
Abstract
Naturally green-colored domestic silk demonstrates remarkable antioxidant activity due to the presence of intrinsic antioxidant and pigmented substance flavonoids. Oxidative damage is one of the main causes of sun-induced skin damage. To investigate the potential protective effect of naturally green-colored domestic silk on skin cells against ultraviolet damage, the antioxidant was extracted and analyzed. An in vitro experiment which involves introducing ultraviolet damage to the human keratinocyte HaCaT cell line was used for the research. By adding the extract into HaCaT culture medium and then subjecting to ultraviolet irradiation, the data evidently showed that the cell apoptosis and growth inhibition of HaCaT were alleviated, indicating the protective effect of the extract to the HaCaT cell against ultraviolet damage. Meanwhile, a large number of the surviving damaged cells stagnated in the diploid (2n) phase, which is the main phase of gene repair during the whole cell cycle. The survival and repair of damaged cells also produced fewer gene fragments caused by ultraviolet damage. The detected reduction of reactive oxygen species and malondialdehyde induced by ultraviolet irradiation in HaCaT confirmed that the antioxidant extract from green silk cocoons does have intracellular and extracellular antioxidant effects. These findings suggest that naturally green silk has the potential to be utilized as a functional textile material for skin sunscreen products, and its extract can be further developed as a source of antioxidants in medical and safety health applications.
Keywords
Naturally colored domestic silks (NCSs) are produced by the color mutations of the silkworm Bombyx mori. As controlled by specific transport genes, these silkworms can uptake natural pigments into the silk gland from mulberry leaves.1,2 NCSs mainly have three color varieties; that is, flavonoid-pigmented yellowish-green silk and carotenoid-pigmented golden yellow and pink silks.3 –5 The carotenoids in golden yellow and pink silks were mainly β-carotene and lutein, which were consistent with those in mulberry leaves. Carotenoids have outstanding biological effects such as protecting eyesight, but carotenoids have a wider range of sources in plants than in silk. 1 The flavonoids in the cocoon shells of green NCSs mainly consist of quercetin, isoquercitrin, and kaempferol, which are different from those in mulberry leaves. The flavonoids from silk have been specially digested and modified by the silkworm, which occupy a content of 33.4 mg/g in the fiber.3,6 It is well known that flavonoids are a class of polyphenols with multiple physiological activities, such as antioxidation, antibacterial, anti-inflammatory, antitumor, and so on. 7 As reported previously, green NCSs showed a scavenging activity 12–24 times that of common white silk against free radicals. 8 It was also found that ethanolic extract from green NCSs showed strong antioxidant activity, and could significantly improve the glucose metabolism of mice with type 2 diabetes mellitus by antioxidation and anti-inflammation. 9 As a result of their great potential to be used in the food and cosmetic industry, the antioxidant substances in the cocoon silks, as byproducts of the silk degumming industry, are drawing increasing interest. Naturally green silk fiber contains a high content of natural antioxidant flavonoids, which gives it a potential application value as a natural antioxidant fiber. In recent years, antioxidant textiles have not only gained commercial application, but also been used in clinical trials to treat certain skin diseases. 10 Therefore, naturally colored silk is not only a kind of ‘green’ environmental protection fiber that does not need dyeing, but is also an excellent natural functional fiber, which has greater application value in the field of healthcare textiles. The antioxidant compositions in the common white silk,11 –13 tasar silk, 14 NCSs,3,4,6,8 and the silk of Rondotia menciana 15 and Sasamayu 16 have been well investigated. Naturally colored silk has similar morphological structure and mechanical properties to ordinary white silk. The main difference is that naturally colored silk contains more such compounds, which is 1.2 to 1.9 times that of ordinary white silk. 4 Among these silk varieties, green NCS has been proved to have the strongest antioxidant activity. The antioxidant activity is related to the structure of the phenolic hydroxyl group in the flavonoid, which can combine with the free radical to form a stable semi-quinone free radical. This combination can terminate the conduction process in the oxidative chain reaction.
Oxidative damage is one of the main reasons for skin damage induced by sunlight. Ultraviolet (UV) irradiation induces the production of massive free radicals, and disturbs the oxidant–antioxidant balance in the skin tissue, thus not only causing photoaging of the skin, but also resulting in some skin diseases, such as lupus erythematosus, solar urticaria, and even skin cancer.17,18 UV irradiation can cause covalent cross-linking within and between DNA strands, mainly forming pyrimidine dimers, resulting in DNA base dimerization. Antioxidants have been widely used as topical drugs to protect the skin from photooxidative damage, or treat damaged skin. 19 External application of exogenous antioxidants enables the penetration of the antioxidants to the dermis from the surface of the skin, and can scavenge the excessive free radicals in the skin tissues, which have resulted from UV exposure. Recently, loading antioxidants onto/into textiles for skin care or treatment purposes has received increasing attention. Natural antioxidants, such as vitamins C and E, flavonoids, tea polyphenols, tannins, gallic acid, and resveratrol have been used for preparing antioxidant textiles.20 –25 The efficacy of some antioxidant textiles has been confirmed by clinical trials and double-blind trials. For example, Martí et al. 24 loaded gallic acid onto cotton fabrics by microcapsule incorporation and a dip-pad-dry process. The fabrics showed a clinically significant effect in decreasing reactive oxygen species (ROS) in the skin tissue and reducing the lipid peroxidation levels by about 10%.
Compared with antioxidant-loaded textiles, green NCSs have intrinsic high-activity antioxidants inside the fiber, and the activity is much more durable to rubbing and repeated home washing. 8 Our recent work confirmed that green NCSs could deliver flavonoids to the human skin with an amount of 5.7 μg/cm2 in 24 h, and could decrease ROS and lipid peroxidation levels by 16.7% and 37.8%, respectively. The antioxidant components in naturally green silk could migrate to the cuticle and dermis of pig skin, and the amount of migration increased with time. The amount of antioxidant components migrating to the skin was closely related to the skin surface moisture, acidity and alkalinity. Humidity and acidity could promote the migration of antioxidant components. The more antioxidant components absorbed by the cortex, the stronger the free radical scavenging ability of the cortex, indicating that the antioxidant activity of each cortex was dose dependent. 26 Current research usually focuses on the prevention of antioxidants toward UV-induced skin oxidative damage at tissue levels. In the present study, the effects of the antioxidants from green NCSs on protecting and repairing human skin against UV damage were investigated at a cellular level.
Materials and methods
Extraction of antioxidant substance
The antioxidants in the green NCSs were extracted by immersing the cocoon shells in a 50% (v/v) aqueous ethanol solution with a solid/liquid ratio of 1:60 at room temperature. After ultrasonic extraction for 120 min, the extract was concentrated under vacuum to reach a total flavonoid content of 10.8 mg/mL, which was determined by a sodium nitrite-aluminum nitrate colorimetric method. 27 The extract was freeze-dried and redissolved in phosphate buffered saline (PBS), and diluted with culture medium according to the required concentration in cell culture.
Preparation of cell culture
HaCaT (derived from American type culture collection) was selected as an in vitro model, and was cultured in Dulbecco's modified eagle medium (Sigma-Aldrich, USA) with 10% wt of fetal bovine serum (Biological Industries, USA) and 5% carbon dioxide (CO2) at 37°C. The UV damage model was set by irradiating the cells with six 9 W UVB lamps for 300 s at a distance of 15 cm. Radiant intensity = 0.8 mW/cm2, UVB dose (mJ/cm2) = UVB radiant intensity (mW/cm2) × time (s).
DPPH and ABTS+ radical scavenging tests
DPPH (1,1-diphenyl-2-picrylhydrazyl radical 2, 2-diphenyl-1-(2,4, 6-trinitrophenyl) hydrazyl) is a stable nitro-centered radical with a single electron, the alcohol solution of DPPH is purple. When free radical scavenger presents, the alcohol solution fades. ABTS (2,2′-azinobis-(3-ethylbenzthiazoline 6-sulphonate)) is a substrate of peroxidase that produces a soluble greenish chelate. Both of them can be quantitatively analyzed by an ultraviolet spectrophotometer. 28
The antioxidant activity of naturally green cocoon shell extract in culture medium was measured by testing the removal of DPPH and ABTS+ free radicals. The scavenging rate of DPPH free radicals (%) = [(Ablank – Asample)/Ablank] × 100, the absorbance was measured at 532 nm. The scavenging rate of ABTS+ free radicals (%) = [(Bblank – Bsample)/Bblank] × 100, and the absorbance was measured at 733 nm.
Detection of ROS and MDA in HaCaT cells
The ROS enzyme linked immunosorbent assay kit (ELISA) (Shanghai Fusheng Industrial Co., Ltd., China) double antibody sandwich method was used to detect overall ROS in cell culture supernatant samples. The lipid peroxidation malondialdehyde (MDA) assay kit (Beyotime Biotechnology) was used for MDA detection. The principle was based on the reaction of MDA and thiobarbituric acid (TBA) to produce red products with maximum absorption at 535 nm. The MDA in HaCaT cell lysis solution was quantitatively detected by colorimetry.
Cell proliferation, apoptosis and cell cycle assays
Cell proliferation was determined using the cell counting kit-8 (CCK-8; Boster Biological Technology, USA.) assays, which reflect cell mass by detecting the metabolism of living cells. 29 Cell samples were prepared in four replicates; 100 μL of cell medium were seeded at about a density of 3000 cells/well in 96-well plates, and determined every 24 h in 5 days. The optical density of the cells was measured at 450 nm by using a SpectraMax i3 fluorescence microplate reader (Molecular Instruments Co., Ltd., USA). The data were analyzed and processed using the t-test.
Cell apoptosis and cycle experiments were detected by flow cytometry, fluorescent isothiocyanate (FITC)-conjugated annexin V and propidium iodide (PI) double staining test kit (Multisciences Biotech Co., Ltd., China) and cell cycle staining kits (Multisciences Biotech Co., Ltd., China).
Results and discussion
Compared with the flavonoids extracted from plants, flavonoids extracted from Bombyx mori cocoons have fewer impurities and high yield or efficacy. These flavonoids might come from Bombyx mori metabolism. Studies on the difference of flavonoid compounds in plants and silk have already been conducted, Tamura et al. 30 identified the structures of flavonoids isolated from cocoon shells through spectroscopic analysis. These flavonoid glycosides, which were not present in mulberry leaves, were considered to be metabolites of silkworms. This metabolism was performed by a glucosyltransferase unique to silkworms, which transferred the glucose portion to quercetin's hydroxyl group, and increased quercetin's hydrophilicity, but the specific biological functions are still not clear. 30
Antioxidant capacity of extract in cell culture medium
To detect the antioxidant benefits of green silk cocoon extract on cells, DPPH and ABTS+ clearance assays were used to detect the antioxidant activity of extract in cell culture medium. ABTS+ and DPPH were added as free radicals to cell culture medium containing extract of different concentrations for co-incubation, and their absorbance was detected. The extract antioxidant capacity, characterized by the removal rate of ABTS+ and DPPH by the extract contained in cells, was then analyzed according to the absorbance of the incubated solution. The data are presented in Figure 2. The results show that the extract exhibited concentration-dependent antioxidant activity in cell culture medium, the highest clearance rates of ABTS+ and DPPH in the culture medium reached 79% and 73%, respectively.
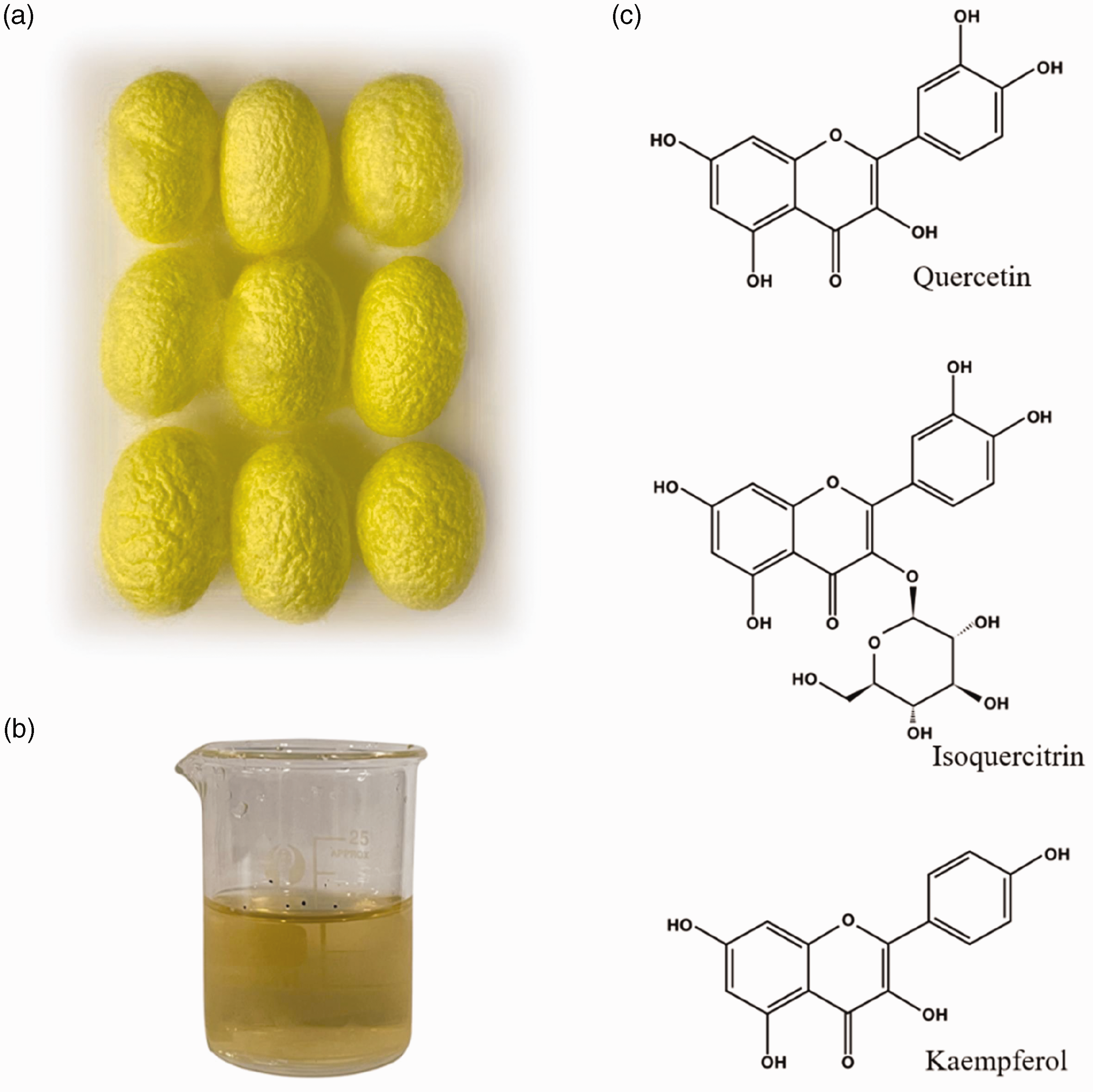
Naturally colored domestic silks and the antioxidants extract. (a) Naturally colored domestic silk cocoons; (b) the antioxidants extract and (c) the chemical structure of the main antioxidant components.
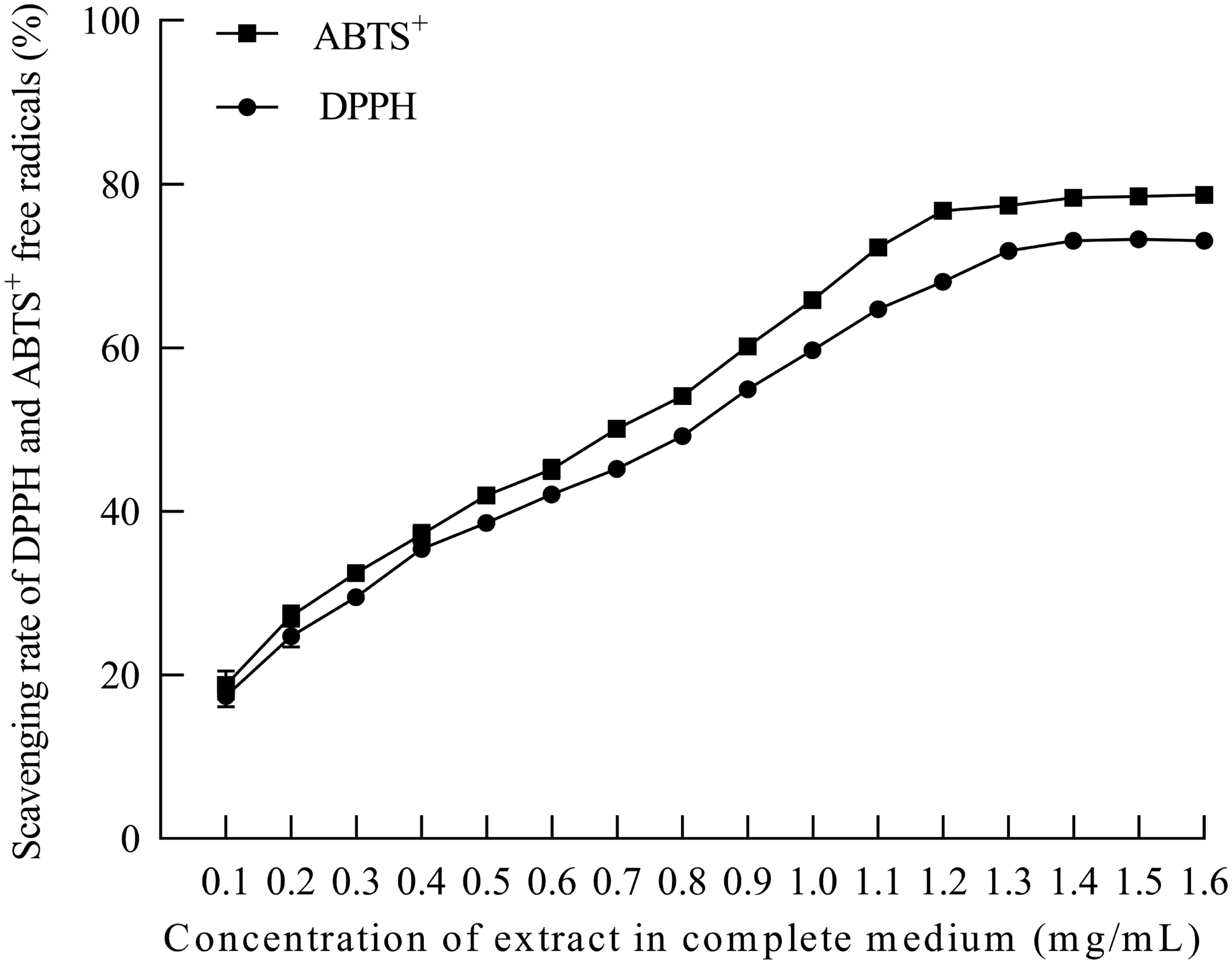
Antioxidant capacity of extract in cell culture medium.
Effects of extract on proliferation of HaCaT
In order to study the direct effect of green silk cocoon extract on the proliferation of HaCaT, we added different concentrations of extract into the cell culture medium, and observed the growth of HaCaT. HaCaT was cultured in complete medium containing 0.1–0.8 mg/mL of green silk cocoon extract for 5 days, and CCK-8 detection was performed every day to analyze the proliferation of HaCaT at different concentrations. As shown in Figure 3, by the third day of culture, the growth of HaCaT cultured with different concentrations of extract gradually showed differences. The results also show that the extract concentration higher than 0.5 mg/mL caused a certain degree of toxicity to HaCaT, and inhibited the proliferation of HaCaT. When the concentration of extract in the medium was less than about 0.4 mg/mL, there was no significant toxicity to HaCaT (unlabeled differences are NS, P > 0.05; *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001).
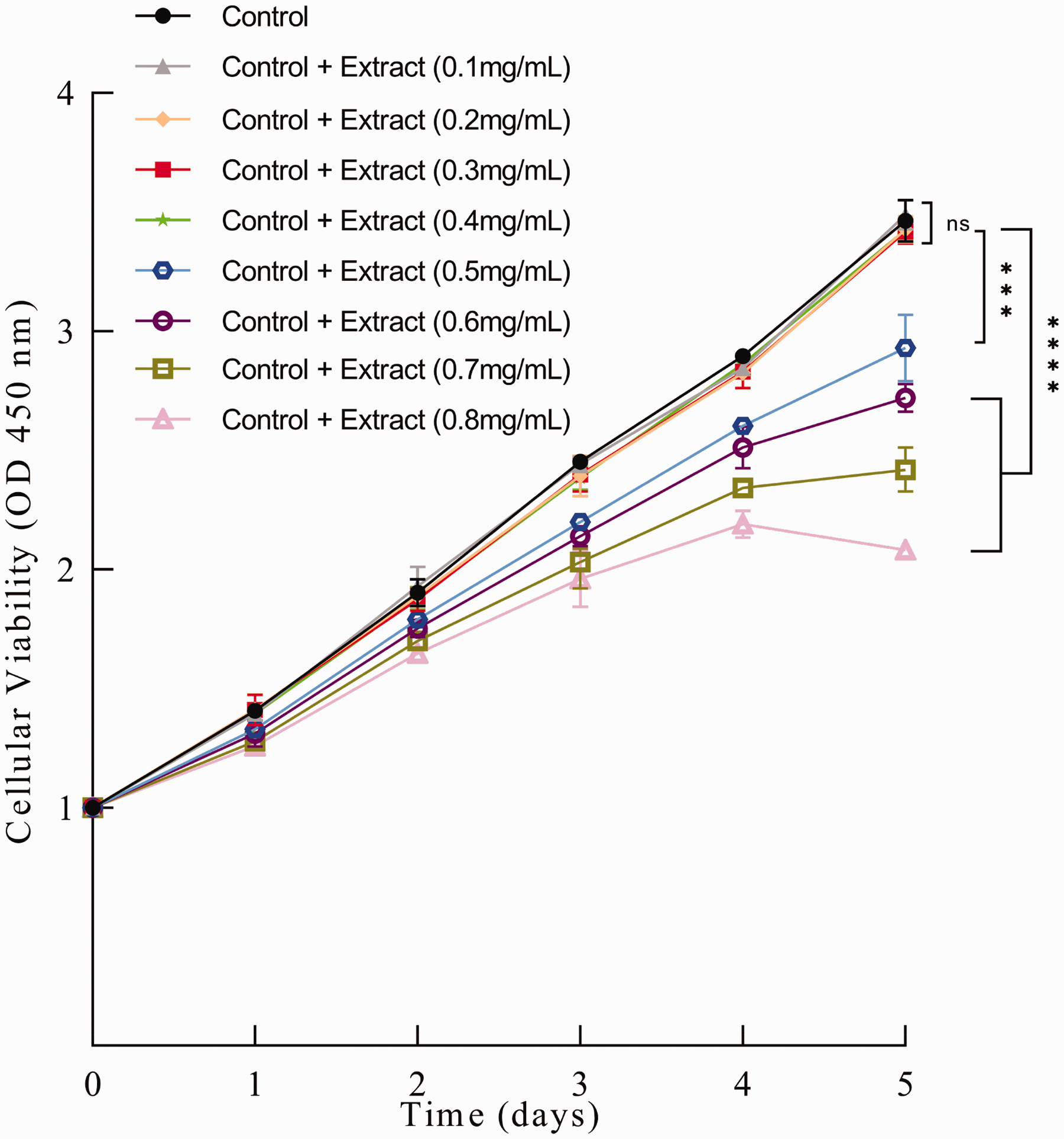
Effect of green silk cocoon extract on proliferation of HaCaT.
Protection of green silk cocoon extract against UV damage to HaCaT
To investigate the protective value of green silk cocoon extract against UV damage, an appropriate amount of extract was added into HaCaT around UV irradiation time, after which the growth and survival of HaCaT were detected. Different concentrations of the extract were added to HaCaT culture medium, and then subjected to UV irradiation before another 3 days culturing. The surviving cells remaining were then measured by CCK-8. As shown in Figure 4, primarily, we found the protective effect of UV damage on HaCaT reached the best level when the extract concentration in the cell culture medium attained 0.4–0.6 mg/mL. Considering that the excessive extract concentration would lead to certain toxic effects, 0.4 mg/mL was a safer effective protective concentration. Subsequently, the extract concentration of 0.4 mg/ml was applied to all other UV damage protection experiments.
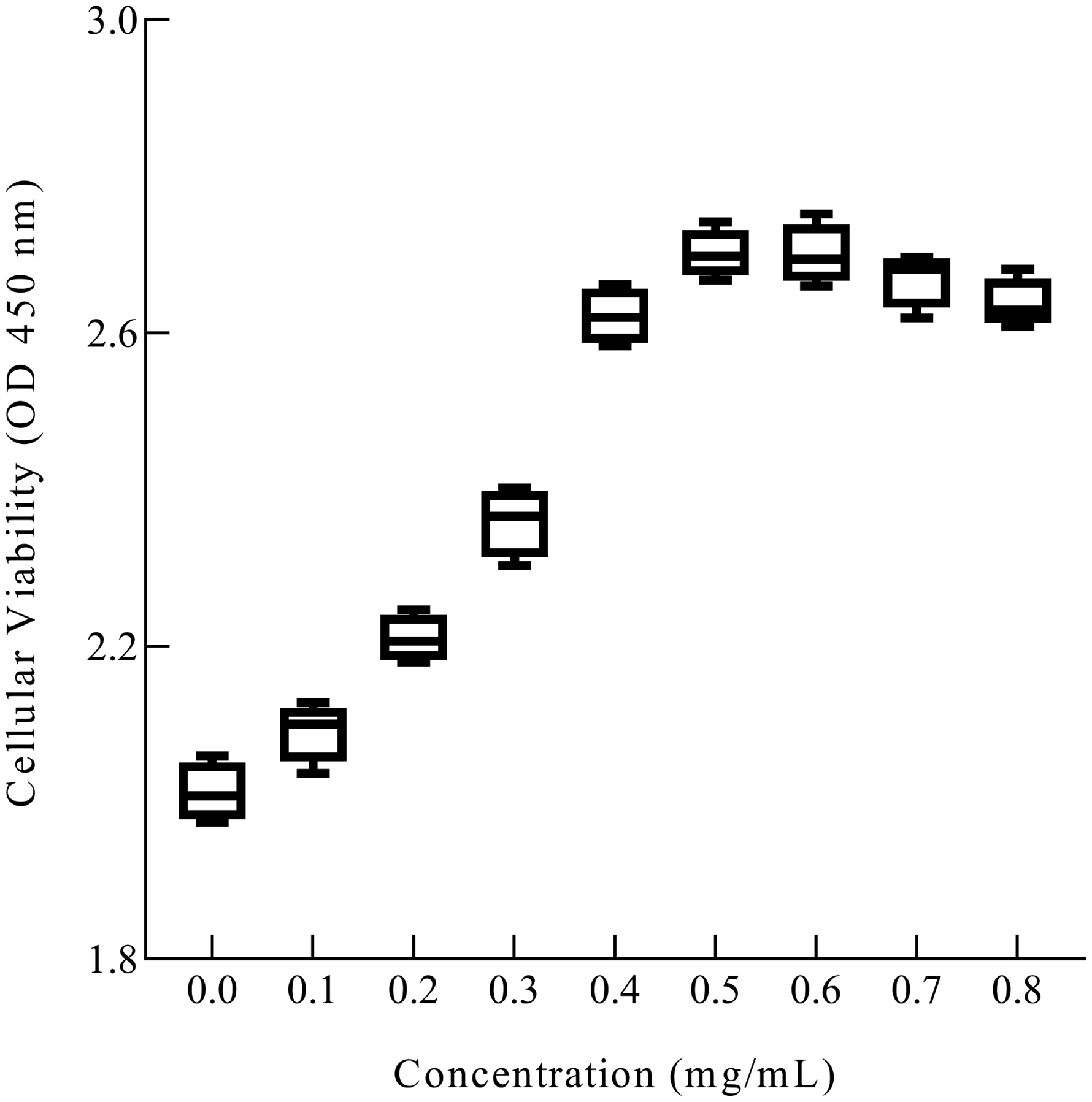
Effect of ultraviolet (UV) irradiation on cell proliferation of HaCaT as a function of the concentration of extract in cell culture medium.
It was found that adding extract at a different time around UV irradiation brought about different protective effects on HaCaT. First of all, the proliferation level of cells that suffer UV damage was increased by the application of the extract. The green silk cocoons extract existing simultaneously with UV irradiation produced the best protective effect on HaCaT against UV damage. Positive protective benefits were also obtained when HaCaT was pre-cultured for 4 h with the extract before UV irradiation, as well as when the extract was added to culture after UV irradiation. Nevertheless, the cells pre-absorption of the extract before UV irradiation had a better protective effect than treatment with the extract after UV damage (Figure 5).
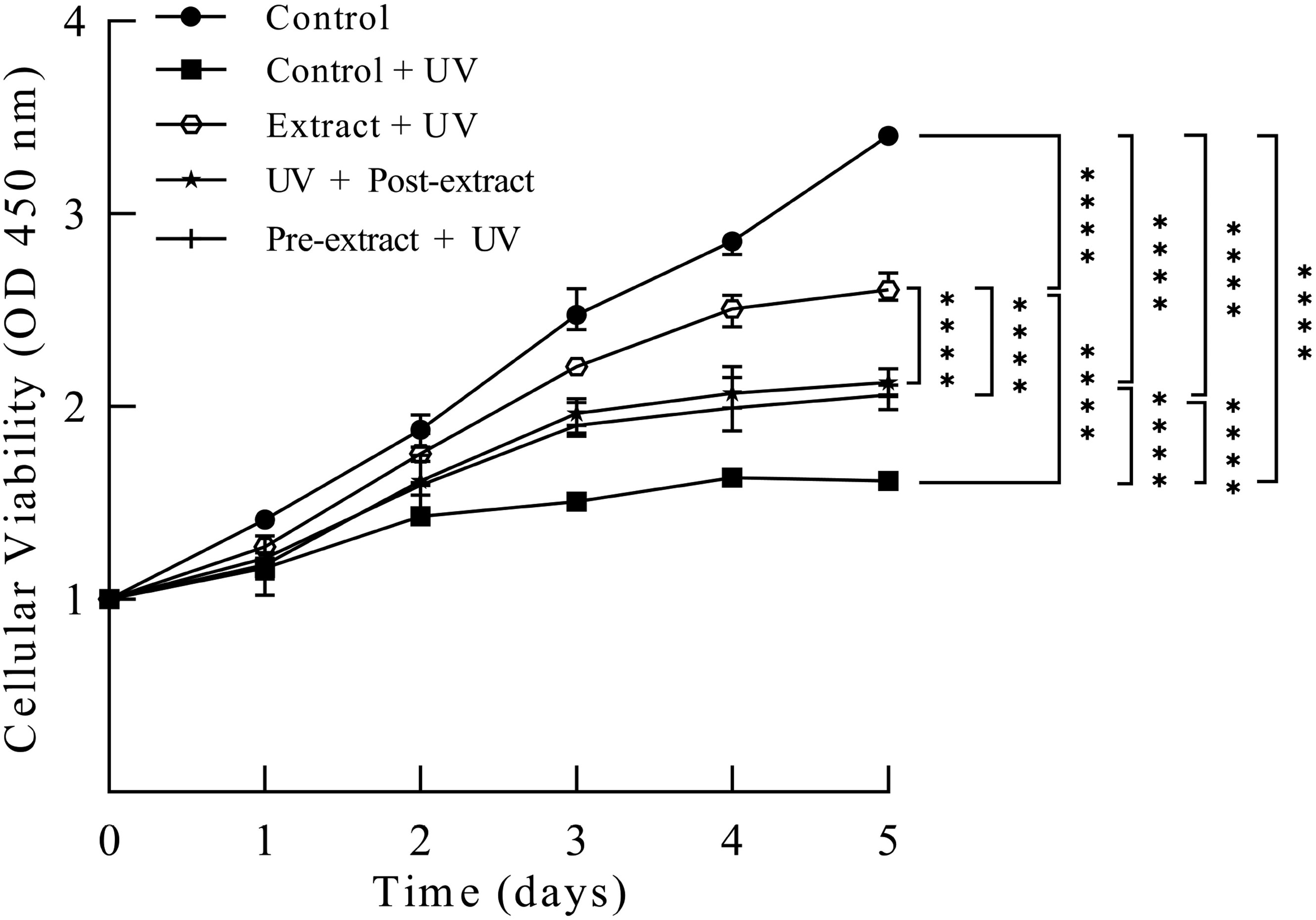
Cellular viability after ultraviolet (UV) irradiation treatment in different modes.
These results indicated that the UV protection mechanism of the extract such as antioxidant might not only exist outside the cells but also inside the cells. The instant protective effect of the extract was more important when the cells were exposed to UV irradiation. The protective effect of the extract added after UV irradiation on damaged cells suggested the protective mechanism of the extract might also be involved in the promotion of recovery.
The control group represented the use of medium with an equal amount of solvent and no UV irradiation. The control + UV group received extra UV irradiation compared with the control group. The extract + UV group contained 0.4 mg/mL extract in medium while being irradiated by UV, after which fresh culture medium was replaced, while the pre-extract + UV group was pre-incubated with the extract for 4 h until UV irradiation. In the UV + post-extract group, the extract was added into the medium after UV irradiation and cultured for another 24 h. The proliferation level of surviving cells after these treatments was measured by CCK-8 reagent.
In addition to the proliferation detection, we also examined cell death through the apoptosis kit to support further the protective effect of the extract on HaCaT against UV damage. The extract culture and UV irradiation were carried out, and the HaCaT survival status was measured using apoptosis detection staining and flow cytometry on the second day after UV irradiation.
Similar to the proliferation detection, the extract presented in the culture medium during UV irradiation protected cells from death, so did the extract pre-cultured and post-cultured treatment groups (Figure 6). The extract-UV synchronized group had 89.81% living cells, the pre-extract + UV group had 83.78% living cells, and the UV+ post-extract group had 73.39% living cells; therefore, the extract-UV synchronized group had the best protective effect. The preprocessing of the extract after UV irradiation showed an effective protection that approximated the pre-extract cultured group except for more early apoptotic cells, which reflected a progressive cell loss following the UV damage. These results further supported the instant protection effect of the extract on HaCaT against UV irradiation in both extracellular and intracellular environments. At the same time, it also showed that the restoration value of the extract after UV damage can not be underestimated.
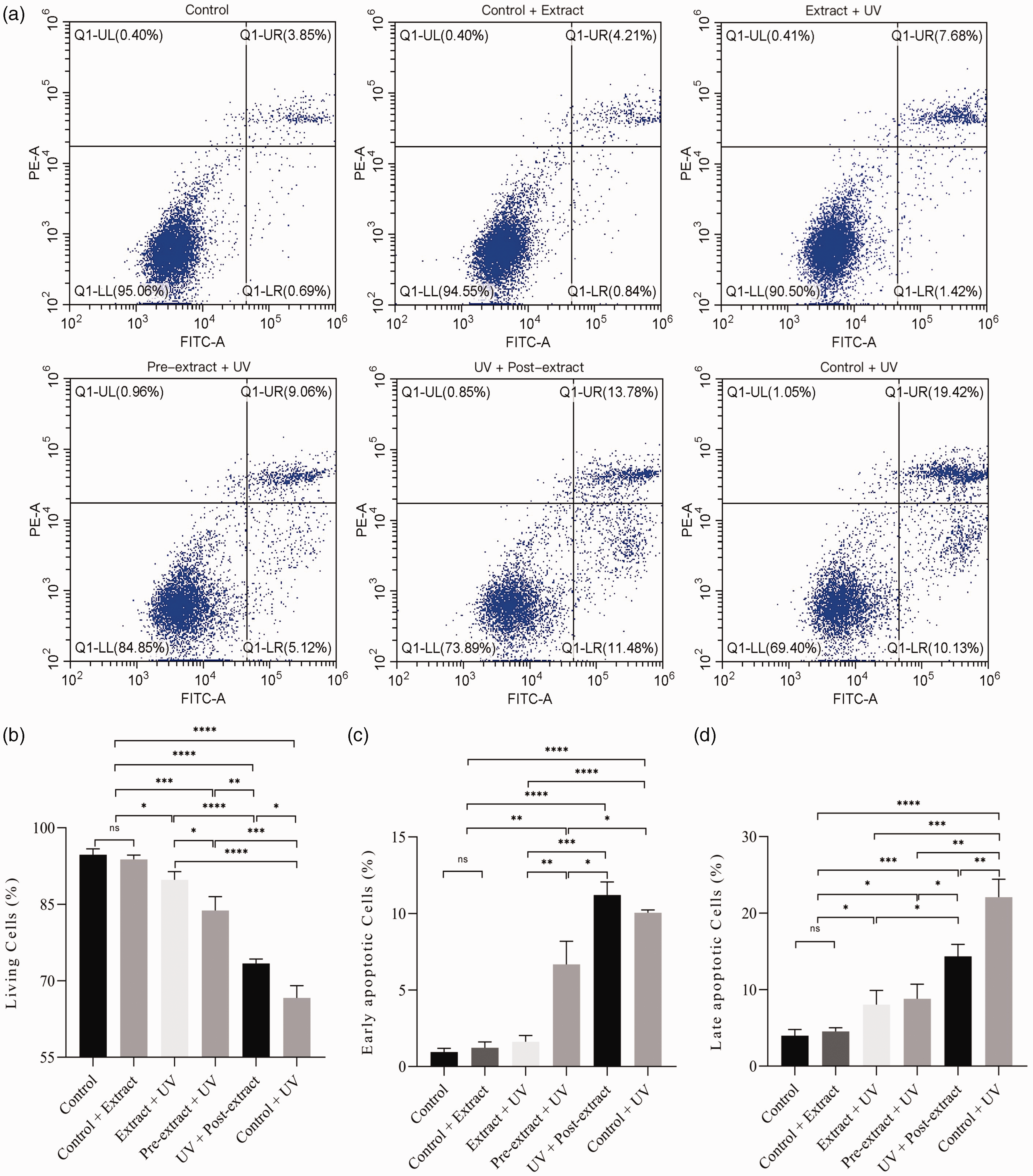
HaCaT cell apoptosis by flow cytometry. (a) Flow cytometry apoptosis of cells; (b) living cells; (c) early apoptotic cells and (d) late apoptotic cells.
Extracts from green silk cocoons are mainly flavonoid substances, including quercetin, kaempferol, and their metabolism products.6,12,16 Flavonoids have been reported to have good antioxidant, anti-sugar, anti-inflammatory, and antitumor biological activities. The content of the flavonoids in green silk cocoons was found to be 12–24 times more than that in the conventional white variety.1,15 Our study suggested that it had anti-ultraviolet ability both inside and outside the cell.
The extract reduced UV damage to DNA of HaCaT
UV damage can lead to cell DNA cross-linking and breakage, and further lead to cell cycle arrest and apoptosis.31,32 Therefore, to study further the damage and division of cells after UV irradiation, we used cell cycle detection to observe the change of the DNA copy number of experimental cells. The change of the DNA copy number of HaCaT was measured using the cell cycle test kit and flow cytometry on the second day after UV and extract treatments planned in this subject.
As a normal keratinocyte with relatively slow proliferation and obvious contact inhibition, HaCaT was mostly in the diploid stage of the nonproliferating state after division, and partly in the predivision tetraploid stage. UV radiation and DNA damage led to the inhibition of cell division, which resulted in a large number of cells stagnating in the tetraploid phase without division. The extract significantly mitigated this phenomenon by protecting the cells against UV damage. Meanwhile, cells exposed to more UV radiation damage produced more genome fragments and cross-linked DNA, which showed series of signals below diploid (Figure 7). Consistent with the results of cell proliferation and death, UV irradiation concurred with the extract addition, and had the least effect on the cell cycle, as well as the least DNA cross-linking and fragmentation. The addition of extract before and after UV irradiation can similarly reduce the inhibition of cell cycle and DNA damage caused by UV irradiation.
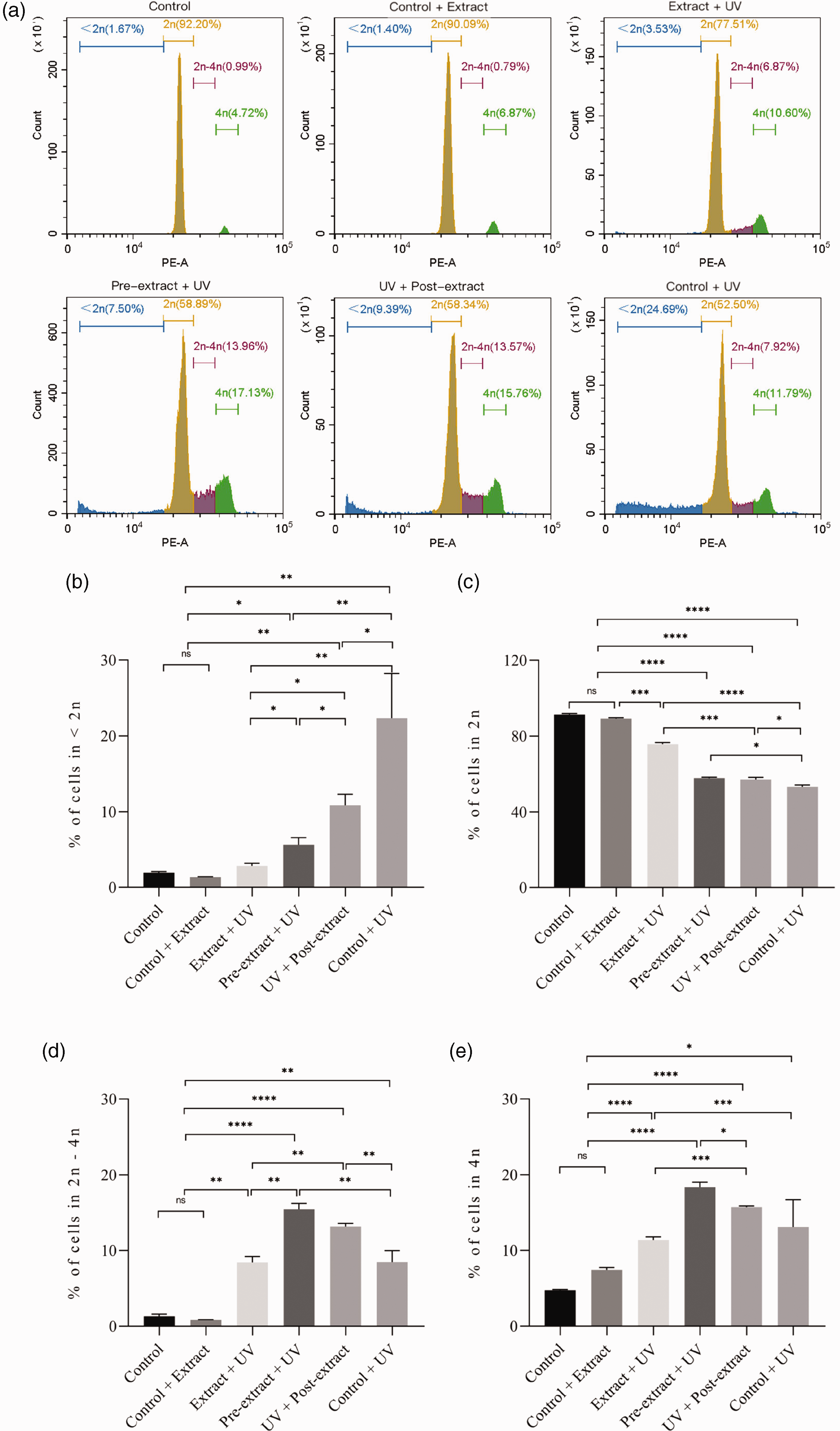
HaCaT cell cycle by flow cytometry. (a) The change of DNA copy number of HaCaT; (b) % of cells in <2 n; (c) % of cells in 2 n; (d) % of cells in 2 n–4 n and (e) % of cells in 4 n.
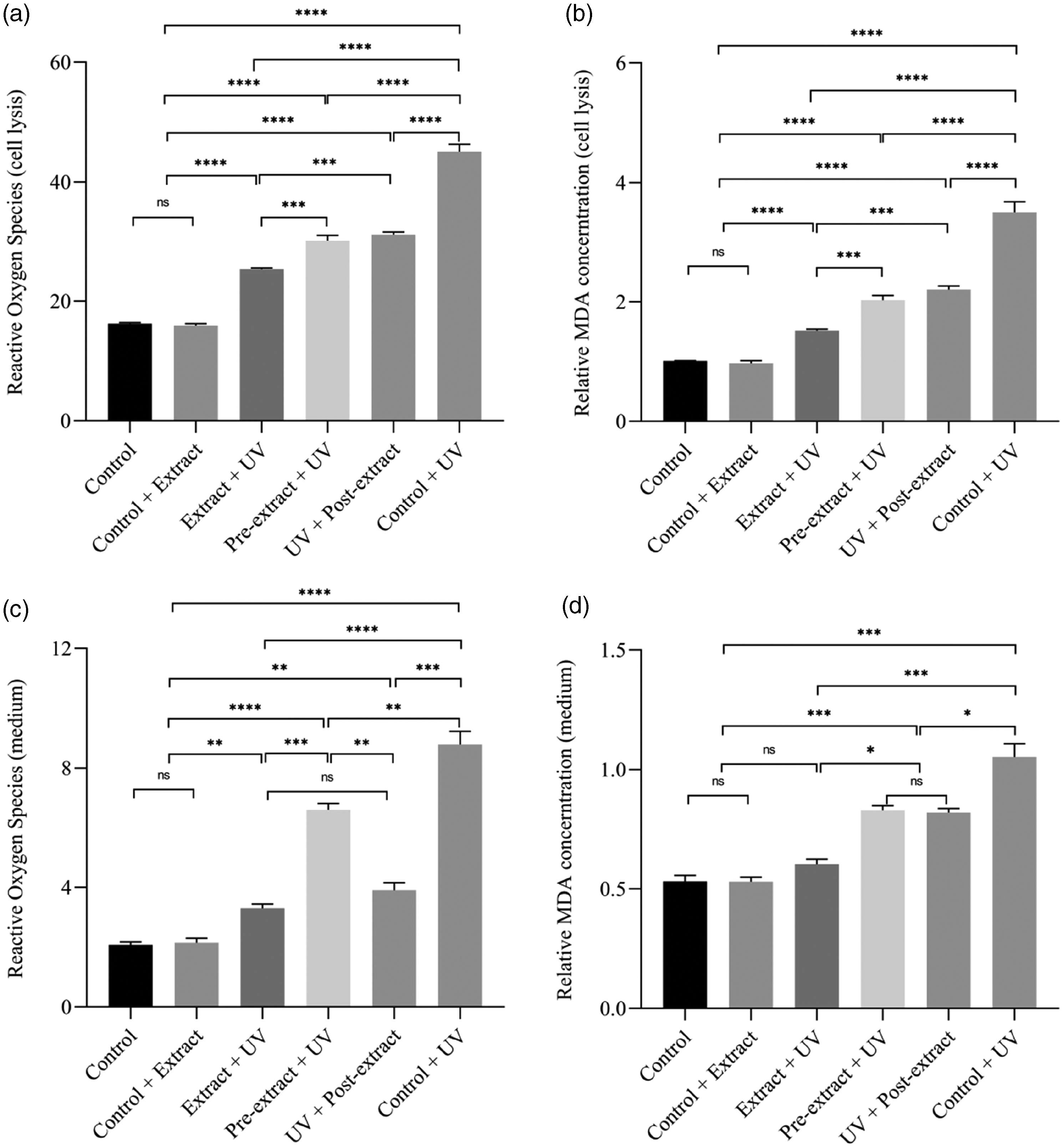
Reactive oxygen species (ROS) and malondialdehyde (MDA) contents in HaCaT cells. (a) ROS in cell lysis; (b) relative MDA concentration in cell lysis; (c) ROS in medium and (d) ROS in medium.
This protective effect existed not only during UV damage, but also after the damage occurred. The post-UV damage protective mechanism we found might be related to the repair promotion of the extract on the damaged cells, which might be a new research direction of the biological function of flavonoid compounds with medical research value.
ROS and MDA changing in HaCaT cells
Flavonoids extracted from green silk cocoons are important antioxidants, which play an important role in the regulation of oxides inside and outside cells during UV irradiation. ROS and MDA were detected as important oxidation markers in living organisms. After the extract culture and UV irradiation, the content of ROS and MDA in culture medium concentrate and cell lysis were detected, respectively. The extract in the culture medium could significantly reduce the ROS increase in the medium after UV irradiation, while the variation range of MDA in the medium was lower. The changes of intracellular ROS and MDA were consistent with phenotypic tests such as proliferation, cell death status as well as DNA damage. These results further demonstrated that the extract could exert antioxidant effects both inside and outside cells, and protect cells from UV damage, but its deep mechanism and other pathways remain to be studied.
The green silk cocoon has a high level of flavonoid extract, and the extract may certainly release slowly when the green silk cocoon manufactured dress was worn, indicating naturally green-colored domestic silk; the dress, of course, has the protective potential of skin cells against UV damage. However, as a clothing fabric, the washing times of silk material are limited. Our study found that when being washed for 15 times, the sericin layer is basically lost, and leaves a 3% DPPH clearance rate. Also, with more detailed and in-depth research, green silk cocoon extract may have the possibility of wide application in medical and safety health fields. In a previous study, naturally green silk antioxidant components were found to be able to migrate to the skin, and the migration amount of antioxidant components was related to the skin contact time, humidity, and acid-base properties of the sample. The friction between human skin and textiles, the relatively high temperature of human skin, and the fluidity of tissue fluids help to penetrate antioxidant components into the skin. 26 In order to achieve the anti-UV effect in in vitro experiments, flavonoids from silk can be concentrated in microcapsules and added to textile dressings.
Conclusions
The green silk cocoon extract had no obvious toxicity to skin cells below 0.5 mg/mL. When the concentration reached 0.4 mg/mL, the extract can protect skin cells from damage caused by UV irradiation, and decrease cell death by 23.16%. Meanwhile, pre-ingestion of the extract also had a certain protective effect of skin cells against UV irradiation, which decreased 17.13% of cell death. The green silk cocoon extract can also promote cell repair, and reduce 6.74% of cell loss of UV damaged cells. Overall, green silk has important research value in the field of UV protection and restoration.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by the National Natural Science Foundation of China (no. 51903220).