
Abstract
The effect of the coupling approach (chemical by using carbodiimide chemistry and grafting-to versus grafting-from synthesis routes, and enzymatic by using transglutaminase) of ɛ-poly-L-lysine (ɛPL) graft yield with wool fibers was studied and evaluated related to their antibacterial activity against gram-negative Escherichia coli and gram-positive Staphylococcus aureus bacteria, after 1–24 h of exposure. While the ɛPL applied was evaluated colorimetrically by wool staining with two acid dyes and quantitatively by evaluation of the basic groups using potentiometric titration, its orientation was determined by Fourier transform infrared spectroscopy and, for the first time, by Electron Paramagnetic Resonance spectroscopy using spin-labeled ɛPL.
The highest (∼99% for E. coli versus ∼92% for S. aureus) and kinetically the fastest (in 3 h) antibacterial activity with ∼83% for E. coli versus ∼64% for S. aureus bactericidal effect was determined for the wool functionalized by the chemical grafting-to approach. Such an effect may be related to both quantitatively the highest (∼62 gɛPL/kgwool) grafting yield of ɛPL and conformationally its highly flexible “brush-like” structure. Comparably, the enzymatic coupling (∼50 gɛPL/kgwool) giving ∼95% and ∼8% reductions of E. coli and S. aureus, respectively, being additionally reduced to ∼74% and ∼78% by using the grafting-from approach (∼34 gɛPL/kgwool), was identified as the less bactericidally effective (∼63% versus ∼58%).
It was also shown that a non-ionic surfactant being used in the durability testing of functionalized wool to washing adheres strongly onto the fibers, thus blocking the amino groups of ɛPL, and, as such, decreases the antibacterial efficiency of the wool, being unaffected in the case when the washing was carried out without surfactant.
Lightness, warmth, softness, and smoothness are the properties that make wool fibers generally used for high-grade textiles. Over the last decade wool keratins have also been studied as potential biomaterials for human clinical applications as wound dressings and implantable devices, being related to both contents of proteins having cell adhesion sequences1,2 and thus being good substrates for fibroblast3,4 and osteoblast cells, 5 as well as being biodegradable in vitro and in vivo. 6 However, as a protein fabric, wool suffers degradation easily, causing skin irritation and/or infection due to the generation and propagation of microorganisms under certain temperatures and humidity. 7 In addition, being used as a biomaterial it has to meet a wide variety of other demands, above all possessing relevant bioactive 8 and antimicrobial activities employing quaternary ammonium salts,9,10 cationic and amphiphilic polymers, 11 and colloidal nanoparticles, such as, for example, SiO2, 12 Cu, and Ag, and using different technologies on nanofunctionalized surfaces.13,14 A range of crosslinking reagents and coupling strategies have been investigated for enabling their covalent attachments; 15 non-covalently bound (mainly embedded in coatings) or encapsulated metal nanoparticles are still the most important subject studies, although they have shown some weaknesses, such as capped long-term effectiveness and the capacity for developing microbial resistance and low efficiency. On the other hand, increasingly applicable antimicrobial active nanoparticles can penetrate through the skin and/or cell membranes and cause vague threats to our health. 16 In addition, as many heavy metals are applied during the coating of fibers via ultrasound irradiation, such treatments have serious limitations due to technical and environmental problems and, therefore, have not been adopted for commercial production.11,17
The desired level of protection is thus to develop materials that would ensure not only effective and long-lasting antimicrobial protection against a wide range of pathogenic bacteria and fungi, but also to maintain a constant and well-defined biological environment. Materials should, therefore, reflect both antimicrobial activity and tissue biocompatibility. Antimicrobially active peptides and proteins are thus being recognized as perspective antimicrobial agents, showing high and specific antimicrobial activities. 18 Amongst them, ɛ-poly-L-lysine (ɛPL) is being used within different fields, including medicine, food, environment, and agriculture, as well as the textile industry, due to its low minimum inhibitory concentration (MIC of < 0.05%) and non-toxic nature. 19 However, amongst the different functionalization procedures that have been studied for the immobilization of ɛPL on various types of fibers and polymers, most of them are non-specific and based on electrostatic interactions between positively charged amino groups of ɛPL and negatively charged polymer surfaces. 20 Stable chemical bonding between lysozyme and wool was also proposed through the crosslinking of lysozymes’ amino groups with available lysine residues within wool fibers 21 by using expensive and toxic glutaraldehyde that, however, may reduce the biocompatibility of the final material. Alternatively, using enzymes working site-specifically and regional-selectively, for example microbial transglutaminase (mTGase; which catalyzes the acyl transfer reaction between the peptide-bound glutamine residues as acyl donors and primary amino groups as receptors), have been shown to be a very useful grafting approach, 22 resulting in a material with ∼96% reduction of E. coli. 23
The aim of this study was thus to define a strategy for the covalent coupling of ɛPL (with the structural formula shown in Figure 1) to the wool whilst maintaining their antibacterial activity. In that respect different coupling approaches were selected and compared for achieving stable and durable functionalization. The grafting-to and grafting-from synthesis routes were applied by using well-known carbodiimide chemistry, and the enzyme-catalyzed approach using a transglutaminase (TGase). Various direct (potentiometric titration) and indirect (colorimetric) analytical methods were used for quantitative determination of the ɛPL applied to the wool, while its orientation was characterized by attenuated total reflectance-Fourier transform infrared (ATR-FTIR) and Electron Paramagnetic Resonance (EPR) spectroscopies using spin-labeled ɛPL. The viability of gram-negative Escherichia coli and gram-positive Staphylococcus aureus bacteria after 1–24 h of incubation with differently treated wool was evaluated for assessing the biocidal versus bacteriostatic effect.
Chemical formula of ɛ-poly-L-lysine.
Experimental details
Materials
The knitted wool fabric used was supplied by Lokateks d.o.o. (Slovenia) and was a fine rib 1:1 knit with a mean fiber diameter of 23.5 µm and mass/unit area of 230 g/m2. ɛPL with a molecular weight of 9.1 kDa (being previously determined by size-exclusion chromatography-high-performance liquid chromatography (SEC-HPLC) using an Agilent BioSEC-3 column with a diameter of 4.6 mm, particle size of 3, µm and pore diameter of 15 nm) was purchased from Zhengzhou Bainafo Bioengineering, China. The EDC (1-ethyl-3 -(3-dimethylaminopropyl)-1-carbodiimide hydrochloride) and sulfo-NHS (sulfo-N-hydroxysuccinimide) were obtained from Sigma-Aldrich (Germany) and used as received.
Tissue TGase from a guinea-pig’s liver of 3.1 U/mg was supplied by Sigma-Aldrich (Germany). Spin label 3-maleimido-2,2,5,5-tetramethyl-1-pyrrolidinyl-oxyl (3-maleimido-PROXYL) was purchased from Sigma-Aldrich (Germany). The Acid Orange 7 (AO7) and Coomassie Brilliant Blue G–250 dyes used for colorimetric quantification of the graft yield were purchased from the Sigma Chemicals Co. Gram-negative Escherichia coli (ATCC 35218) and gram-positive Staphylococcus aureus (ATCC 25923) were provided by the American Type Culture Collection (ATCC). A Live/Dead BacLight kit from Sigma-Aldrich (Germany), consisting of a SYTO 9 dye and Propidium Iodide (PI), was used for bacterial viability testing. Non-ionic surfactant Dehaclin WP 20 was purchased from CHT/Bezema, Germany. Dichloromethane, ethanol, methanol, octanol, and other solvents were of analytical grade and used without further purification.
The water used in all experiments was purified using a Milli-Q direct water purification system (Millipore Direct 8) with a resistivity of 18.2 MΩ cm. Spectra/Por dialysis membranes with a Molecular Weight Cut Off (MWCO) of 2 kDa from regenerated cellulose were acquired from Fisher. Other chemicals used in this work were all of chemical grade without additional purification.
Methods
Pre-treatment of wool samples
In order to improve the availabilities of functional groups for covalent attachment of ɛPL, the wool was extracted sequentially with dichloromethane according to the ISO 3074:2014 standard, followed by rinsing with Milli-Q water and dried at 40℃ to remove surface lipids and contaminants. Such wool was marked as a reference sample (Ref-WO) and was used in all further treatments.
Functionalization of wool with ɛPL
Wool is a proteinaceous fiber with an isoelectric point (pI) at a pH of ∼4.9 (confirmed by potentiometric titration) that offers many reactive sites for its functionalization with ɛPL, as schematically presented in Figure 2. According to the chemical approach, two different coupling routes were applied, being generally available in the synthesis of peptide-polymer conjugates using water-soluble EDC and N-hydroxysuccinimide (NHS) as zero-length and site-specific reaction mediators.24,25 The carboxyl groups (-COOH) in the C-terminal of the wool (grafting-from) or ɛPL (grafting-to) were converted into an unstable amino-reactive intermediate (O-acylisourea) that reacted readily with the available and non-protonated amino (-NH2) groups of ɛPL or wool (R-NH2 of lysine or arginine residues), respectively, thus forming a stable amide linkage.
26
However, it has to be considered that the N-terminal ɛ-amino group of ɛPL has a higher pKa (∼10.5) than the numerous sided α-amino groups (pKa ∼9.0). This renders the lysine amino side groups effectively more nucleophilic and thus highly reactive in their reactions.
Schematic illustration of three routes for ɛ-poly-L-lysine (ɛPL) coupling onto the wool protein using both the chemical (grafting-to and grafting-from) and enzymatic (transglutaminase (TGase)) approaches. EDC: 1-ethyl-3 -(3-dimethylaminopropyl)-1-carbodiimide hydrochloride; NHS: N-hydroxysuccinimide.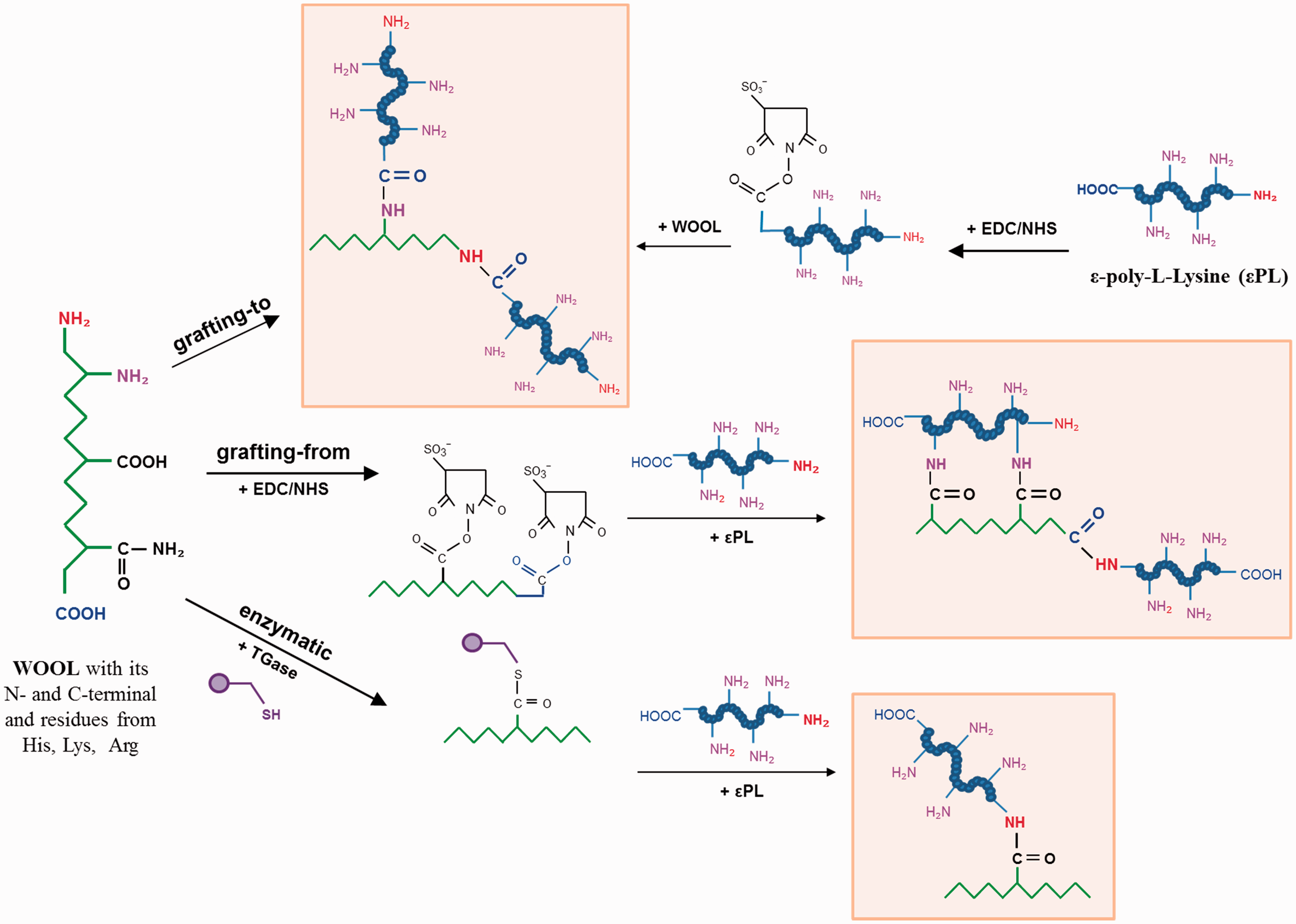
In the case of the grafting-from approach, 1 g of wool was thus activated with EDC/NHS at a molar ratio of 10:10:1 with respect to the available (i.e. dissociated) wool carboxyl groups (∼585 mmol/kg wool as determined by potentiometric titration) in a 100 mM phosphate buffer of pH 4.5 containing 200 mM NaCl and 50% methanol for 24 h at 23℃ by constant shaking. The fabrics were then added to an equimolar amount (5 g/L) of ɛPL with respect to its amino groups, previously dissolved in 100 mM carbonate/bicarbonate buffer of pH 10. In the other way around, that is, the grafting-to approach, 5 g/L of ɛPL was incubated with EDC/NHS at a molar ratio of 10:10:1 with respect to the ɛPL acidic groups (∼8.22 mmol/kg ɛPL, determined by potentiometric titration) in 100 mM phosphate buffer of pH 4.5 containing 200 mM NaCl for 2 h at 23℃ by constant shaking. The solution with an activated terminal carboxylic group of ɛPL was then incubated with 1 g of wool (with respect to an equimolar amount of its basic groups determined by potentiometric titration), previously soaked in 100 mM carbonate buffer of pH 10.5. The incubation was carried out for 24 h at 23℃ by constant shaking. In both cases the samples were post-treated by rinsing them in a low pH (pH 2) and high salt concentration (2 M NaCl) to remove residual (non-reacted or ionically adsorbed) ɛPL, as well as isourea as a by-product being originated during the chemical reaction.
The enzymatic approach of ɛPL grafting was performed by TGase, which catalyzed the reactions between the ɛ-amino groups of ɛPL and the glutamine residues of the wool in a two-step reaction, resulting in the formations of new isopeptide bonds 27 ; in the initial step the active site of TGase cysteine binds to the γ-carboxyamide of the glutamine residue by a thioester bond, forming an acyl-enzyme intermediate that interacts forward with the nucleophilic ɛ-amino group of the lysine residue, accelerating the release of ammonia. For that purpose, 1 g of wool was incubated with 5 g/L of ɛPL and 2 U of TGase in 100 mM phosphate buffer of pH 7.5 containing 5 mM CaCl2 for 24 h at 50℃. The reaction was stopped by heating the samples for 5 min at 95℃ and post-treating according to the same procedure as described above to remove residual (non-reacted or ionically adsorbed) ɛPL as well as the ammonia by-product.
In all types of treatments, parallel experiments were conducted being denoted as reference wools, treated in the same way as other samples but with the reagents (Ref-EDC/NHS) or enzyme (Ref-TGase) only and in the absence of ɛPL. In addition, the wool was treated with ɛPL but without the reagents or enzyme, being denoted as Ref-ɛPL.
Colorimetric evaluation of wool by its staining with anionic dyes Coomassie Brilliant Blue G–250 and Acid Orange II, and indirect quantification of (all) basic groups
The ɛPL deposition on the wool was evaluated directly (colorimetric evaluation of the wool) and indirectly (dye concentration evaluation in the treating solutions) after its staining with two acids dyes (Coomassie Brilliant Blue G–250 and AO7) being related to Van der Waals attractions and ionic interactions between the dye’s sulfonate groups (–SO3–) and the available weak base protonated groups (–NH3+) of wool protein (arising from side groups of histidine, lysine, arginine and the α-NH2 groups at the chain ends) 28 in acidic media.
The staining of the wool with an already prepared pre-mixed solution of Coomassie Brilliant Blue G–250 with unknown concentration and pH ∼3 was performed at 50℃ for 30 min using a liquor-to-fabric ratio of 15:1, followed by washing with distilled water for two cycles and then drying at room temperature. 29 The reflectance (R) of the dyed fabrics was measured using an SF 600+ (Datacolor) Spectrophotometer over a wavelength range of 360–700 nm. The relative color depths of the stained samples, expressed as K/S, were established according to the Kubelka–Munk equation, K/S = (1 – R) 2 / 2 R, 30 where K and S are the absorption and scattering coefficients, respectively.
The staining of wool samples with 1% of AO7 dye solution was performed at pH ∼2.2 (being adjusted with 0.05 M sodium acetate and acetic acid, to ensure that the amino groups of ɛPL and wool were fully protonated) and 50℃ for 30 min using a liquor-to-fabric ratio of 50:1. The remaining solution was analyzed spectrometrically at 486 nm to determine the amount of the dye in the solution by calculating the dye concentration based on a standard curve made of dye in the acetic acid. By subtracting the remainder from the initial dye concentration, the amount of dye adsorbed onto the wool was obtained. Data shown are the means of four experiments performed in duplicate, containing the standard deviation. In parallel, the color depth of stained fabrics, expressed as K/S, were established as described under the previous section.
Quantification of (some) basic groups on wool by potentiometric titration
Potentiometric titrations of wool samples, before and after chemical modification with ɛPL, were performed using a dual-burette instrument (Mettler Toledo T-70) equipped with a combined glass electrode (Mettler TDG 117). The burettes were filled with 0.1 M HCl (Merck, Titrisol) and 0.1 M KOH (Baker, Dilut-it). Samples were rinsed in low pH (0.01 M HCl) to convert polypeptide basic and acidic residues into a protonated form, soaked in Milli-Q water and, afterwards, dried at 40℃. Titration was carried out at room temperature (23 ± 1℃) in forward and back runs between pH 3 and pH 11. From the potentiometric titration data, the molar concentration relating to the overall charge of the weak ions (arising from side groups of histidine and lysine, and the α-NH2 groups at the chain ends) was calculated, a method described in detail by Čakara et al. 31 All the reported values are the mean values of duplicate determinations, containing the standard deviation.
Attenuated total reflectance-Fourier transform infrared spectroscopic analysis of wool
FTIR spectroscopy was used to identify spectral differences associated with the polymer orientation differences amongst the wool samples. For that purpose, a PerkinElmer IR Spectrometer was used with a Golden Gate ATR attached to a diamond crystal. The spectra were accumulated under ambient conditions from accumulating 16 scans at resolutions of 4 cm–1 within a region of 4000–650 cm–1, with air spectrum subtraction performed in parallel as a background. The PerkinElmer Spectrum 5.0.2 software program was applied for the data acquisition analysis. A linear baseline was subtracted and the resulting spectra were normalized before determining the differences between the secondary structures of the samples by using Fourier self-deconvolution of the amide I band components (1700–1570 cm−1), processed by using Peakfit 4.12 software (Galactic Industries Corporation, USA). The amide I band was resolved by the second-order derivative with respect to the wavelength. In addition, the spectra were evaluated quantitatively by calculating the intensity ratio between the amide I and amide II bands (Aamide I/Aamide II).
Electron Paramagnetic Resonance spectroscopic analysis of wool using spin-labeled ɛPL
Site-directed spin-labeling of ɛPL amino groups with 3-maleimido proxyl (MP) was performed at room temperature for 24 h in 100 mM phosphate buffer containing 200 mM of NaCl (pH 8) at a ratio of spin label to ɛPL amino groups of 1:10 (mol/mol). The free spin label was removed by dialysis at room temperature against a 100-fold excess of 200 mM NaCl using the Spectra/Por membrane of MWCO 2 kDa. The dialysis buffer was changed four times every 12 h.
The wool pieces were crushed into a powder to be homogenized for EPR analysis. The samples were put into quartz capillary tubes of 1 mm in diameter and transferred into the EPR resonator for analysis. X-band (9.3 GHz) continuous-wave Electron Paramagnetic Resonance (cw-EPR) spectra were recorded at room temperature using a Bruker EleXsys E500 Spectrometer (Germany) equipped with a high-sensitivity resonator (SHQEW0401) applying a scan range of 100 G, 1.5 G amplitude of modulation, 41.94 s of sweep time, 40.96 ms of time constant, and microwave power of 20 mV. Differently immobilized ɛPL on wool was expected to bring a local constriction that would restrict the possible motions of the ɛPL and their associated spin labels. These changes in motion result in changes in the determined rotational correlation times (
Antibacterial activity determination
The antibacterial efficacies of wool were assessed under dynamic contact conditions by using the dynamic shake flask test method ASTM E2149-10. 32 Stock solutions of selected bacteria (gram-negative E. coli ATCC 35218 and gram-positive S. aureus ATCC 25923) were streaked on Petri dishes and cultured for 24 h at 37℃, before fully grown single colonies were inoculated in sterilized nutrient broth and cultured for 24 h. The incubated culture solutions were diluted using a sterilized physiological saline (0.9% NaCl) in order to give a final concentration of 1.5–3.0 × 105 colony-forming units (CFU)/mL being used as a working bacterial solution. Each sample (∼0.1 g) was transferred to a sterile 150 mL Erlenmeyer flask containing 10 mL of the working bacterial solution. Control experiments were run during which no wool samples were present within the bacteria culture and treated in the same way. All flasks were capped loosely and incubated for 1, 3, 6, and 20 h, respectively, on a rotary shaker (120 min–1) at 37℃. At different contact periods, 1 mL of the bacterial suspension was withdrawn, serial diluted using the physiological saline, transferred onto the prepared Petri dish, and spread over the nutrient agar using a hockey stick. The inoculated plates were incubated at 37℃ for 24 h and the surviving cells counted. The values were converted to CFU/mL in the flasks by multiplying by the dilution factor, and the mean values of percent reduction were calculated using the following equation: reduction (R) = (A – B) / A × 100, where A and B are the numbers of bacteria colonies on the samples before and after the shake flask test, respectively. Experiments were performed in duplicate and the data are shown as the mean values, containing the standard deviation.
Bacterial viability and wool bactericidal assessment using the Live-Dead test
The working suspensions of both bacteria, E. coli (2 × 108 bacteria/mL) and S. aureus (2 x 107 bacteria/mL) were prepared as reported. 33 After 24 h of inoculating wool substrates with bacteria cells using working bacterial suspensions, a stock solution (1.5 mL) of Live/Dead BacLight staining reagent mixture (6 µM SYTO 9 and 30 µM PI) was mixed with 1.5 mL of each bacterial suspension, which remained after shake flask testing. The solutions were incubated at room temperature in the dark for 15 min and then each pipetted (100 µL) into separate wells of 96-well flat-bottomed micro-plates and measured using the fluorescence mission spectrum using a Tecan Spectrophotometer. The viable bacteria (live cells) were stained with SYTO 9 to a green fluorescent color, whereas the bacteria with damaged membranes (dead cells) were stained with PI to a red fluorescent color. By using an excitation wavelength centered at 480 nm, the integrated intensities of the green (G; 530 nm) and red (R; 630 nm) emission were acquired, and the green/red fluorescence ratios (Ratio G/R) were calculated for each suspension. The biocidal effects (%) within 24 h against E. coli and S. aureus were determined by calculating the relationship between Live (G, x) bacteria and G/R fluorescence ratio (y) as [y1 = 0.0632x + 0.0614] for E. coli and [y2 = 0.0151x + 0.3369] for S. aureus.
The biocidal efficiencies of the wool samples were examined further by viability tests performed by using confocal microscopy and fluorescence spectroscopy. After 24 h of exposure to the media inoculated with E. coli (105 CFU/mL) or S. aureus (105 CFU/mL), the wool samples were stained with a 3.4 µM of SYTO 9 and 6.0 µM of PI, respectively, for 30 min in the dark at room temperature, washed with Milli-Q water, air-dried, and then visualized by fluorescence microscopy consisting of a Nikon TE2000-E inverted microscope, lambda LS Xe-Hg arc lamp, BD Carv II fluorescence, confocal unit, and Rolera MGi EMCCD camera. The images were excited using 420 or 488 nm excitation filters and the emissions collected using a UBG triple-band-pass emission filter cube. Experiments were performed in duplicate and the data are shown as the mean value, containing the standard deviation.
Durability testing of functionalized wool to washing
The durability of the wool functionalization activity was assessed by washing the samples within the beaker dyeing system Labomat (Werner Mathis) according to standard ISO 6330:2012. 34 For that purpose, the samples were submitted to five or 20 washing cycles at 40℃ for 30 min using the non-ionic surfactant Dehaclin WP 20 at a concentration of 1 g/L. In addition, the washing cycles were performed in the same way but without using the washing agent. The samples were then subjected to antibacterial testing using ASTM E2149-10, 32 as described above, in order to analyze the bacterial growth. Experiments were performed in duplicate and the data are shown as the mean value, containing the standard deviation.
Wool extraction
The extraction process was undertaken to evaluate the possible adsorption of non-ionic Dehaclin WP 20 surfactant onto differently processed woolen samples during the washing. The extraction was performed with dichloromethane as the extraction solvent, 35 followed by rinsing with Milli-Q water and drying at 40℃. The extracts were filtered and concentrated to dryness immediately by rotary evaporation. The extracts were maintained under vacuum overnight in a desiccator in order to eliminate moisture traces and, finally, weighed. The yielded dichloromethane extractable matter is given as a percentage of the dry mass of the extracted specimen, using the following formula: 100 × ·m1 / m2, where m1 (g) is the mass of the dichloromethane-soluble extract and m2 (g) is the dry mass of the extracted test specimen. Experiments were performed in duplicate and the data are shown as the mean value, containing the standard deviation.
Results and discussion
The influence of the coupling strategy of an ɛPL graft yield onto wool
Direct and indirect analytical methods were used in order to evaluate the coupling efficacy of ɛPL to wool. Direct methods were related to colorimetric evaluation of the wool stained with different acid dyes, as well as its quantification by potentiometric titration, while the indirect method gave us the information about the remaining AO7 dye content in the treatment bath after the staining.
The potentiometric titration curves (Figure 3) of the reference wool (Ref-WO) and differently ɛPL-treated wool using both chemical and enzymatic approaches were performed to observe the charge reversal in the case of wool functionalization. The pI of the wool functionalized with ɛPL using the grafting-to approach is the highest and occurs at pH ∼10, similar to the free ɛPL (pI ∼9.5, pre-determined by zeta potential; data not shown), while the pI of the Ref-WO appeared at pH of ∼4.9, which is in accordance with the previous results.2,36 The weak basic (arising from side groups of histidine and lysine residues, as well as the α-NH2 groups at the chain ends of the wool) group content (Table 1), being determined from the back-titration data, thus increased by ∼72% (to ∼ 1005 mmol/kg) and by ∼42% (to ∼ 833 mmol/kg) by the grafting-to and grafting-from approaches, respectively, compared to the Ref-WO, where ∼585 mmol/kg basic groups were evaluated. Furthermore, the quantity of these groups on the wool treated by the enzymatic approach increased by ∼ 55% (to ∼ 909 mmol/kg). The control sample treated without EDC/NHS (Ref-ɛPL) did not bring any increase of the basic group’s content (∼563 mmol/kg), which means that the ionic adsorption of ɛPL to the wool is negligible and that all the non-covalently bound ɛPL was removed successfully by its rinsing at very low pH (pH 2), at which the strong electrostatic interaction are disrupted.
37
On the other hand, a significant reduction of basic groups (∼172 mmol/kg) was detected for Ref-EDC/NHS, indicating the crosslinking between the available carboxylic and amino groups of wool protein residues
26
; a similar but much less pronounced effect can be observed for the Ref-TGase sample, also being well in agreement with other studies, which have shown that keratin can be self-crosslinked by microbial TGase, resulting in less lysine amino group availability.38,39
Titration curves of the reference and differently ɛ-poly-L-lysine-functionalized wool samples. TGase: transglutaminase. The coloration yield (K/S) and quantity of basic groups (mmol/kg wool) of the references and differently ɛ-poly-L-lysine (ɛPL)-functionalized wool samples, being evaluated through their staining with different acid dyes as well as by potentiometric titration TGase: transglutaminase; EDC: 1-ethyl-3 -(3-dimethylaminopropyl)-1-carbodiimide hydrochloride; NHS: N-hydroxysuccinimide.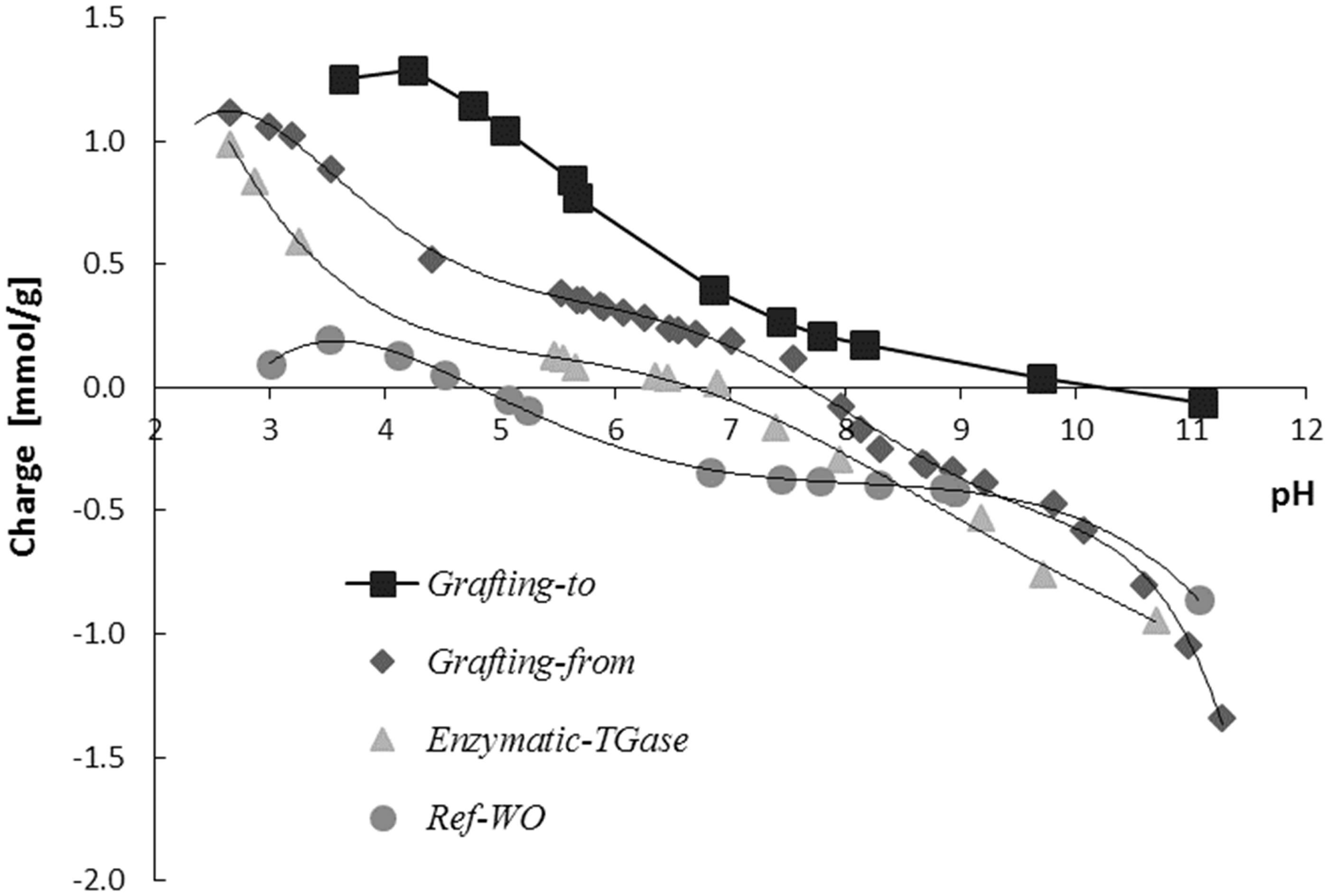
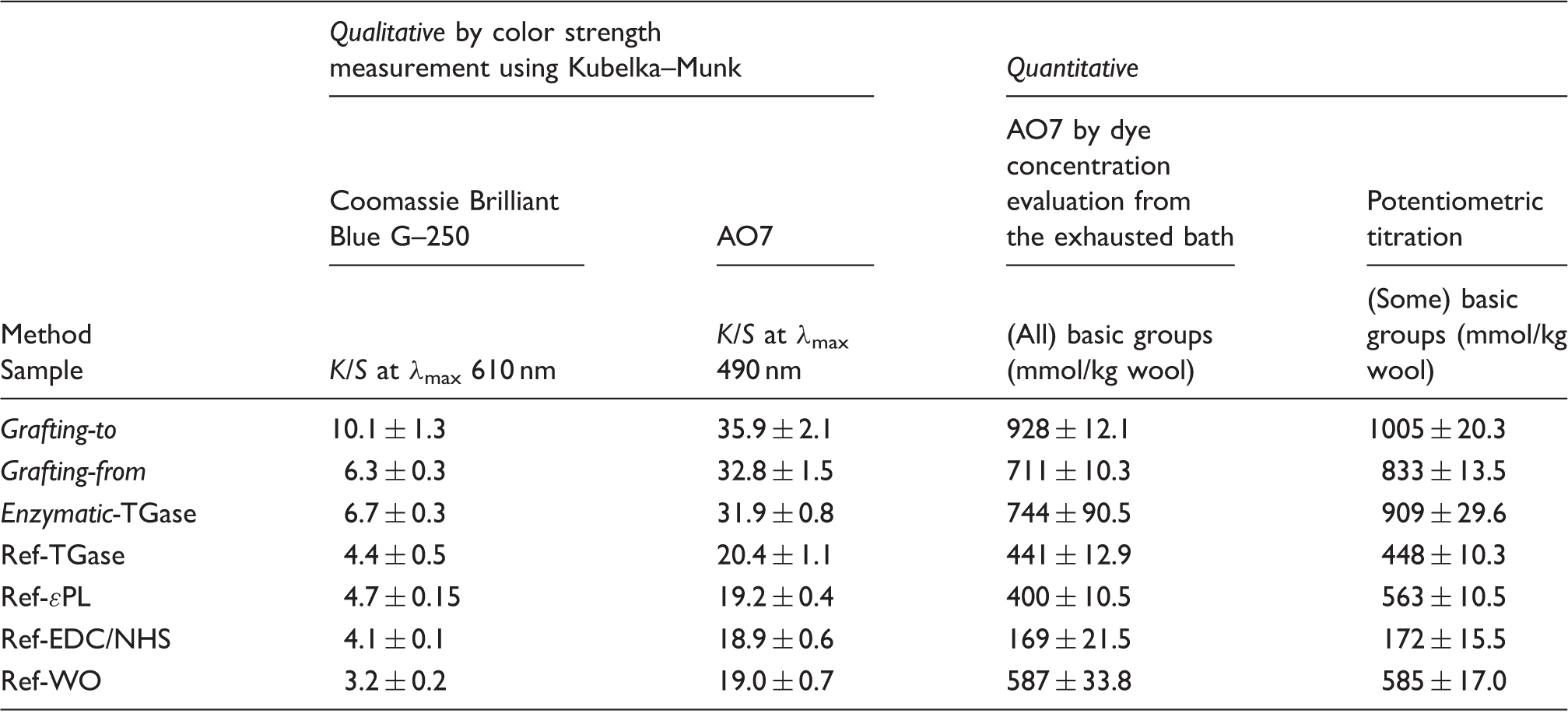
Indirectly quantified basic groups (arising from side groups of histidine, lysine, and arginine residues, as well as α-NH2 groups at the chain ends) of the wool (by determining the AO7 dye concentrations in the solutions of wool samples after their staining at very low pH, where the wool is fully protonated and thus available for ionic interactions with acid dye) showed a similar trend. As shown in Table 1, the quantity of basic groups present on the wool was the largest (∼58% higher) when using the EDC/NHS coupling mechanism and grafting-to approach (∼928 mmol/kg), compared to the reference (∼587 mmol/kg) as well as grafting-from (∼711 mmol/kg) and the TGase catalyzed coupling mechanism (∼744 mmol/kg). The basic group reduction was observed for all the reference samples, except for Ref-ɛPL, showing the group’s content of ∼400 mmol/kg being comparable to that of the Ref-WO. The values are well correlated to that from the potentiometric titration analysis.
On the other hand, direct evaluation of the ɛPL grafting yield by the staining of available amino groups on wool with both acid dyes (Coomassie Brilliant Blue G–250 and AO7, respectively) showed increased color strength (K/S) values for all ɛPL-treated wools (Table 1) as a result of an increased amount of dyes applied. 40 The EDC/NHS treatment resulted in more intensive coloration in the case of grafting-to (K/S of 10.1 and 35.9, respectively) compared to the grafting-from (K/S of 6.3 and 32.8, respectively) mechanism and the TGase approach (K/S of 6.7 and 31.9, respectively), being again significantly higher compared to all the references showing K/S values between 3.2 and 4.4 for dyeing with Coomassie Brilliant Blue G–250 and between 19 and 20.4 for AO7. The dyeability (K/S) of all AO7 dyed samples is related to much higher initial dye concentration compared to the Coomassie Brilliant Blue G–250, which is available only as a pre-mixed solution with unknown concentration.
It may be concluded that ɛPL immobilization on wool is achieved by all three coupling strategies, being, however, the highest by using EDC/NHS coupling chemistry and the grafting-to approach, promoting the basic group’s content of the wool–ɛPL conjugate. In addition, the resistance of ɛPL desorption upon the exposure of functionalized wools to a strong acidic solution under which all the ionic interactions that can cause non-specific attachment should be broken, indicating the specific attachment of the ɛPL to the wool surface via amide bond formation.
The orientation and localization of ɛPL on a wool surface
The ATR-FTIR spectra of the referenced and differently treated wool samples were used to evaluate the interactions of ɛPL within the wool protein. The spectra of all the samples presented in Figure 4 exhibited similar absorption bands with a slight difference in two primary amide bands, namely the amide I (at ∼1630 cm–1) and amide II (at ∼1513 cm–1) regions, being used frequently to assess polymer orientation and their conformations
41
; hence, an increase or a decrease in the ratios of their intensities (IAI/IAII) could be attributed to the changes in the orientation of secondary structures. Furthermore, the change in the amide I band shape is expected to have the same functional dependence as the amide I/amide II intensity ratio.
42
In that respect, the peaks were characterized using the ratio of the peak areas between amide I and amide II, as seen in Table 2. The assignments of intensities were determined between the local minima at 1470 and 1577 cm–1 for the amide II and at 1577 and 1717 cm–1 for the amide I ranges.
Attenuated total reflectance-Fourier transform infrared spectra of the references and differently ɛ-poly-L-lysine (ɛPL)-functionalized wool. TGase: transglutaminase; EDC: 1-ethyl-3 -(3-dimethylaminopropyl)-1-carbodiimide hydrochloride; NHS: N-hydroxysuccinimide. Relative peak areas of amide I and amide II regions of Fourier transform infrared spectra for the references and differently ɛ-poly-L-lysine (ɛPL)-functionalized wool samples TGase: transglutaminase; EDC: 1-ethyl-3 -(3-dimethylaminopropyl)-1-carbodiimide hydrochloride; NHS: N-hydroxysuccinimide.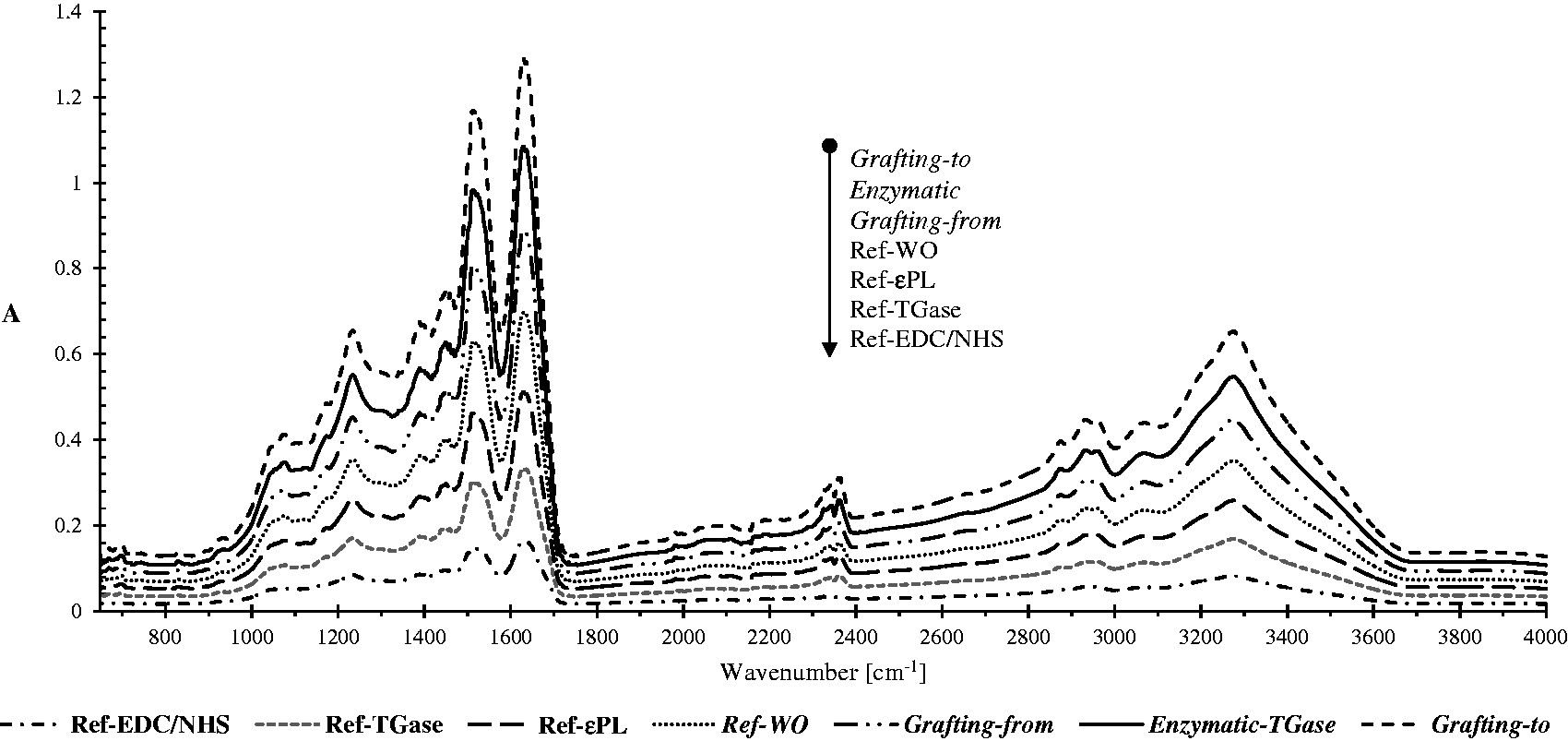
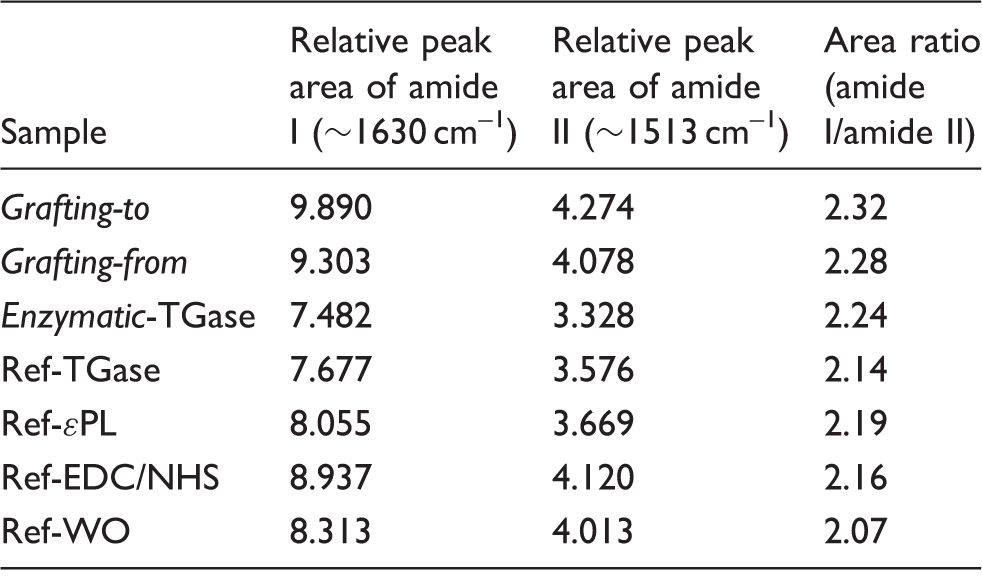
All functionalized samples had higher peak area ratios than the references, indicating that the grafting yield affected the protein amide I and II peak areas. The wool being functionalized chemically by the grafting-to approach exhibited the highest peak area ratio (A = 2.32), which had increased by ∼12% in comparison with the reference wool (A = 2.07) and by ∼7% compared to all the other references. In the case of grafting-from (∼10% increase) or the enzymatic (∼9% increase) coupling approach, the peak area ratios were lower in comparison with the grafting-to chemistry, which was also consistent with the quantification of amino groups, evaluated by the spectroscopic method and potentiometric titration.
The amide I absorption band that originates mainly from the C = O stretching of the peptide linkages (approximately 80%) is especially sensitive to the secondary structures of the proteins and could be used to determine changes in protein after functionalization, as each type of secondary structure gave rise to somewhat different C = O stretching frequencies due to their unique molecular geometries and hydrogen bonding patterns.
43
Accordingly, the curve fitting procedure was used to quantify the area of α-helix, β-sheet, random, and turn structures, representing the secondary structures of the wool protein. As seen in Figure 5, 20 different peaks were identified in the second derivative spectra within the 1570–1700 cm–1 amide I region, being assigned to various types of polymer orientation. The band centered at ∼1633 cm−1 was attributed to the intramolecular β-sheet; the peak at ∼1689 cm−1 and the bands within the 1618–1627 cm−1 regions were due to the intermolecular β-sheets. The contribution of the random coil conformation was identified by a peak centered at ∼1647 cm–1, the α-helix centered at ∼1651 cm–1,
44
and disordered conformations being assigned to the bands between 1675 and 1700 cm–1.
45
The 310-helix with 15–20% of all helixes is the fourth most common type of secondary structure in proteins after α-helices, β-sheets, and reverse turns, and is commonly found as N- or C-terminal extensions to the α-helix.
46
Fourier self-deconvoluted Fourier transform infrared spectra (shown as absorbance) of the amide I region (1700–1570 cm−1, C = O stretching) with relative areas of assigned secondary structures for the reference and differently ɛ-poly-L-lysine-functionalized wool samples. TGase: transglutaminase.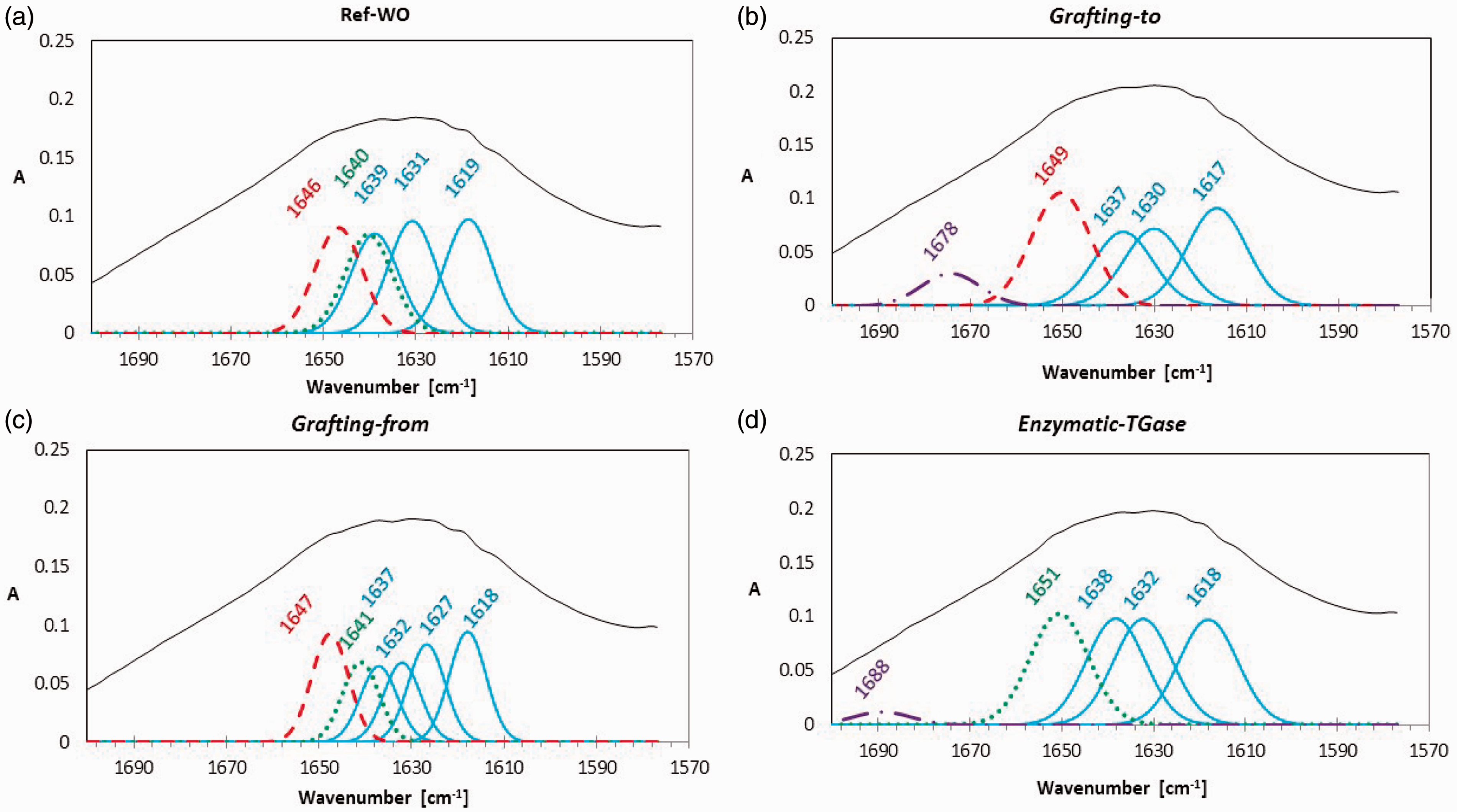
The results of the relative areas of the de-convoluted absorbance bands, shown in the inserted table in Figure 5, revealed that a change in the secondary structures occurred in all ɛPL-functionalized wool samples. Whilst the ß-sheet content increased for all the grafting mechanisms, the 310-helix structure from the Ref-WO diminished to about one fifth or even disappeared during the grafting process. New specific bands also occurred after its functionalization with ɛPL: the band at ∼1651 cm–1 corresponding to the α-helix structure 43 appeared only in the case of the enzymatic grafting approach, while the peak for β-turn conformation occurred in the case of the grafting-to and enzymatic coupling approaches. Besides, the band at ∼1647 cm–1, which is characteristic for random coil formation, was the highest in the wool functionalized by the grafting-to mechanism, which can be assigned to the fact that the grafting-to synthesis approach uses available amino groups of wool reacting with the terminal carboxylic group from the ɛPL; thus, the remaining free amino groups from the ɛPL, which are charged, can affect the forming of random coil structures. In contrast, the grafting-from approach uses an initiated terminal carboxylic site on the ɛPL to form a peptide bond with the wool amino groups, resulting in the formation of both helical (at ∼1641 cm–1) and random coil (at ∼1646 cm–1) structures.
Finally, the contribution of relative areas of the de-convoluted absorbance bands within the amide I region can thus be attributed to the molecular orientation of the grafted ɛPL
EPR spectroscopy is a widely used spectroscopic technique for determining the structure–function correlations in protein macromolecules.47,48 As the mobility of a spin-label MP being covalently attached to ɛPL is influenced by its micro environment, the analysis of the EPR spectra provides information on the orientation of site-specific immobilized ɛPL-MP on wool. Presuming that all MP molecules (1000/1 molar excess of available groups over the amount of MP added) are covalently tethered to ɛPL through their amino group availabilities, line-shape differences among samples thus yield information about their structural environments. 49
The rotational correlation time (τc, which may be viewed as the time taken for an axis of the nitroxide group of MPs to travel through one radian) is highly sensitive to the motion of the chain.
50
In the fast tumbling limit (10–11 < τc < 10–9 s), the EPR spectrum of the MP’s nitroxide consists of three anisotropic lines, the broadenings of which generally indicate MP immobilization, whereas their sharpening indicates an increase in MP mobility. If the MP is thus attached to a molecule that is sufficiently small or flexible, it exhibits an isotropic EPR spectrum, as shown in Figure 6, presenting a clean triplet signal of ɛPL-MP, being typical for nitroxide within the fast motional regime where τc is below 2 ns. In contrast, if the mobility of the MP’s nitroxide is restricted, a less isotropic EPR line-shape would be observed due to the unpaired electron of the MP residing in a molecular orbital 2 p along the N–O bond.
51
Electron Paramagnetic Resonance spectra of 3-maleimido-PROXYL spin-labeled ɛ-poly-L-lysine (ɛPL-MP) in comparison with the wool functionalized with ɛPL-MP using different coupling approaches.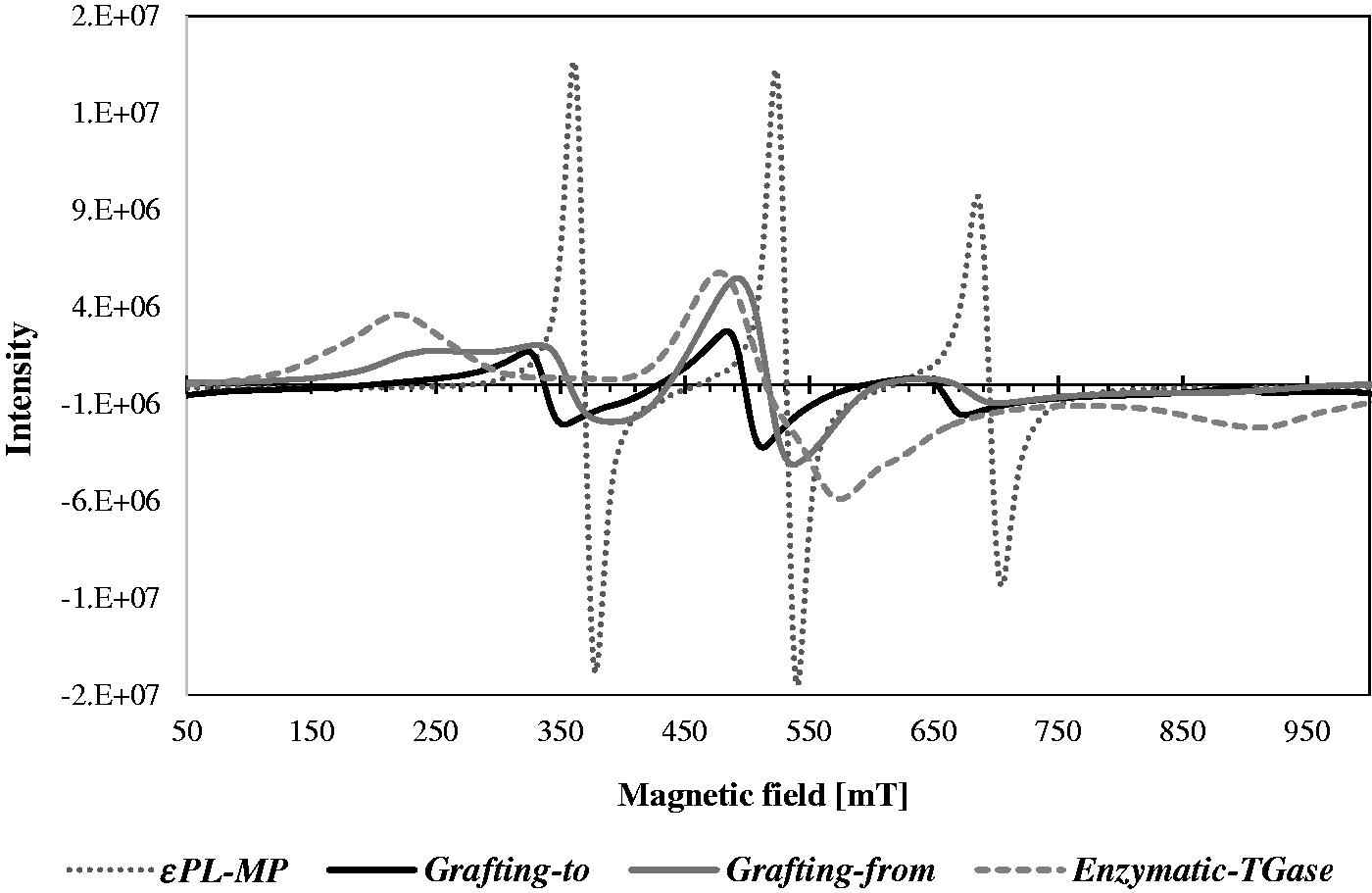
The EPR spectra of the wool being functionalized with ɛPL-MP by different approaches displayed different patterns of the characteristic MP nitroxyl triplet signal. Rotational correlation times of ɛPL-MP functionalized wool were significantly higher than those for ɛPL-MP (τc ≈ 0.11 ns). In addition, the comparison between spectra shows a dramatic change in spectral line-shape, which can be associated with the modes of coupling chemistry and the mechanism (synthesis route) used. The MP attached to a molecule with a sufficiently slow τc (>0.1 ns) exhibits a broad EPR spectrum typical for a completely immobilized sample, as in the case of TGase grafted ɛPL-MP (τc ≈ 1.13 ns). Depending on the degree of ɛPL-MP immobilization, partially broadened spectra may also be observed in the chemical grafting-from approach, indicating the mobility of an intermediate time scale (τc ≈ 1.05 ns). As the τc of the ɛPL-MP functionalized wool by the grafting-to mechanism is lower (τc ≈ 0.39 ns), it suggests that a grafted-to ɛPL-MP has higher mobility and thus indicates that ɛPL molecules are linked to a wool surface within a highly flexible “brush-like” structure. On the other hand, in the case of the grafting-from mechanism, the ɛPL-MP may also be coupled to wool by ɛPL side amino groups, leading to a splitting of grafted ɛPL chains into several parts forming loops or tails similar to the brush-like layer. 24
The results are well correlated with the FTIR spectra analysis, also indicating the fact that the conformation of ɛPL onto the wool is affected not only by the coupling approach, but also by the pH used for the grafting. Namely, at lower pH values (pH 4.5), the ɛPL possesses mainly the PII (polyproline II helical) and β structures, 52 which are very likely to have been formed in the grafting-to approach where the highest flexibility was observed. In contrast, at higher pH values (pH 10) the α-helical conformation may also occur, leading to a more restricted motional regime, as in the case of the grafting-from and enzymatic coupling approaches.
Antibacterial activity of ɛPL-functionalized wool
The results of the in vitro growth-inhibitory activities of differently treated wool substrates are shown in Figure 7. For satisfactory antimicrobial activity the bacteria reduction should exceed 60%, which is represented by the dashed line. As can be seen, all ɛPL-functionalized wool showed satisfactory bacteria reductions (≥60%), being reached much faster kinetically (after 3 h) for gram-positive S. aureus compared to Gram-negative E. coli bacteria, reaching this value after 6 h of inoculation. However, the functionalization of wool by the grafting-to approach was the most effective, giving ∼99% of both bacteria reductions, although the grafting-from and TGase-catalyzed approaches also gave satisfactory results with the reduction abilities of ∼74% and ∼86% for E. coli and, even higher, ∼92% and ∼82% for S. aureus, after 24 h of exposure. The apparent time-related viability decrease reflects the higher resistance of gram-negative E. coli, being attributed to its complex cell wall structure that is comprised of multiple layers in which an outer membrane layer containing long chain lipopolysaccharides in its outer leaflet and phospholipids in the inner leaflet lies on top of a thin peptidoglycan layer (which is much thicker in gram-positive bacteria). Such a thick cell wall structure restricts the penetrations of chemicals as well as antimicrobial agents. On the other hand, gram-positive bacteria have a mesh-like peptidoglycan layer, which is more accessible to permeation by the agents.
53
Time-dependent reduction of (a) gram-negative E. coli and (b) gram-positive S. aureus bacteria on the references and differently ɛ-poly-L-lysine (ɛPL)-functionalized wool samples, being evaluated by using the ASTM E2149-10 shake flask method
32
and corresponding bactericidal effect after 24 h of bacterial inoculation. EDC: 1-ethyl-3 -(3-dimethylaminopropyl)-1-carbodiimide hydrochloride; NHS: N-hydroxysuccinimide; TGase: transglutaminase.
As already discussed, the ɛPL was initially described to show the analogical antimicrobial kinetics against different bacteria by causing irreversible changes in both the inner and outer cell membranes of pathogens. However, as all immobilized ɛPL on the wool is bonded covalently, its good antibacterial activity can be attributed to its electrostatic interactions with the bacterial membrane, followed by stripping off the membrane and distribution of their cytoplasm. 54 For that purpose, the staining of bacteria after 24 h of inoculation was performed by using the Live/Dead BacLight bacterial viability staining test for detecting live cells with an intact membrane stained green by PI, and dead cells with a damaged membrane stained red by SYTO9.
The data inserted in the graphs in Figure 7 show an increase in bactericidal effect, as a result of the relative decrease in green fluorescence intensity when wool is functionalized with ɛPL. These values for gram-negative E. coli were comparatively higher and increased faster from enzymatic (∼63%) via grafting-from (∼70%) to grafting-to (∼83%) coupled ɛPL compared with those for gram-positive S. aureus (∼58% versus ∼54% versus ∼64%). However, there is a clear need for critical evaluation of the results obtained from combined staining, as previous studies have shown that SYTO9 could not be effective in staining some intact gram-negative bacteria, being related to its permeability problems through the double-membrane layer or its exporting from the bacteria cytoplasm, 55 while a decrease in green fluorescence intensity can be related to the entrance of PI. 56 The confocal fluorescence microscopy imaging of the samples was performed despite the fact that the combined fluorescent staining may lead to results with some deviations.
In Figure 8 the presented representative fluorescence images after exposure to the bacterial strain show green labeled cells on the surface of all the tested samples, being much more intensive on the Ref-WO when compared to the other modified wool samples, where the presence of red bacteria indicates that ɛPL may destroy the cytoplasmic membranes of bacterial cells before lysis. Similar observations were reported recently by Zhou et al.,
57
where various damage to the E. coli cell envelope by ɛPL demonstrated detachment of the outer membrane, swelling of the inner membrane, apical bursting of cells, and leakage of cytosol at a minimal inhibitory concentration. This hypothesis may also be supported by the already approved bactericidal effect of ɛPL by the Federal Drug Administration,
58
indicating that the bactericidal action of ɛPL against both bacteria is mediated by physical ionic interactions with the microbial cell wall inducing pore formation and/or disintegrating the cell membrane. Cytoplasmic membrane damage and cell permeabilization caused by ɛPL would thus be responsible for PI entry and cell staining. Furthermore, the Live/Dead confocal micrographs confirmed the highest cell density on the wool being ɛPL-functionalized by the grafting-to approach as compared to the other two mechanisms used. In addition, the ratio between the green (living) and red (dead) cells (i.e. cell viability) varied proportionally with the quantity of bound ɛPL, being the highest for the sample functionalized by the grafting-to coupling approach.
Confocal microscopy images of the ɛ-poly-L-lysine-functionalized wool samples by grafting-to ((a) and (b)), grafting-from ((c) and (d)), and enzymatic transglutaminase (TGase) - catalyzed ((e) and (f)) coupling approaches in comparison with the reference sample (Ref-WO, (g) and (h)), after exposure to a media inoculated with gram-negative E. coli ((a), (c), (e), (g)) or gram-positive S. aureus ((b), (d), (f), (h)) for 24 h, and a Live/Dead BacLight bacterial viability staining test. Representative scans were taken at 60 × magnification showing a green color for Live and a red color for Dead bacterial cells. (Color online only.)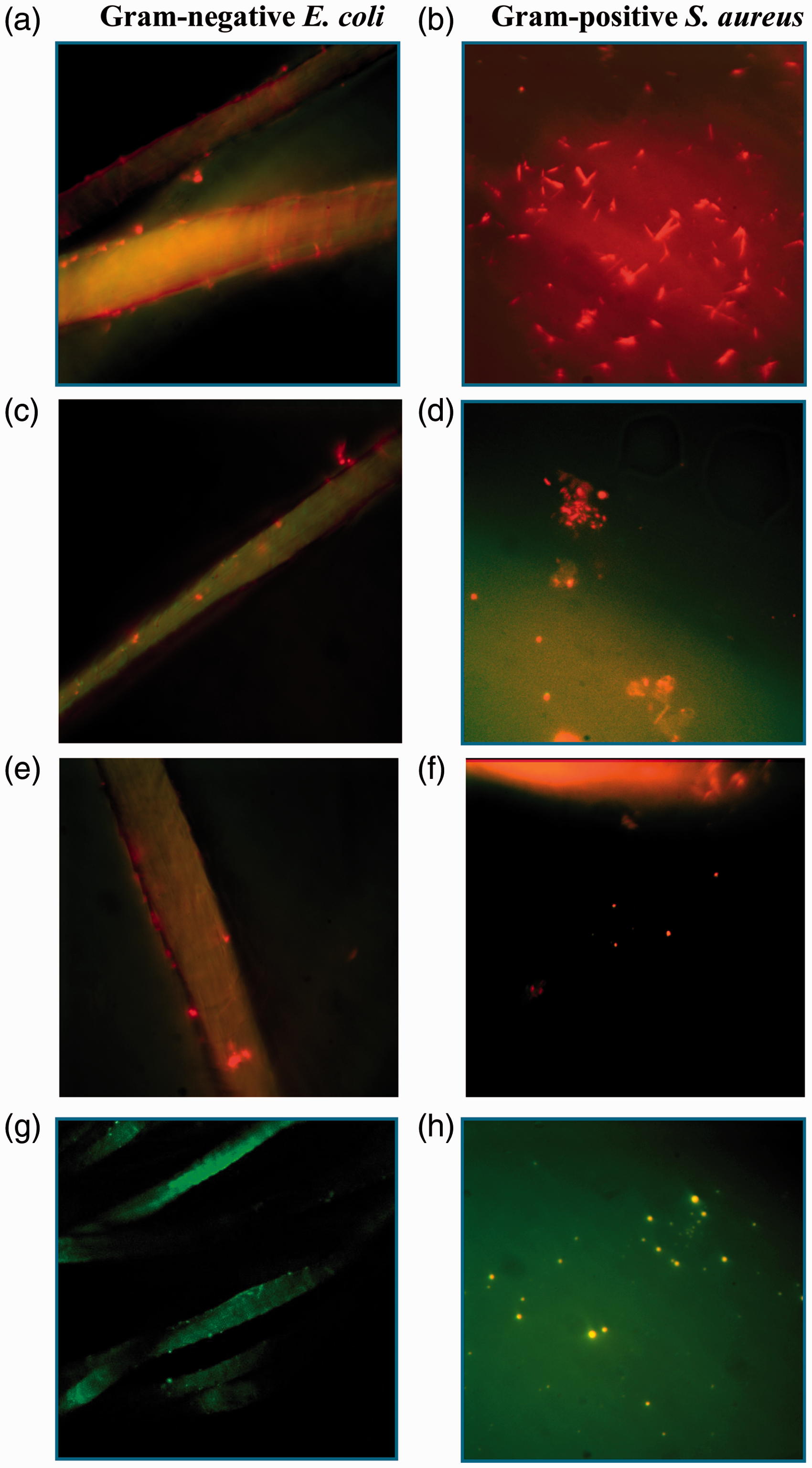
Durability of wool’s antibacterial activity to washing
In order to evaluate the durability of the wool’s antibacterial effect after washing, the ɛPL-functionalized wool substrates were washed according to the standard ISO 105-D01:2010 test method.
59
As shown in Figure 9, the wool functionalized by the grafting-to approach still provided ∼86% reduction of gram-negative E. coli after five washing cycles, being slightly lower for ɛPL coupled by the grafting-from approach but still satisfactory, while showing a minimal but still sufficient effect in the case of enzymatic-TGase functionalized wool. However, although the ɛPL was immobilized covalently, an increased number of washes (20 washing cycles) caused progressively reduced antimicrobial efficacy. As the presence of residues of non-ionic surfactant after washing may reduce the antimicrobial activity of the wool,
60
the samples were extracted by Soxhlet extraction using dichloromethane as the extraction solvent, and the non-ionic detergent measured gravimetrically as solvent extractable material after removing the solvent using a rotary evaporator under vacuum. The results presented in Table 3 show that wool adsorbed appreciable quantities of surfactant (i.e. ∼0.016 g/g of wool and ∼0.027 g/g of wool, submitted to five and 20 washing cycles, respectively), which was in accordance with the previous studies showing that non-ionic surfactants are adducts that can form complexes with wool protein by means of hydrophobic interaction between hydrophobic sites,
61
thus blocking the functional groups of wool. In addition, the ɛPL-functionalized wool washed with 20 cycles without the surfactant showed good wash fastness properties, indicating that ɛPL continuously inhibits the growth of bacteria in contact with the surface.
62
The grafting-to approach was the most effective, providing ∼98% reduction against E. coli and ∼79% against S. aureus. The influence of the surfactant chosen for washing seems to be crucial for both the effectiveness of the finishing (in our case antibacterial activity) and the durability evaluation, requiring additional studies.
The reduction of gram-negative E. coli and gram-positive S. aureus on the reference and differently ɛ-poly-L-lysine-functionalized wool samples, being washed with and without non-ionic surfactant according to ISO 105-D01:2010 for five and 20 cycles, evaluated by using the ASTM E2149-10 shake flask method. TGase: transglutaminase. The amount of non-ionic surfactant extracted from ɛ-poly-L-lysine-functionalized wool by the grafting-to coupling approach and submitted to five and 20 washing cycles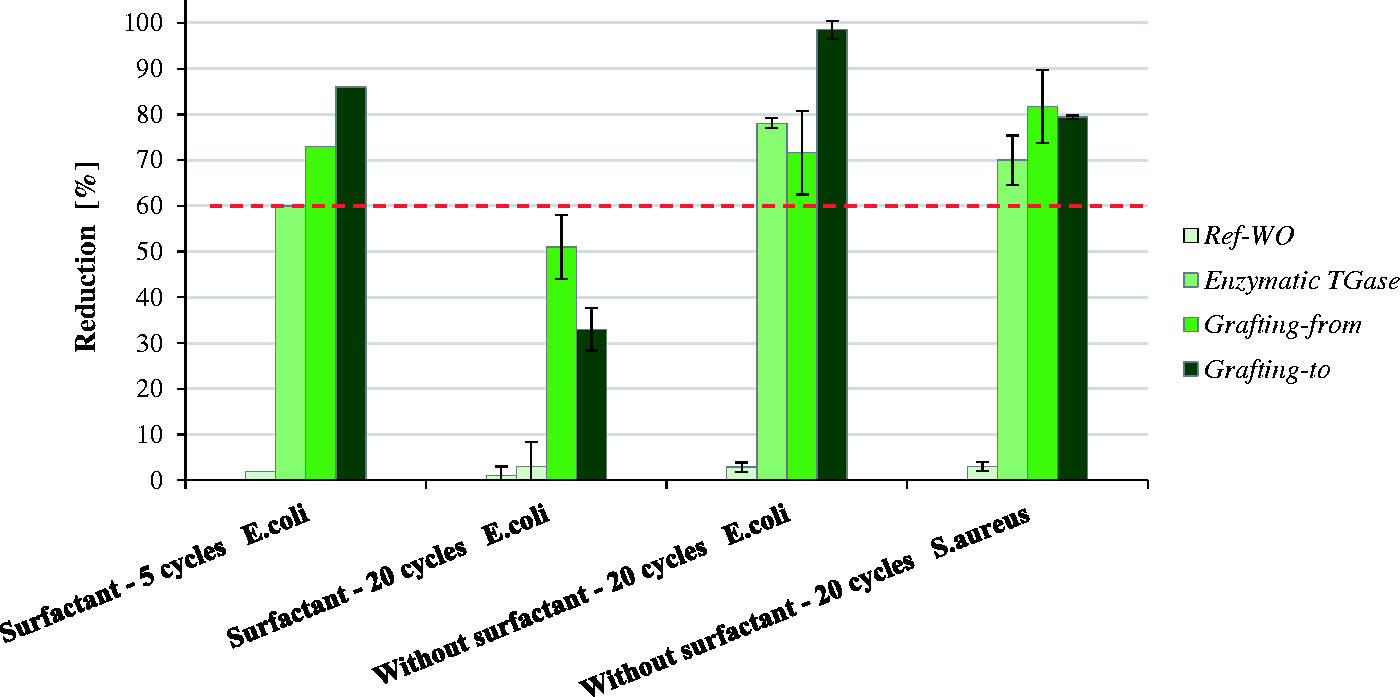

Conclusions
The current study presents the effect of the coupling strategy (grafting-to, grafting-from, enzymatic using TGase) of ɛPL to the wool using the non-toxic and environmentally friendly EDC/NHS mechanism, leading to antibacterially protective wool. While the quantity of ɛPL applied was evaluated by different direct and indirect analytical methods using spectroscopic techniques and potentiometric titration, the detailed structural characterization of the wool–ɛPL conjugate was undertaken by FTIR and EPR spectroscopies. The most efficient and conformationally the most flexible “brush-like” structure of ɛPL binding to the wool was achieved by using the grafting-to approach, resulting in the highest antibacterial activity against gram-positive S. aureus and gram-negative E. coli, being further confirmed by the Live/Dead bacterial viability testing, exhibiting adequate bactericidal properties of the wool as a result of bacterial cell membrane leakage.
In addition, the non-ionic surfactant used for testing of the wool’s antibacterial effect durability to washing was shown to adsorb strongly onto the wool fibers, thereby blocking the functional sites of ɛPL and thus preventing its interaction with bacteria. The influence of the surfactant chosen for washing seems to be crucial for both the effectiveness of the finishing (in our case antibacterial activity) as well as the durability evaluation, requiring additional studies.
Finally, this work brought new knowledge into the designing of antimicrobially active materials and offers the potential for creating multifunctional and specific antimicrobial protection.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Ministry of Higher Education, Science and Technology and 7FP (Era-Net Matera Plus ANTIMICROB PEPTIDES grant number 3211-10-000369).