
Abstract
Both innate superiority and negative frequency-dependent advantage (NFDA) hypotheses have been proposed to explain the left-hander advantage in interactive sports such as fencing. However, empirical evidence on the specific technical and tactical patterns potentially underlying this advantage remains limited. Therefore, this study analyzed match performance of elite female épéeists at the 2017–2019 World Championships using notational analysis. We examined the prevalence and winning rates of left- and right-handed fencers across tournament stages, as well as the distributions of performed and scored actions in terms of technique, target area, and piste zone. Results revealed that left-handers were consistently overrepresented relative to population prevalence, and their winning rates increased up to the semifinals (80%) but matched right-handers in the finals (50%). Technique use was highly consistent across all handedness matchups, with attacks being the most frequent action. While the inner upper body remained the primary target overall, asymmetrical matchups (left-vs.-right and right-vs.-left) were associated with a shift toward the outer upper body and lower body; this pattern was attenuated in left-handers facing left-handers. Crucially, left-handers consistently demonstrated a more invasive spatial strategy across all matchups, showing a greater tendency to operate and score in the opponent’s middle zone. Taken together, the left-hander’s advantage in elite female épée appears to be underpinned by an interplay between context-dependent target adaptation, aligning with the NFDA account, and a generalized proactive spatial exploitation, as proposed by the innate superiority account.
Introduction
In the general population, the prevalence of left-handedness is estimated at around 10–13%, with some cultural variations (Gilbert & Wysocki, 1992; McManus et al., 2010; Papadatou-Pastou et al., 2020). More specifically, the best overall estimate is 10.6%, ranging from 9.3% under the most stringent criterion to 18.1% under the most lenient criterion (Papadatou-Pastou et al., 2020). Notably, left-handed athletes are often overrepresented at the elite level in certain interactive sports, where the action of one directly affects the action of the opponent (Grouios et al., 2000; Hagemann, 2009; Harris, 2010; Loffing, 2017; Loffing & Hagemann, 2012, 2016; Loffing & Schorer, 2021; Raymond et al., 1996; Simon et al., 2025).
Fencing provides a particularly striking example. The terms left-handed and right-handed refer specifically to weapon-hand laterality, namely the hand used to hold the weapon during competition, rather than the general handedness in everyday activities. Historical surveys of elite épée fencers consistently report an overrepresentation of left-handers. For example, among male champions from 1979 to 1993, 24% were left-handed, rising to 38% among the top eight finishers (Azémar & Stein, 1994; cited in Raymond et al., 1996). At the 1996 Olympics Games, the proportion of left-handers increased from 26% in the opening round to 44% in the final (Azémar, 2003). This pattern persisted in datasets spanning 2007–2016 (Loffing & Schorer, 2021). More recently, analysis of a larger dataset covering 2004–2023 confirmed the overrepresentation of left-handers in both épée and foil, particularly among higher-ranked fencers (Simon et al., 2025).
Why do left-handed fencers appear to have a competitive advantage? This question has long been debated in terms of nature and nurture explanations (Grouios, 2004; Harris, 2010; Loffing & Hagemann, 2012, 2016; Simon et al., 2025). The nature hypothesis proposes that left-handers possess innate superiority in aspects such as reaction time, motor control, attention, or interhemispheric transfer speed. For example, left-handed fencers showed faster finger and hand movements in peg-moving or finger-tapping tasks (Annett & Manning, 1989; Kilshaw & Annett, 1983; Nalcaci et al., 2001), as well as shorter response times in fencing-specific target-hitting tasks (Witkowski, Tomczak, Łuczak, et al., 2020), compared to right-handers. However, reaction time, response time or accuracy in fencing-specific actions, such as the lunge, has not been found associated with competitive success (Ketlinski & Pickens, 1973; Singer, 1968). An attentional advantage has also been suggested. Left-handed fencers exhibited faster reaction times in unattended conditions of the Posner task (Bisiacchi et al., 1985), and showed earlier visually evoked potentials (Taddei et al., 1991). Nevertheless, these claims largely stem from early psychomotor research with small sample sizes (e.g., Bisiacchi et al., 1985; Taddei et al., 1991), limiting their generalizability. Moreover, if left-handers possessed a broad innate superiority in fundamental motor or cognitive functions, one might expect them to be consistently overrepresented across a wide range of sports. However, empirical evidence indicates that left-handers are primarily overrepresented in certain interactive sports, particularly fencing (épée and foil) and table tennis, while the effect is weaker or inconsistent in other interactive sports such as badminton and largely absent in non-interactive sports like swimming, cycling, or gymnastics (Grouios et al., 2000; Hagemann, 2009; Loffing, 2017; Loffing & Hagemann, 2012, 2016; Loffing & Schorer, 2021; Raymond et al., 1996; Simon et al., 2025).
On the other hand, the nurture hypothesis, often referred to as negative frequency-dependent advantage (NFDA), suggests that left-handers benefit from their rarity in the general population: particularly in interactive sports, right-handers encounter left-handed opponents relatively infrequently and therefore may be less familiar with their perceptual cues and movement patterns (Grouios et al., 2000; Loffing & Hagemann, 2016; Loffing & Schorer, 2021; Raymond et al., 1996). As a result, left-handers may benefit from right-handers’ less developed perceptual skills (e.g., anticipation), motor responses, and tactical adaptations that are typically refined through repeated interactions (Coren, 1993; Loffing & Hagemann, 2016; Loffing & Schorer, 2021; Roi & Bianchedi, 2008). If this hypothesis were fully sufficient to explain the phenomenon, the left-hander advantage should diminish at the later stages of tournaments, such as semifinals or finals, where left-handers are more likely to compete against each other (Loffing & Schorer, 2021). However, both earlier and more recent data do not support this prediction (Raymond et al., 1996; Azémar, 2003; Simon et al., 2025), showing greater overrepresentation among higher-ranked players in fencing (épée and foil) and table tennis. Taken together, it seems both innate superiority and NFDA alone could not fully account for the left-hander advantage (Groothuis et al., 2013; Simon et al., 2025).
In the literature, analyses of winning rates associated with handedness remain relatively scarce, with only a few examples reported in boxing (Gursoy, 2009) and wrestling (Ziyagil et al., 2010). It is possible that the left-hander advantage arises from multiple interacting mechanisms, including elements of both innate superiority and NFDA, as well as differences in technical and tactical patterns (Groothuis et al., 2013; Simon et al., 2025). However, the specific technical and tactical processes potentially underlying this advantage remain poorly understood. While some studies have explored related perceptual strategies, such tracking visual fixations in foil fencing (Witkowski et al., 2018; 2020b), empirical data on actual behavioral execution during matchplay remain limited. Analyzing match performance indicators, such as techniques used and those leading to successful outcomes across different contexts, may provide more direct and practical insights into how left-handers maintain their advantage (Hughes & Franks, 2007; O'Donoghue, 2014). Similar approaches have been applied in other interactive sports, including cricket (Brooks et al., 2004), table tennis (Malagoli Lanzoni et al., 2019) and tennis (Loffing et al., 2009, 2010). Therefore, this study investigated the match performance of elite female épée fencers with different weapon-hand laterality using notational analysis.
The study had two main aims: first, to examine the prevalence and winning rate of left- and right-handed fencers across different tournament stages; and second, to investigate their technical and tactical patterns when facing left- and right-handed opponents. To address these aims, we examined the techniques, target areas on the opponent’s body, and the spatial zones of actions on the field of play (i.e., piste) of fencers across different dyadic combinations, including right-handers facing left- and right-handers, as well as left-handers facing left- and right-handers. Both actions performed and scored were examined.
Based on the historical overrepresentation data (Raymond et al., 1996; Azémar, 2003; Simon et al., 2025) and findings from other combat disciplines (Gursoy, 2009; Ziyagil et al., 2010), we hypothesized that left-handed fencers would be overrepresented and demonstrate higher winning rates, particularly at the later stages of tournaments. We further anticipated that this competitive advantage would be partially explained by different technical and tactical behaviors, including technique selection, target areas, and the use of piste zones. Specifically, if left-handers possess generalized perceptual-motor or attentional advantages as proposed by the innate superiority account (e.g., Grouios, 2004; Harris, 2010; Simon et al., 2025), they should show more proactive behaviors (e.g., performing more attacks and counterattacks), more flexible target selection and greater offensive piste control regardless of opponent handedness. In contrast, the NFDA account (e.g., Loffing & Hagemann, 2016; Loffing & Schorer, 2021; Raymond et al., 1996; Simon et al., 2025) predicts that such technical and tactical differences should emerge primarily in left-vs.-right dyads, where right-handers are less familiar with left-handed perceptual cues and movement patterns, but should diminish in left-vs.-left dyads (e.g., Loffing & Schorer, 2021; Simon et al., 2025).
Methods
Sample
This study analyzed a previously published dataset (Hsu & Chen, 2026), which comprised match of elite female épée fencers competing from the top 16 through to the final bouts at the World Championships in 2017, 2018, and 2019. Match videos obtained from the official YouTube channel of the International Fencing Federation (Fédération Internationale d'Escrime, FIE). Ethical approval was granted by the International Review Board of Fu Jen Catholic University (No: C110181).
Procedure
The present study used the same notational procedure reported in Hsu and Chen (2026). For completeness, the key elements of the coding procedure are summarized here. All bouts were analyzed using a systematic notational approach in which each scoring event was coded according to match metadata, fencer information, and technical-tactical variables. Match metadata included the year of competition (2017–2019), tournament stage (round of 16, quarterfinal, semifinal, final), and bout phase (first, second, third, or sudden-death overtime). Information regarding the participating fencers was also recorded, including their names and weapon-hand laterality (left- or right-handed). Technical actions were classified following FIE definitions as attack, riposte, counterattack, or “others” (including remise, redoublement, reprise of the attack, and counter-time actions) (FIE, 2023). Target areas were categorized as inner upper body, outer upper body, or lower body. The upper body was further subdivided into inner and outer areas based on fencer’s engarde weapon position. Specifically, body areas inside the guard were define as inner upper body, corresponding to the areas typically defended by the fourth position (quarte), whereas body areas outside the guard were defined as outer upper body, corresponding to areas typically defended by the sixth position (sixe) (Figure 1). The location of each action on the piste was determined by the midpoint between the fencer’s two feet at the moment the hit occurred. The piste was divided into six zones based on the official on-guard, warning, and end lines on both sides. These zones were sequentially coded from the focal fencer’s side to the opponent’s side as the focal fencer’s end, middle, and central zones (FE, FM, and FC, respectively), followed by the opponent’s central, middle, and end zones (OC, OM, and OE, respectively) (Figure 1). Additional variables included the hit outcome (single hit, double hit, or no hit), the score at the time of the action (0 to 15) and the final bout outcome. To ensure reliability of the notation, bot intra- and inter-rater reliability were assessed as described in Hsu and Chen (2026). Two experienced analysts independently coded the final bout of the 2015 World Championships twice with a two-week interval. Reliability for all categorical variables was assessed using Cohen’s kappa, showing excellent agreement (Cohen’s kappa = 0.90–1.00). Schematic illustration of target body areas and piste zones in an asymmetrical matchup. The piste is sequentially divided into six zones: FE (focal fencer’s end zone), FM (focal fencer’s middle zone), FC (focal fencer’s central zone), OC (opponent’s central zone), OM (opponent’s middle zone), and OE (opponent’s end zone). Target body areas are classified into the inner body (blue), outer upper body (red), and lower body (white); the mask is not highlighted due to its rare occurrence despite being a valid scoring target in épée. In the illustrated example, the left-handed fencer is designated as the focal fencer and scores via a counterattack to the right-handed opponent’s outer upper body in FC, while the right-handed fencer scores via an attack to the focal fencer’s inner upper body in OC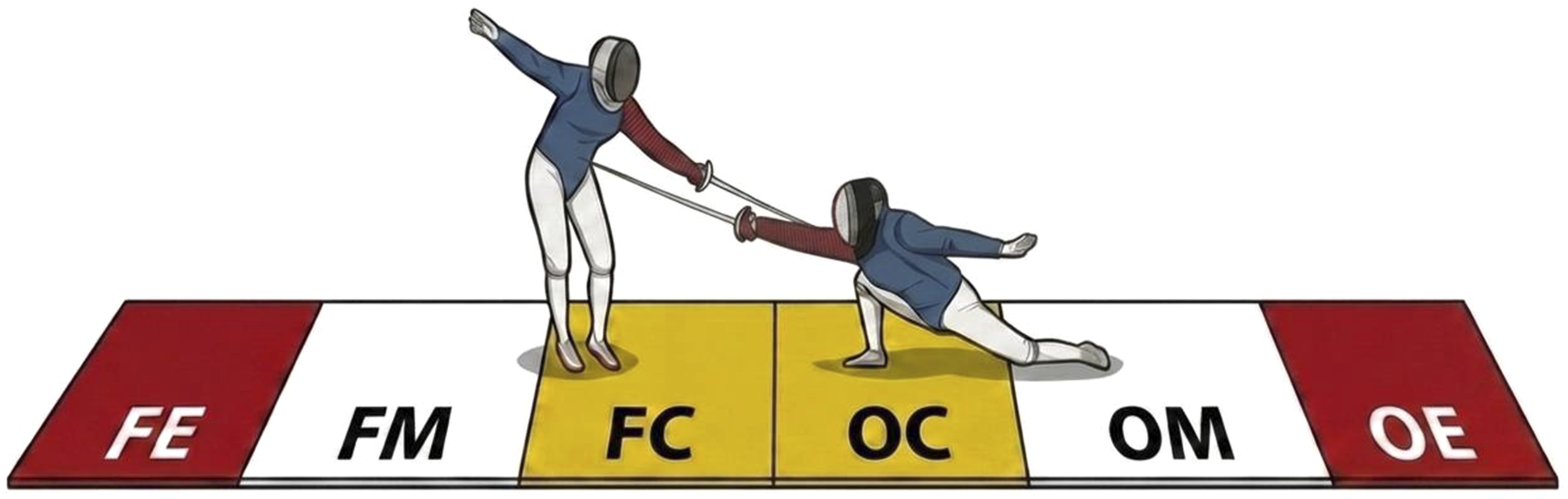
Data Analysis and Statistics
One-sample exact binomial tests were conducted to examine whether left-handed fencers were overrepresented relative to the population-level prevalence of 10.6% (Papadatou-Pastou et al., 2020) across tournament stages. Because several fencers competed in multiple bouts (e.g., across different years or tournament stages), overall left-handed prevalence was analyzed based on both total appearances and unique individuals. Exact (Clopper-Pearson) 95% confidence intervals (CI) for the observed proportions and effect sizes (Cohen’s h) (Cohen, 2013) were reported. Winning rates of left-handed fencers were subsequently analyzed across tournament stages. To examine technical and tactical behaviors, two separate sets of generalized linear mixed models (GLMMs) were conducted for performed and scored actions, respectively. This modelling approach was selected to account for the nested structure of the data, specifically the repeated observations within fencers, and to control for individual variability across the sample. In each set, the models assessed whether the distribution of techniques (4 categories), target areas (3 categories), and piste zones (6 categories) could be predicted by the focal fencer handedness, the opponent handedness, and their dyadic interaction. Each model specified a multinominal distribution with a generalized logit link function. The handedness of the focal fencers, the opponent, and their interaction were included as fixed effects, while fencer identity was included as a random intercept to account for the dependency between observations from the same individuals. All statistical analyses were conducted using IBM SPSS Statistics (version 29.0; IBM Corp., Armonk, NY, USA), with the significance level set at alpha = .05.
Results
The dataset initially included a total of 1,840 events across 45 matches: 12 right-vs.-right, 24 left-vs.-right, and 9 left-vs.-left matches. Among these, 4 events that occurred during sudden-death overtime rounds were excluded because they were not captured in the official video recordings. Additionally, 6 actions were excluded due to non-combativity, defined as a two-minute period without any valid touch under the regulations at that time. These consisted of 4 yellow-card and 2 red-card penalties, with the latter resulting in one point awarded to each fencer. As these instances did not involve any technical-tactical actions, they were excluded from the analysis. The final dataset comprised 1,830 notated actions.
The Prevalence and Winning Rate of Left-Handed Fencers Across Tournament Stages
Overall and Stage-specific Prevalence and Winning Rates of Left- and Right-Handed Fencers Across Tournament Stages, and Statistical Comparisons Between Left-Handed Prevalence and the Population-Level Rate (10.6%; Papadatou-Pastou et al., 2020)
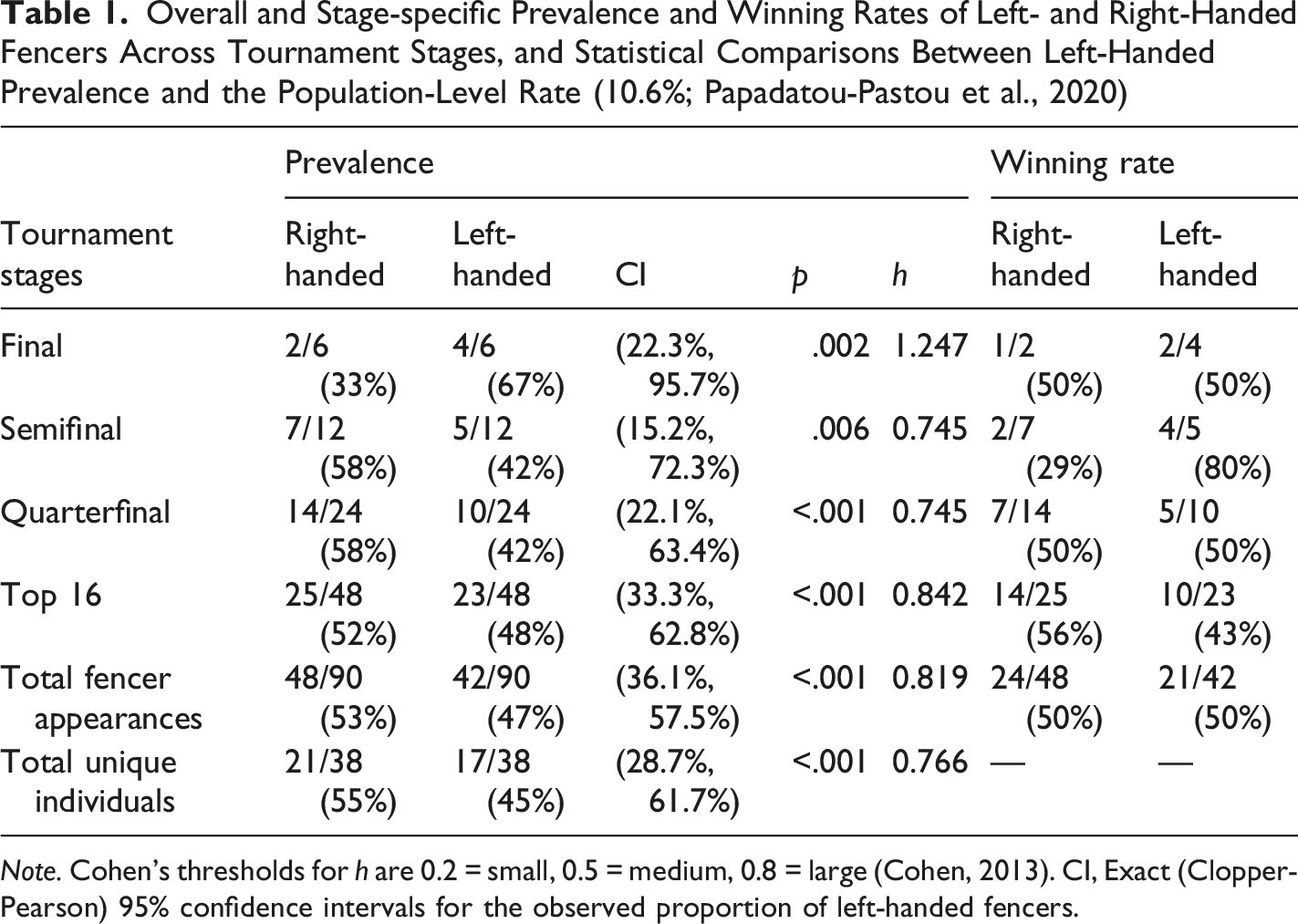
Note. Cohen’s thresholds for h are 0.2 = small, 0.5 = medium, 0.8 = large (Cohen, 2013). CI, Exact (Clopper-Pearson) 95% confidence intervals for the observed proportion of left-handed fencers.
Technical and Tactical Behaviors at the Level of Individual Actions
The distribution of the 1,830 actions performed across matchups was as follows: 492 actions from right-handed fencers against right-handed opponents, 982 actions from asymmetric matchups (491 from right-handed fencers against left-handed opponents, and equivalently, 491 actions from left-handed fencers against right-handed opponents), and 356 actions from left-handed fencers against left-handed opponents. Among these 1,830 actions, 1,111 were scored actions, consisting of 480 actions from right-handed fencers against right-handed opponents, 338 actions from right-handed fencers against left-handed opponents, 158 actions from left-handed fencers against right-handed opponents, and 135 actions from left-handed fencers against left-handed opponents. Figure 2 illustrates the absolute counts (panels A and B) and relative ratio (panels C and D) of techniques used and scored of fencers across the four handedness combinations. Total counts of techniques used (A) and scored (B), alongside the corresponding ratios of techniques used (C) and scored (D) across four handedness combinations. Data indicate the actions performed or scored by the focal fencers against the opponents (R = right-handed fencers; L = left-handed fencers)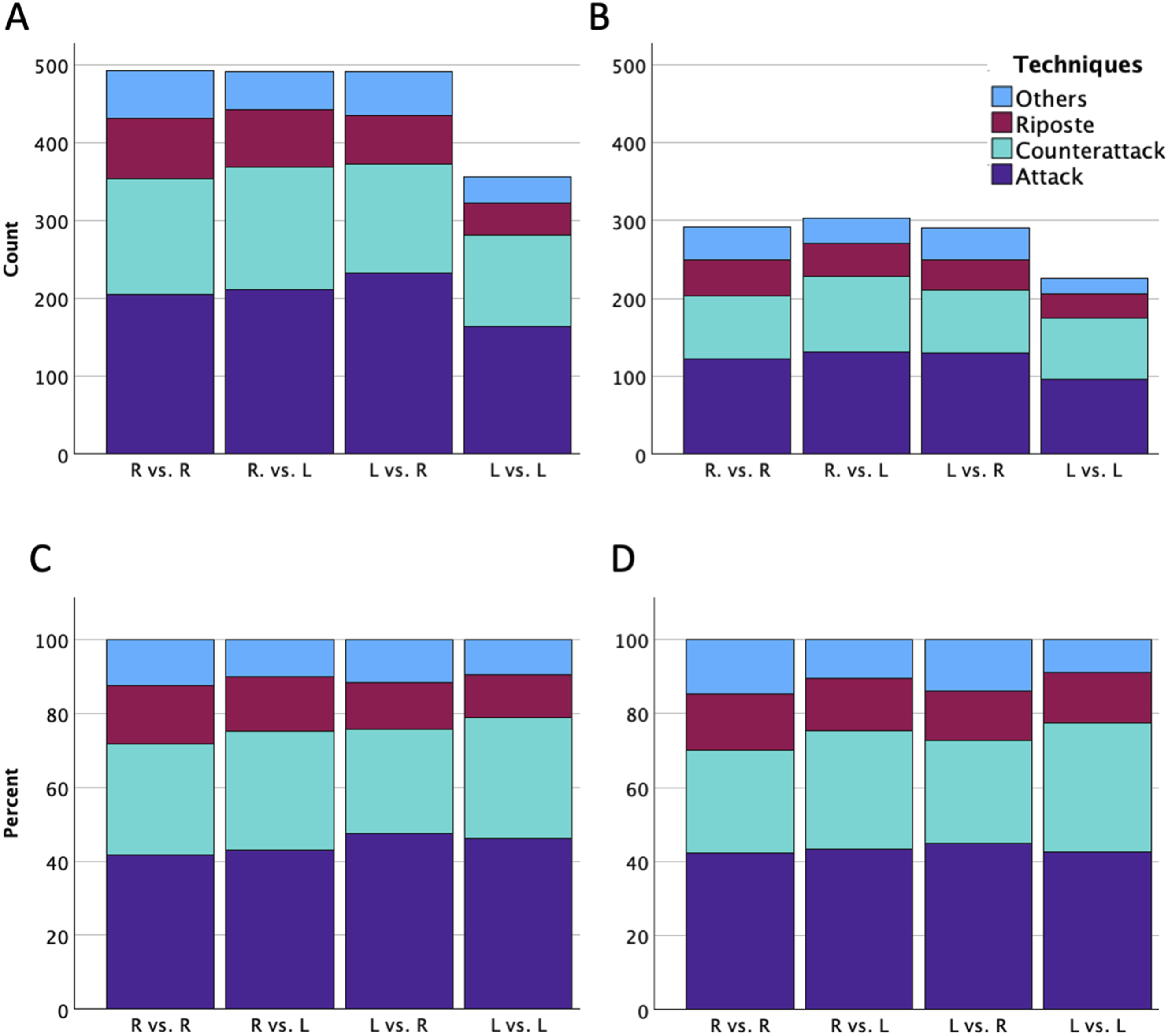
Techniques Used
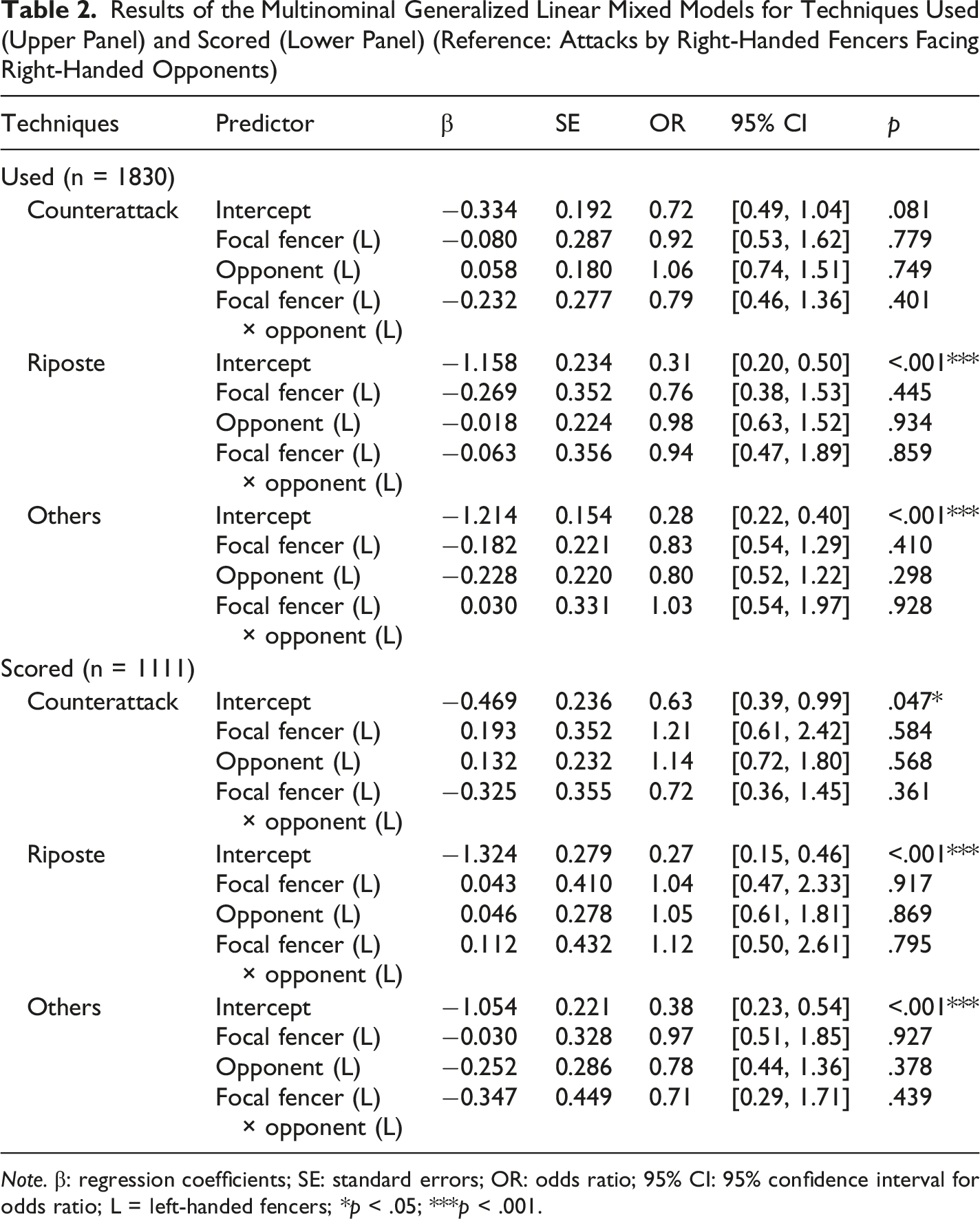
Results of the Multinominal Generalized Linear Mixed Models for Techniques Used (Upper Panel) and Scored (Lower Panel) (Reference: Attacks by Right-Handed Fencers Facing Right-Handed Opponents)
Note. β: regression coefficients; SE: standard errors; OR: odds ratio; 95% CI: 95% confidence interval for odds ratio; L = left-handed fencers; *p < .05; ***p < .001.
Techniques Scored
The GLMM of techniques scored revealed similar results to the GLMM of techniques used. The overall fixed effects were not significant for the focal fencer’s handedness (F(3,1099) = 0.233, p = .873), the opponent’s handedness (F(3,1099) = 1.454, p = .226), or their interaction (F(3,1099) = 0.508, p = .677). Baseline parameter estimates (Table 2, lower panel) showed that under the right-vs.-right condition, attacks remained the dominant scoring technique, with counterattacks (OR = 0.63, 95% CI [0.39, 0.99], p = .047), ripostes (OR = 0.27, 95% CI [0.15, 0.46], p < .001), and other techniques (OR = 0.38, 95% CI [0.23, 0.54], p < .001) being significantly less likely to score. Crucially, the lack of significant parameter estimates for the left-handed predictors and their interaction (all p values >.298) indicated that this scoring hierarchy remained highly stable across all handedness combinations. Similarly, the random effect of the fencer was significant for counterattacks (Variance = 0.603, p = .005) and ripostes (Variance = 0.680, p = .019), but not for others (Variance = 0.247, p = .151).
Body Areas Targeted
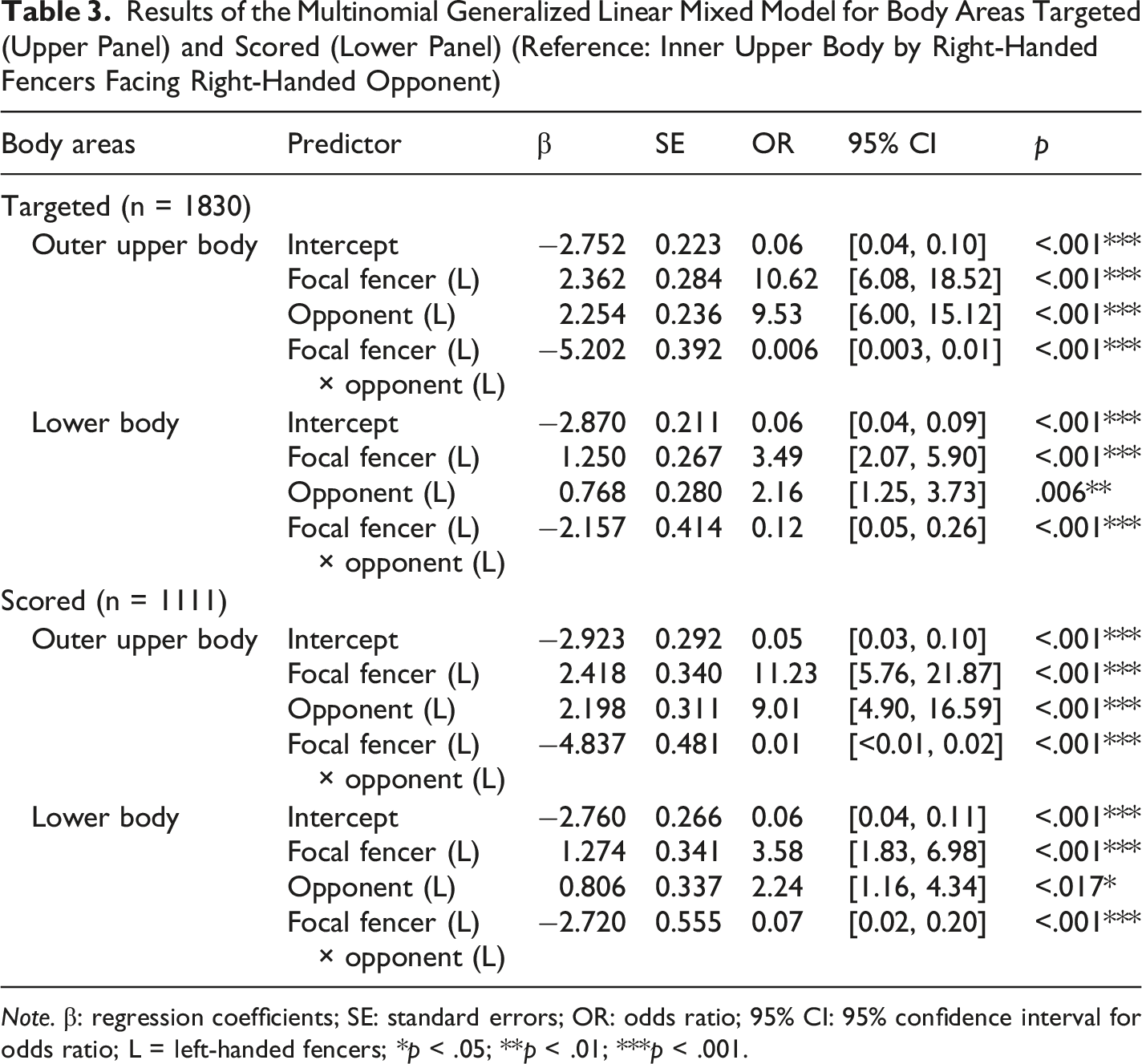
Figure 3 illustrates the absolute counts and corresponding relative ratio of the body areas targeted (panels A and C) and scored (panels B and D) of fencers across four handedness combinations. The multinomial GLMM of body areas targeted revealed a significant interaction effect between the handedness of the focal fencer and the opponent (F(2,1822) = 96.732, p < .001), whereas the main effects of the focal fencer’s handedness (F(2,1822) = 0.832, p = .435) and the opponent’s handedness (F(2,1822) = 2.496, p = .083) were not significant. This prominent interaction indicated that tactical targeting was fundamentally driven by whether the matchup was symmetrical (right-vs.-right, left-vs.-left) or asymmetrical (right-vs.-left, left-vs.-right). With the “inner upper body” specified as the reference category, baseline parameter estimates (Table 3, upper panel) indicated that it was the dominant target in symmetrical matchups. In asymmetrical matchups, fencers shifted their strategy toward open angles: left-handed fencers facing right-handed opponents showed significantly higher odds of targeting the outer upper body (OR = 10.62, p < .001) and lower body (OR = 3.49, p < .001). Similarly, right-handed fencers facing left-handed opponents showed higher odds of targeting the outer upper body (OR = 9.53, p < .001) and lower body (OR = 2.16, p = .006). Crucially, when two left-handed fencers faced each other, the strong negative interaction effects (beta = −5.202 for the outer upper body and beta = −2.157 for the lower body) countered these main effects. This interaction led to a massive reduction in the odds of targeting the outer upper body (OR = 0.006, p < .001) and lower body (OR = 0.12, p < .001). Consequently, the tactical targeting distribution of left-vs.-left matchups reverted to a profile highly resembling that of right-vs.-right matchups. The random effect of the fencer was significant for outer upper body (Variance = .220; p = .049), but not for lower body (Variance = .033; p = .719). Total counts of body areas targeted (A) and scored (B), alongside the corresponding ratios of body areas targeted (C) and scored (D) across four handedness combinations. Data indicate the actions performed or scored by the focal fencers against the opponents (R = right-handed fencers; L = left-handed fencers) Results of the Multinomial Generalized Linear Mixed Model for Body Areas Targeted (Upper Panel) and Scored (Lower Panel) (Reference: Inner Upper Body by Right-Handed Fencers Facing Right-Handed Opponent) Note. β: regression coefficients; SE: standard errors; OR: odds ratio; 95% CI: 95% confidence interval for odds ratio; L = left-handed fencers; *p < .05; **p < .01; ***p < .001.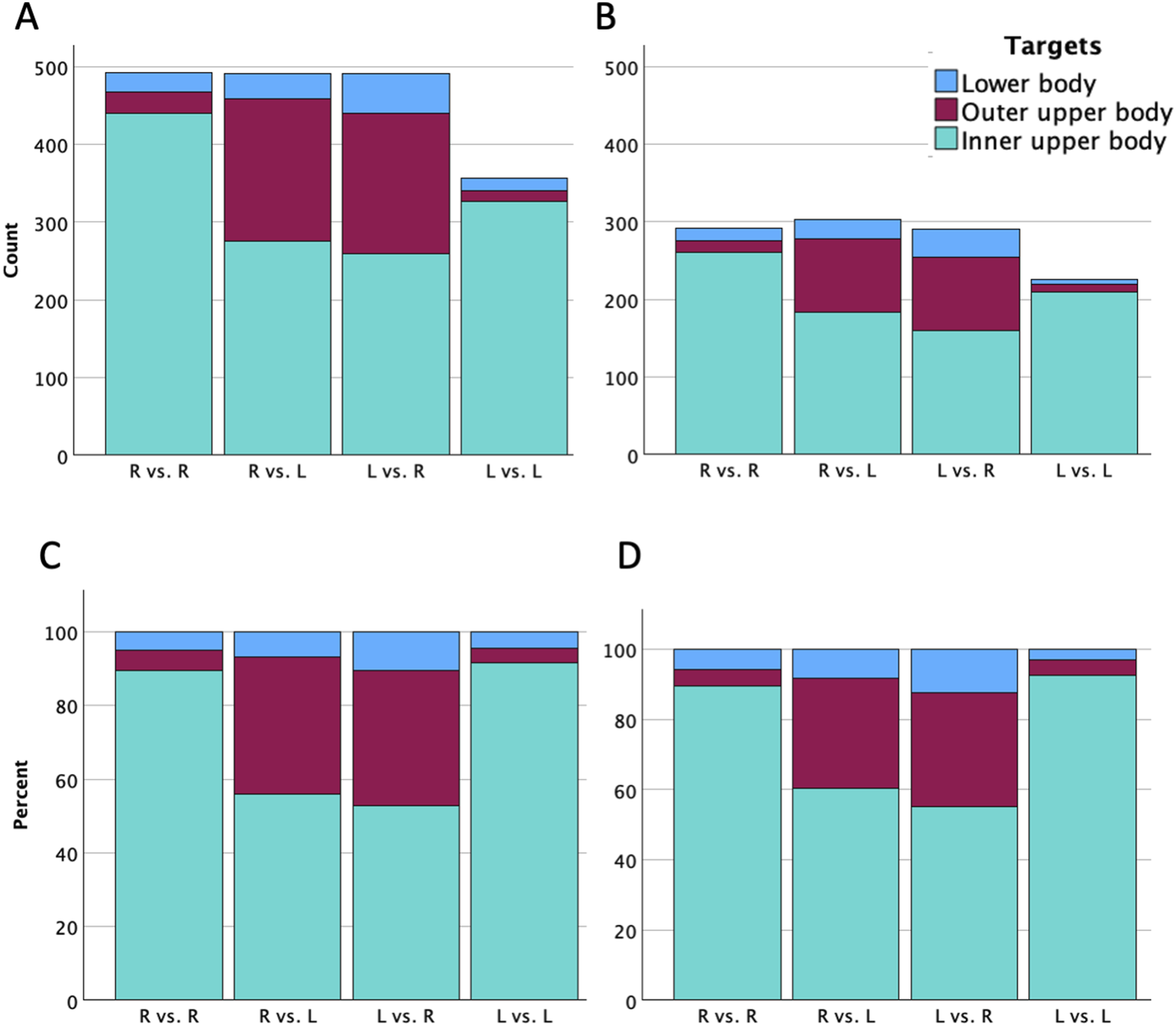
Body Areas Scored
Mirroring the targeting patterns, the GLMM for body areas scored revealed a highly significant interaction between the focal fencer and the opponent’s handedness (F(2,1103) = 59.011, p < .001), while the main effects remained non-significant (focal fencer’s handedness: F(2,1103) = 0.042, p = .959; opponent’s handedness: F(2,1103) = 2.286, p = .102). The parameter estimates closely followed the targeting model (Table 3, lower panel). Left-handed fencers in asymmetrical dyads successfully scored on the outer upper body (OR = 11.23, p < .001) and lower body (OR = 3.58, p < .001) at significantly higher rates. However, in left-vs.-left (symmetrical) matchups, this tactical advantage disappeared due to the significant interaction effect (outer upper body: beta = −4.837, OR = 0.01, p < .001; lower body: beta = −2.720, OR = 0.07, p < .001), causing the scoring distribution to match the baseline right-vs.-right profile. Unlike the targeting model, the random effect of fencer was not significant no matter for outer upper body (Variance = 0.139, p = .185) or lower body (Variance = 0.108, p = .418).
Piste Zones Used
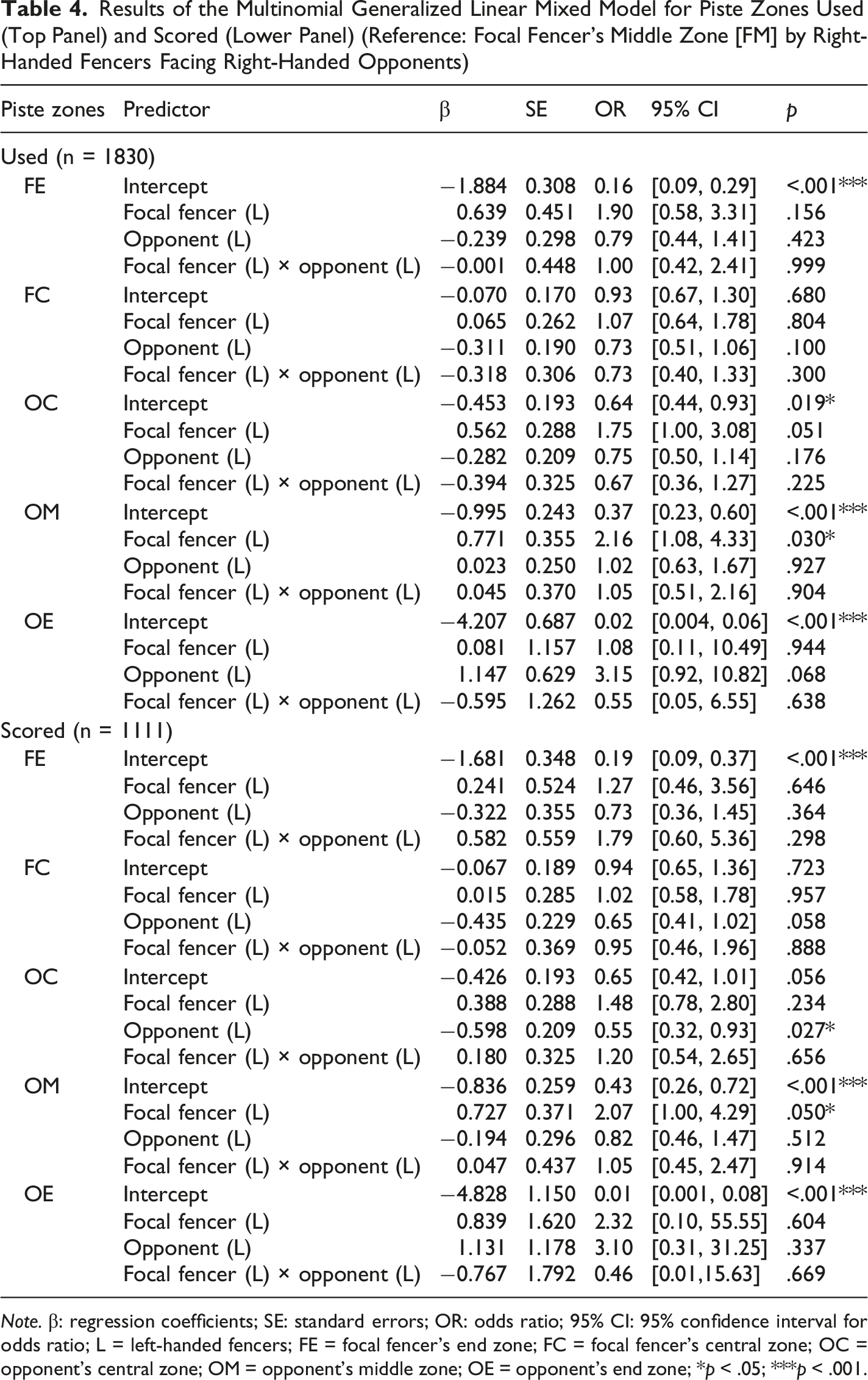
Figure 4 presents the absolute counts and corresponding relative ratio of the piste zones used (panels A and C) and scored (panels B and D) of fencers across handedness combinations. The fixed effect of the opponent’s handedness (F(5,1810) = 2.738, p = .018) was significant, whereas the focal fencer’s handedness (F(5,1810) = 1.191, p = .053) and the interaction effect (F(5,1810) = 0.526, p = .756) were not. With the FM specified as the reference category under the baseline right-vs.-right condition, parameter estimates (Table 4, upper panel) indicated that the FM was the most frequently used area, followed sequentially by the FC, OC, OM, FE, and OE. The odds of executing actions in other zones were significant lower compared to the FM (OR values <0.64, p values <.019), except for the FC (OR = 0.93, p = .680). Moreover, left-handed fencers were significantly more likely to take action in the OM (OR = 2.16; p = .030), and descriptively higher in the OC (OR = 1.75; p = .051), than right-handed fencers. No other parameter comparisons reached statistical significance. The random effect revealed significant individual variability across all piste zones (FC: Variance = .253; p = .022; OC: Variance = .351; p = .009; OM: Variance = .606; p = .007; FE: Variance = .882; p = .006; OE: Variance = 2.849; p = .012). Total counts of piste zones used (A) and scored (B), alongside the corresponding ratios of piste zones used (C) and scored (D) across four handedness combinations. Data indicate the actions performed or scored by the focal fencers against the opponents (R = right-handed fencers; L = left-handed fencers; FE = focal fencer’s end zone; FM = focal fencer’s middle zone; FC = focal fencer’s central zone; OC = opponent’s central zone; OM = opponent’s middle zone; OE = opponent’s end zone) Results of the Multinomial Generalized Linear Mixed Model for Piste Zones Used (Top Panel) and Scored (Lower Panel) (Reference: Focal Fencer’s Middle Zone [FM] by Right-Handed Fencers Facing Right-Handed Opponents) Note. β: regression coefficients; SE: standard errors; OR: odds ratio; 95% CI: 95% confidence interval for odds ratio; L = left-handed fencers; FE = focal fencer’s end zone; FC = focal fencer’s central zone; OC = opponent’s central zone; OM = opponent’s middle zone; OE = opponent’s end zone; *p < .05; ***p < .001.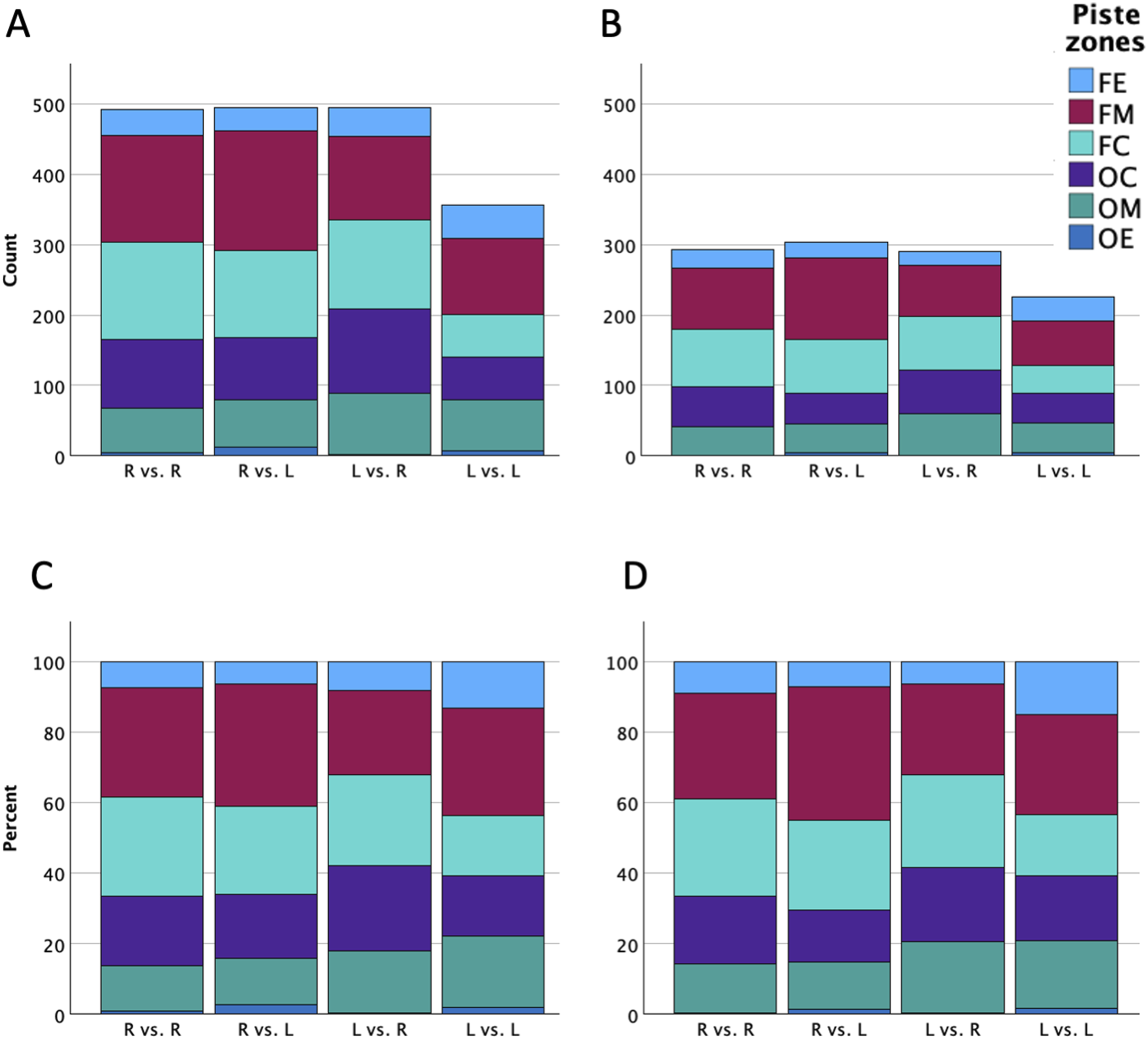
Piste Zones Scored
Consistent with the patterns observed for piste zones usage, the GLMM for piste zones scored revealed a significant fixed effect of the opponent’s handedness (F(5,1091) = 2.250, p = .047), whereas the main effect of the focal fencer’s handedness (F(5,1091) = 1.855, p = .099) and the interaction effect (F(5,1091) = 0.323, p = .899) were not significant. The pattern of the parameter estimates remained largely consistent with that observed in the GLMM for piste zones usage (Table 4, lower panel). Notably, the differences between the FM and OC was no longer statistically clear as the confidence interval expanded slightly (OR = 0.65, 95% CI [0.42, 1.01], p = .056), highlighting a descriptive drop in scoring efficiency with the OC. Left-handed fencers maintained a higher tendency to score in the OM (OR = 2.07, p = .050) compared to right-handed fencers. This advantage was not observed in the OC (OR = 1.48, p = .234); instead, right-handed fencers showed a lower tendency of scoring in the OC (OR = 0.55, p = .027). The random effect of the fencer was significant across all piste zones (OC: Variance = .309; p = .034; OM: Variance = .467; p = .033; FE: Variance = .908; p = .014; OE: Variance = 2.961; p = .044), except for FC (Variance = .177; p = .121).
Discussion
While the innate superiority hypothesis and NFDA hypotheses have long surrounded the left-hander advantage in interactive sports such as fencing (Harris, 2010; Loffing & Hagemann, 2016; Loffing & Schorer, 2021; Raymond et al., 1996; Simon et al., 2025), the technical and tactical patterns potentially underlying this advantage remain poorly understood. To tackle this, we examined the match performance of elite female épée fencers in the 2017–2019 World Championships using notational analysis. The results are discussed as follows.
Left-Handers Showed Elevated Prevalence Across Tournament Stages, With Their Winning Rate Increasing up to the Semifinals
The prevalence of left-handers among the top 16 fencers was approximately 50%, whether calculated based on total appearances (47%) or unique individuals (45%). This finding corroborates the well-documented overrepresentation of left-handers in interactive sports relative to the general population (Hagemann, 2009; Loffing & Hagemann, 2016; Loffing & Schorer, 2021; Raymond et al., 1996; Simon et al., 2025). Notably, left-handed prevalence remained relatively stable from the round of 16 to the semifinals (42%–48%) but increased sharply to 67% in the finals. This narrative directly supported our first hypothesis and aligned with previous research showing a progressive rise in left-handed representation at later tournament stages; for example, 24%–38% in the top 8 (Azémar & Stein, 1994; Raymond et al., 1996), and 26%–44% in Olympic finals (Azémar, 2003). The differences in magnitude between studies may reflect gender or temporal variations: our data comprised female épée fencers competing between 2017 and 2019, whereas earlier data focused on male épée fencers from previous decades (e.g., 1979–1993 in Azémar & Stein, 1994). Although all stages showed large effect sizes (Cohen’s h > 0.8), the finals exhibited an exceptionally large effect (1.247), reflecting the markedly high observed proportion (67%) relative to expected general population rate of 10.6%. However, our sample size for the finals was limited (6 fencers from 3 matches); thus, these results should be interpreted with caution. Future research using broader datasets across multiple years is warranted to verify whether this increasing trend among female épée fencers has strengthened in recent years.
The NFDA hypothesis has been used to account for the left-hander’s advantage, suggesting that this advantage might diminish at later tournament stages as left-handers increasingly encounter one another (e.g., Harris, 2010; Loffing & Hagemann, 2016; Raymond et al., 1996). To the best of our knowledge, no previous study has directly examined left-handers’ winning rates when facing other left-handers. Our findings provided empirical support for this hypothesis: when left-handers became heavily overrepresented in the finals (67%), their winning rate dropped to match that of right-handers (50% vs. 50%), contrasting with the clear advantage they maintained in the semifinals (80% vs. 29%). Nevertheless, a larger dataset is needed to confirm this pattern more conclusively.
Congruence Between Performed and Scored Actions
We examined whether the distributions of the technique types, target body areas, and piste zones, across both performed and scored actions, were influenced by the handedness of the focal fencer, the opponent, or their interaction. A close correspondence between the models of performed and scored actions was consistently observed across the three domains, suggesting that the technical-tactical selection and spatial positioning are key determinants of scoring success. Nevertheless, several subtle discrepancies emerged.
In terms of technique, analyzing the parameter magnitudes (effect sizes) revealed a consistent trend of reduced scoring efficiency for counterattacks relative to attacks. Under the baseline condition, the odds of executing a counterattack were descriptively lower than those of an attack, though the confidence interval marginally spanned the baseline threshold (OR = 0.72, 95 % CI [0.49, 1.04], p = .081). Importantly, this gap widened for scoring outcomes, where odds of successfully scoring with a counterattack dropped significantly lower than with an attack (OR = 0.63, 95 % CI [0.39, 0.99], p = .047). This downward shift in the odds ratios (from 0.72 to 0.63) underscores a descriptive decrease in tactical efficiency for counterattacks.
A similar pattern driven by effect magnitudes was observed for piste zones. Actions were performed more frequently in the FM than in the OC (OR = 0.64, 95 % CI [0.44, 0.93], p = .019). When examining successful touches, however, the confidence interval expanded to span across the baseline threshold (OR = 0.65, 95 % CI [0.42, 1.01], p = .056), suggesting a descriptive drop in scoring efficiency within the OC. This trend may be related to the lower tendency of right-handed fencers scoring in the OC when facing left-handed opponents, a pattern that was not evident in the distribution of executed actions. Taken together, these discrepancies between performed and scored actions indicated that, even at the elite level, certain technical and spatial contexts are associated with reduced efficiency in converting actions into successful outcomes (Hughes & Franks, 2007; O'Donoghue, 2014).
Technique Distribution: The Influence of Individual Style Over Handedness
Our results confirmed a highly consistent pattern across all handedness combinations: Attacks were the most frequently used and scored technique, followed by counterattacks and ripostes. This distribution aligns with established fencing doctrine, which emphasizes the predominance of offensive actions in épée (Czajkowski, 2009; Garret et al., 1994; Barth & Beck, 2006; Harris, 2010; Sowerby, 2014; Szabo, 1977). Notably, counterattacks accounted for approximately one-third of total actions, highlighting the unique nature of épée. The allowance of double hits and the absence of right-of-way rules promote reactive scoring strategies, making counterattacks a particularly viable and frequently employed option (e.g., FIE, 2023; Szabó, 1997).
Contrary to our second hypothesis, which anticipated distinct technical choices driven by literalities, the absence of significant fixed effects of handedness suggested that overall technique selection was not systematically influenced by whether a fencer is left- or right-handed, nor by the handedness pairing. This directly refuted a generalized innate superiority explanation. However, this finding should be interpreted with caution, as it may partially reflect the relatively broad categorization of techniques used in the present study. In practice, attacks and ripostes include both simple and compound forms, while counterattacks encompass various stop-hits made with or without opposition and within a defined fencing time (FIE, 2023). It is therefore possible that more fine-grained differences exist at the subcategory level, which could not be captured in the current dataset. Future research should incorporate more detailed technical classifications to better resolve these distinctions.
Importantly, the significant random effects for counterattacks and ripostes indicated substantial inter-individual variability in the use of these techniques. This suggested that elite fencers may exhibit distinct tactical profiles (Hughes & Franks, 2007; O'Donoghue, 2014), with some acting as “counterattack specialists” or “riposte-oriented” performers, while others remain predominantly attack-focused, regardless of handedness. Conversely, the non-significant variance for the “others” category indicated that these less frequent techniques were used at a relatively consistent baseline level across fencers, reflecting limited individual differences.
Aiming Adaptation: Target Selection and Scoring
Although the entire body constitutes a valid target in épée, the inner upper body emerged as the dominant area for targeting and scoring across all handedness combinations. This likely reflects its larger surface area and its alignment with the most direct attack trajectories. The finding validating our second hypothesis was the significant interaction effect between the focal fencer’s and opponent’s handedness. The absence of significant main effects indicated that neither “being left-handed” nor “facing a left-handed opponent” independently determined targeting and scoring behavior. Rather, tactical decisions emerged from the matchup type. Specifically, compared to right-handed fencers facing right-handed fencers, both being left-handed and facing a left-handed opponent were associated with substantially increased odds of targeting and scoring the outer upper body and lower body (ORs ranging from 2.16 to 10.62 for targeting, and from 3.58 to 11.23 for scoring). This pattern suggested that the asymmetrical matchups (right-vs.-left or left-vs.-right) altered the geometry of interaction, effectively opening the outside line. Because guard positions do not mirror each other in these configurations, the outer upper body becomes a more accessible and strategically viable target for bypassing the opponent’s blade.
In contrast, when left-handed fencers faced other left-handed opponents, the likelihood of targeting and scoring both the outer upper body (OR = 0.006 for targeting; OR = 0.001 for scoring) and lower body (OR = 0.12 for targeting; OR = 0.07 for scoring) decreased markedly. This sudden tactical reversion in same-handed dyads provided evidence for the NFDA account over the innate superiority hypothesis. Because left-handed fencers are more accustomed to facing right-handed opponents; thus, when encountering a same-handed opponent, their typical perceptual-motor advantages regarding the outside line no longer find an uncalibrated opponent to exploit (Harris, 2010; Loffing & Hagemann, 2016; Raymond et al., 1996), compelling them to revert to a more conservative, centrally focused strategy highly resembling the baseline right-vs.-right profile.
The random effects analysis further clarified these patterns. Significant between-fencer variability was observed only for targeting the outer upper body, but not for lower body targeting or for scoring in either region. This suggested that while some variability in targeting the outer upper body may reflect individual preferences, successful scoring in these peripheral areas was less a matter of individual style and more a context-driven tactical necessity. From an applied perspective, these findings have some implications (Hughes & Franks, 2007; O'Donoghue, 2014). Right-handed fencers should be prepared for increased attacks to their outer upper body side and lower limbs when facing left-handed opponents, with training emphasizing outside-line control and leg defense in asymmetrical matchups. Left-handed fencers should adopt similar preparations when facing right-handed opponents. Conversely, left-handed fencers may benefit from specifically developing strategies to exploit outer and lower targets in left-vs.-left matchups, where these options appear underutilized.
Spatial Adaptation: Piste Zones Utilization and Scoring
The present findings provided insight into how fencers regulate their spatial positioning along the piste. Across all matchups, the FM was the most frequently used and most successful location, followed by adjacent central zones (FC and OC) and OM, and finally the end zones. From a tactical perspective, the central 4 meters represents a balance between risk and opportunity, allowing fencers to initiate attacks while preserving sufficient space to retreat or respond to opponent actions. The multinomial GLMM for piste zones revealed only a significant main effect of the opponent’s handedness, with no significant effect of the focal fencer’s handedness or their interaction. Intriguingly, this lack of interaction complicated a pure NFDA interpretation, which would predict spatial anomalies to disappear in left-vs.-left matchups. Instead, the data revealed that left-handed fencers consistently demonstrated a more invasive spatial strategy across all dyads, operating significantly more in the OM (OR = 2.16, 95 % CI [1.08, 4.33], p = .030) and marginally in the OC (OR = 1.75, 95 % CI [1.00, 3.08], p = .051). This persistent shift toward the opponent’s half side of the piste offered partial support for an innate or deeply ingrained proactive spatial control mechanism (e.g., Grouios et al., 2000; Simon et al., 2025). By occupying more advanced piste zones (i.e., closer to the opponent’s side), left-handed fencers may be better able to impose pressure, reduce reaction time for their opponent, and capitalize on disrupted perceptual-motor calibration.
Moreover, the significant random effects across nearly all piste zones (particularly with high variance in the OE: Variance = 2.849 and 2.961 for piste zones used and scored, respectively), indicated substantial inter-individual variability. Some fencers may prefer to engage at longer distances and retreat toward their own end, whereas others may consistently press forward and operate in advanced zones. The particularly large variance observed for the OE suggested that extreme spatial strategies (e.g., pushing the opponent to the back of the piste) may be characteristic of specific individuals rather than a generalizable pattern.
From a coaching perspective, these results highlighted the importance of training spatial adaptability rather than fixed positioning strategies. Given the high inter-individual variability, fencers should be encouraged to develop flexible distance management skills that allow them to operate effectively across different zones of the piste. Coaches should prepare right-handed fencers for the “territorial loss” they are likely to experience against left-handers. Training should focus on “fencing from the back”, maintaining composure and scoring while being pushed into the FM or FC zones. Concurrently, left-handed fencers should continue to leverage their tendency to occupy the opponent’s half of the piste, as this is where they are statistically most active and successful at scoring.
Conclusions
This study confirmed the well-documented overrepresentation of left-handed fencers in elite female épée (Loffing & Schorer, 2021; Raymond et al., 1996; Azémar, 2003; Simon et al., 2025). While left-handers were more prevalent in high-level populations, their winning rates in final matches were comparable to those of right-handers. However, this pattern needs to be validated by future studies involving a larger sample size. The technical repertoire, specifically the frequency of attacks, counterattacks, and ripostes, remained remarkably consistent across all handedness matchups, indicating the weapon-hand laterality does not dictate basic technique choice. While the inner upper body remained the universal primary target, asymmetrical matchups (left-vs.-right and right-vs.-left) triggered a significant shift toward the outer upper body and lower body, a pattern that is attenuated in left-handers facing left-handers. Crucially, the left-handed fencers demonstrated a more invasive spatial strategy, exhibiting a higher propensity to engage and score in the opponent’s middle zone regardless of matchup symmetry. Collectively, these findings suggest that the left-hander’s advantage in elite female épée is driven less by technique selection, but more by interplay between context-dependent target adaptation and a generalized proactive spatial exploitation. These findings indicate that while aiming strategies align more closely with the NFDA hypothesis, spatial control elements preserve features consistent with an innate superiority account (Groothuis et al., 2013; Simon et al., 2025). However, it must be noted that this study analyzed data at the action level, future research should transition to match-level analyses to determine how these granular tactical patterns and sequential interactions cumulatively influence match outcomes.
Footnotes
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.