
Abstract
Objective
To examine the systemic biological effects of excess dietary iron and elevated hemoglobin in the context of the 2024 World Health Organization revision of anemia thresholds. This study integrates experimental evidence from BALB/c mice with previously proposed diagnostic indices (SF/TSAT and SF×TSAT) to distinguish iron overload from inflammation.
Methods
This study is based on an experimental murine model, with human evidence incorporated to provide contextual interpretation. BALB/c mice were exposed to graded oral iron supplementation, and hematocrit and glycemia were evaluated across different metabolic conditions. The findings were interpreted in the context of established human diagnostic criteria and recognized modulators of iron absorption.
Results
Iron supplementation induced a dose-dependent increase in hematocrit, with higher values observed in operationally defined obese mice. Hematocrit–glycemia coupling strengthened following supplementation (overall r = 0.62; p < 0.01), with stronger correlations in operationally defined obese mice (r = 0.74) and in females (r = 0.72), suggesting that both metabolic status and sex may modulate this relationship. The 2024 WHO revised hemoglobin thresholds are expected to reduce global anemia prevalence estimates, potentially modifying population-level interpretations. Dietary ascorbic acid intake and circadian hepcidin rhythms emerged as relevant determinants of iron absorption and systemic iron dynamics.
Conclusions
Both iron deficiency and iron excess are associated with adverse outcomes, reflecting a U-shaped relationship between iron status and risk. These findings support precision-based iron supplementation strategies that incorporate validated diagnostic markers, while considering metabolic context and chronobiological regulation. Such approaches may improve anemia management while minimizing unintended iron excess in non-deficient populations.
Keywords
Introduction
Following our previous work, 1 in which we introduced diagnostic indices to distinguish iron overload from inflammation in children (serum ferritin/transferrin saturation [SF/TSAT] and serum ferritin × transferrin saturation [SF×TSAT]), the present study investigates the systemic effects of excess dietary iron. We examine its potential contribution to oxidative stress, insulin resistance, and metabolic dysregulation, emphasizing the need for selective, diagnosis-based supplementation strategies. In addition, we integrate experimental animal data with observational human evidence to evaluate the implications of the updated World Health Organization definition of anemia at high altitude (HA).
Anemia affects approximately 1.92 billion people worldwide, with a particularly high prevalence among children aged 6–59 months, approaching 40%. This global burden has led to widespread implementation of iron supplementation programs, multiple micronutrient powder distribution, and food fortification initiatives. In 2023, 35.5% of pregnant women aged 15–49 years were diagnosed with anemia globally. 2 However, distinguishing the underlying causes of anemia remains essential, as its clinical significance and management differ substantially across physiological and pathological conditions.
Physiological anemia may result from hemodilution during pregnancy or from the transition between fetal hemoglobin (HbF) and adult hemoglobin (HbA) in infancy, without indicating disease. In contrast, pathological anemia arises from diverse causes including bleeding, chronic inflammation, renal disease, hemoglobinopathies, infections, and parasitic conditions. 3 Despite extensive global efforts over the past two decades, progress in reducing anemia has been modest. The WHO has emphasized that anemia should be addressed comprehensively rather than solely through iron supplementation 4 and achieving the 50% reduction target set by the Sustainable Development Goals remains unlikely. 5
These limitations have prompted renewed scrutiny of current diagnostic frameworks, particularly the reliance on hemoglobin as a sole indicator. Hemoglobin alone cannot distinguish iron deficiency from other physiological or pathological processes, and similarly, elevated hemoglobin or hematocrit values do not differentiate true iron overload from redistribution or erythropoietic stimulation. 6
In March 2024, the WHO revised hemoglobin thresholds for anemia, lowering the diagnostic cut-off to 10.5 g/dL for children aged 6–23 months and pregnant women in the second trimester, and introducing altitude adjustments for populations living above 500 m. 7 While these changes refine prevalence estimates, controversy remains regarding the 0.4 g/dL decrease applied at 500–999 m, where oxygen saturation is minimally affected.3,6 Application of these updated criteria has substantially reduced anemia prevalence estimates in several countries, including India, Argentina, and Peru.8–10
These findings suggest that previous thresholds may have contributed to overestimation of anemia prevalence and, consequently, to widespread iron supplementation practices. Given that excessive iron intake is associated with oxidative stress, metabolic disturbances, and increased susceptibility to infection, the updated WHO framework underscores the need to reassess current prevention strategies. 11 Within this context, it is also essential to examine the opposite end of the hemoglobin spectrum, including conditions of elevated hemoglobin and iron excess, and their clinical implications.
Methods
Study design
Because universal iron supplementation policies limit the feasibility of controlled trials in non-deficient humans, an experimental murine model was used to evaluate hematologic and metabolic responses to graded iron exposure.
Animals and experimental model
Eighty BALB/c mice (40 males and 40 females; 8 weeks old; 25–30 g) were maintained under standard laboratory conditions with free access to water and chow (∼50 ppm iron). This strain was selected for its stable metabolic and hematologic profile, allowing assessment of moderate metabolic heterogeneity without the severe obesity phenotype observed in C57BL/6J mice.12–15
To account for metabolic variability, half of the animals were exposed to a high-fat hypercaloric diet, resulting in increased body weight compared with controls. Because direct quantification of adiposity (e.g., fat mass or body composition) was not performed, obesity in this study was operationally defined based on body weight divergence between diet groups. Accordingly, the term “operationally defined obese mice” is used throughout the manuscript to reflect this definition.
All groups consumed identical standard chow with an iron content corresponding to standard maintenance levels (∼50 ppm), and no diet-based iron manipulation was performed. Therefore, intergroup differences cannot be attributed to variability in dietary iron composition.
Intervention
Iron was administered as ferrous sulfate by daily orogastric gavage at graded doses (1, 2, 4, and 5 mg/kg/day) for 30 days. Housing and feeding conditions were identical across groups, ensuring uniform baseline iron exposure and endogenous recycling, including coprophagic behavior inherent to rodent physiology.16,17
Measurements
Animals were allocated sequentially to treatment groups without formal randomization, which should be considered when interpreting potential allocation-related variability. However, given the controlled experimental conditions, standardized housing, and identical feeding across groups, this allocation approach is unlikely to introduce systematic bias in the dose–response comparisons.
The final design included 80 mice distributed across 20 groups (n = 4 per group), allowing evaluation of iron dose and metabolic status.18,19 Blood samples (∼50 µL) were collected from the tail vein without anesthesia using minimally invasive techniques. Animals were gently restrained to minimize stress. Baseline measurements were obtained prior to iron administration; however, the primary analyses focused on post-intervention values to assess dose–response relationships.
Ethics
All procedures were approved by the Ethics Committee for the Use of Animals of the Universidad Peruana Cayetano Heredia on 4 January 2023 (approval code: DC: 001-01-2023) and complied with ARRIVE 2.0 guidelines. 20 Animals were euthanized by intraperitoneal pentobarbital sodium (200 mg/kg), with death confirmed by absence of cardiac activity and reflexes.
Statistical analysis
Data met parametric assumptions (Shapiro–Wilk and Levene tests, p > 0.05). Differences were evaluated using two-way ANOVA (Group × Dose). Within-group dose effects were assessed using one-way ANOVA followed by Tukey’s HSD post hoc test. Associations between glycemia and hematocrit were assessed using Pearson correlation coefficients. Human data were not part of the experimental design and are included solely for contextual interpretation in the Discussion. Statistical significance was set at p < 0.05.
Results
Dose–response relationship between iron intake and hematocrit values in healthy and operationally defined obese male and female mice after 30 days of iron supplementation.

Data are expressed as mean ± SEM (n = 4 per subgroup). Within-group dose effects were analyzed using one-way ANOVA followed by Tukey’s HSD post hoc test. A global two-way ANOVA (Group × Dose) was performed to assess main effects and interaction. Partial eta-squared (η2p) was calculated as effect size. Significant dose effects were observed in operationally defined obese mice (F = 10.23; p = 0.000014) and healthy mice (F = 4.38; p = 0.0057). A significant main effect of metabolic status was detected (F = 52.66; p = 3.25 × 10-10). No statistically significant sex differences were detected at any iron dose (all p > 0.05). In operationally defined obese mice, hematocrit increased significantly from 0 mg/kg to all higher doses, with 5 mg/kg also exceeding 1–2 mg/kg. In healthy mice, only 4 and 5 mg/kg differed significantly from baseline. Effect sizes were large for Group (η2p = 0.425) and Dose (η2p = 0.433), whereas the Group × Dose interaction was small (η2p = 0.037). Observed power was high for main effects (≈1.00) but low for interaction (0.16).
Dose–response relationship between iron intake and blood glucose levels in healthy and operationally defined obese male and female mice after 30 days of iron supplementation.
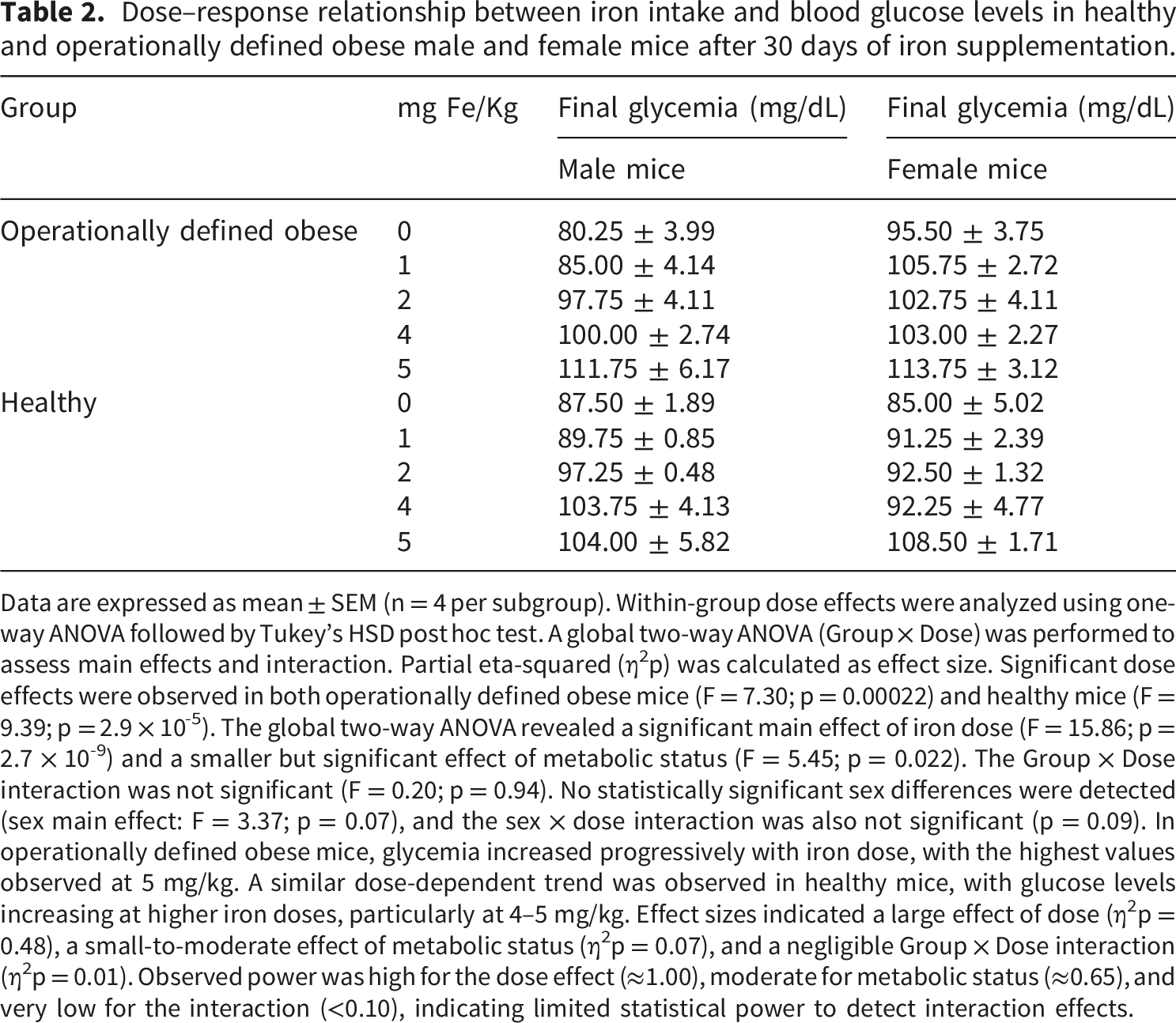
Data are expressed as mean ± SEM (n = 4 per subgroup). Within-group dose effects were analyzed using one-way ANOVA followed by Tukey’s HSD post hoc test. A global two-way ANOVA (Group × Dose) was performed to assess main effects and interaction. Partial eta-squared (η2p) was calculated as effect size. Significant dose effects were observed in both operationally defined obese mice (F = 7.30; p = 0.00022) and healthy mice (F = 9.39; p = 2.9 × 10-5). The global two-way ANOVA revealed a significant main effect of iron dose (F = 15.86; p = 2.7 × 10-9) and a smaller but significant effect of metabolic status (F = 5.45; p = 0.022). The Group × Dose interaction was not significant (F = 0.20; p = 0.94). No statistically significant sex differences were detected (sex main effect: F = 3.37; p = 0.07), and the sex × dose interaction was also not significant (p = 0.09). In operationally defined obese mice, glycemia increased progressively with iron dose, with the highest values observed at 5 mg/kg. A similar dose-dependent trend was observed in healthy mice, with glucose levels increasing at higher iron doses, particularly at 4–5 mg/kg. Effect sizes indicated a large effect of dose (η2p = 0.48), a small-to-moderate effect of metabolic status (η2p = 0.07), and a negligible Group × Dose interaction (η2p = 0.01). Observed power was high for the dose effect (≈1.00), moderate for metabolic status (≈0.65), and very low for the interaction (<0.10), indicating limited statistical power to detect interaction effects.
For hematocrit (Table 1), a clear increase was observed with higher iron doses. One-way ANOVA demonstrated significant dose effects within both operationally defined obese mice and healthy mice. The global two-way ANOVA (Group × Dose) confirmed significant main effects of iron dose and metabolic status, whereas the interaction term was small. Effect size analysis demonstrated large effects for both Group (η2p = 0.425) and Dose (η2p = 0.433), while the Group × Dose interaction was small (η2p = 0.037). Operationally defined obese mice consistently displayed higher hematocrit values than healthy mice across iron doses, with the highest values observed at 5 mg/kg. Post hoc comparisons indicated that, in operationally defined obese mice, hematocrit increased significantly from baseline (0 mg/kg) to all higher doses, whereas in healthy mice significant increases were mainly observed at 4 and 5 mg/kg.
Blood glucose levels also increased with iron dose (Table 2). Within-group analyses revealed significant dose effects in both operationally defined obese mice and healthy mice. The global two-way ANOVA confirmed a strong main effect of iron dose (η2p = 0.48) and a smaller but significant effect of metabolic status (η2p = 0.07), while the Group × Dose interaction was negligible (η2p = 0.01). Unlike hematocrit, sex differences were not detected at any iron dose. In both operationally defined obese mice and healthy animals, glucose levels tended to rise progressively with increasing iron intake, with the highest concentrations observed at 5 mg/kg.
Before iron supplementation, glycemia and hematocrit were not correlated (r = –0.008; p > 0.05). After iron supplementation, hematocrit and glycemia were positively correlated (Figure 1). The overall correlation coefficient was r = 0.62, indicating a moderate-to-strong association. Subgroup analyses showed stronger correlations in metabolically altered (operationally defined obese) mice (r = 0.74) and in females (r = 0.72). Linear regression analyses confirmed significant associations in both females (β = 0.28, p < 0.001) and males (β = 0.15, p < 0.001). Sex-stratified analyses showed stronger associations in females than males, with a modest but statistically significant difference in slopes (p = 0.027). Correlation between blood glucose and hematocrit values in healthy and operationally defined obese male and female mice after 30 days of iron supplementation. A positive linear association between glycemia and hematocrit was observed (overall r = 0.62; p < 0.01). Linear regression analyses showed significant relationships in both females (y = 0.28x + 31.71; r = 0.72; p < 0.01) and males (y = 0.15x + 44.66; r = 0.53; p < 0.01). Comparison of regression slopes using an interaction model (Sex × Glycemia) indicated a modest but statistically significant difference between sexes, with a steeper slope in females. Similarly, operationally defined obese mice showed a stronger correlation (y = 0.20x + 41.50; r = 0.74; p < 0.01) than healthy mice (y = 0.15x + 43.17; r = 0.47; p < 0.01), indicating that metabolic status modulates the glycemia–hematocrit association.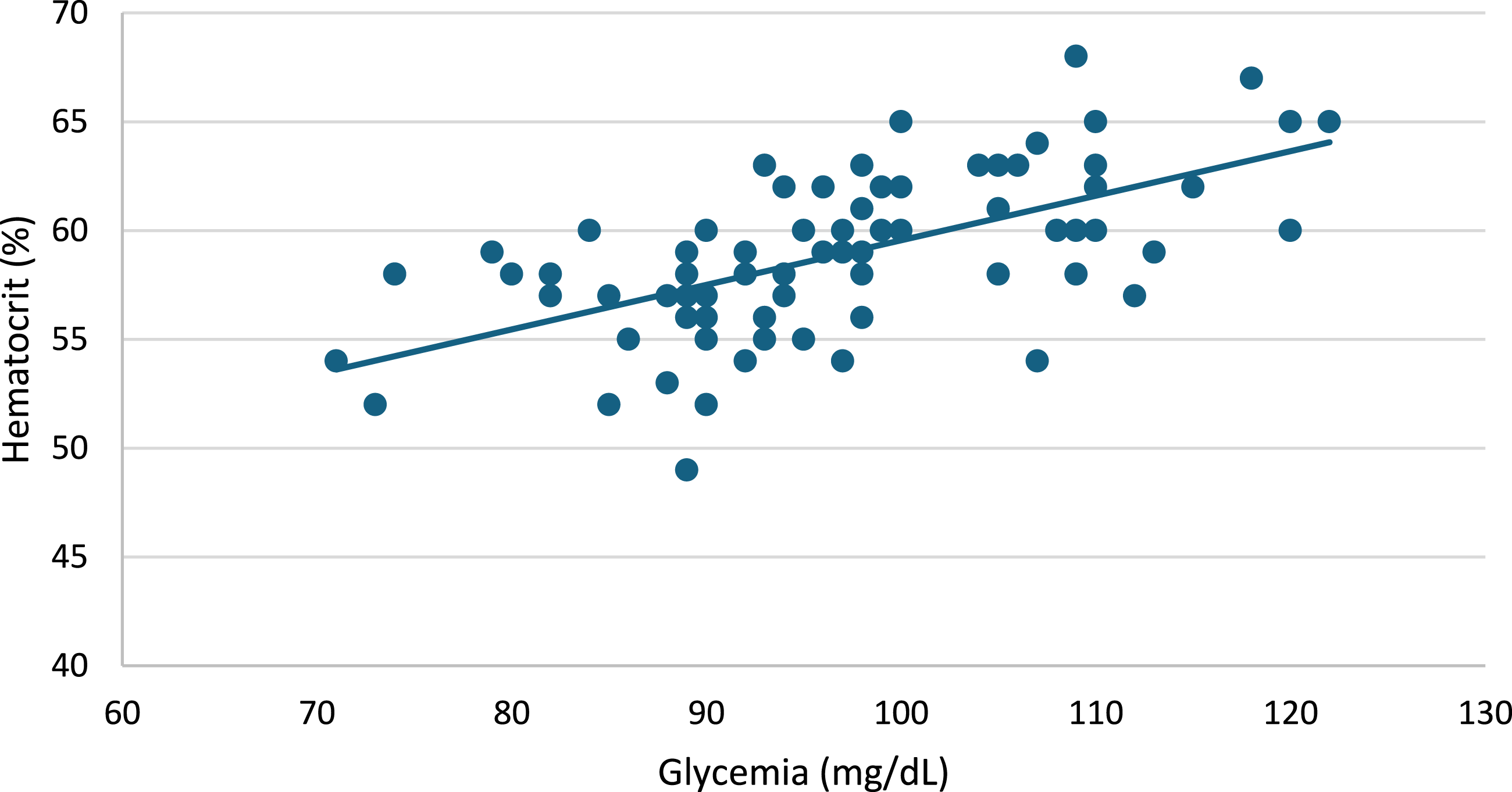
Discussion
This study demonstrates that sustained high-end nutritional iron exposure is associated with dose-dependent increases in hematocrit and glycemia, with a stronger hematologic response observed in metabolically altered (operationally defined obese) mice. In addition, iron supplementation strengthened the association between glycemia and hematocrit, with more pronounced relationships observed in metabolically altered animals and in females.
Although the association between glycemia and hematocrit appeared strengthened following iron supplementation, this relationship should be interpreted with caution. The observed correlations (including the overall and subgroup-specific coefficients) do not establish a direct mechanistic link between glucose metabolism and erythropoietic response. Rather, they may reflect parallel physiological processes influenced by shared metabolic or inflammatory pathways. Therefore, while these findings support a potential interaction between iron status and metabolic regulation, they should not be construed as evidence of causality, and further mechanistic studies are required to clarify the underlying biological relationships.
The experimental model used in this study is supported by prior evidence demonstrating that BALB/c mice provide a stable background for metabolic and hematologic research, allowing the assessment of graded physiological responses without the pronounced obesity phenotype observed in other strains.12–15 The induction of metabolic heterogeneity through dietary manipulation has been widely used to explore interactions between nutritional factors and systemic metabolism.16–19 Within this experimental setting, the present design enables evaluation of iron dose–response relationships under controlled conditions, while minimizing confounding effects related to diet composition or environmental variability. In addition, the methodological approach, including standardized housing, controlled feeding, and peripheral blood sampling, aligns with established experimental protocols and ethical guidelines for translational metabolic research. 20
In our experimental study, iron exposure was designed to approximate the upper physiological range of nutritional intake rather than pharmacologic or toxic dosing. When adjusted using allometric scaling, the highest dose corresponds to approximately 28 mg/day in a 70-kg adult, a level near the upper range of habitual intake but below established safety thresholds. 21 This distinction is essential, as chronic high-end nutritional intake differs substantially from acute pharmacological iron loading in terms of absorption dynamics and systemic regulation.22,23
Excessive iron intake refers to iron consumption exceeding physiological requirements and approaching or surpassing established nutritional thresholds. In humans, the Recommended Dietary Allowances (RDA) are 8 mg/day for adult men and postmenopausal women, 18 mg/day for premenopausal women, and 27 mg/day during pregnancy. The Tolerable Upper Intake Level (UL) set by the NIH/Institute of Medicine is 45 mg/day for adults, above which gastrointestinal, oxidative, and metabolic adverse effects are more likely. 24
It is important to distinguish between chronic high-end nutritional iron intake and acute pharmacological iron loading when interpreting the thresholds proposed by NIH/IOM. While values such as 45 mg/day represent the tolerable upper intake level for sustained dietary exposure, they are not directly comparable to short-term, higher-dose pharmacological interventions used in clinical or experimental settings. These two contexts differ substantially in terms of absorption dynamics, regulatory responses, and systemic iron handling. Failure to differentiate them may lead to misinterpretation in translational scaling, particularly when extrapolating findings from experimental models to human populations.
Although the European Food Safety Authority (EFSA) has not established a formal UL due to high interindividual variability, chronic high intake is recognized to increase the risk of systemic iron accumulation in susceptible individuals, with a proposed safe intake of 40 mg/day. 25
The observation that high-end nutritional iron exposure, without exceeding established tolerable upper intake levels, was associated with increases in hematocrit and strengthened glycemia–hematocrit coupling suggests that metabolically relevant alterations may occur even within nutritionally relevant exposure ranges. Experimental and clinical evidence has linked elevated iron stores with insulin resistance, obesity, and metabolic syndrome through pathways involving oxidative stress, mitochondrial dysfunction, altered glucose metabolism, and activation of hypoxia-inducible factor 1 alpha (HIF-1α) signaling.22,23,26,27
Iron excess has also been shown to activate an intestinal HIF-2α–dependent hyperabsorptive response regulated by the hepcidin–ferroportin axis, generating a hepatic–intestinal feedback loop that promotes systemic iron accumulation. 28 In this context, sustained iron exposure may partially override normal hepcidin-mediated regulation, particularly in metabolically altered states characterized by low-grade inflammation or altered iron trafficking. Even modest increases in ferritin and hemoglobin within reference ranges have been associated with oxidative stress and alterations in insulin signaling, 29 providing a plausible explanation for the strengthened glycemia–hematocrit coupling observed after iron supplementation.
These findings support the concept that iron availability is a key determinant of erythropoiesis. Persistent exposure to high nutritional iron may promote incremental increases in hematocrit, particularly in metabolically susceptible states. The amplified hematologic response observed in operationally defined obese mice is consistent with human evidence showing that obesity disrupts iron homeostasis, often leading to elevated ferritin, soluble transferrin receptor (sTfR), and hepcidin levels. 30
Conventional clinical thresholds for iron overload (e.g., transferrin saturation >45% and ferritin >200 ng/mL in women or >300 ng/mL in men) are primarily intended to identify overt iron excess. However, such cutoffs may not adequately distinguish inflammatory hyperferritinemia from true iron overload, nor do they capture metabolically relevant intermediate states.
This limitation underscores the importance of contextual and combined biomarker interpretation. These obesity-associated disturbances in iron homeostasis provide a plausible biological basis for understanding the heightened hematologic responsiveness to iron supplementation observed in operationally defined obese mice in our study.
To contextualize these findings within human physiology, observational data from children living in the highlands of Puno provide complementary evidence. In this population, total body iron increased significantly with higher dietary ascorbic acid intake despite similar iron consumption across groups, indicating enhanced intestinal iron absorption.
31
Using previously published data from children living in the highlands of Puno, we illustrate the relationship between dietary ascorbic acid intake and total body iron (y = 8.7152ln(x) - 1.9785. R2 = 0.9519; p<0.001. Figure 2). These findings reinforce the concept that iron bioavailability, rather than absolute intake alone, is a critical determinant of systemic iron status. Total body iron (mg/kg) in children aged 6–59 months from Puno according to levels of dietary ascorbic acid intake. Total body iron increased with higher ascorbic acid intake despite similar dietary iron consumption across groups, suggesting enhanced intestinal iron absorption. These previously published observational data were adapted from Ref. 31 and are presented solely for contextual interpretation; they are not part of the current experimental study or dataset. Source: Authors’ own elaboration based on data reported by Choque-Quispe et al. (Reference 31), published under the Creative Commons Attribution 4.0 License (CC BY 4.0).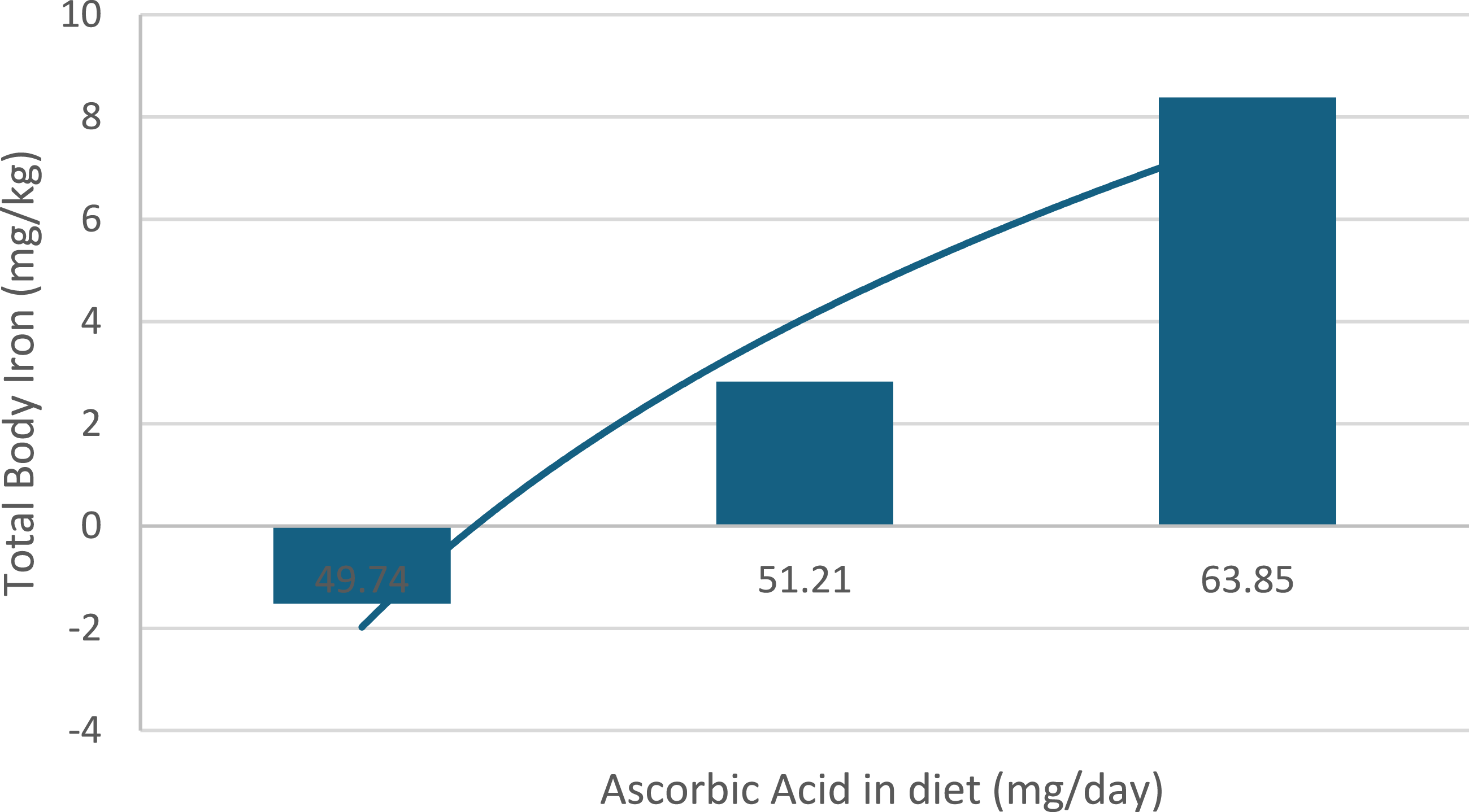
Oral iron supplementation, although widely implemented, is not without biological cost. Even in healthy individuals, excessive intake can induce the formation of non–transferrin-bound iron (NTBI), a reactive species linked to oxidative and vascular toxicity.32,33
These hemoglobin extremes persist across altitudes, linking observations from low-altitude conditions such as preeclampsia with those described in high-altitude populations. At low altitude, both low and elevated hemoglobin levels have been associated with adverse maternal and fetal outcomes, reflecting disturbances in oxygen delivery and vascular adaptation. 34 Similarly, at high altitude, excessive erythrocytosis and suboptimal hemoglobin levels may represent maladaptive responses to hypoxia rather than purely physiological adaptation.35,36 This continuity across altitudes supports the concept that hemoglobin extremes, regardless of environmental context, may signal underlying pathophysiological processes rather than optimal oxygen transport. 37
The updated 2024 altitude adjustment proposed by the World Health Organization introduces a modest hemoglobin correction of −0.4 g/dL at 500–999 m; however, its physiological relevance at these altitudes remains limited. Arterial oxygen saturation is largely preserved within this range, and the hypoxic drive for erythropoiesis is minimal, suggesting that the magnitude of this adjustment may not reflect a biologically meaningful change in oxygen transport, and may instead introduce classification bias in populations residing at low-to-moderate altitudes.3,6
Consequently, applying this correction may disproportionately influence anemia classification without corresponding alterations in tissue oxygenation. This distinction is critical, as even small numerical shifts in hemoglobin thresholds can substantially affect population-level prevalence estimates, potentially reinforcing supplementation strategies in individuals without true physiological need.
The temporal dimension further modulates iron metabolism. Hepcidin exhibits circadian variation, with lower levels in the morning favoring increased absorption.38–40 Ignoring these chronobiological patterns in supplementation strategies may contribute to gradual iron accumulation over time.
Ferritin, transferrin saturation, hepcidin, and tissue iron quantification were not assessed in this study. Therefore, the observed hematological changes cannot be directly attributed to confirmed systemic iron overload and may instead reflect adaptive erythropoietic responses or functional alterations in iron utilization. Consequently, these findings should be interpreted as indicative of iron-responsive physiological effects rather than definitive evidence of iron overload. Future studies incorporating comprehensive iron biomarkers will be necessary to distinguish between true iron excess, redistribution, and erythropoietic stimulation. Importantly, this limitation does not compromise the internal validity of the observed dose–response relationships.
Finally, chronic excess iron may have significant metabolic consequences. Sustained hyperferritinemia has been implicated in the progression from simple steatosis to metabolic dysfunction–associated steatohepatitis (MASH), promoting oxidative stress, inflammation, fibrosis, and ferroptosis. 41
Together, these findings challenge the prevailing paradigm that higher iron exposure is uniformly beneficial and instead support a paradigm in which iron status must be tightly regulated within an optimal physiological range to avoid both deficiency- and excess-related risks.
Conclusion
These findings indicate that higher iron exposure is not invariably associated with improved health outcomes and that elevated hemoglobin concentrations may reflect maladaptive rather than beneficial responses. Our results, together with evidence from previous studies, suggest the possibility of a U-shaped relationship in which both iron deficiency and excess may be associated with increased risk. This proposed model should be interpreted as a hypothesis requiring confirmation in further experimental and clinical studies. Considering recent revisions by the World Health Organization, these observations underscore the need to reassess current approaches to anemia prevention and iron supplementation. Future strategies should adopt a precision-based approach incorporating validated diagnostic markers, metabolic determinants, and physiological regulation, with the aim of optimizing benefit while minimizing unintended iron excess in non-deficient populations and balancing the risks of deficiency and excess across diverse populations and environmental settings.
Footnotes
The authors acknowledge Dr. Manuel Gasco for training Jeannett Rivera in animal handling and animal use procedures.
Ethical considerations
All animal procedures were approved by the Ethics Committee for the Use of Animals of the Universidad Peruana Cayetano Heredia on 4 January 2023 (approval code: DC: 001-01-2023) and were conducted in accordance with ARRIVE 2.0 guidelines.
Author contributions
Funding
The authors received no financial support for the research, authorship, and/or publication of this article: This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Declaration of conflicting interests
The authors declare no conflicts of interest in relation to this manuscript.
Data Availability Statement
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Use of AI tools
The authors used ChatGPT (OpenAI) solely for language editing and improvement of readability. All scientific content, interpretation, and final manuscript preparation were performed by the authors.