
Abstract
Introduction
Understanding postural control in older adults is crucial, given the progressive decline in sensory systems with aging, particularly vision, somatosensation, and the vestibular system (Nashner, 1976; Winter, 1995). Among these, deteriorating visual function is a key factor contributing to reduced balance and increased fall risk in older individuals (Seidler et al., 2010). Older adults show delayed saccadic eye movements (Irving et al., 2006) and diminished ability to maintain stable posture (Malcolm et al., 2021), suggesting that changes in visual processing may compromise postural control. The interaction between vision and postural stability has long been a subject of empirical investigation. According to ecological optics theory, postural coordination emerges from real-time interaction with the visual environment, where visual input acts as a primary source of information for movement regulation (Gibson, 2014; Wade & Jones, 1997). Experiments such as the “swinging room” (Lee & Lishman, 1975; Stoffregen, Hove, et al., 2006) and visual frequency modulation studies (Ravaioli et al., 2005) have demonstrated that visual stimuli dynamically influence postural sway. Furthermore, blocking or degrading visual input has been shown to increase postural instability (Paulus et al., 1984), reinforcing the role of vision in stabilizing posture.
Recent research has examined how complex visual tasks affect balance, particularly in older adults. Tasks involving cognitive load, such as the Simon-Flanker paradigm, have been associated with prolonged postural correction time in older adults (Kwag et al., 2024). Likewise, optical flow manipulations have shown that visual stimulus direction significantly alters center of pressure (COP) behavior in older adults at fall risk (Phu et al., 2023). These findings highlight that postural control is not only a motor function but also closely intertwined with visual-cognitive processing. It is well established that older adults rely more heavily on visual information to maintain postural stability (Paulus et al., 1984; Phu et al., 2023). However, most previous studies have quantified this visual dependence using amplitude- or frequency-based measures of postural sway. While these measures are useful for characterizing the magnitude and oscillatory properties of sway, they are limited in their ability to capture the organizational patterns, adaptability, and qualitative aspects of postural control.
To quantitatively evaluate these interactions, researchers have increasingly turned to entropy-based measures. Entropy reflects the degree of irregularity or unpredictability in a physiological signal and is widely used to assess system complexity. Sample entropy (SampEn), in particular, quantifies the unpredictability of time-series data such as COP trajectories and has proven to be a reliable indicator of postural control complexity (Dinkel et al., 2017; Lakhani & Mansfield, 2015; Ramdani et al., 2009; Richman & Moorman, 2000; Yamagata et al., 2017). Higher SampEn values suggest more irregular, less predictable sway often interpreted as a more automatic and adaptive control state (Fischer et al., 2023; Kędziorek & Błażkiewicz, 2020).
SampEn reflects the complexity of postural control rather than task difficulty itself. This metric captures how attention and control strategies influence the organization of postural sway. Under constrained conditions (e.g., eyes closed or unstable surfaces), reduced SampEn indicates a simplified, less complex control mode dominated by conscious stabilization (Donker et al., 2007; Franco et al., 2018). In contrast, higher SampEn values in dual-task or external-focus conditions reflect greater system complexity and more automatic, adaptive regulation (Becker & Hung, 2020; Potvin-Desrochers et al., 2017; Rhea et al., 2019; Richer & Lajoie, 2020; Wulf, 2013). That is, lower SampEn reflects a more constrained and less complex control mode dominated by conscious stabilization, whereas higher SampEn indicates greater system complexity and a higher degree of automatic control. However, increased entropy observed in contexts such as aging or seated instability may reflect maladaptive noise or loss of structured variability rather than functional complexity (Borg & Laxåback, 2010; Rhea et al., 2011; Roerdink et al., 2011). In older adults, sensory reweighting and increased reliance on visual input may elevate complexity in postural control, which can either represent compensatory adaptation or degraded regulation when dominated by random fluctuations (Liu et al., 2024).
Parallel to COP complexity, gaze behavior can also be quantified using entropy measures. Gaze entropy captures the variability and predictability of eye movement patterns and serves as an index of visual search strategy and cognitive load (Bhavsar et al., 2017; Ebeid et al., 2018; Ryu et al., 2016). Higher gaze entropy reflects wider visual exploration, while lower entropy suggests focal attention on specific areas (Jungk et al., 1999; Song & Han, 2024). In occupational and sports contexts, experts tend to exhibit lower gaze entropy due to more structured scanning behavior (Bhavsar et al., 2017), whereas novices show higher gaze entropy, reflecting broader and less efficient visual sampling.
Despite the utility of both SampEn and gaze entropy in understanding movement and attention, few studies have explored their interactive relationship, particularly under gaze-controlled conditions. Gaze behavior plays a central role in stabilizing posture, especially in environments that require dynamic visual scanning (Balaban & Thayer, 2001). However, whether manipulating gaze entropy directly alters postural complexity (i.e., COP, SampEn), and whether this effect differs by age, remains unclear.
Beyond quantifying visual dependence through sway magnitude or frequency, examining the dynamic coupling between gaze behavior and postural fluctuations offers a novel perspective for understanding visuomotor integration in aging. According to previous studies, older adults exhibit stronger sway–gaze coupling and longer fixation durations than younger adults, particularly under attentionally demanding conditions (Walsh, 2021), indicating age-related changes in how visual and postural control systems interact. In addition, reduced coherence and increased cross-entropy between visual motion and body sway have been reported in older adults (Sotirakis et al., 2016), suggesting decreased efficiency in integrating visual information for postural regulation. Building on these findings, the present study aims to explore how gaze entropy representing the variability and distribution of visual exploration relates to the SampEn of COP in both younger and older adults. This approach provides a novel framework to characterize the complexity and adaptability of visuomotor coordination beyond traditional sway-based indicators, thereby contributing to a more comprehensive understanding of age-related changes in postural control.
Therefore, this study aims to investigate how changes in gaze behavior operationalized as gaze entropy affect the irregularity of postural control (measured via COP, SampEn) in both younger and older adults. By manipulating gaze movement conditions, this study explores whether increased visual complexity translates to changes in postural complexity, and whether these effects differ by age. This approach not only expands upon previous research that passively measured vision–posture coupling, but also introduces an active, task-driven manipulation of gaze to examine its causal role in balance regulation.
Hypotheses
(1) Older adults will exhibit higher COP SampEn than younger adults under all gaze conditions, reflecting age-related changes in postural control. (2) Gaze entropy manipulations will influence COP SampEn, with increased gaze entropy (reflecting greater visual exploration complexity) leading to increased postural irregularity. (3) The relationship between gaze entropy and COP SampEn will be more pronounced in older adults, reflecting their greater reliance on visual input for postural regulation.
Methods
Participants
A total of 30 participants were recruited, consisting of 15 younger adults (9 males, 6 females; mean age: 31.33 ± 5.74 years) and 15 older adults (6 males, 9 females; mean age: 66.53 ± 1.77 years). An a priori power analysis using G*Power 3.1 indicated that a total sample size of 34 would be required to achieve a power of 0.80 for detecting medium-sized effects (f = 0.25) in a 2 × 2 mixed-design ANOVA. Due to recruitment limitations, the final sample consisted of 30 participants, yielding an estimated power of 0.75. Despite this slight reduction in power, effect sizes are reported alongside p-values to provide additional information on the magnitude and robustness of the observed effects, allowing for cautious interpretation.
The younger adult group comprised healthy individuals under the age of 40, primarily university students and graduate students. The older adult group consisted of healthy individuals aged 65 or older residing in the community, including those who had retired from office work or were engaged in freelance or light production activities. All participants possessed sufficient physical fitness to perform the single-leg stance task without difficulty and had corrected vision adequate for wearing and utilizing the eye-tracking device without any issues.
Apparatus
Eye-Movement Tracking Device
To measure eye movements, the Eye Tracker Dikablis Glasses 3 (Ergoneers, Germany) was used. This device can track both vertical and horizontal eye movements, allowing for real-time analysis of participants’ gaze shifts. In this study, binocular tracking was performed, and data were collected at a sampling frequency of 60 Hz. Additionally, an external webcam was connected to simultaneously record participants’ single-leg stance movements and gaze behavior. The collected video data were analyzed using D-Lab 3.0 (Ergoneers, Germany). D-Lab 3.0 automatically corrected minor artifacts such as eye blinks and pupil instability. As a result, the gaze trajectories of both eyes were successfully obtained for more than 97% of the total recording duration. During the preprocessing stage, calibration and pupil tracking accuracy were carefully verified, and no substantial data loss or abnormal signal was observed during the experimental trials.
Force Plate
To measure the COP, two BMS400600 force plates (AMTI, USA) were used. These devices precisely measure the ground reaction forces (GRF) in vertical, anterior-posterior, and medial-lateral directions. Data were collected at a sampling frequency of 100 Hz using two synchronized force plates.
Procedures
Experimental Task
The experimental task was developed using MATLAB 2024a, referencing previous studies to align with the research objectives.
The eye-tracking task required participants to follow a red target circle among ten circles presented on a screen using only their gaze (Alvarez & Franconeri, 2007; Pylyshyn & Storm, 1988). Following previous studies, the non-target circles were initially red but quickly changed to blue after task onset, requiring participants to actively track the designated red target among moving distractors. This design ensured that the target’s movement was sufficiently complex while preventing participants from anticipating its trajectory, thereby engaging visual attention and tracking mechanisms. The size of all circles was set within a visual angle of 3° (Thomas et al., 2016), and the red target moved at a speed of 60 pixels per second to facilitate smooth pursuit eye movement. To minimize head rotation and measure postural sway, the red target was restricted to movements within a visual angle of 15° (Glasauer et al., 2005; Stoffregen, Bardy, et al., 2006, 2007). Although the primary purpose of this task was to manipulate visual attention and gaze behavior, it was also expected to subtly influence postural control by engaging the participants’ visual tracking system during standing. The height of the visual stimulus was standardized for all participants, with no individual adjustments. Participants were instructed to maintain an upright posture with their heads held high and to minimize head movement throughout the task.
The postural control task required participants to wear an eye-tracking device while performing a single-leg stance for 20 seconds, continuously focusing on a red target displayed on a 120-inch screen positioned 2 meters in front of them (Figure 1). The single-leg stance task was chosen based on findings by Thomas et al. (2016), which indicated that a two-leg stance did not yield significant differences in postural control between younger adults and older adults. Experimental setup
For the single-leg stance, participants initially stood barefoot (or wearing thin socks) on a force plate with both feet evenly supported. They crossed their arms over their chest, and upon the presentation of the eye-tracking task, they lifted their preferred foot to maintain balance (Song et al., 2022). Several studies have recommended a minimum trial duration of 30–60s for reliable COP-based measurements (Jonsson et al., 2004; van der Kooij et al., 2011). In older adults, COP measurements obtained during 30s double-leg stance tasks have been reported to provide adequate reliability (Moghadam et al., 2011; Qiu & Xiong, 2015). However, in the present study, participants were not only older adults but also required to perform single-leg stance tasks, for which prolonged trials may pose physical and ethical concerns. Pilot testing indicated that maintaining balance beyond 30 s frequently led to loss of stability or excessive sway, making it difficult to distinguish whether such fluctuations were due to fatigue or gaze manipulation. Therefore, each participant performed the same task twice per session, resulting in a total of four trials. To ensure data consistency, only trials in which participants maintained full balance were included in the analysis; any trial involving loss of balance or compensatory movement (e.g., hopping or stepping) was repeated immediately after a brief rest period. Such cases occurred in fewer than two instances overall, suggesting negligible influence on data reliability.
Experimental Procedure
After signing the informed consent form, participants received an explanation of the experimental procedure from the researcher. The experiment began with the participant sitting on a chair positioned behind the force plate while wearing the eye-tracking device. Once the device was fitted, participants performed a calibration process by fixating on four designated points without moving their heads. The researcher ensured that the participant’s gaze aligned with the crosshairs displayed on the monitor.
Following calibration, participants stood barefoot (or wearing thin socks) on the force plates in a bipedal stance, and the force plate measurement commenced. They were then instructed to locate the red target among the ten circles on the screen. Once they identified the target, they verbally confirmed their selection and initiated the single-leg stance at their own pace. The red target simultaneously changed to the same blue color as the other circles and began to move as the participant lifted their foot. All participants lifted their foot within 3 seconds of identifying the target. Gaze recording via the eye-tracking device began at the moment of foot lift and continued for the 20-s duration of the task. Participants were instructed to raise their non-supporting foot to approximately knee height of the opposite leg, although no specific height was strictly imposed.
The task consisted of two gaze tracking conditions differing in gaze entropy levels:
Narrow gaze tracking, where the red target moved regularly within a small range, following a predictable sinusoidal path within a visual angle of ±5°, and Wide gaze tracking, where the target moved irregularly over a larger range, following a pseudo-random path up to ±15°, simulating irregular visual exploration. Each condition was repeated twice to prevent learning effects, and task order was counterbalanced across participants. To avoid bias related to foot dominance, the dominant leg was not specifically identified, and participants were allowed to use whichever leg they felt more comfortable supporting. All trials were conducted using the same leg. To mitigate fatigue effects, a 1-min rest period was provided between trials. All participants successfully completed the experiment without falls.
Data Analysis
Gaze Entropy
To calculate gaze entropy, the Shannon entropy method was used, as shown in equation (1). Gaze entropy was computed based on the fixation frequency of more than 0.1 seconds on an area of interest (AOI) (Shiferaw et al., 2019; Song & Han, 2024).
The AOI was defined by dividing the screen used in the eye-tracking task into a 9-grid pattern, ensuring uniform segmentation (Cui et al., 2024).

In the entropy equation, H represents the entropy value, N denotes the number of possible states, and Pᵢ refers to the probability of state i occurring. Thus, gaze entropy was derived based on the probability of fixations within each AOI region.
COP & Sample Entropy
The data collection period was defined as starting the moment the participant lifted one foot—specifically, at the moment when the vertical ground reaction force (Fz) of the lifted foot dropped below 10 N. This threshold was applied consistently across all participants. After collecting all raw data, a 4th-order double-pass Butterworth filter was applied with a cut-off frequency of 10 Hz for filtering (Borg & Laxåback, 2010).
Additionally, to ensure consistency in analysis, the first 5 seconds of the single-leg stance, where the most significant postural sway occurs (Jonsson et al., 2004), and the last 5 seconds before the trial ended were excluded. As a result, a total of 10 seconds of data from each trial was used for analysis.
SampEn was calculated using COP data. The calculation assumes that the recorded COP trajectory consists of N data points, sampled at a constant sampling frequency (Fs). The total signal duration (T) is defined as T = N/Fs, where T is measured in seconds. The values used for the SampEn parameters were m = 2 and r = 0.2 × SD, in accordance with previous postural control studies (Borg & Laxåback, 2010; Ramdani et al., 2009).
At each time n/Fs, the COP coordinates were extracted, and equation (2) represents the center of pressure displacement in the medial-lateral (ML) and anterior-posterior (AP) directions, respectively (Quijoux et al., 2021).

To calculate SampEn from the collected COP data, equation (3) was applied.
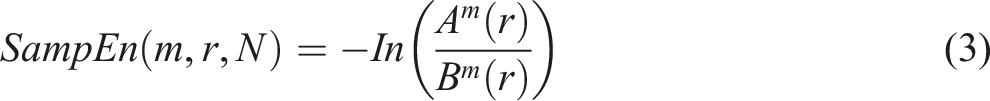
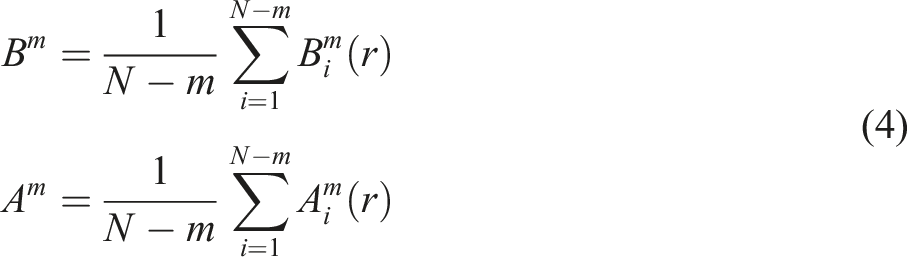
SampEn is used to quantify the variability of COP over time and is defined as the negative natural logarithm (-ln) of the probability that a data sequence of a certain length remains similar when an additional data point is included (Estrada et al., 2017; Richman & Moorman, 2000).
In this study, a custom MATLAB script (MathWorks, USA) was used to calculate SampEn in both the AP and ML directions (Borg & Laxåback, 2010). Here, m represents the length of the data sequence being compared, r is the similarity threshold, and N is the total data length.
After filtering the COP data, SampEn was calculated. Additionally, the same analysis was performed on the unfiltered COP data to assess the impact of filtering. The results showed absolute differences depending on the filtering process; however, the overall trends across conditions remained consistent. Although COP signals were not normalized to unit variance, the tolerance parameter (r) was defined relative to each signal’s standard deviation (SD) to compensate for amplitude differences across conditions.
Statistics
Data processing was conducted using engineering software (MATLAB, 2024a, MathWorks, USA) and Microsoft Excel (MS Excel, Microsoft, USA), while statistical analysis was performed using SPSS 26.0.
To assess the relative reliability of the COP and Sample Entropy measures across repeated trials, intraclass correlation coefficients (ICC (2, 2)) for the mean of multiple trials were calculated using a two-way random-effects model with absolute agreement (Pineda et al., 2020). Prior to the main analysis, the Shapiro–Wilk test was used to assess the normality of all dependent variables. A two-way mixed-design ANOVA was then conducted to examine the effects of group (younger vs. older) and gaze-tracking task (narrow vs. wide gaze tracking) on gaze entropy, COP, and SampEn. If a significant main effect was found for any factor, a Bonferroni post hoc test was performed. The statistical significance level was set at p < .05.
Results
Gaze Entropy
The analysis of gaze entropy revealed no significant differences between groups [F (1, 28) = .202, p = .657, A two-way mixed-design ANOVA Results for Gaze Entropy Measures Across Groups and Tasks. The error bar is the standard deviation. * indicate a statistically significant difference (p < .05)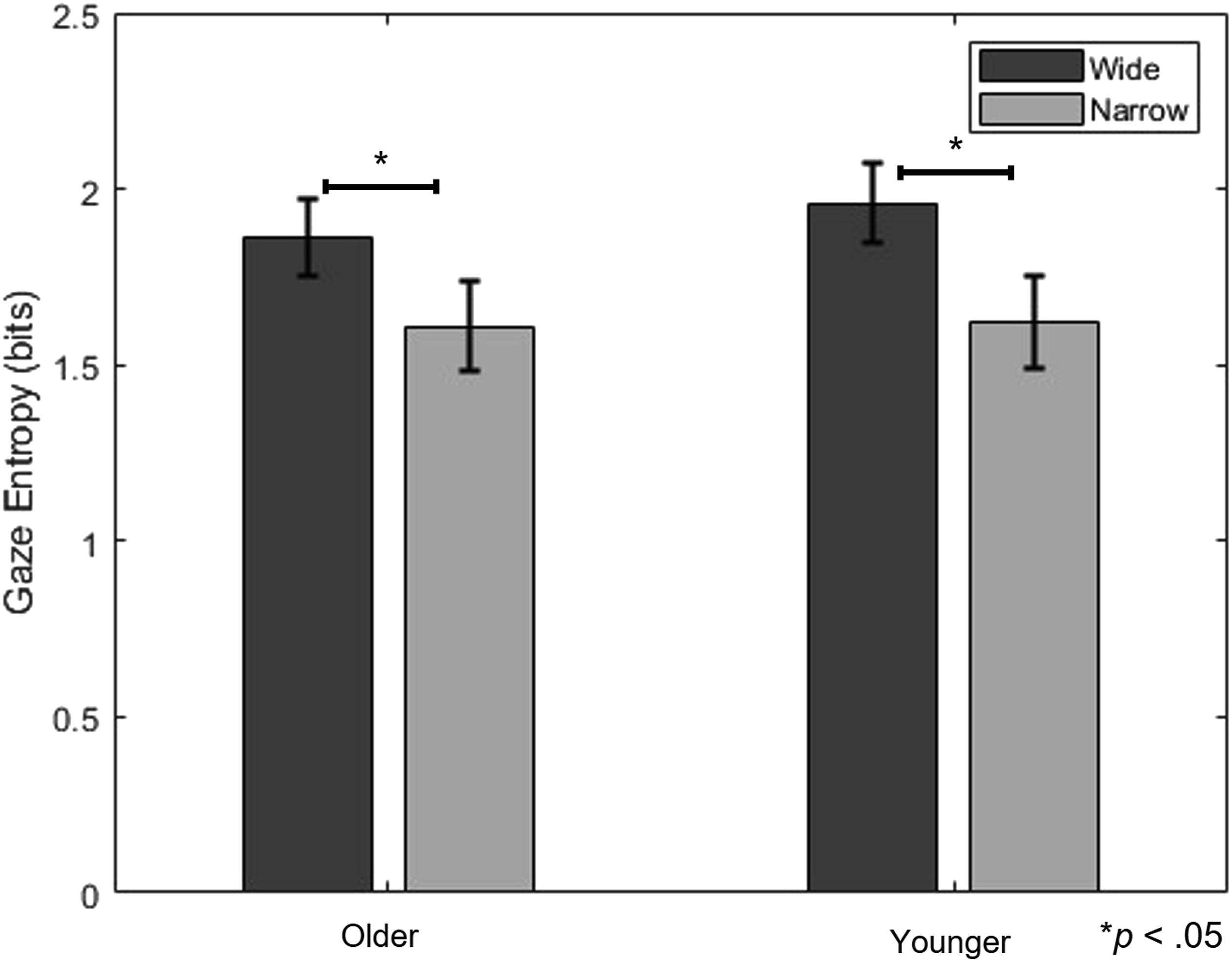
Cop
A Two-Way Mixed-Design ANOVA Results for Gaze and COP Measures Across Groups and Tasks
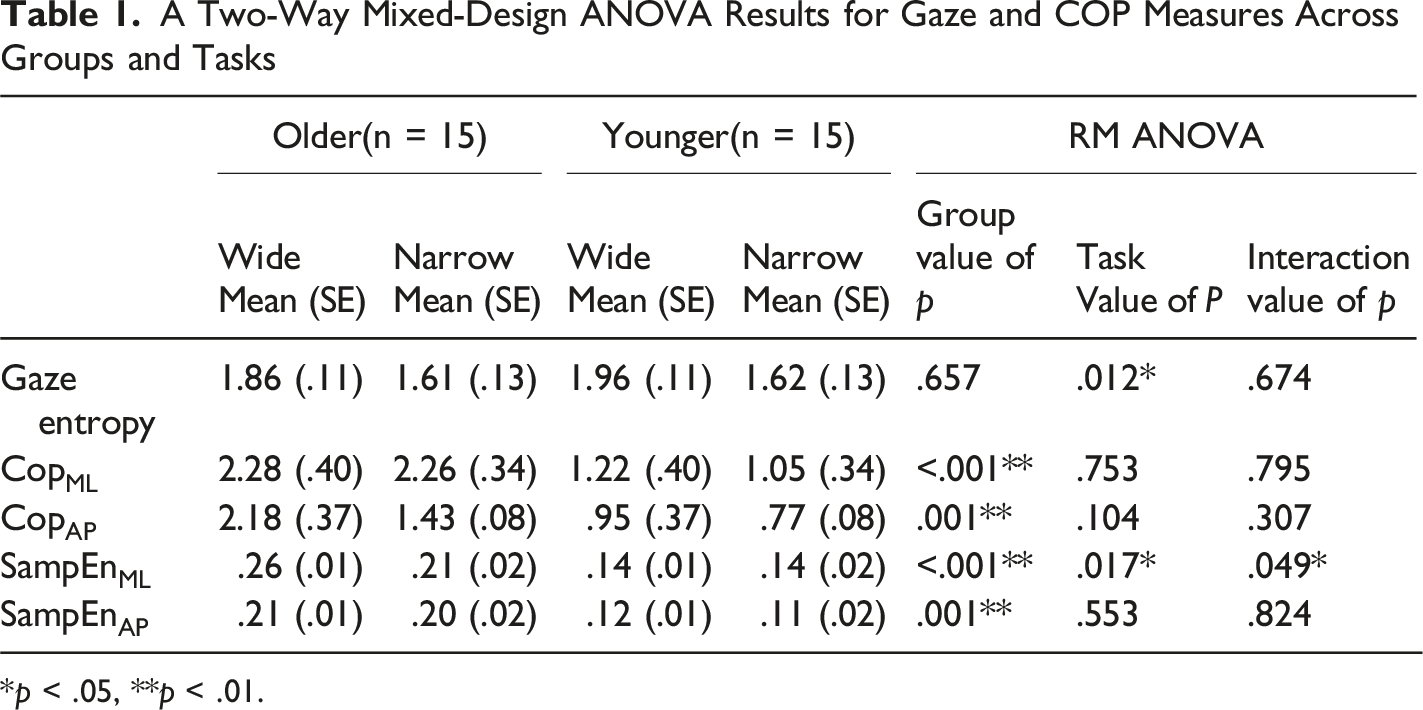
*p < .05, **p < .01.
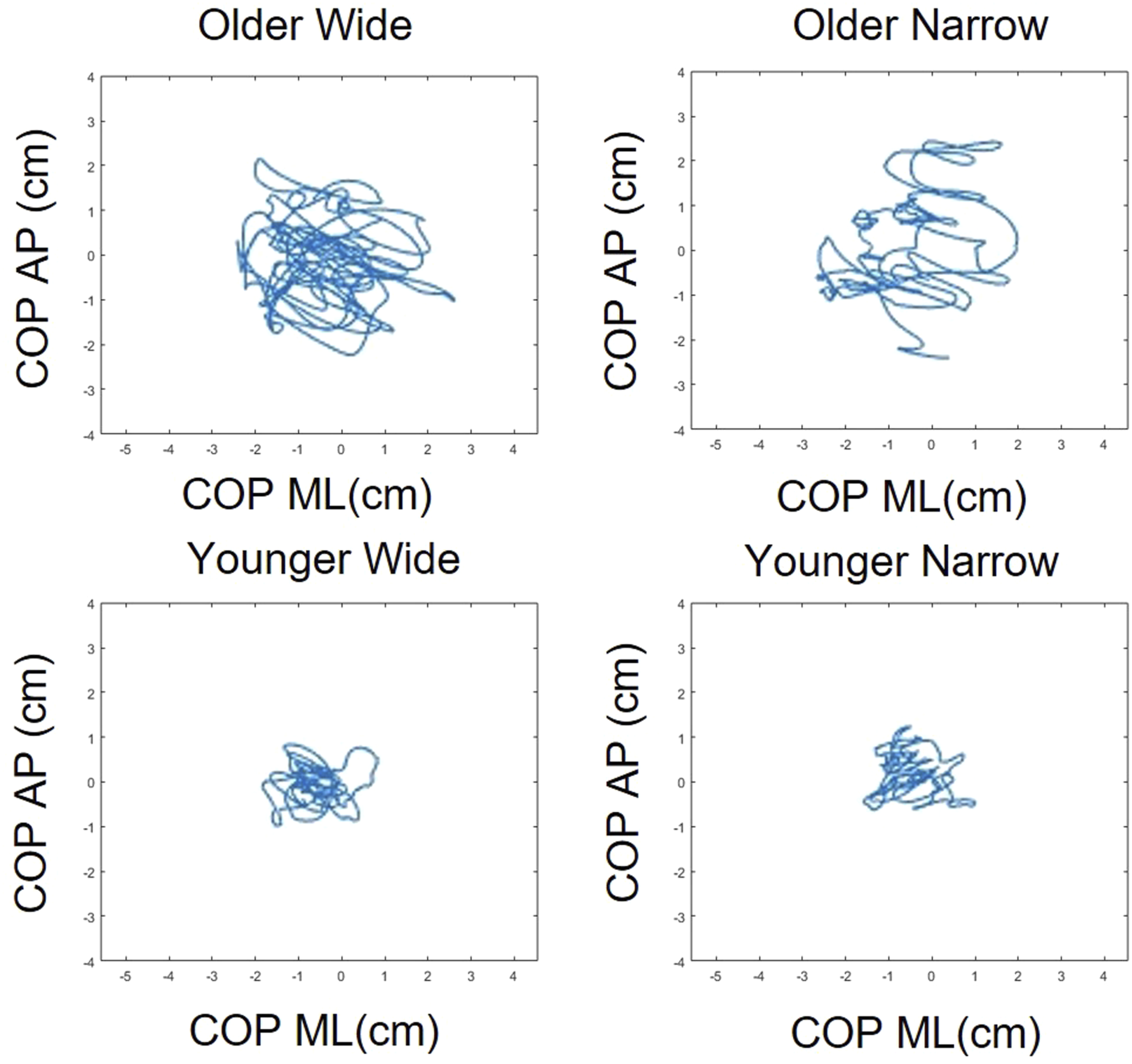
Individual COP results for younger and older adults
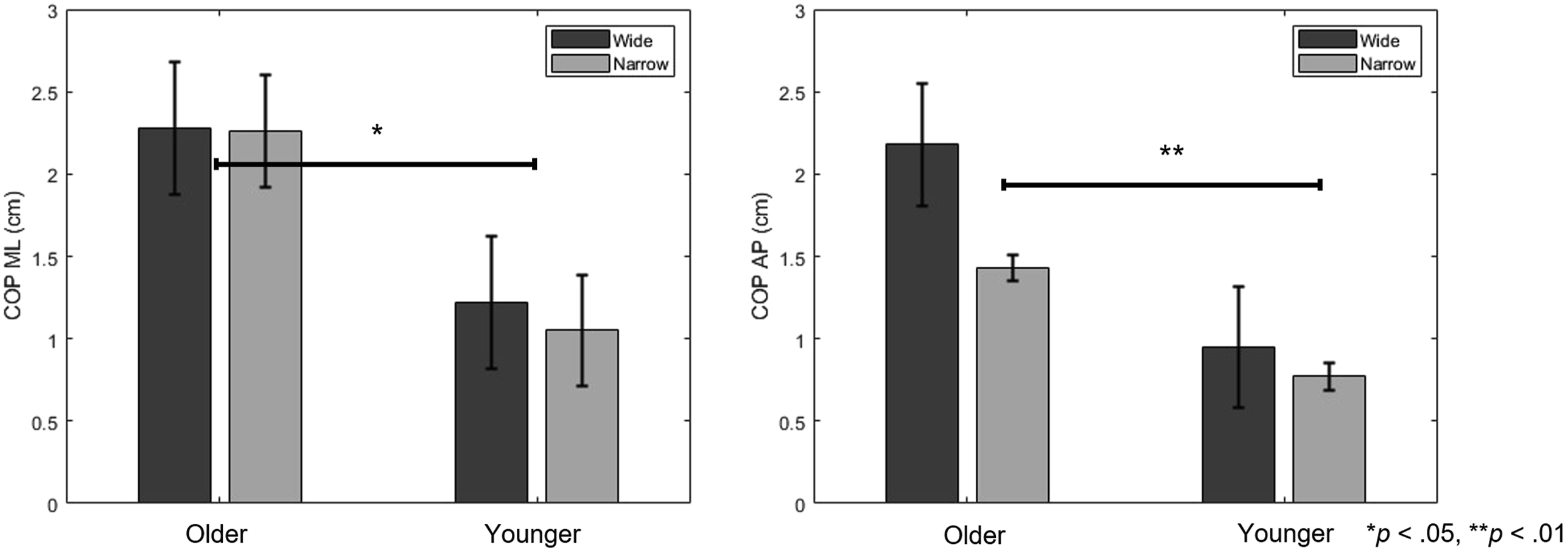
A two-way mixed-design ANOVA Results for COP measures Across Groups and Tasks. The error bar is the standard deviation. * indicate a statistically significant difference (p < .05). ** indicate a statistically significant difference (p < .01)
Sample Entropy
The ICC results are as follows. Sample Entropy showed ICCs of .812 (SampEn-ML) and .652 (SampEn-AP) for the wide task, and .704 (SampEn-ML) and .765 (SampEn-AP) for the narrow task, indicating measurement reliability. Older adults exhibited significantly higher SampEn values than younger adults in both ML [F(1, 28) = 31.462, p < .001, A two-way mixed-design ANOVA Results for SampEn Measures Across Groups and Tasks. The error bar is the standard deviation. * indicate a statistically significant difference (p < .05). ** indicate a statistically significant difference (p < .01)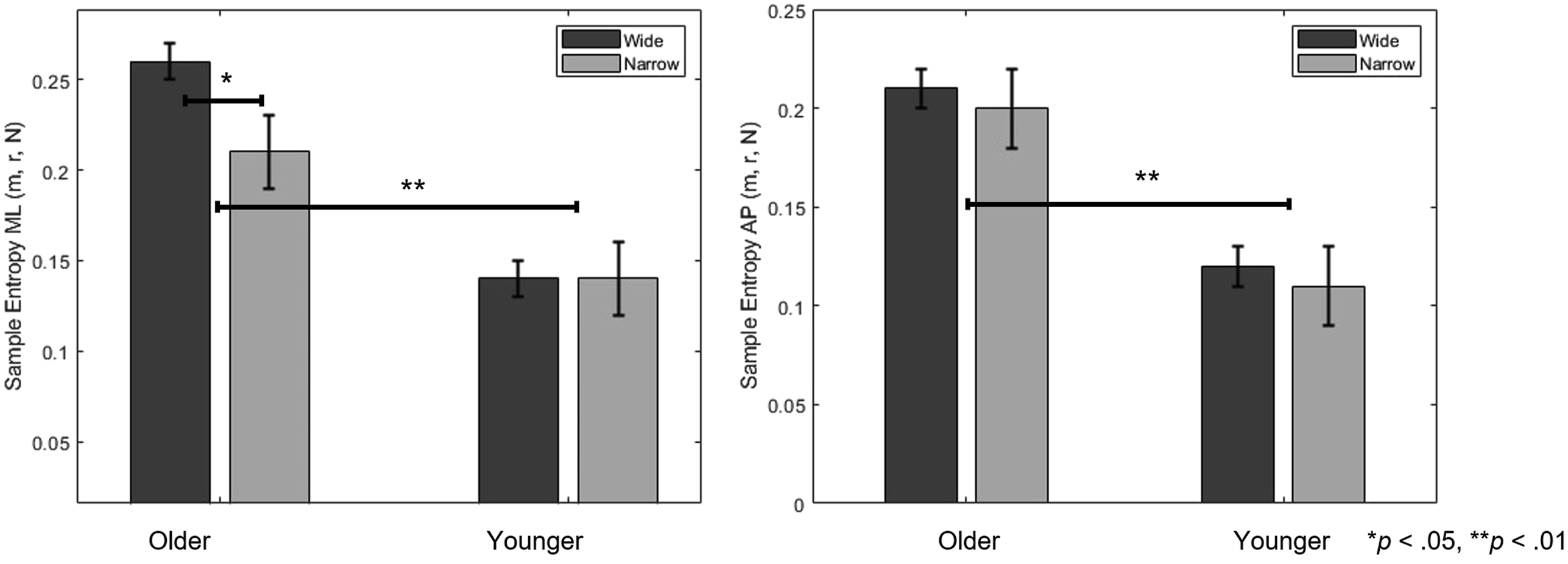
Discussion
This study aimed to examine how task-driven manipulation of gaze behavior, quantified by gaze entropy, influences the complexity of postural control measured by SampEn of COP in younger and older adults during a single-leg stance. This approach expands on previous studies that treated visual input as a static factor, by introducing controlled variability in visual search behavior. Our findings offer three key insights related to age-related postural control, gaze behavior, and their interaction.
First, gaze entropy significantly differed by task condition but not by age group, indicating that our manipulation (narrow vs. wide gaze tracking) effectively altered visual search complexity. This supports the notion that visual exploration can be experimentally modulated (Bhavsar et al., 2017; Foulsham, 2015) and that both younger and older adults retained the cognitive flexibility to adapt to this manipulation under controlled conditions. Contrary to assumptions that aging leads to consistently reduced gaze complexity (Jungk et al., 1999; Land, 2006), older adults in this study exhibited gaze entropy levels comparable to those of the younger adult group, suggesting preserved visual adaptability when task difficulty is systematically managed.
Second, postural measures both COP displacement and SampEn were significantly higher in the older adult group, consistent with prior research showing greater sway and control irregularity in aging populations (Borg & Laxåback, 2010; Jonsson et al., 2004). SampEn, often interpreted as a measure of system irregularity or complexity (Richman & Moorman, 2000), was elevated in older adults regardless of gaze task. This may reflect compensatory neuromuscular strategies or reduced sensory integration, rather than simple balance deterioration. Previous research supports this view, showing that older adults exhibit increased irregularity in postural sway particularly under attention-demanding conditions (Pellecchia, 2003; Shumway-Cook & Woollacott, 2000). Previous studies have shown that older adults’ ability to increase postural sway complexity during dual-tasking may be constrained by their functional balance capacity (Kal et al., 2022). From this perspective, the increased irregularity observed in the present study can be interpreted not as simple postural deterioration, but as a functional adaptation reflecting the reallocation of attentional resources to the secondary task and a shift toward automatic control.
Third, a significant interaction was observed in SampEn in the ML direction, where older adults showed greater SampEn under wide gaze entropy conditions. In contrast, younger adults’ SampEn remained stable across conditions. This suggests that older adults dynamically adjust postural control complexity in response to increased visual task demands possibly reflecting increased reliance on visual input for balance (Balaban & Thayer, 2001; Ravaioli et al., 2005). This is conceptually aligned with the U-shaped nonlinear interaction model (Lacour et al., 2008), where moderate task demands can either hinder or enhance postural automaticity depending on cognitive load. In our case, the wide gaze condition may have promoted external focus and automatic stabilization, leading to increased entropy (McNevin & Wulf, 2002; Wulf et al., 2004). However, SampEn does not linearly map to balance quality. While higher SampEn has been linked to more adaptive control in some contexts (Becker & Hung, 2020; Kędziorek & Błażkiewicz, 2020), in others, it may signify instability or system noise (Donker et al., 2007; Roerdink et al., 2011). Hence, our results should not be interpreted as “better” or “worse” control based on entropy alone, but rather as reflecting different control strategies a key point raised by previous studies (Ramdani et al., 2009; Guerraz & Bronstein, 2008).
Additionally, the lack of interaction in AP direction suggests directional specificity in how visual exploration affects balance. While ML control is often interpreted as being more dependent on active sensory integration and cognitive attention than AP control, it is important to note that in single-leg stance, ML stability is mechanically more critical due to the narrower base of support. Therefore, the observed directional specificity may also reflect adaptive changes related to task mechanics rather than solely increased reliance on sensory or attentional processes (Haid & Federolf, 2018; Hansen et al., 2017).
Motor behavior and the use of visual information occur in complex environments and inevitably involve dynamic interactions with both internal and external factors (Zentgraf et al., 2011). Understanding these complex interactions and developing cognitive abilities to regulate perception-action processes allow individuals to more accurately interpret the outcomes of their actions. Previous studies have also highlighted the need to explore how such complexity influences the optimization of human behavior (Renaud et al., 2003, 2007). Increases in the complexity of gaze behavior can influence sensorimotor processes related to postural control, and some studies have reported that visual stimuli affect postural regulation (Giveans et al., 2011; Jeka et al., 2006; Stoffregen, Bardy, et al., 2006). However, other studies have failed to find clear evidence that eye movements directly affect postural control (Haworth et al., 2014), possibly because the experimental tasks did not provide sufficient challenge for participants. In light of this, the present study employed a more challenging single-leg stance task instead of a double-leg stance. Single-leg stance has been used as an indicator to identify groups with impaired balance (Drusini et al., 2002; Vellas et al., 1997) and to assess fall risk in older adults (Hurvitz et al., 2000).
The results of the present study suggest that, in the older adult group, increases in gaze entropy were generally associated with higher COP and SampEn values, which may indicate greater postural irregularity. Importantly, these findings do not necessarily reflect simple balance deterioration but could reflect a functional adaptation, whereby attentional resources are reallocated to visual exploration tasks and automatic postural control strategies may be engaged. In particular, the observed increases in postural irregularity in both the ML and AP directions during the single-leg stance could reflect compensatory neuromuscular control and altered sensory integration. These findings highlight a potential coupling between gaze behavior and postural control and provide insight into how visuomotor integration strategies may be adjusted with aging. That is, older adults may not simply exhibit increased postural variability but may adopt adaptive balance strategies that utilize visual information. Taken together, these findings indicate that gaze entropy manipulation can be a useful approach to probe sensorimotor integration, particularly in older adults. Moreover, using entropy-based metrics such as SampEn may allow for detecting subtle, strategy-level adaptations that might not be apparent in linear COP measures.
Despite promising results, this study had some limitations. A limitation of this study is that only the central 10-s segment of each trial was used to compute COP and SampEn. Although this approach minimized artifacts and reduced the burden for older adults, short epochs can compromise the reliability of COP measures. In the present study, most ICC values were above 0.7, indicating good reliability; however, SampEn in the anteroposterior direction (SampEn-AP) showed relatively lower reliability, warranting caution in interpreting the results. Future studies should consider longer recording durations or additional trials to enhance the reliability of postural control metrics. Additionally, in this study gaze and force-plate signals were synchronized using an external camera trigger rather than a dedicated hardware integration. However, because our primary aim was to compare sample entropy of COP across gaze-entropy conditions using fixed-length analysis windows, precise sample-by-sample hardware synchronization was not required to test our main hypotheses. The present study included 30 participants (15 per group), slightly fewer than the 34 initially recruited. The original sample size was based on previous studies and estimated to provide sufficient statistical power (≥0.8) for detecting interaction effects; however, due to attrition, the final sample was slightly smaller, which may have slightly reduced the achieved power. However, given the complex nature of entropy-based metrics and the slight reduction in analyzed participants, a formal power analysis based on actual effect sizes and inter-trial correlations is warranted in future studies to confirm the adequacy of the sample size. In addition, we used fixed SampEn parameters (m = 2, r = 0.2 × SD); future work should explore multiscale entropy or complementary complexity metrics to validate findings (Costa et al., 2002). Further studies should also incorporate measures of gaze-posture temporal coupling, muscle activation, or balance success rates to determine whether increased entropy reflects adaptive control or emerging instability. Such integrative analysis will clarify whether gaze-modulated postural complexity represents a compensatory shift or a marker of control deterioration in aging.
Footnotes
Ethical Considerations
All procedures were approved by the Research Ethics Committee of Jeonbuk National University (JBNU 2024-07-028-001) and conducted in accordance with the Declaration of Helsinki.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the [Republic of Korea and the National Research Foundation of Korea] under Grant [NRF-2024S1A5B5A16022239].
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.