
Abstract
This study presents a comparative evaluation of chitosans derived from fungi (Agaricus bisporus) and mealworms (Tenebrio molitor) as sustainable antibacterial finishing agents for textile applications. The structural properties of fungal and insect chitosans were determined using Fourier transform infrared (FTIR) spectroscopy, scanning electron microscopy (SEM), thermogravimetric analysis, and elemental analysis. The applicability of fungal and insect chitosans to cotton fabrics, the antibacterial activity they impart to the fabric, and the durability of this antibacterial activity after washing were evaluated using the standardized Japanese Industry Standard L 1902 methodology. The analysis results showed that mealworm chitosan exhibited higher thermal stability (maximum degradation temperature of 325°C for mealworm and 309°C for fungal) and a higher degree of deacetylation (79.65% for mealworm and 8.7% for fungal) than fungal chitosan. FTIR spectroscopy and SEM analyses demonstrated that fungal and insect chitosans were physically bound to the fabric surface. Both chitosans were observed to impart good antibacterial activity (A = 3.86 for mealworm and A = 3.61 for fungal; ≥99.9% reduction) to the fabric surface. Consequently, this study demonstrates the usability of alternative chitosan sources in the textile industry while also identifying the limitations of these systems in terms of wash resistance, thereby providing a concrete roadmap for future research.

Keywords
Natural fibers such as cotton, hemp, and silk are widely used in furniture, medical treatment, clothing, and other fields due to their lightness, softness, and biodegradability. 1 Cotton textiles are susceptible to microbial growth, which can lead to odor, staining, and fabric degradation. The moisture-absorbing properties of cotton textiles create favorable conditions for microorganisms and pose potential health risks. 2 Pathogenic bacteria can be transferred from contaminated textiles and clothing to the skin via sweat, where they can colonize 3 and cause allergic sensitivities, eczema, or asthma. 4 Interest in antibacterial textiles has increased, particularly since the COVID-19 pandemic, due to heightened public awareness of hygiene and a significant rise in infections. 5 Antibacterial textiles are of great importance in protecting human health and in medical care, as they help prevent the growth and proliferation of microorganisms such as bacteria and fungi; this, in turn, helps reduce the incidence of infectious diseases. 6 Today, the production of antimicrobial textiles is one of the most promising areas for new textile materials. In particular, antibacterial properties are among the preferred comfort features in colored textile products.
In textile products, the use of many antimicrobial agents is avoided due to their potential harmful or toxic effects. 7 Therefore, the textile sector has turned to natural products that can offer various opportunities and benefits for both producers and consumers, reducing the use of synthetic chemicals and minimizing harmful environmental impacts.8-10 Research has shown that natural materials such as walnut shells, pomegranate peels, and madder root can be used as both natural dyes and antibacterial agents.11-14 In addition, the use of sustainable natural biomaterials such as chitosan, obtained from different sources, has gained increasing attention in recent years.15,16
Chitosan is a cationic polymer composed of N-acetyl-D-glucosamine and D-glucosamine units. 17 It is produced by the deacetylation of chitin, which is a component of the outer skeletons of insects and crustaceans and the cell walls of fungi. 18 Chitosan has high solubility in weak acids. In addition to being biocompatible and biodegradable, it is a polymer with high bacteriostatic and antimicrobial activity thanks to its cationic structure. 19 It is considered a potential antibacterial agent for various biomedical and pharmacological applications. 20 The performance of chitosan has been examined in various studies on the development of antibacterial textiles.16,18,21
Although chitosan is generally obtained from crustaceans, in recent years alternative chitosan sources have been sought for various reasons. Insufficiently purified shrimp chitosan may cause allergic reactions in sensitive individuals due to the tropomyosin protein residues it contains. 22 In addition, finding alternative chitosan sources has become important for vegetarians and vegans who do not consume animal-derived chitosan, as well as for people complying with religious restrictions. 23 Fungal chitosan is known to not contain protein residues that cause allergic reactions and is a vegan-friendly material. 19 Additionally, mushrooms have attracted significant attention as a nonanimal chitosan source among vegans. Compared to crustaceans, insects have short life cycles and high reproductive rates and can be easily mass-reared. 24 This makes insects a new ecologically and economically sustainable source of chitosan. 20
Every year, millions of tons of mushrooms are produced worldwide, and tons of waste are generated that can be used in chitosan production.23,25 Considering that approximately 1–15% of the mass of the cell walls of fungi is chitin, 26 fungal waste appears to have great potential for chitosan production. 19 The most widely cultivated mushroom, Agaricus bisporus, is suggested as an alternative chitin source due to its high production capacity (32% of total global production). 27
Insect breeding farms are being established on an industrial scale around the world for purposes such as disposing of organic waste and finding new protein sources for animal and human diets.24,28,29 Insects must constantly renew their exoskeletons to grow from larva to adult. Since the exoskeletons of insects have a high chitin content, they are suggested as a new source of chitin and chitosan. 28 Thus, chitin-rich waste biomass produced in insect farms constitutes a potential source for chitosan production.20,24 The yellow mealworm (Tenebrio molitor), which has a high growth rate and reproductive capacity, has recently attracted attention as a new potential source of protein for food and animal feed.29,30 Considering that T. molitor larvae have a high chitin content (18.01%), they have been suggested for use in the production of chitin and chitosan. 20
Although there are numerous studies on the extraction of chitin from fungi and insects, which are considered to be alternative sources to crustacean-derived chitin, as well as on the production of chitosan and its properties, studies on the applications of insect- and fungus-derived chitosans are limited. 17 The aim of this study is to comparatively examine the applicability of chitosan obtained from fungi and mealworms as a natural antibacterial finishing agent for textile materials. In this study, chitosan samples were characterized using Fourier transform infrared (FTIR) spectroscopy, scanning electron microscopy (SEM), thermogravimetric analysis (TGA), and elemental analysis and were then applied to white cotton fabric using the impregnation method. The antibacterial activities of the treated fabrics against Staphylococcus aureus were evaluated before and after washing according to the Japanese Industry Standard (JIS) L 1902 method.
Materials and methods
White cotton, from which sizing, oil, and other impurities had been removed and whose hydrophilicity had been increased, was coated in chitosan to produce the test samples. The fabric was 140 cm wide and weighed 75 g/m2.
Chitin extraction and chitosan production from mushrooms and mealworms
Chitin was isolated from the mushroom A. bisporus according to the method used by Ifuku et al., 31 with some modifications. Mushrooms were cleaned by washing and dried at room temperature for 1 week. First, the dried mushrooms were blended and soaked in 1 L of water overnight. Then, they were treated with 2.5% NaOH at 100°C for 16.5 h and filtered after washing with distilled water. Next, the material was kept in 350 mL of 2M HCl at room temperature for 2 days. After that, it was treated with a mixture containing 0.5% NaClO2 and 1% acetic acid for 1 day. Finally, it was treated with 2% NaOH at 100°C for 19 h and washed and filtered again. The resulting chitin was dried in an oven at 55°C. Dry chitin was deacetylated using 65% NaOH at 150°C and 600 r/min for 5 h. After that, the produced chitosan was washed, filtered, and dried at 55°C for 2 days. Mealworms (T. molitor) were cleaned by washing and dried in an oven at 45°C for a few days. First, the dried mealworms were treated with 5% NaOH at 70°C for 1 h and then washed and filtered. Next, they were refluxed with 2M HCl at 70°C while stirring at 600 r/min for 5 h. After that, the residue was filtered and washed with distilled water. Finally, it was treated with 2M NaOH at 80°C and 700 r/min for 21 h. The resulting chitin was filtered again and dried at 50°C for 2 days.
Mealworm-derived chitin was deacetylated using 70% NaOH at 800 r/min for 4 h. It was then washed and filtered through filter paper. After that, it was decolorized using a mixture of distilled water, methanol, and chloroform (4:2:1, v/v/v) at 700 r/min and 30°C for 1 h. Finally, the obtained chitosan was washed, filtered, and dried at 50°C for 2 days.
Coating of white calico fabrics with chitosan gel
Chitosan was dissolved in 1% (v/v) acetic acid to obtain a 10% (w/v) solution. 15 The coating solution was prepared by stirring at 40°C and 500 r/min. The fabric was then weighed using a precision balance. After determining the initial weight, the fabric was impregnated with the prepared chitosan solution and passed through rollers to remove excess solution. The fabrics were weighed immediately after impregnation to calculate the pickup amount. The pick-up was found to be 60%. Chitosan-coated fabric samples were dried at 100°C and cured at 110°C for 1 min. Wet and dry fabric weights were used to calculate the percentage squeezing pressure of the rollers.
Fourier transform infrared spectroscopy
FTIR spectra of the mealworm and fungal chitosan and the chitosan-coated white calico fabrics were recorded in the spectral range of 4000–625 cm–1 using a PerkinElmer Attenuated Total Reflectance-Fourier Transform Infrared (ATR-FTIR) spectrometer.
Scanning electron microscopy
Surface images of mealworm and fungal chitosan and chitosan-coated white calico fabrics were examined at different magnifications using a scanning electron microscope (FEI QUANTA FEG 250). Additionally, Energy-dispersive X-ray diffraction (EDX) analyses of the chitosan-coated white calico fabrics were performed.
Elemental analysis
An elemental analyzer (Thermo Finnigan Flash EA 1112) was used to determine the percentage of carbon (C), hydrogen (H), and nitrogen (N) in the mealworm, and the fungal chitosan samples. The degree of deacetylation (DD) of the chitosan samples was determined using the following equation: 32
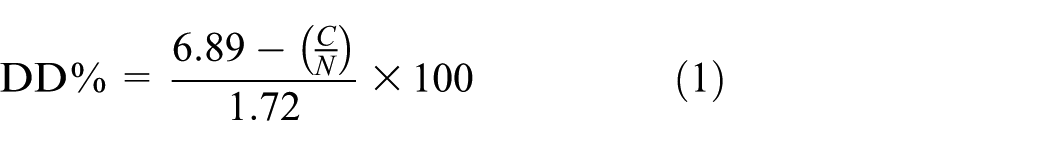
where DD% is the degree of deacetylation and C/N is the carbon-to-nitrogen ratio derived from the elemental analysis of the chitosan samples.
Thermogravimetric analysis
The thermal stability of mealworm- and mushroom-derived chitosans was determined using a TGA analyzer (PerkinElmer Pyris 1). The samples were heated up to 650°C in a nitrogen atmosphere at a heating rate of 10°C/min.
Determining the antibacterial efficiency
The JIS L 1902:2015 (Measurement of Antibacterial Activity of Textiles) was used to determine the efficacy of chitosan-coated fabrics in reducing bacterial growth. 33 The antibacterial activity was tested against the pathogenic bacteria S. aureus (ATCC 6538 Gram (+)) and the antimicrobial assay was performed according to Erdoğan and Becenen. 18 Firstly, chitosan-coated fabrics (0.4 g) were sterilized in test tubes at 121°C for 15 min prior to the experiment. Then, the fabrics were inoculated with 0.2 mL of S. aureus at a concentration of 3 × 105 Colony-Forming Units (CFU). Initial bacterial concentrations were measured at time zero immediately after inoculation. After that, both control and test specimens were inoculated with S. aureus and incubated in closed vials at 37°C for approximately 24 h. The final microbial concentrations were measured after incubation. The reduction in bacterial growth was determined based on the control sample and initial concentrations. The coated fabrics were washed once and then tumble dried at 40°C, 34 after which they were tested again for antibacterial efficacy.
The bacterial growth rate was determined using the following formula for the control sample:
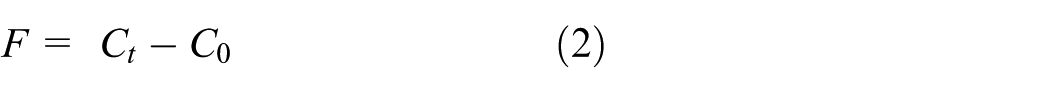
The bacterial growth rate for the chitosan-coated fabrics was determined using
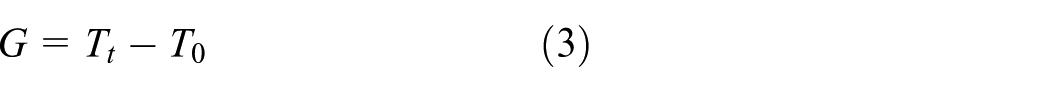
The following formula was used to calculate the antibacterial efficiency value:

In the above, A is antibacterial activity, C0 the logarithm of the number of bacteria in the control sample right after inoculation, Ct the logarithm of the number of bacteria in the control sample after a 24 h incubation, T0 the logarithm of the number of bacteria in the test specimen right after inoculation, and Tt the logarithm of the number of bacteria in the test specimen after a 24 h incubation. Regarding antibacterial efficacy, A < 2 denotes no efficacy, 2 ≤A <3 denotes efficacy, A≥3 denotes full efficacy.
Results and discussion
Physicochemical properties of the mealworm and the fungal chitosan
FTIR spectroscopy
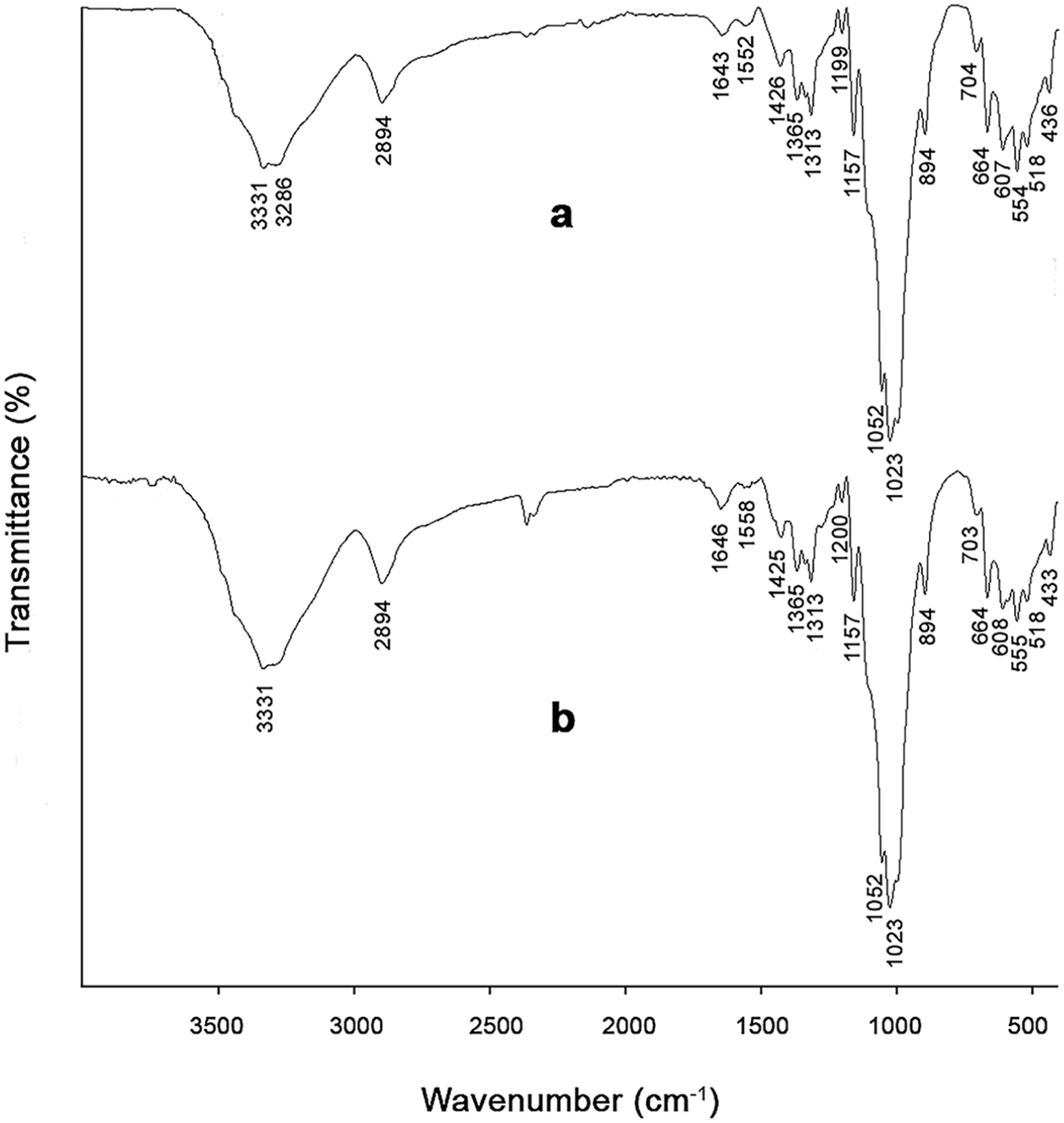
In the FTIR spectra, chitosan-specific carbonyl absorption peaks (C=O stretching) were observed at 1647 and 1646 cm–1, and amide II absorption peaks (N–H bending) were observed at 1583 and 1586 cm–1 for mealworm and fungal chitosans, respectively (Figures 1(a) and 1(b)). When the FTIR spectra of insect and fungal chitosans obtained in this study were compared with commercial chitosan data reported in the literature, it was observed that both chitosan types were highly consistent in terms of chemical structure with other fungal and insect chitosans,35,36 as well as with commercial chitosan.32,37,38 In particular, the presence of amide I (∼1650 cm–1), amide II (∼1585 cm–1), C–O–C (∼1150 cm–1), and β-glycosidic bond (∼894 cm–1) bands confirms that both bioderived chitosans retain the characteristic chitosan structure. However, some band shifts and intensity differences reflecting differences in chitosan source and structure were also observed. Accordingly, peaks corresponding to O–H and N–H stretching vibrations of overlapping –OH and –NH2 groups were observed around 3425, 3359, and 3292 cm–1 in insect chitosan, 3360 and 3291 cm–1 in fungal chitosan, and approximately 3354–3363 and 3291–3292 cm–1 in commercial chitosan.32,38 Overall, both biosourced chitosans showed a chemical structure highly similar to that of commercial chitosan. Peaks attributed to aliphatic C–H stretching vibrations corresponding to –CH and –CH2 groups in the polysaccharide backbone were observed around 2874 cm–1 for insect chitosan, 2917 and 2870 cm–1 for fungal chitosan, and approximately 2916 and 2870 cm–1 for commercial chitosan. 37 Fungal chitosan, in particular, showed a very good overlap with commercial chitosan. 38 Amide I and amide II bands, representing residual N-acetyl groups, were recorded at around 1647 and 1583 cm–1, 1646 and 1586 cm–1, and approximately 1650–1656 and 1584–1590 cm–1 for insect chitosan, fungal chitosan, and commercial chitosan, respectively. 32 These bands in insect- and fungus-derived chitosan are in good agreement with the commercial chitosan ranges, and the preservation of peak positions indicates that the characteristic structure of chitosan is retained in both samples. Bands corresponding to CH2 and CH3 deformation vibrations and C–N stretching were observed at 1421 and 1376 cm–1 in insect chitosan, 1455, 1419, 1375, and 1320 cm–1 in fungal chitosan, and approximately 1422, 1377, and 1320 cm–1 in commercial chitosan, 37 and were found to be conserved across all samples. However, the band at 1455 cm–1 was slightly more prominent in fungal chitosan, possibly due to structural impurities. A strong overlap was observed among all three chitosan types in terms of C–O–C and C–O stretching vibrations (insect: 1149, 1077, 1028 cm–1; mushroom: 1150, 1059, 1027 cm–1; commercial: ∼1150, ∼1062, ∼1022–1027 cm–1), corresponding to glycosidic (β-(1→4)) linkages in the polysaccharide backbone. 32
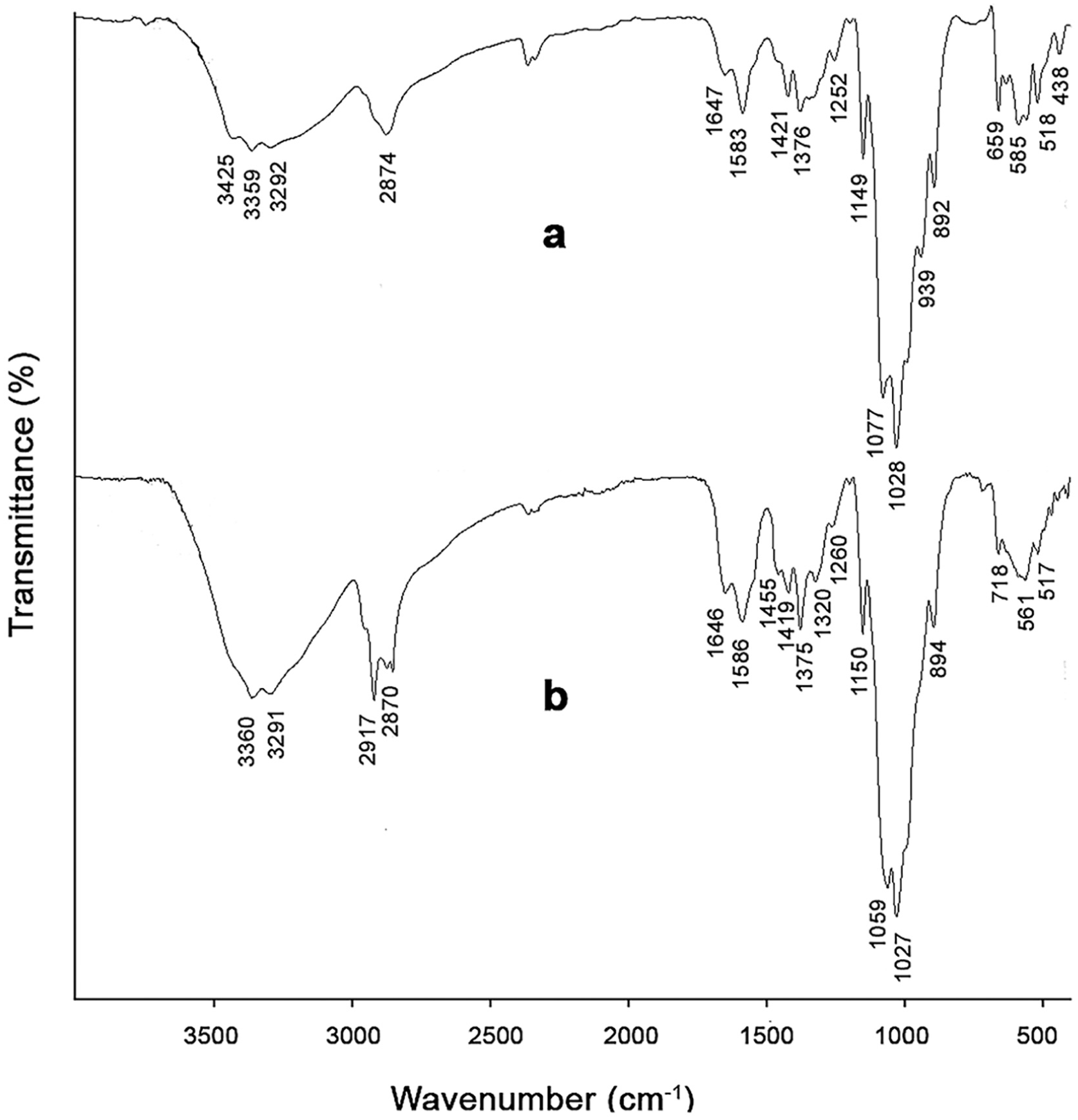
Fourier transform infrared spectra of chitosan samples: (a) mealworm chitosan, (b) fungal chitosan.
In conclusion, mealworm and fungal chitosan exhibited FTIR spectra that were highly consistent with those of commercial chitosan. The characteristic amide I and amide II bands appeared at approximately 1650 and 1585 cm–1, respectively. The preservation of key polysaccharide backbone vibrations (1150–1020 cm–1) and β-glycosidic bonds (∼894 cm–1) confirms the structural integrity of chitosan. The minor peak shifts are likely due to differences in pigment residues in insect-derived chitosan and β-glucan residues in fungal chitosan, as well as variations in degree of deacetylation, molecular weight, and source-dependent hydrogen bonding interactions.
SEM
Mealworm chitosan exhibits a structure consisting of densely packed, small pores, while also containing fine fibrous features (Figure 2(a)). In contrast, fungal chitosan shows a structure in which the fiber layer is interwoven with a β-glucan layer, and no distinct fibrous or pore-like structures are observed (Figure 2(b)). The surface morphologies of mealworm- and fungus-derived chitosans in this study reveal significant similarities and differences compared with commercial chitosan. Insect chitosan contains fine fibrous structures along with a densely located structure of small pores. This morphology is consistent with the nanofibrous and porous structures of commercial chitosans derived from shrimp and crayfish.15,39 However, the denser and more frequent pore distribution in insect chitosan differs from that of crustacean chitosans, which exhibit wider spacing and larger pore diameters. Fungal chitosan, on the other hand, shows a distinctly different morphology from both insect and commercial chitosans. It displays a compact structure in which the fiber layer is interwoven with β-glucan, and no distinct pores or fibrous structures are visible. Therefore, fungal chitosan differs from porous commercial chitosans in this respect.37,38 However, the surface morphologies of mealworm- and fungus-derived chitosans in this study are consistent with those reported in the literature for insect and fungal chitosans.24,36 Consequently, while insect chitosan shows similarity to commercial chitosan in terms of its porous and fibrous structure, fungal chitosan exhibits a different structural character due to its more compact and nonporous morphology.

Surface images of the chitosan samples: (a) mealworm, (b) mushroom.
TGA
While mealworm chitosan underwent mass loss in two stages, fungal chitosan exhibited an additional mass loss (Figure 3). Mealworm chitosan lost 7.473% of its mass at temperatures up to 125°C due to moisture loss, and between 125°C and 650°C, it lost 62.010% of its mass due to the decomposition of the chitosan polymer. The residual amount of mealworm chitosan was 27.487% at 650°C (Figure 3(a)). The mass loss of fungal chitosan due to water loss in the first stage was around 3% (up to 150°C). Fungal chitosan started to decompose after 200°C and lost 55.475% of its mass due to the degradation of the chitosan polymer up to 425°C. Afterward, it had an additional mass loss between 425°C and 650°C, losing 13.130% of its mass. The residual amount of the fungal chitosan was determined as 31.247% at 650°C (Figure 3(b)).
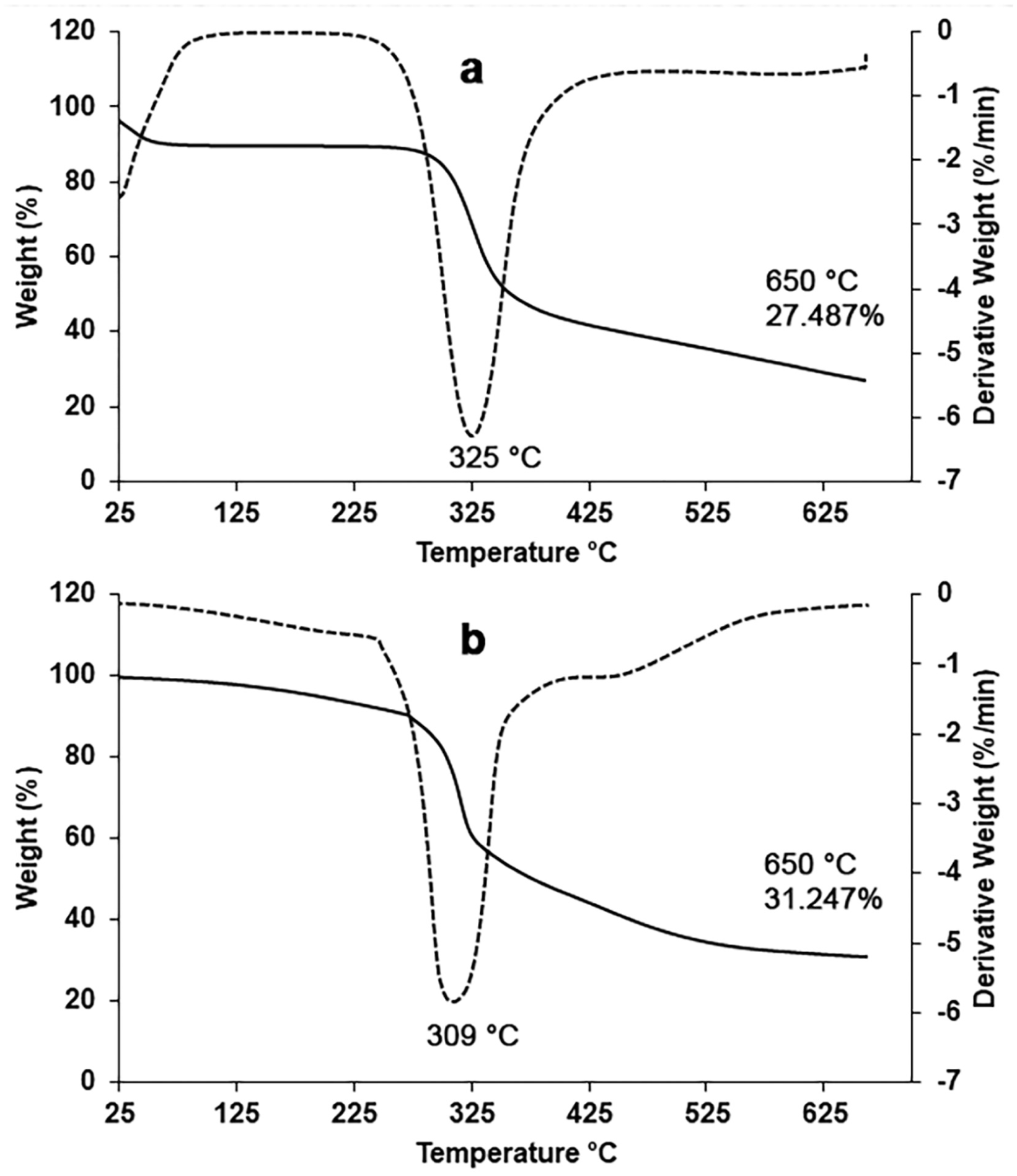
Thermogravimetric analysis curves of chitosan samples: (a) mealworm chitosan, (b) fungal chitosan.
Comparison based on literature data shows significant differences between the thermal stability of insect and fungal chitosans in this study and that of commercial chitosan. The mealworm chitosan in this study exhibits thermal behavior similar to that of commercial chitosan, with a two-stage degradation profile and significant mass loss at high temperatures. The fungal chitosan in this study has an additional degradation step and exhibits a three-stage degradation behavior. The additional degradation step of mushroom chitosan points to a more heterogeneous or complex structure. This likely results in fungal chitosan forming a relatively higher thermal residue. It may also be related to lower crystallinity and weaker hydrogen bonds, as noted in the literature. 40 The degradation ranges of commercial chitosan are attributed to moisture loss at 50–100°C and to polymeric structure degradation at 400–500°C.32,38,41 The absence of any significant thermal event above 150°C in commercial chitosan indicates high stability at this temperature, associated with high crystallinity and strong –OH/–NH2 hydrogen bonding. In this respect, fungal chitosan differs from commercial chitosan. The thermal degradation behavior of mealworm- and fungus-derived chitosans in this study is also consistent with those reported in the literature for other insect and fungal species.23,42
In this study, the maximum degradation temperature (DTGmax) of the mealworm chitosan was measured as 325°C, while the DTGmax value of the fungal chitosan was measured as 309°C. It has been stated that the low DTGmax value of chitin and chitosan obtained from fungi is attributed to the glucan residues remaining in the chitin structure after extraction. 43 These values are consistent with those reported in the literature for various insect, fungal, and crustacean species.15,42,44
Elemental analysis
The N, C, and H contents of chitosan samples were measured as 7.12%, 39.37%, and 6.61% for mealworm chitosan and as 6.05%, 42.64 %, and 7.15% for fungal chitosan, respectively. In the literature, the N, C, and H contents of commercial chitosan are reported as 7.2%, 40.4%, and 6.4%, respectively. 32 The C/N ratio, which reflects changes in chemical composition during chitosan deacetylation, is directly related to the removal of acetyl groups (–COCH3) from the polymer chain. 45 As acetyl groups are removed from the structure, the carbon content decreases while the nitrogen content remains relatively constant. This allows the DD of chitosan to be estimated using empirical relationships. Therefore, the DDs of insect and fungal chitosans were calculated according to equation (1) using percentages of C and N obtained from elemental analysis.
The DDs were calculated as 79.65% for mealworm chitosan and 8.7% for fungal chitosan. Insect chitosan exhibits a highly deacetylated structure. This value is quite consistent with those reported in the literature for medium-molecular-weight commercial chitosan (72.6%) and crustacean chitosan (83%).32,38 The DD value of fungal chitosan is relatively low. Elemental analysis (for C, H, N), due to its widespread use in the literature, provides comparable results and determines the total C and N content in the sample with high accuracy. However, this method can be affected by moisture, inorganic content, or impurities, leading to deviations in the calculation. In this study, impurities originating from β-glucan affected the calculated value for fungal chitosan. Since β-glucan does not contain nitrogen but does contain carbon, it may have artificially increased the C/N ratio in the elemental analysis, leading to deviations in the calculation. Therefore, it is observed that the fungus-derived chitosan used in this study is significantly different from those reported in the literature and has a considerably lower DD value. 25 The DD value of the insect chitosan in this study is consistent with the literature data. 28
The DD can significantly affect the solubility, crystallinity, hydrophilicity, and bioactivity of chitosan. 20 The DD is affected by deacetylation conditions such as reaction time, NaOH concentration, and temperature, and increases at higher temperatures. 24 An increase in deacetylation time has also been reported to increase the DD of fungal chitosan. 46 Increased positively charged amino groups due to an increase in DD give chitosan its antibacterial activity. 17
Characterization and antibacterial efficacy results of the coated fabrics
FTIR spectroscopy results
The IR spectra of white calico fabrics coated with mealworm and fungal chitosans are presented in Figure 4. FTIR spectra of chitosan-coated fabrics showed limited changes in the positions and intensities of some characteristic peaks compared with those of pure mealworm- and fungus-derived chitosans. Owing to the overlap of broad –OH/–NH stretching vibrations, the broad band observed around ∼3350 cm–1 in fungal and insect chitosans shifts to a lower wavenumber (∼3300 cm–1) in chitosan-coated fabrics (Figures 1 and 4). This indicates the formation of strong hydrogen bonds between chitosan and cellulose. The shift and decrease in intensity of the C=O (amide I) band, one of the characteristic functional groups of chitosan, from ∼1647 to ∼1643 cm–1 in the chitosan-coated fabric, demonstrates intermolecular interactions (hydrogen bonding and electrostatic interactions) between chitosan and the cellulose fiber surface (Figures 1 and 4). The shift of the amide II band (around 1580 cm–1) (Figure 1), originating from chitosan’s N–H bending and C–N stretching vibrations, to approximately 1550 cm–1 and its decrease in intensity in the coated fabrics supports the idea that chitosan’s –NH2 groups form hydrogen bonds with cellulose. Similarly, peaks observed around 1376 cm–1 in the chitosan spectra were found to shift to 1365 cm–1 in the coated fabrics. In contrast, peaks associated with C–H stretching vibrations, observed at approximately 2870 cm–1, were found to shift towards 2894 cm–1 after coating, accompanied by an increase in intensity. Furthermore, shifts in the C–O–C and C–O stretching peaks in both cellulose and chitosan indicate interactions between polymer chains. These spectral changes confirm the successful incorporation of chitosan into the textile substrate. These shifts point to changes in the hydrogen-bonding structure of cellulose after coating white calico fabric with chitosan and demonstrate the physical bonding between chitosan and the fabric. 47 The preservation of chitosan-specific peaks in the FTIR spectra of the coated fabrics indicates that chitosan was successfully immobilized on the fabric surface, while the absence of a new absorption band in the spectra suggests that physical interactions (hydrogen bonds and van der Waals forces) are dominant, rather than chemical (covalent) bond formation between chitosan and cotton fibers. Furthermore, the absorption peaks exhibited by mealworm- and fungal-chitosan-coated fabrics are similar to those of cotton fabrics coated with shrimp and crayfish chitosan reported in previous work. 16
Fourier transform infrared spectra of chitosan-coated white calico fabrics: (a) mealworm-chitosan-coated fabric, (b) fungal-chitosan-coated fabric.
White calico fabric is a cellulose-based cotton woven fabric. Cellulose is a polymer with three hydroxyl groups (at the C2, C3, and C6 positions) in each glucose unit. These groups form both intramolecular and intermolecular hydrogen-bond networks.19,48 The most fundamental interaction between chitosan and cellulose is hydrogen bonding; O–H···O- and N–H···O-type hydrogen bonds are formed between the –NH2 and –OH groups of chitosan and the –OH groups of cellulose (Figure 5).3,49 Since chitosan is dissolved in acetic acid to coat the fabric, it becomes protonated in a weakly acidic environment and takes the form of –NH3+, gaining a positive charge. 17 The cellulose surface, on the other hand, generally exhibits a negative character in aqueous environments. 50 In this case, additional electrostatic attractive forces are also formed between chitosan (cationic) and cellulose (anionic).17,49
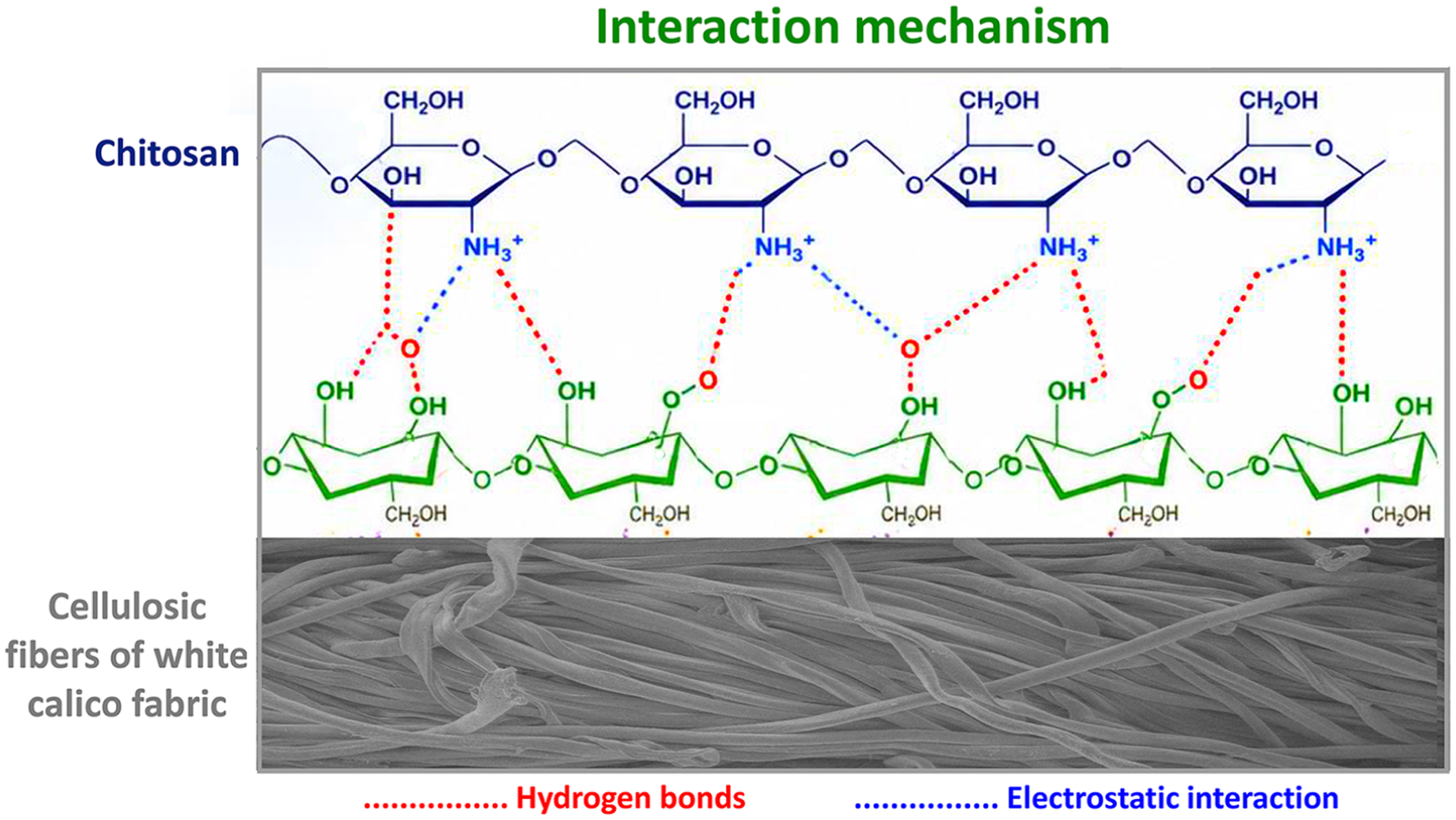
A schematic diagram illustrating the interaction mechanism between chitosan and cellulosic fibers of white calico fabric.
SEM and EDX results
The surface structure of mealworm- and fungal-chitosan-coated fabrics was examined by SEM analysis. SEM images (Figure 6) revealed a relatively smooth and clean surface for the untreated fabric, while the fibers of the chitosan-coated samples showed a tightly adhered appearance. Mealworm- and mushroom-derived chitosans were coated onto the white calico fabric as a thin layer, and no chitosan accumulation or agglomeration was observed on the fabric surface (Figures 6(c), 6(d), 6(e), and 6(f)). It is reported in the literature that a liquid chitosan solution dissolved in 1% acetic acid coats the fabric surface as a thin film layer. 18

Scanning electron microscopy images of chitosan-coated white calico fabrics: (a) and (b) uncoated fabric, (c) and (d) mealworm-chitosan-coated fabric, (e), and (f) fungal-chitosan-coated fabric.
The EDX analysis results revealed that the fungal-chitosan-coated fabric contained 45.34 wt.% C, 0.98 wt.% N, and 53.68 wt.% O, while the mealworm chitosan-coated fabric contained 45.44 wt.% C, 1.01 wt.% N, and 53.55 wt.% O. The natural cellulose structure (C6H10O5) does not contain nitrogen (Figure 7(a)). Therefore, the presence of nitrogen detected in the EDX analysis indicates the presence of fungal and insect derived chitosan (–NH2 groups) bound to the fiber surface (Figures 7(b) and 7(c)).
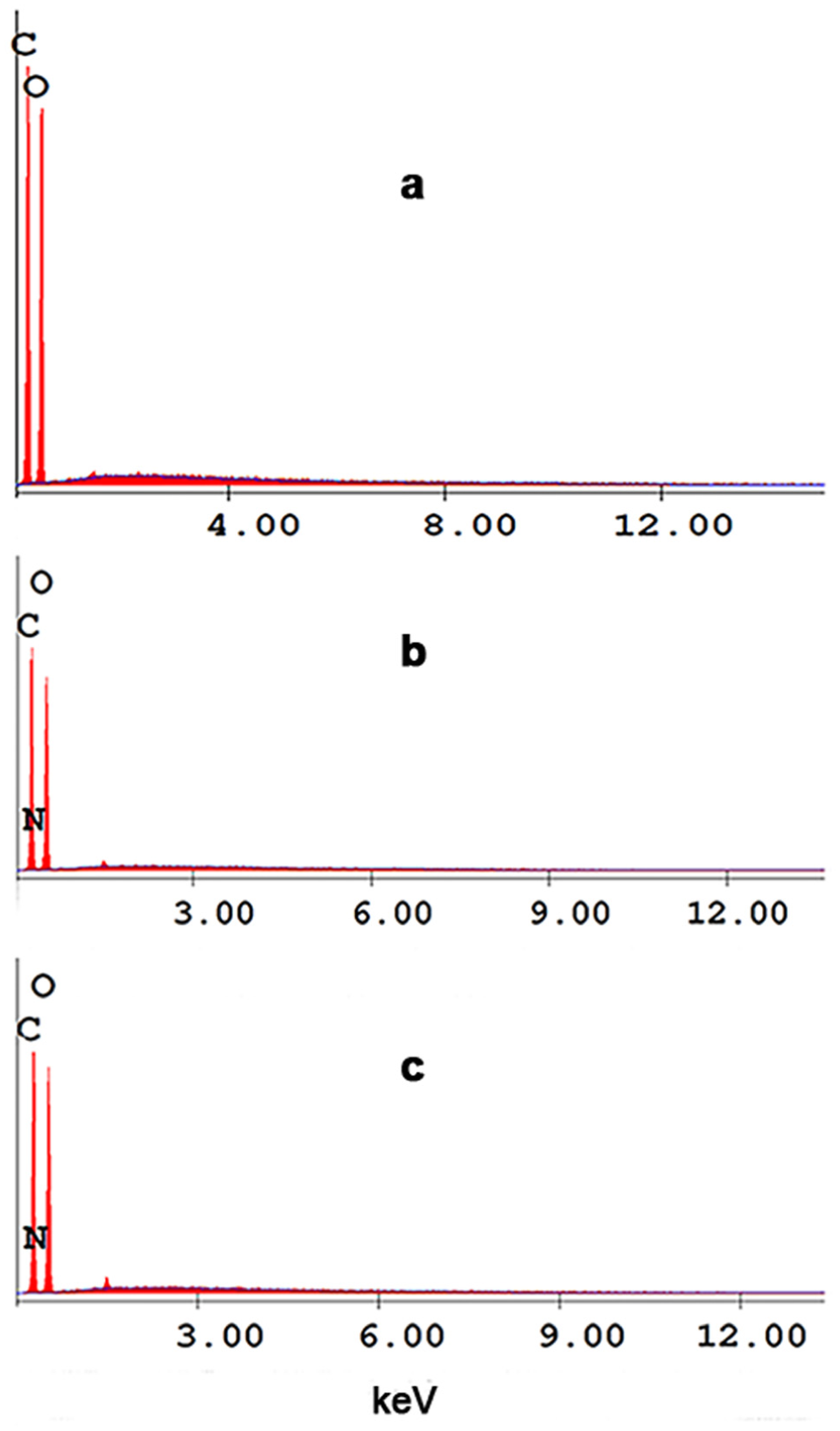
EDX graphs of coated and uncoated fabrics: (a) uncoated fabric, (b) mealworm-chitosan-coated fabric, (c) fungal-chitosan-coated fabric.
Antibacterial activity results
The JIS L 1902 challenge test measures the antimicrobial activity of a material after 24 h of direct contact with microorganisms. 33 Since the results are taken as microbial growth inhibition on a log scale, they allow a quantitative assessment of antimicrobial activity. 51 The antibacterial activity value (A) is defined as the logarithmic decrease in the number of viable bacteria in the sample after incubation (JIS L 1902:2015). Therefore, A > 3 indicates a logarithmic reduction of ≥3 (≥99.9% decrease) compared with the untreated control and reflects strong antibacterial performance.
The antibacterial activities of white cotton fabrics coated with chitosan derived from fungi and mealworms were determined after incubation with a bacterial suspension of 3 × 105 CFU/mL (Table 1). The A value, indicating the antibacterial activity of the mealworm-chitosan-coated fabric, was calculated as 3.86, while that of the fungal-chitosan-coated fabric was found to be 3.61. According to the standard classification, both values correspond to “very strong” or “fully effective” antibacterial performance (≥99.9% reduction). The A values obtained in this study show that both types of chitosan exhibited strong antibacterial activity against S. aureus, with mealworm-derived chitosan showing slightly higher performance (Table 1). Although antibacterial activity against Gram-negative bacteria was not measured using the JIS L 1902:2015 method in this study, the antibacterial effect of shrimp and crayfish derived chitosan-coated cotton fabrics against Escherichia coli ATCC 8739 was investigated in our previous studies using the same method. 16 It was found that both chitosans exhibited very strong antibacterial activity against E. coli (A = 5.74 and A = 3; A≥ 3, ≥99% reduction).
Antibacterial activity results of mealworm and fungal chitosan coated fabrics based on the Japanese Industry Standard (JIS) L 1902:2015 standard analysis method
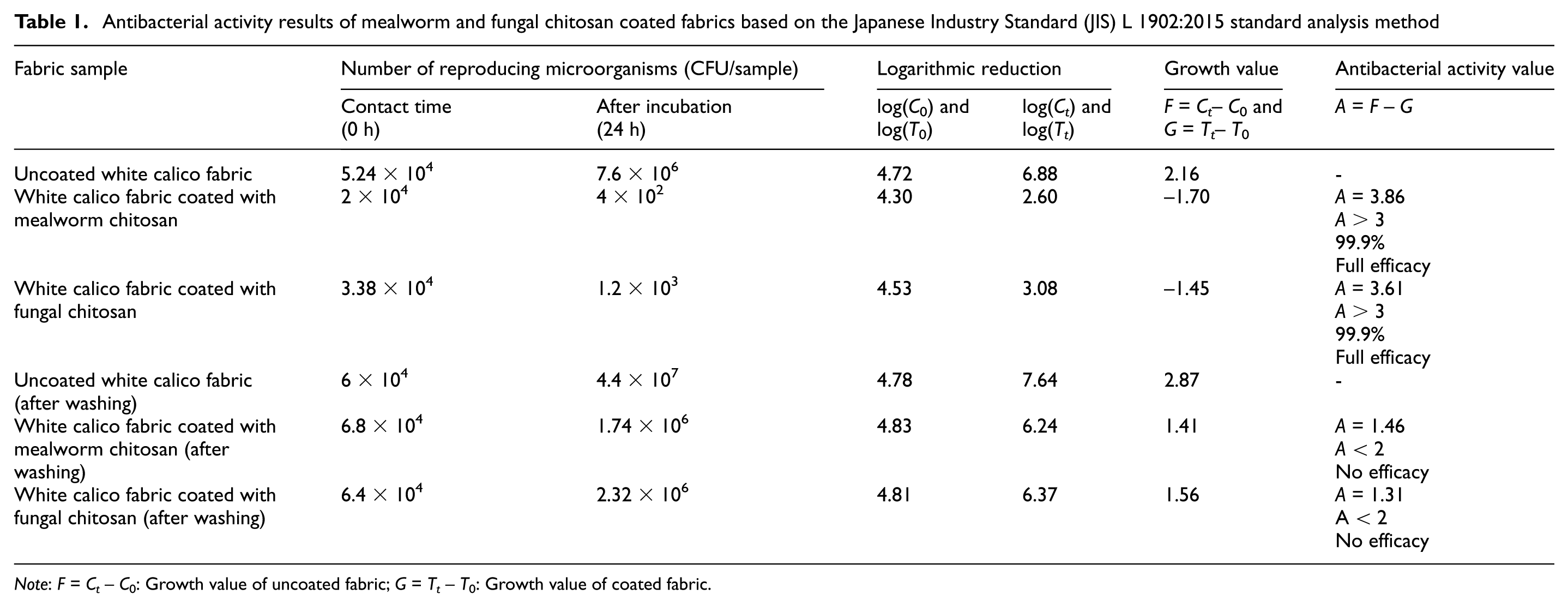
Note: F = Ct–C0: Growth value of uncoated fabric; G = Tt–T0: Growth value of coated fabric.
Antibacterial activity is also largely influenced by the structural and physicochemical properties of the chitosan source. Among these structural properties, the DD and molecular weight directly affect antibacterial activity, while other factors such as the crystallinity and concentration of chitosan, and target microorganism species also influence antibacterial activity.17,18 In general, higher DD values increase the density of protonated amino groups (–NH3+), thereby strengthening electrostatic interactions with negatively charged bacterial cell walls and enhancing antimicrobial efficacy. 52 Chitosan with a high positive charge density (DD: 97.5%) exhibited greater antibacterial activity against S. aureus compared to chitosan with a relatively lower positive charge density (DD: 83.7%). 53 However, there are also reports in the literature indicating that even low-DD chitosan systems can exhibit significant antibacterial activity depending on molecular architecture and source-specific properties. 16 In addition, Kaya et al. 32 observed that both low-molecular-weight scorpion chitosan and medium-molecular-weight commercial chitosan exhibited similar antimicrobial activities against Gram-positive and Gram-negative bacteria. In this study, the DD of insect chitosan was calculated as 79.65%, while that of fungal chitosan was calculated as 8.7%, and both chitosans were found to exhibit strong antibacterial activity against S. aureus. In addition, as demonstrated by FTIR analysis, the presence and intensity of amide and amine bands (∼1646, 1647 cm–1 and ∼1583, 1586 cm–1) in the FTIR spectra of both chitosans indicate the presence of functional groups responsible for antimicrobial activity.
Chitosan exhibits antibacterial activity through various mechanisms. The primary mechanism is electrostatic interaction. The amino groups (–NH2) present in chitosan become protonated in an acidic environment, forming –NH3+ (positively charged). These protonated –NH3+ groups interact with the negatively charged bacterial cell membranes, disrupting membrane integrity. As a result, membrane permeability is disrupted, leading to leakage of intracellular contents and ultimately bacterial cell death. Habeeba et al. 54 reported that the antibacterial activity of chitosan-coated cotton fabrics is mainly based on these electrostatic interactions. Insect-derived chitosan generally exhibits antibacterial activity through electrostatic interactions with bacterial cell membranes due to the higher availability of protonatable amino groups (DD: 79.65%). The second mechanism is that chitosan binds metal ions, thereby inhibiting bacterial enzyme activity and disrupting cellular metabolism, which results in an antibacterial effect. Low-molecular-weight chitosan can penetrate bacterial cells and inhibit essential biological processes, thereby exerting antibacterial activity. Fungal chitosan generally has a lower molecular weight and a more amorphous structure. 40 This leads to increased solubility of chitosan. As a result, fungal chitosan can more easily diffuse into bacterial cells, and its mechanism of action is not limited to the cell surface; it interacts with intracellular components, leading to a more profound antibacterial effect. The third mechanism is that chitosan, particularly when coated onto fabric surfaces, forms a film that acts as a barrier for bacteria and limits nutrient transport to bacterial cells. High-molecular-weight chitosan exerts its effect by forming a film on the bacterial cell surface. Insect chitosan generally exhibits higher molecular weight and a more crystalline structure. 40 This restricts polymer chain mobility, resulting in a more rigid structure. Consequently, instead of penetrating bacterial cells, insect chitosan tends to interact with the cell surface through electrostatic interactions.
After one washing cycle at 40°C followed by drying, a significant decrease in the antibacterial performance of chitosan-coated fabrics was observed. The A value decreased to 1.46 for mealworm-chitosan-coated fabric and to 1.31 for fungal-chitosan-coated fabric, indicating a loss of effective antibacterial activity. The reduction in antibacterial activity after washing is not related to the antimicrobial potential of fungal and insect chitosans, but rather to coating stability. As also shown by FTIR spectroscopy results, insect- and fungus-derived chitosans were attached to the cotton fabric through physical adsorption based on hydrogen bonding and electrostatic interactions; therefore, they were removed from the surface during washing, leading to a marked decrease in antibacterial performance. The results indicate that more advanced coating strategies are needed between chitosan and cotton fabric to improve the limited wash resistance of fungal and insect chitosans.
To improve the wash resistance of chitosan coatings, fiber-chitosan coating stability can be enhanced using surface activation techniques such as chemical crosslinking, layer-by-layer bonding, plasma applications, enzymatic modifications, and resin-based binding agents.54,55 These methods can improve the coating durability and consequently the antibacterial activity of chitosan. However, these applications increase the coating thickness, hindering the breathability of the fabric.
Furthermore, the use of chemical crosslinking agents increases the chemical load associated with textile waste, thus harming the environment. This study aimed to enhance the functionality of cotton fabric using the minimum amount of chemicals possible, employing an environmentally friendly approach. Although crosslinking was not systematically investigated in our study, low-toxicity, environmentally friendly crosslinking agents such as genipin, citric acid, and sodium tripolyphosphate, used in textile applications, can promote covalent bond formation between the amino (–NH2) and hydroxyl (–OH) groups of chitosan and functional groups present in textile fibers, resulting in durable coatings with high wash resistance. Future studies should systematically evaluate the relationship between crosslinking intensity and antibacterial resistance after repeated washing cycles.
Unlike previous studies that have focused primarily on chitosans derived from crustaceans, this study presents a novel comparative evaluation of fungus- and mealworm-derived chitosans, which have rarely been examined in the literature for textile applications, using a standardized antimicrobial testing methodology. 23 The study systematically demonstrates the applicability, antibacterial efficacy, and postwash durability limitations of fungus- and insect-based chitosan systems as alternatives to commercial chitosan on cotton fabrics.
Conclusion
In this study, chitosan was produced from fungi and insects, characterized, and coated onto white cotton fabric using an impregnation method. The antibacterial activity of chitosan-coated fabrics against S. aureus was measured before and after washing according to the JIS L 1902 method, and their effectiveness was compared. The data obtained from the study showed that both types of chitosan could be easily coated onto the fabric and that both chitosans provided antibacterial activity on the fabric surface. The findings revealed that insect- and fungus-derived chitosans are suitable as functional textile finishes and can be used as alternatives to crustacean-derived chitosan. In this respect, the study contributes to the evaluation of new raw material sources. Since insects and fungi are cultivable, they constitute a sustainable source of chitosan. In the future, comparative studies should be conducted using chitosans with different degrees of deacetylation and different linker systems. Their effects on fabric adhesion, wash resistance, and antibacterial activity should be systematically investigated. This will contribute to the development of sustainable and long-lasting functional textile coatings.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by the Scientific Research Projects Unit of Trakya University (Project No: TUBAP 2018/198), Türkiye.
Data availability statement
The data supporting the findings of this study are available within the article.