixation was a peripheral square that was one degree wide and located at seven degrees eccentricity. In one task, one of the peripheral squares brightened before the appearance of a visual target (80% of the time) on the same side as the peripheral square. During the second task, the brightening of the peripheral square predicted the appearance of the visual target on the opposite side (80% of the time). These tasks were designed to elicit attention orienting eye movements. Eye movement shifts to the right visual field tended to activate attentional regions bilaterally and shifts to the left visual field predominately activated right hemisphere areas, meaning that there was relatively greater right hemisphere involvement across the visual attention tasks.
Among the brain areas that became relatively more active during Nobre et al.’s (1997) visual processing tasks were the right anterior cingulate gyrus and the posterior parietal cortex. These activations are of particular interest because some research has suggested that attentional networks can be roughly divided into (a) alerting, (b) orienting, and (c) executive functioning (Fan et al., 2002). The alerting system is associated with activity in the right frontal cortex and the right interior parietal lobe. Orienting activity is associated with activity in the temporal parietal junction (part of the inferior parietal cortex). Executive control is associated with activity in the anterior cingulate cortex (ACC) and the dorsolateral prefrontal cortex (DLPFC) (Chica et al., 2012; Fan et al., 2002). Furthermore, Chica et al. (2012) found that inducing phasic alertness (activating the alerting system) helped patients with right hemisphere damage and varying degrees of hemineglect reduce their left-sided orienting deficits. Thus, these three attentional networks seem to interact such that increasing alerting activity may facilitate performance among the other attentional systems.
Bilateral saccadic eye-movements performed by participants in the aforementioned studies might have engaged the orienting and executive control networks. Edlin and Lyle (2013) showed that bilateral saccadic eye movements improved participants’ performance on the attentional control (executive function) aspect of the Attention Network Test (Fan et al., 2001) though these bilateral saccadic eye movements seemed to have no effect on participants’ performance on the orienting attention part of the test. This mixed finding is consistent with Lyle and Edlin’s (2015) observation that increased top-down attentional control underlies the effects of eye movements on memory.
Importantly, recent evidence also suggests that disrupting attention has a detrimental effect on memory retrieval for visual information (Craik, 2014). Craik (2014) reviewed a series of studies showing that participants with their eyes closed better recalled and recognized visual information than participants who were visually distracted. Seemingly, visual attentional resources were recruited during the eyes closed retrieval processes, while visual distractions disrupted the retrieval process. The effect of distractors seemed to be modality specific (i.e., visual) as memory for visual information was not disrupted by auditory distractors (which would not have recruited as extensively from the right hemisphere). Craik also noted that researchers in prior studies disrupted performance of attention demanding secondary tasks during encoding but only disrupted retrieval if the secondary task and the material to-be-remembered were similar. He argued that memory encoding competes with other tasks for general resources, while retrieval competes for domain specific resources.
Collectively, these various threads of prior research create an emerging picture. Most prior eye movement research required participants to remember visual information (e.g., Christman et al., 2003; Lyle et al., 2008), and recall of this information should heavily recruit right hemisphere brain processes (Nobre et al., 1997), especially those associated with orienting attention (the right temporal parietal junction) and top down attentional control (Chica et al., 2012; Fan et al., 2002). Thus, eye movements should enhance the retrieval of visual information by recruiting right hemisphere visual-spatial and attentional processing, especially among consistent handers whose access to right hemisphere processing is less facile (Prichard et al., 2013).
It is important to note here that hemispheric “access” need not mean “callosally mediated” access. Inconsistent handers, with easier right hemisphere access, might be expected to have a more active attentional control network. Remember that, although Edlin and Lyle (2013) found that eye movements specifically improved attentional control, but not orienting attention, Chica et al. (2012) showed that activating the alerting system can modulate activity in the other systems. Possibly, inconsistent handers may most activate the brain’s alerting system, but more research is needed to be certain of this. From research to date, we would expect inconsistent handers to show stronger right hemisphere mediated attentional processes, especially those related to attentional control, in their everyday cognitive functioning. This might explain why inconsistent handers have shown a general advantage for episodic recall (see Christman & Propper, 2010; Prichard et al., 2013 for reviews). Similarly, eye movements might have relatively greater retrieval benefits for consistent handers whose bilateral eye movements then facilitate access to right hemisphere functioning by recruiting domain-specific attentional right hemisphere resources that enhance retrieval, even while they compete with encoding for general resources, perhaps leading to the interfering effect shown by Christman and Butler (2005). Thus, inconsistent handers might show little relative retrieval benefit from eye movements, given their already active attentional networks.
These possible implications of the foregoing research have not yet been fully tested during reading for which saccadic eye-movements are critical (Rayner, 1998). The direction of saccadic eye movements during reading may be associated with the selective activation of different brain hemispheres. Thus, the question arises as to whether actively manipulating a reader’s eye movements by manipulating the text layout on a page might affect the reader’s comprehension and recall. In the present study, we set up an experimental scenario in which a text passage was presented with different text widths in order to manipulate how much bilateral eye movement occurs during reading. Since continuous eye movements in one direction can activate the contralateral hemisphere (Ornstein, 1997), continuous leftward jumps might activate the right hemisphere (just as attention related saccadic jumps have been associated with increased right hemisphere activity). Also, according to the HERA model (Habib et al., 2003), the left hemisphere is relatively more active during encoding memory. One means of manipulating readers’ right or left hemisphere activity and their associated memory retrieval and encoding processes might be to alter saccadic eye movements by changing the text column width of the prose being read. More specifically, a wide text column layout induces long continuous rightward eye movements that would activate left hemisphere controlled encoding processes and compete with the right hemisphere dominated visual attention network for general resources. By contrast, a narrower text column layout results in more frequent bilateral and leftward eye-movements that orient attention to the start of a new line of text, and this process might be disruptive for the encoding process, especially for consistent handers because it would activate right hemisphere attentional processes that compete with left hemisphere encoding for general resources
The purpose of the present paper was to test the hypothesis that text layout might affect what readers can remember, specifically with respect to episodic and cued recall. Because episodic memory has a greater visuospatial component, it should rely more on recruitment from right hemisphere controlled visual attention processes, meaning that any effect of text column width should be larger with respect to episodic recall and when readers are consistent handers. Additionally, recent research suggests that eye-movement effects are specific to conceptually explicit memories, such as for cued recall (Parker et al., 2018). Accordingly, the present study adapted the methods and materials from a recently developed paradigm used to investigate handedness consistency differences in cued recall for prose. Our specific hypothesis was that consistent handers who read information presented in narrow columns would show relative memory deficits because eye movements during encoding would selectively activate right hemisphere processes in consistent handers. Conditions which would falsify our hypothesis include:
Any main effects of handedness (e.g., inconsistent handers performing better across conditions), but no handedness by condition interaction. A condition by handedness interaction in which consistent handers outperformed inconsistent handers in any condition. A condition by handedness interaction in which consistent handers in the wide columns condition performed worse than consistent handers in the narrow columns condition. A failure to obtain any significant effects.
Method
Participants
Our study participants were 120 students (Mage = 19.23 years, SD = 1.63; 76 females, 44 males) from a large Midwestern University who were enrolled in an introductory psychology class. All participants gave their written informed consent to participate in the study, and the study’s protocol was approved by the University’s Institutional Research Board.
Materials
Edinburgh Handedness Inventory
To measure the participants’ handedness consistency, we used The Edinburgh Handedness Inventory (Oldfield, 1971), frequently referred to as the EHI. This is a 10-item questionnaire listing 10 common manual tasks and or items that require the use of one hand. The EDI version used in the present study was slightly modified from the original in its content and scoring system (for information regarding the factor structure of the version used in the present study, see Christman et al., 2015). The 10 items/tasks were Writing, Spoon, Open Jars, Toothbrush, Throwing, Comb Hair, Scissors, Kinfe, and Striking a Match. Participants had five response options for each task: always left, usually left, no preference, usually right, and always right. For each item a score of −10 was assigned for the always left option, a score of −5 was assigned for the usually left option, a score of 0 was assigned for the no preference option, a score of 5 was assigned for the usually right option, and a score of 10 was assigned for the always right option. EHI item total raw scores ranged from −100 to 100. We assigned participants to consistent handedness and inconsistent handedness groups by taking a median split of the EHI’s absolute value scores such that participants who scored in the lower half of the test (suggesting a less lateralized hand preference) were considered inconsistently handed, and participants who scored in the upper half (suggesting a more lateralized hand preference) were considered consistently handed.
Research comparing consistent and inconsistent handers has frequently used a median split of handedness data in order divide participants into groups (Prichard et al., 2013). This method warrants further discussion, as, by this definition, an inconsistent hander is not necessarily ambidextrous. A participant might use their non-dominant hand for only one or two tasks to be classified as inconsistently handed. Although the distinction between consistent handedness and inconsistent handedness may seem small, there is empirical evidence that this method of grouping yields meaningful differences. Prichard et al. (2013) reviewed literature showing that this method of grouping participants predicted participant differences in multiple research paradigms whereas previous research that compared left versus right handers yielded few or no correlates on some of these variables of interest. In addition, by grouping participants in this manner, Christman et al. (2015) demonstrated that consistent and inconsistent handers showed different factor structures in their responses to the EHI. Specifically, consistent handers showed a one factor solution, while inconsistent handers showed a two factor solution, suggesting that handedness is more complicated for inconsistent handers. When classifying participants into consistent versus inconsistent handedness through discriminant function analysis, those EHI items that loaded onto the second factor were the items that discriminated between consistent and inconsistent handers (Prichard et al., 2020). Thus, a median split on the EDI is an effective handedness consistency classification system.
Importantly for this study, Lyle et al. (2012) investigated whether or not handedness consistency and direction of handedness interacted to predict memory performance. They found that inconsistent handers showed better memory performance irrespective of whether or not they were left or right-handed. For the purpose of memory research, current evidence suggests that these variables can be considered orthogonal and non-interactive. For this reason, we only recorded and considered consistency and not direction of handedness.
Reading Sample: Spinoza Story Island and Questions
All participants read a 229-word story about a fictional island called Spinoza Island. Later, at testing, participants were asked to write their responses to 20 questions about the story. The questions were designed to force participants to recall story details. These materials were also used in a previously published study (Prichard & Christman, 2017). The story is included in Table 1, and the 20 questions are included in Table 2. Just as with the procedure used by Prichard and Christman (2017), we gave our participants a set amount of time to write down as many answers to the questions as they could remember. Responses did not have to be in complete sentences, nor did respondents have to use correct spelling, as long as the intended response could be ascertained. For example, “Eistien” would be considered a correct response to question number two, even though it is a slight misspelling.
Island Story From Prichard and Christman (2017).

Island Story Questions From Prichard and Christman (2017).

Questions were scored with a “1” for a correct response and a “0” for an incorrect response. The dependent variable was the percentage of correct responses. This was found by dividing the number of correct responses by 20 and multiplying the quotient by 100. The primary purpose of the questions was to gauge declarative memory for factual information about the story, as opposed to comprehension. Although the initial reading was manipulated by width of text presentation, all participants received questions with lines of the same character length, which was equal to the default character length for a Microsoft Word document written in Times New Roman 12-point font.
Procedure
At the experiment’s onset, the experimenter greeted the participants and instructed them to sit in one of two designated seats at either end of a long table. After obtaining participants’ signed consent, the experimenter provided instructions, telling the participants that they “may be asked some questions about the story” they were to read. Hence, the memory test was not a surprise.
We tested participants 1–2 at a time in the same room. Participants received the story face down. Half of the participants were randomly assigned to a reading condition in which the story text was presented in narrow columns and half were randomly assigned to text presented in wide columns. Wide columns had a width of ∼120 characters and narrow columns consisted of columns of ∼28 characters. According to McConkie and Rayner (1975) 12–15 characters is the maximum character length at which information useful to reading can be discerned. The 28-character columns were based on twice this number. This would yield approximately two rightward eye movements for every leftward jump back to the next line. The 120 characters were based on the maximum number of characters that could fit across a page. This would yield approximately 10 rightward eye movements for every leftward jump. All participants were given two minutes to read the story. After two minutes, participants were given 10 minutes to complete both the EHI and another questionnaire used as part of another study. After the 10-minute questionnaire period, participants had two minutes to answer as many questions about the story as they could. Unanswered questions were considered incorrect. In both studies, the timing was based on several pilot sessions in which the goal was to keep the experiments short enough to minimize participant boredom and to maximize efficiency, while still giving participants enough time to read through and respond to the materials. Participants were then debriefed by the experimenter.
Data Analyses
We analyzed data using a three-way between participants analysis of variance (ANOVA): 2 (Story Margin Width: Wide vs Narrow) × 2 (Handedness: Inconsistent vs Consistent) × 2 (Sex: Male vs Female). Sex was included as a factor because pilot data from an earlier study in which the “Island Story” materials were under development suggested that participant sex may modulate handedness differences (Prichard & Christman, 2012). Additionally, the first published study to use the “Island Story” also included sex as a factor (Prichard & Christman, 2017).
The first dependent variable was the percentage of correctly answered questions. The second dependent variable was a measure of signal to noise sensitivity. In addition to calculating the percentage of responses that were correct responses, we also counted the percentage of responses in which participants wrote an incorrect answer. This was important because some questions could have yielded correct responses through guessing. For example, question two asked participants to name a famous scientist. Some participants may have written “Einstein,” because they remember reading his name. Others may have just guessed “Einstein,” because he is such a famous scientist that he is the first to come to mind. People with a high tendency to guess might have had an artificially high hit rate. To account for guessing, we subtracted the z-scores for each participant’s number of incorrect guesses from the z-score for their number of correct responses, yielding a value similar in concept to d’ from signal detection theory. However, since d’ is typically calculated using recognition tests with critical lures, our value was not d’ per se. Instead, we called it “Signal Sensitivity,” and defined it as the rate of correct responses when accounting for the tendency to guess.
Prior to testing our main hypothesis, we conducted preliminary analyses to test the dependent variable’s data distribution for violations of the assumptions of normality. When the dependent variable was the percentage of correct responses, the overall mean percentage of questions answered correctly was 43.6% (SD = 1.6). The Shapiro-Wilk test for violations of normality failed to reject the null (p = .116), indicating that the data distribution was roughly normal. When the dependent variable was Signal Sensitivity, the mean was 0 (SD = 1.69), indicating that a participant’s z-score for correct responses was equal to their z-score for incorrect responses, a chance level of performance. A positive score indicated that they averaged either more correct or fewer incorrect guesses, suggesting a higher signal to noise ratio. A negative score indicated an average of either fewer correct responses or more incorrect guesses, or a higher noise to signal ratio. The Shapiro-Wilk test for violations of normality failed to reject the null (p = .200) indicating an approximately normal distribution.
As the median absolute EHI score was 80, we categorized scores of 80 and below as inconsistent handers and those with a score of 81 or above as consistent handers (A cell refers to a condition in a factorial design. A 2×2 design has four cells). We analyzed data using IBM SPSS statistics version 25. We set statistical significance at p < .05.
Results
Main Analyses
For the main three-way ANOVA’s [2 (Condition: Wide vs Narrow)×2 (Handedness: Consistent vs Inconsistent)×2 (Sex: Male vs Female)], the first dependent variable was the participants’ total percentage of correct responses. While this ANOVA yielded no significant main effects, there was a significant condition by handedness interaction, F(1, 112) = 3.9, p = .05, partialη2 = .034. After assigning participant data to cells that represented both handedness and text format conditions, there were 31 inconsistent and 29 consistent handers in the narrow text condition; and there were 33 inconsistent and 27 consistent handers in the wide text condition. Post-hoc analyses using the least squared difference test revealed that the recall performance of consistent handers in the narrow text condition (M = 36.3%, SD = 17.8) was poorer than the performance of participants in all three other cells, consisting of inconsistent handers in the narrow text condition (M = 46.4%, SD = 17.5; p = .026, d = .57, η2 = .075) consistent handers in the wide text condition (M = 46.2%, SD = 16.4; p = .034, d = .58, η2 = .078), and inconsistent handers in the wide text condition (M = 45.3%, SD = 17.2; p = .043, d = .50, η2 = .059). A graphical representation of this interaction is presented in Figure 1.
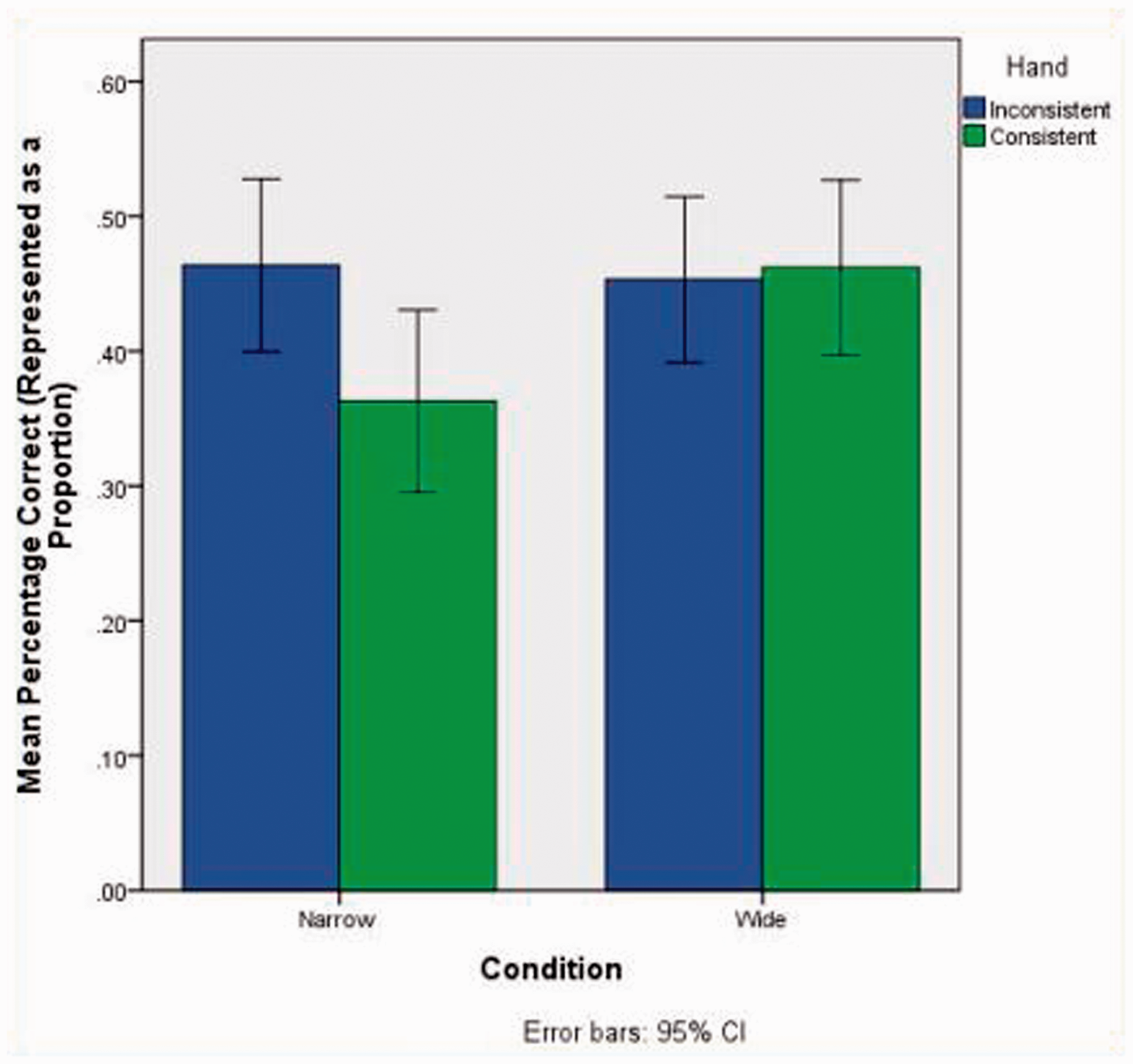
Condition by Handedness Interaction.
For the second dependent variable, signal sensitivity, there were no statistically significant main effects. However, there was a significant handedness by text condition interaction, F(1, 112) = 4.69, p = .033, partialη2 = .040. None of the paired comparisons were statistically significant. Regarding the practical significance of these effects, we calculated effect sizes and used Cohen’s d and η2 to compare the consistently handed/narrow text group (M = −0.426, SD = 1.76) to (a) inconsistent handers in the narrow column condition (M = 0.20, SD = 1.58; d = .375, η2 = .034); (b) consistent handers in the wide text condition (M = 0.38, SD = 1.51; d = .49, η2 = .057); and (c) inconsistent handers in the wide text condition (M = −0.126. SD = 1.83; d = .17, η2 = .007). Overall the signal sensitivity effect sizes were weaker than the correct response effect sizes. However, both measures indicated that consistent handers in the narrow condition performed more poorly.
To assist future replication research, we conducted two power analyses to suggest the needed participant sample size, assuming a significance level of p < .05 and power equal to 0.80. For each dependent variable, we conducted power analyses using free a power/sample size calculator offered by the University of British Columbia (https://www.stat.ubc.ca/∼rollin/stats/ssize/n2.html). For both dependent variables, the largest group differences were between the consistent handers in the narrow text condition and the wide text condition. Based on our reported group means and standard deviations, for percent of correct responses 48 participants per cell or 192 total participants are required. For signal sensitivity, 65 participants per cell or 260 participants would be required. Thus, any replication study should have somewhere in the range of 192--260 participants.
Discussion
In this study, consistently handed participants who read a text laid out in a narrow columnar format remembered less information than either participants who read the text from a wide text layout or inconsistently handed participants who read it from a narrow text layout. Our proposed explanation for these results is that inconsistent handers had more resting brain activity in the right cerebral hemisphere regions that are associated with visuospatial attention, particularly the attentional processes in the frontal and parietal regions (the brain’s alerting system) and the frontal and posterior brain regions (the brain’s attentional systems for orienting and executive control). The narrower text columns elicited readers’ leftward attention orienting saccadic eye movements and activated orienting areas of the brain such as the TPJ and executive control in the ACC. The effects of manipulating text column width and eye movement with associated brain activation was stronger for consistent handers who otherwise show less right hemisphere activation because their natural predilection is toward left-hemisphere based encoding that tends to compete for general cognitive resources with right hemisphere retrieval (Craik, 2014). Among consistent handers, wider text columns recruited novel right hemisphere activity that interfered with this predilection toward disruptive encoding activity. A visual of this model can be seen in Figure 2.
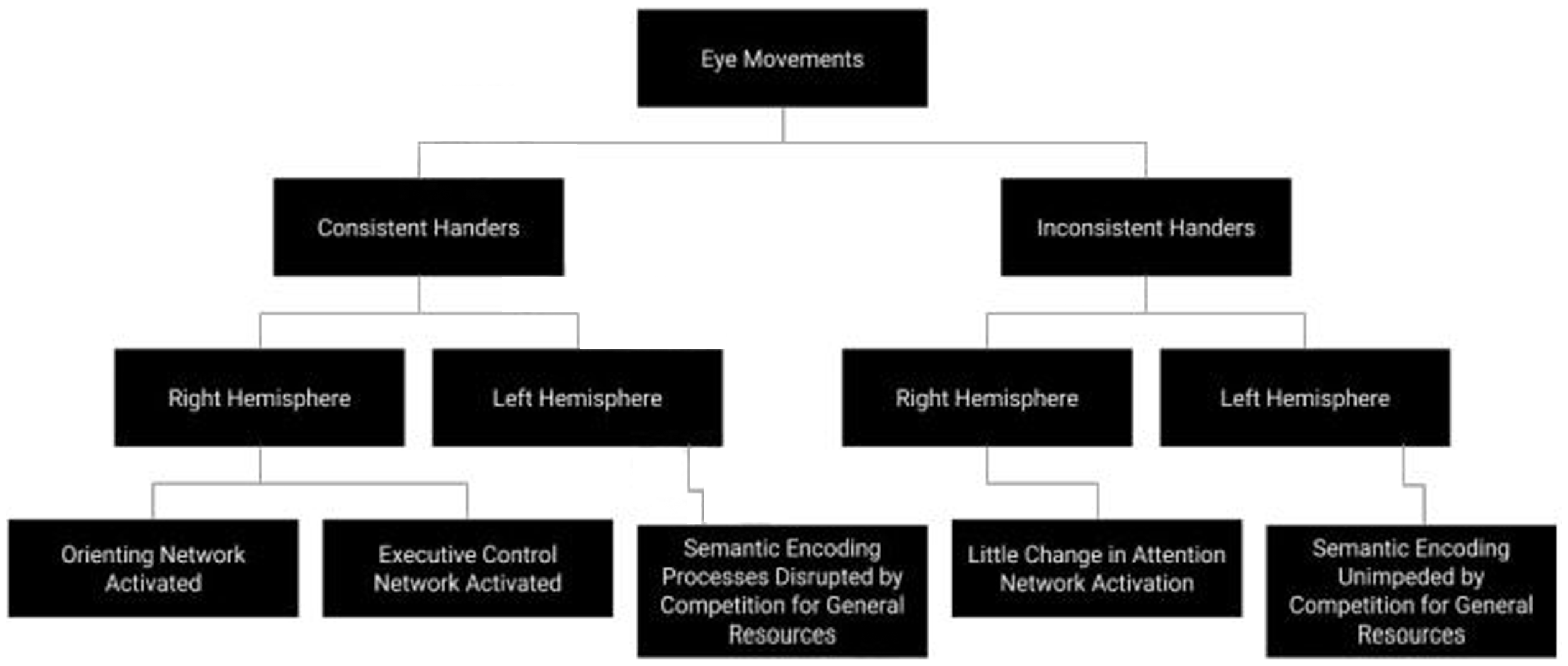
Model of Proposed Mechanism.
These findings and our account of them have several implications. First and foremost, our study suggests that both the HERA model (Habib et al., 2003) and the attentional processing model of brain functioning (e.g., Lyle & Martin, 2010) have roles in explaining the effects of eye movements and handedness consistency on brain activation and text recall. The attentional model may actually help explain the HERA model. While early critics of the attentional model often noted its apparent failure to explain memory for nonverbal information (Fletcher & Henson, 2001; Owen, 2003; Propper & Christman, 1999), Craik (2014) noted that visual attentional disruptions specifically disrupted explicit memory, even for visually presented verbal material. Parker et al. (2018) then found that eye movements enhanced explicit recall of conceptual material. Hence, eye movements induced a relative increase of right hemisphere brain activity during explicit recall that might relate primarily to increased activity in the attentional regions of the right hemisphere, for which a higher resting activity among inconsistent handers presented a relative explicit, but not implicit, memory advantage (Christman & Propper, 2010). This advantage, in turn, accounted for greater functional access to the right hemisphere experienced by inconsistent handers, though it did not imply interhemispheric interaction. Hence accounts of attentional processing, right hemisphere access, and HERA may all be different ways of saying the same thing: explicit recall for visually presented verbal material relies heavily on right hemisphere dominated visuospatial attentional networks.
From an applied cognition perspective, the results of this study highlight the importance of column width on the readers’ eye movements and on associated brain processing during reading. A recent study on persuasion through reading a text passage also employed this research paradigm by which text column width was manipulated (Christman & Sabry, 2009). Participants who read a passage in a narrow versus wide text columns were more persuaded by the text content of the passage. These authors interpreted their finding as reflecting greater left hemisphere activation in the wide column condition, arising from predominantly rightward eye movements while reading. As belief updating and persuasion tend to be handled by the right hemisphere (Christman et al., 2008; Ramachandran & Blakeslee, 1998), this left hemisphere activation worked against updating beliefs. Thus, reading text with narrow columns may have interfered with memory, at least among consistent-handers, even while it may have increased persuasion.
Limitations and Directions for Future Research
Among our study’s most notable limitations was our participant sample size, increasing our risk for failing to detect significant group differences. Our post-hoc power analyses suggested that future replications should have around two hundred participants. A second major limitation is that we could not test our interpretation of hypothesized brain activity, as we did not directly measure brain activity. EEG or fMRI studies might provide supporting or disconfirming evidence for our proposed brain activity hypotheses. Thirdly, the materials used in this study are relatively new in the literature (Prichard & Christman, 2017), and their full psychometric properties for measuring memory are not yet fully understood. However, using these materials, our result was in line with our predictions. Future researchers can use the materials reproduced here and by Prichard and Christman (2017) to replicate or expand upon our results. Also, our study only used verbal stimuli, and future researchers might determine whether a different effect may be obtained using non-verbal visual stimuli, such as an array of shapes arranged in a wide or narrow groupings. Future research should take these findings and their interpretations squarely into the realm of applied cognitive psychology. Following a conceptual replication of these results, it will be crucial for future studies to use more naturalistic materials such as frequently available magazines, newspapers, and cell phones with their skinny paragraphs versus textbooks with their wider columns. Additionally, in the current study we only manipulated the width of text at encoding. It remains an open empirical question as to what would happen if we had manipulated the width of text at retrieval and/or presented the memory assessment questions in different text widths. Other questions to be addressed include: (a) Do people use compensatory strategies when reading from narrow columns, and how widespread are these strategies? (b) What is the temporal course of the text column width effect for inconsistent and consistent handers? and (c) What are text column width and handedness consistency interaction effects on comprehension and persuasion?
Conclusion
In this study, we showed eye-movement effects on certain readers’ (i.e., consistent vs. inconsistent handers) memories of text material. We presented a novel research paradigm and discussed several different theoretical brain activation mechanisms for our findings. These results extend and help further clarify the theoretical debate surrounding eye-movements and memory by lending support to primarily right cerebral hemisphere mediated attentional processing. Finally, the methodology in this study offers an avenue for more applied research on the recall, comprehension, and persuasive effects of text layout on read material.