
Abstract
Moffitt’s developmental taxonomy has sparked much attention among criminologists interested in explaining the etiology of life-course-persistent (LCP) offending. The taxonomy suggests that genetic factors influence LCP offending, that genetic risk factors will be mediated by neuropsychological deficits, and that genetic factors interact with environmental factors to influence LCP offending. Various behavior genetic methodologies were used to estimate the genetic influence on LCP offending, to determine whether these genetic factors were mediated by the presence of neuropsychological deficits, and to control for genetic factors while simultaneously estimating the impact of numerous environmental influences. The findings suggested that genetic factors influence persistent offending and that these influences are partially mediated by levels of self-control. No parental influences predicted persistent offending after controlling for genetic effects, no Gene × Environment interactions were found, and few environmental influences operated as a nonshared environmental predictor of persistent offending.
Criminologists have long noted that the best predictor of future criminality is past criminality (Nagin & Paternoster, 2000; Robins, 1978). This finding has sparked myriad lines of research into the developmental origins of offending behaviors, the life course trajectories of offending behaviors, and the correlates of long-term offending (DeLisi & Piquero, 2011). One of the most consistent findings to emerge from this research is that a small portion of the population is far more likely to be involved in criminal behavior beginning in adolescence and continuing into adulthood (Moffitt, 2006). This group is commonly referred to as life-course-persistent offenders (LCP; Moffitt, 1993).
LCP offenders (or LCPs) are grossly overrepresented in crime statistics (DeLisi, 2005). Scholars argue that roughly 10% of the general population are LCP offenders (Moffitt, 1993), but their criminal behavior is believed to account for more than 50% of all crimes (Piquero, 2011). Not only are LCP offenders more involved in crime than other members of the population, but they also tend to commit interpersonal violent crimes at a higher rate (Moffitt, 2006; Moffitt, Caspi, Harrington, & Milne, 2002). When one ties all of this together, it is easy to see why criminologists have devoted much attention to these offenders (DeLisi & Piquero, 2011).
Though researchers have explored many aspects of LCP offending, one of the more pressing questions that has yet to be fully addressed is, “What causes persistent offending?” A number of theories have been proffered, and the potential explanations include familial or parental influences (Gottfredson & Hirschi, 1990), progression and state dependence effects (Loeber, 1996; Sampson & Laub, 1993), and the presence of neuropsychological deficits (Moffitt, 1990). Although each of the extant theories offers a unique perspective on the etiology of persistent offending, one of the most prominent explanations comes from Moffitt’s (1993) developmental taxonomy.
The following sections will provide an overview of Moffitt’s (1993) theory, with particular emphasis on her hypotheses concerning the etiology of LCP offending. Next, an analysis will be presented using a probability sample from the United States. The analysis will test different hypotheses laid out in the taxonomy. Finally, the results of the analysis are discussed in terms of their importance for theoretical advancement.
Moffitt’s Developmental Taxonomy
In 1993, Moffitt set forth a theory of criminality that has drawn attention from scholars around the world (Bartusch, Lynam, Moffitt, & Silva, 1997; DeLisi & Piquero, 2011). At the most basic level, Moffitt hypothesized that aggregate crime statistics, such as the age-crime curve, masked important variation in offending patterns. Her main thesis was that two types of offenders are identifiable in the population: adolescence-limited (AL) offenders and LCP offenders. The AL category encompasses the vast majority of offenders. Indeed, Moffitt argued that roughly 90% of the offending population are AL offenders (or ALs). Although this group is large in number, the group’s offending is age patterned and consists of minor deviant and delinquent acts. Perhaps most importantly, AL offenders cease their involvement in criminal activity promptly after entering early adulthood (around age 20).
Research generally supports the existence of an AL offender group (Moffitt, 2006), but some studies have painted a more complicated picture suggesting that other groups exist (Eggleston & Laub, 2002; Laub & Sampson, 2003; Skardhamar, 2009). Regardless, the criminological literature has produced mounds of studies finding that for most individuals, youthful delinquency peaks in adolescence and returns to preadolescent levels quickly during early adulthood (Wright, Tibbetts, & Daigle, 2008). Also, AL offenders are less likely to be involved in violent and aggressive crime (Piquero & Brezina, 2001) and appear to be influenced by a combination of social and biological influences referred to as the maturity gap (Barnes & Beaver, 2010).
Although AL offenders represent an important and large piece of the criminal careers puzzle (DeLisi & Piquero, 2011), the vast majority of research into Moffitt’s (1993) theory has centered on the LCP offender group (Moffitt, 2006). LCP offending is said to originate early in the life course, within the first few years of life (Farrington, 1998). During early childhood, LCP offenders will develop difficult temperaments (Moffitt, 1990), will display aggressive and violent behavior (Tremblay et al., 1999), and will display a limited ability to regulate their attention and behavior (Gottfredson & Hirschi, 1990; Moffitt & Caspi, 2001). In early adolescence, these individuals will begin their foray into deviant and criminal activity. LCPs will initiate themselves into drug use, sexual activity, and interpersonal crime at a much earlier age than their AL counterparts (Moffitt et al., 2002; Odgers et al., 2007). During late adolescence and early adulthood, around the time AL offenders are desisting from crime, LCPs will maintain their frequent involvement in criminal activity and may progress to more serious criminal activity (Loeber, 1996). Although conflicting interpretations of the theory abound (Laub & Sampson, 2003), Moffitt (1993) argued that LCP offenders are unlikely to completely cease their involvement in antisocial behavior, although their crime involvement may wane with old age (Moffitt, 2006). LCP offenders have drawn much attention from criminologists and with good reason, given their heightened frequency of offending and their overinvolvement in serious and violent activity (Raine, Brennan, & Mednick, 1994).
Etiology of LCP Offending
The etiology of LCP offending has roots in two factors: genetic-biological influences and early-rearing environment influences (Moffitt, 1993). Stated succinctly, LCP offenders are born with neuropsychological deficits that increase the risk that antisocial behavior will develop (see, generally, Hirschi & Hindelang, 1977; Ogilvie, Stewart, Chan, & Shum, 2011; Utendale, Hubert, Saint-Pierre, & Hastings, 2011). At the same time, LCPs are born into adverse rearing environments that are unable to respond to their behavior in a prosocial manner (Moffitt & Caspi, 2001). The confluence of these two factors (i.e., neuropsychological deficits and adverse home environment) works to increase the chances that an LCP offending pattern will develop beginning in early childhood.
A large literature has examined Moffitt’s (1993) hypotheses concerning the origins of LCP offending. In broad strokes, research has revealed that LCP offenders are more likely to suffer from neuropsychological deficits (Piquero, 2001; Raine et al., 2005), to be born to at-risk parents (Tibbetts & Piquero, 1999), and to suffer from a combination of neurocognitive impairments and environmental risk factors (Raine et al., 1994; Turner, Hartman, & Bishop, 2007). Raine et al. (1994) reported that fewer than 5% of their sample suffered from both birth complications and early maternal rejection. Nonetheless, these individuals accounted for roughly 20% of all violent offenses. Tibbetts and Piquero (1999) reported that low birth weight (a proxy for neuropsychological deficits) interacted with familial socioeconomic status (SES) and with a measure of family structure in predicting an early onset of offending. In line with the theory, both interactions revealed that the impact of neuropsychological deficits (i.e., low birth weight) on offending were greater in the presence of an adverse rearing environment (i.e., low SES or weak family structure).
In a similar analysis, Gibson, Piquero, and Tibbetts (2000) assessed whether children born to mothers who smoked while pregnant (a proxy for neuropsychological deficits; but see McGloin, Pratt, & Piquero, 2006) were more likely to display an early onset of delinquency. Their regression models revealed that maternal smoking predicted early onset, as did low birth weight and SES. Raine et al. (2005) analyzed a group of adolescents who were on the LCP pathway for a range of neuropsychological deficits. These authors reported that LCP offenders had lower IQ scores than ALs and controls, they had greater memory impairments as compared to control participants, they had neurocognitive impairments, and they were more likely to have suffered head injuries that resulted in unconsciousness. In all, these studies have revealed strong support for the neuropsychological element of LCP offending. Also, many have supported the interactional hypothesis between neuropsychological deficits and an adverse rearing environment (e.g., Raine et al., 1994).
Genetic Influences on LCP Offending
Although an impressive literature has analyzed the impact of neuropsychological deficits on LCP offending (DeLisi & Piquero, 2011; Gibson et al., 2000; Raine et al., 2005), few have traced these effects to genetic factors. This oversight is surprising given that neuropsychological deficits are likely the result of environmental and genetic influences (Beaver, Wright, & DeLisi, 2007; Raine, 2008). Brain structure and function appear to be regulated largely via genetic factors (Devlin, Daniels, & Roeder, 1997; Thompson et al., 2001; Toga & Thompson, 2005), although some environmental influences are believed to impinge directly on neurocognitive functioning (Wright, Boisvert, & Vaske, 2009; Wright, Dietrich, et al., 2008). Recall that one of the hallmarks of LCP offending is an early onset of antisocial behavior. Researchers have investigated the genetic influences on early childhood antisocial behavior, and the results have been surprisingly consistent; childhood antisocial behavior is, at least partially, the result of heritable factors (Arseneault et al., 2003; Jaffee et al., 2005; van Beijsterveldt, Bartels, Hudziak, & Boomsma, 2003; Van Hulle et al., 2009). Taylor, Iacono, and McGue (2000), for example, reported that genetic factors were more prominent for youth who displayed an early onset into delinquency as compared to those who had a late onset. These findings hint that LCP offending may be partially driven by genetic influences.
Perhaps the clearest indication that genetic factors underlie LCP offending comes from a recent analysis by Barnes, Beaver, and Boutwell (2011). Drawing on data from the National Longitudinal Study of Adolescent Health (Add Health; K. Harris, 2009), Barnes et al. first separated the sibling subsample into three groups: LCPs, ALs, and abstainers. Next, the authors estimated the genetic influences on each offending pattern. The findings from this portion of the analysis revealed that the LCP pattern was between 56% and 70% heritable, with the remaining variance being attributable to nonshared environmental factors. In other words, genetic influences accounted for more than half of the variance in being identified as an LCP offender. Nonshared environmental influences—environmental experiences that are unique to each individual—accounted for the remaining variance.
The Current Study
Juxtaposing the findings from behavioral genetic research (e.g., Taylor et al., 2000) with Moffitt’s (1993) theory raises several interesting and important questions. First, if genetic factors underlie LCP offending, how do these effects operate? As noted above, any genetic influence on LCP offending is almost certainly mediated by the brain and is likely tied to Moffitt’s arguments regarding neuropsychological dysfunction. Put another way, genetic factors identified by previous studies may indirectly affect LCP offending via their impact on neuropsychological deficits (Wright, Tibbetts, et al., 2008). See Figure 1 for a graphical depiction of the hypothesized relationship. Notice that the figure includes three latent terms: genetic factors, neuropsychological deficits, and LCP offending. Genetic factors are hypothesized to affect neuropsychological deficits, which in turn influence LCP offending. When this relationship is taken into account, the link between genetic factors and LCP offending is substantially weakened or is reduced to zero. Thus, the arrow connecting genetic factors directly to LCP offending is displayed as a broken line. The current study will analyze these relationships.
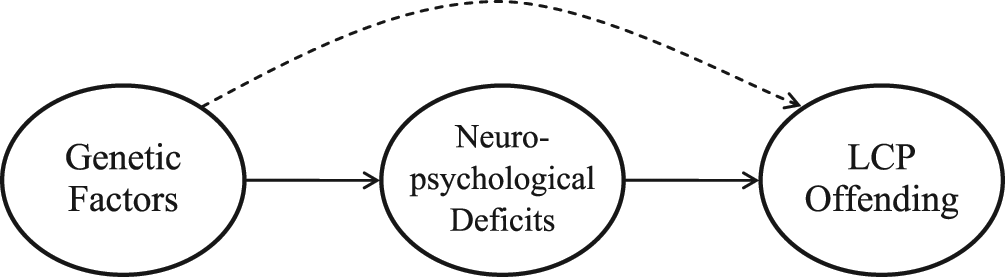
Graphical Depiction of the Hypothesized Relationship Between Genetic Factors, Neuro- psychological Deficits, and Life Course–Persistent Offending
The second question that emerges from contemporary research concerns the nonshared environmental factors that influence LCP offending. The nonshared environment has been shown to affect LCP-like offending (Barnes et al., 2011; Taylor et al., 2000) and is believed to account for the majority of environmental influences on antisocial behavior generally (Moffitt, 2005). Although scholars agree that the nonshared environment is of utmost importance (J. Harris, 1998), it is not yet clear what these environments are (Plomin, Asbury, & Dunn, 2001). Nonshared environments are defined as any factor (other than genetics) that is different between two siblings and operates to make them less similar to one another (Plomin & Daniels, 1987). Although defining nonshared environments was simple, the empirical search for them has been substantially more difficult (Plomin et al., 2001; Turkheimer & Waldron, 2000).
Moffitt (1993) hypothesized that LCP offending would arise as the result of neuropsychological deficits and an adverse rearing environment. Adverse rearing environments may tap anything from poor parenting to being raised in a disadvantaged neighborhood (e.g., Tibbetts & Piquero, 1999). The current study will examine whether a host of parental influences operates as nonshared environmental influences on LCP offending. Although these constructs may be viewed as shared environmental factors (i.e., they, theoretically at least, should lead siblings to be more similar to one another), scholars have argued for the investigation of whether ostensibly shared environmental factors actually operate as nonshared influences. This may occur for several reasons, not least of which is that two people who experience the same event (e.g., parental discipline) may have nonshared interpretations of that event, leading to a nonshared environmental effect (Turkheimer & Waldron, 2000).
Parental influences are one of the cornerstones of criminological research (e.g., Hirschi, 1969). The effect that parenting has on LCP offending, however, has yet to be analyzed after controlling for genetic influences. This is an important oversight because as prior scholars have noted (Beaver, 2008; DiLalla, 2002; Jaffee & Price, 2007; Kendler & Baker, 2007; Plomin & Bergeman, 1991; Walsh, 2002; Wright & Beaver, 2005), environmental influences cannot be analyzed properly unless genetic factors are accounted for first. The current study will estimate the effect of various parental factors (all of which have been linked with offending behaviors by prior research) on the probability of being an LCP offender. These effects will be observed after controlling for genetic factors that influence LCP offending.
In summary, four hypotheses drawn either directly from Moffitt’s (1993) theory or by combining current research findings with Moffitt’s theory are analyzed in the current study.
Hypothesis 1: The effect of genetic risk factors on LCP offending will be mediated by the presence of neuropsychological deficits.
Hypothesis 2: Parental influences, even after accounting for genetic risk, will predict LCP offending.
Hypothesis 3: Genetic risk factors for LCP offending will interact with parental and neighborhood influences such that the presence of poor parenting or the presence of a disadvantaged neighborhood will exacerbate genetic risk.
Hypothesis 4: Measures of neuropsychological deficits and parental influences will operate as nonshared environmental factors on LCP offending risk.
To date, no research has directly examined any of the above hypotheses.
Method
Data
Data for the current study come from Add Health (K. Harris, 2009). As a longitudinal and nationally representative sample of adolescents, the Add Health is an ideal data set for the current focus. The Add Health study unfolded in a number of steps, beginning with a survey of more than 90,000 students who were attending 132 different schools in 1995. This round of data collection is referred to as the in-school survey and provided the sampling frame from which the longitudinal portion of the study was drawn.
Immediately after the in-school surveys were completed, a subsample of the students who completed the questionnaire were contacted and asked to complete a follow-up interview, along with their primary caregiver, in their homes. Information from 20,745 adolescents and 17,700 primary caregivers was gathered during this round of data collection. This round of surveys is referred to as Wave 1. The Wave 1 surveys were designed to gain more detailed information about the adolescent, his or her social experiences, and his or her rearing environment. Approximately 1 year after the Wave 1 interviews were completed, 14,738 of the respondents were again interviewed in their homes. This round of surveys is known as Wave 2. Only a short amount of time elapsed between Wave 1 and Wave 2, and as a result, the questionnaires remained very similar. For instance, respondents were asked about their behaviors and their ability to get along with others.
Nearly 6 years after Wave 1 interviews were conducted (and roughly 5 years after Wave 2), a third round of interviews took place with 15,197 respondents (i.e., Wave 3 in-home interviews). By this time, the respondents had reached early adulthood. To account for these age differences, the survey was redesigned to include age-appropriate questions. For example, respondents were asked about their employment histories, their marital relationships, and their involvement in criminal behavior. Finally, a fourth round of interviews was completed between 2007 and 2008. Roughly 12 years had passed since Wave 1 interviews were conducted, and all of the respondents had reached adulthood. The age range at Wave 4 was 24 to 34 years. Similar to Wave 3, participants were asked questions that tapped their employment histories, romantic relationships, and involvement in criminal behavior. Wave 4 surveys were completed by 15,701 participants.
Nested within the Add Health data is a subsample of sibling pairs who resided in the same household at Wave 1. This subsample of sibling pairs is used in the current analysis. During Wave 1 in-home interviews, all respondents who lived with an identical twin (monozygotic [MZ]), a fraternal twin (dizygotic [DZ]), a half sibling, or a stepsibling were identified, and their sibling was automatically included in the study. Additionally, full siblings were included in the sample, but these pairs entered the subsample as a result of chance. The current study was limited to MZ twins, DZ twins, and full siblings. All other sibling pairs were removed from the sample to limit the possibility that assortative mating effects would artificially bias heritability estimates.
Measures
Persistent offending
LCP offenders were identified by following two steps. First, a scale of each respondent’s involvement in crime and delinquency at Wave 1, Wave 2, Wave 3, and Wave 4 was generated. Participants were asked about their involvement in 17 different delinquent activities at Wave 1 and Wave 2 (questions were identical at both waves). During both waves, the reference period for each of the delinquency questions was “during the past 12 months.” The Wave 1 scale was created by summing each respondent’s answers to the 17 questions so that higher values reflected a greater involvement in delinquency (α = .85). These same questions were asked at Wave 2, allowing for the calculation of a Wave 2 delinquency scale by summing across the 17 items (α = .81). During Wave 3 interviews, respondents were asked about the frequency with which they had engaged in 12 criminal behaviors in the past 12 months. As with the previous scales, each respondent’s answers to the 12 questions were summed together to create the Wave 3 criminal behavior scale so that higher values indicated a greater involvement in crime (α = .71). Twelve questions were asked about the respondent’s involvement in criminal activities during Wave 4 interviews. When summed together, higher scores reflected more involvement in criminal activity at Wave 4 (α = .71).
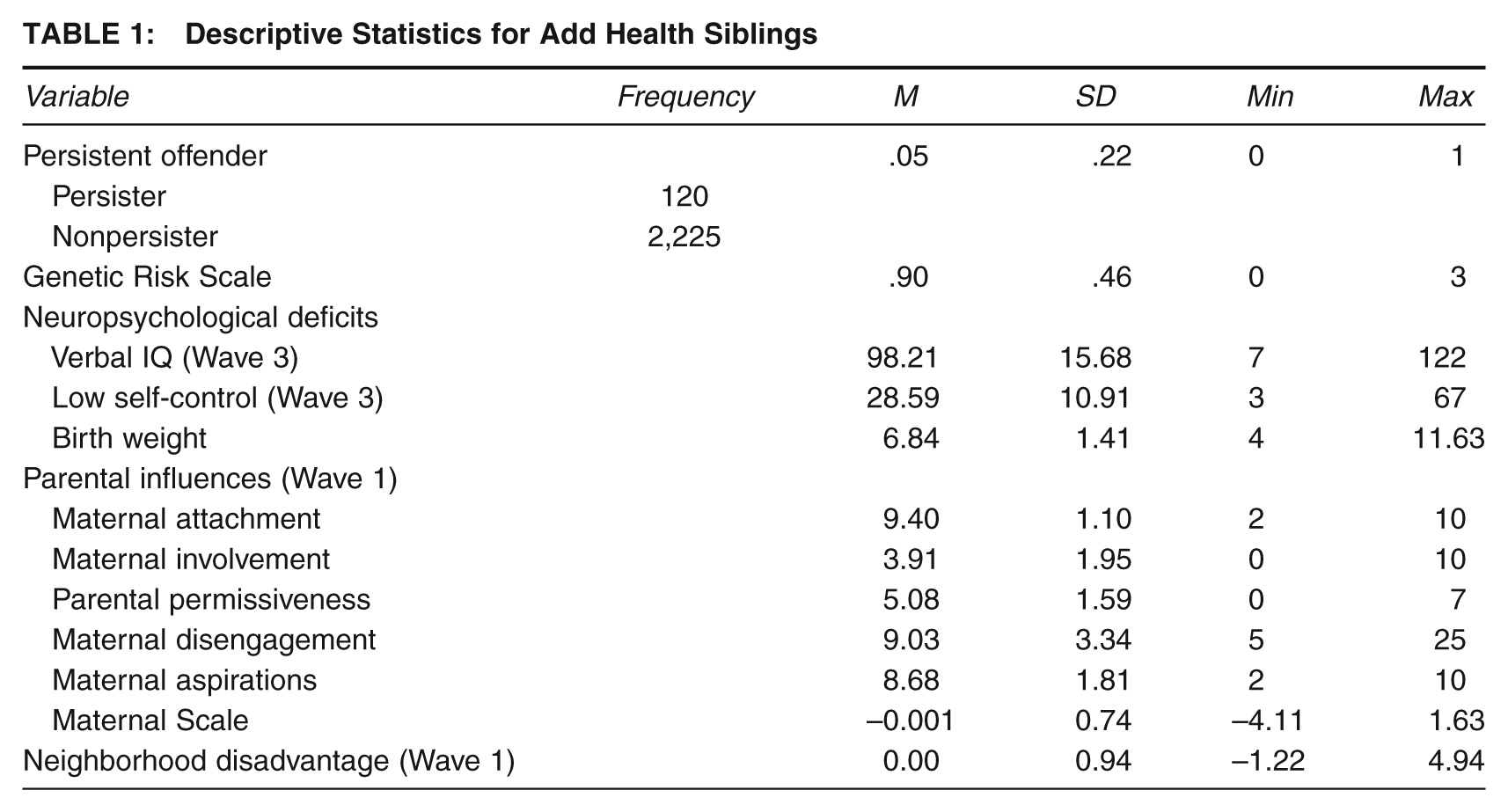
The second step toward identifying LCP offenders was carried out by generating a new variable whereby a 1 was assigned to all respondents who scored 1 or higher on each of the four crime and delinquency scales (Barnes et al., 2011; and see Turner et al., 2007, for a similar coding strategy). Respondents who did not score 1 or higher across all four scales were assigned a value of 0. As shown in Table 1, this approach led to roughly 5% of the sample being identified as an LCP offender.
Descriptive Statistics for Add Health Siblings
Genetic Risk Scale
Behavioral genetic scholars have developed myriad ways to measure and control for genetic risk factors. The current study used an established strategy for capturing genetic influences on a dichotomous trait (Beaver, Barnes, May, & Schwartz, 2011; Kendler et al., 1995). Building on the knowledge that MZ twins share 100% of their DNA and DZ twins and full siblings share 50% of their distinguishing DNA (on average; Carey, 2003), a genetic risk continuum can be constructed by combining genetic relatedness information with information about the cosibling’s status as a persistent offender.
Generating the scale involved five steps. First, the data were arranged so that each sibling appeared once as the target sibling and once as the cosibling. Second, because MZ twins share 100% of their DNA, any target MZ twin whose cotwin was identified as a nonpersistent offender would have the lowest genetic risk for becoming a persistent offender. Thus, all target MZ twins whose cotwin was not a persistent offender were coded as 0. Third, target DZ twins and full siblings whose cotwin or sibling was not a persistent offender have the next lowest genetic risk of becoming a persistent offender themselves. These participants, therefore, were coded as 1. Fourth, target DZ twins and full siblings whose cotwin or sibling was identified as a persistent offender have a higher genetic probability of being a persistent offender. To account for these influences, all target DZ twins and full siblings who had a cotwin or sibling who was identified as a persistent offender were coded as 2. Finally, target MZ twins whose cotwin was identified as a persistent offender have the highest genetic liability toward persistent offending. Thus, these respondents were coded as 3.
The Genetic Risk Scale has been used previously to control for genetic influences on a range of outcomes, such as childhood externalizing problems (Boutwell, Franklin, Barnes, & Beaver, 2011), depression (Kendler et al., 1995), and psychopathy (Beaver et al., 2011). Important for the current focus, the Genetic Risk Scale will allow for the examination of the link between genetic risk factors for LCP offending and neuropsychological deficits (see Figure 1).
Verbal IQ
Moffitt (1993, p. 681) noted that a low verbal IQ may reflect neuropsychological dysfunction, and prior scholars have linked verbal IQ with problems with self-control (Ratchford & Beaver, 2009), psychopathy (Johansson & Kerr, 2005), and persistent delinquent involvement (Moffitt, Lynam, & Silva, 1994). In other words, scholars have used verbal IQ as a proxy for neuropsychological dysfunction in prior research (Piquero, 2001). The current study included a measure of verbal IQ gleaned from the Wave 3 interviews. During these interviews, respondents were administered the Peabody Picture Vocabulary Test. Standardized scores were used so that the distribution for the full Add Health sample was normal with a mean of 100 and a standard deviation of 15 (the mean and standard deviation are slightly different for the sibling subsample). Higher scores reflected a greater verbal IQ.
Low self-control
Moffitt (1993, p. 681) noted that individuals with neuropsychological deficits may display signs of low self-control. In other words, measures of self-control may be used as an indicator of neuropsychological dysfunction. Building on this hypothesis, a measure of self-control was constructed using the Wave 3 survey data. Specifically, Beaver, Ratchford, and Ferguson (2009) identified 20 items that, when combined, tap each respondent’s level of self-control. Factor analysis indicated that all 20 items hung together on a single construct, and the alpha coefficient indicated a strong degree of reliability (α = .83). To generate the scale, responses to the 20 items were summed so that higher values reflected lower levels of self-control.
Birth weight
Previous research has used birth weight as an indicator of neuropsychological deficits (Tibbetts & Piquero, 1999). Following the lead of these scholars, a measure of the respondent’s birth weight was included in the current analysis. During Wave 1 interviews, primary caregivers were asked to report on their child’s (i.e., the target respondent’s) birth weight. Responses were recorded in pounds and ounces. As a result, the birth weight measure was created whereby whole numbers reflected pounds and ounces were coded as a fraction of a pound. The substantive conclusions were unchanged when the birth weight measure was dichotomized into an indicator of low birth weight (i.e., those born at or below 5.5 pounds).
Parental influences
Moffitt (1993) noted that “parents of children who are difficult to manage often lack the necessary psychological and physical resources to cope constructively with a difficult child” (p. 681). Clearly, Moffitt anticipated a parental influence on the child’s probability of becoming an LCP offender. For capturing these influences, a series of parenting scales was constructed and included in the analysis. First, a measure of maternal-child attachment was constructed. During Wave 1 interviews, target respondents were asked about the level of closeness they felt toward their mothers and how much they thought their mothers cared about them. Both items were coded so that higher values indicated a greater bond between the respondent and his or her mother. Responses to these two questions were summed together to create the scale (α = .63).
The second parenting scale indexed the level of involvement that mothers had with their children. At Wave 1, target respondents were asked whether they had taken part in 10 activities with their mother in the past 4 weeks. Activities referenced were shopping, going to a religious service, and going to a movie, among others. Each measure was coded dichotomously so that 0 = the respondent did not participate in the activity with his or her mother and 1 = the respondent did participate in the activity with his or her mother. To generate the scale, responses to the 10 items were summed so that higher values reflected greater maternal involvement (α = .53).
The third parenting scale measured the amount of autonomy granted to the child by his or her parents at Wave 1. Target respondents were asked to reflect on whether their parent(s) allowed them to make decisions regarding their curfew, their peer group, and the clothes they wore. Each of the seven questions were coded dichotomously (0 = no, 1 = yes) so that when summed, higher values reflected greater parental permissiveness (α = .63).
A fourth measure of parenting practices tapped the level of disengagement between the mother and the target respondent. Five questions measured the level of warmth expressed by the respondent’s mother, the level of encouragement offered by the mother, and the level of satisfaction with the mother-child relationship. Each question was asked to the target respondent at Wave 1. Answers to the five items were summed, and higher values indicated more maternal disengagement (α = .82).
The fifth parental measure tapped the mother’s aspirations for the target respondent’s future. Importantly, the questions were asked to the target respondent during Wave 1 so they reflected the participant’s perceptions of his or her mother’s aspirations. The first question asked how disappointed the respondent’s mother would be if he or she did not graduate from college (ranged from 1 = low disappointment to 5 = high disappointment). The second question was similar, but the reference was high school. Responses to these two items were summed so that higher values reflected higher perceptions of maternal aspirations (α = .57).
The final parental measure was a composite scale constructed from the items mentioned above. Specifically, each of the five scales outlined above was subjected to a factor analysis. The results from this analysis indicated that three of the scales (Maternal Attachment, Maternal Involvement, and Maternal Disengagement [reverse coded]) could be combined together into a single index of maternal influence. Because each scale was measured on a different metric, the scales were first standardized and then an average score was generated. The resulting Maternal Scale was coded so that higher values reflected “better” or more positive relations between mother and child (α = .59).
Neighborhood disadvantage
A Neighborhood Disadvantage Scale was created by combining information from the following five variables taken from the 1990 Census: unemployment rate, proportion of residents on public assistance, proportion of residents living below the poverty line, proportion of residents who are Black, and proportion of female-headed households (Sampson, Raudenbush, & Earls, 1997). Each of the constituent variables, which were measured at the block-group level, was factor analyzed, and the results suggested that the variables be combined into a single construct. Thus, one factor was extracted using regression scoring techniques (α = .79). Higher values reflected a greater degree of neighborhood disadvantage.
Analysis
The analysis unfolded in three broad steps. The first step was to analyze the relationship between genetic risk and the persister status variable. Two analyses were conducted for this step. First, the behavior genetic technique known as the ACE model was estimated. The ACE model has been described at length elsewhere (Neale & Maes, 2004; see Wright, Beaver, DeLisi & Vaughn, 2008, for a recent example of the ACE model in criminology). Briefly, the ACE model will decompose the variance in the persister status variable to determine whether genetic factors (i.e., A) are operative. The ACE model will also estimate the degree to which shared environments (i.e., C; environments that operate to make siblings more similar to one another) and nonshared environments (i.e., E; environments that operate to make siblings dissimilar) are operative. The threshold ACE model was estimated to account for the dichotomous coding of the outcome variable (Derks, Dolan, & Boomsma, 2004). Whether the ACE model revealed genetic influences on the persister status variable was critical to the remainder of the analysis—indeed, the saliency of the Genetic Risk Scale rests on the assumption that genetic factors are operative on the outcome of interest (i.e., persister status). The findings from the ACE model, therefore, paved the way for the remaining analyses, which used the Genetic Risk Scale.
The next set of analyses employed the persister status indicator as the dependent variable, and the Genetic Risk Scale was used as the primary predictor variable. The bivariate relationship between these two variables was examined first. This model provided a baseline relationship that could then be compared to subsequent models (i.e., when the other covariates were entered). Recall that prior work has identified a strong genetic link to LCP offending (Barnes et al., 2011) and that this relationship is most likely mediated via neuropsychological deficits (Moffitt, 1993). To test this hypothesis (Hypothesis 1), the three neuropsychological deficit measures (i.e., verbal IQ, low self-control, and birth weight) were entered into the model in a stepwise fashion. Entering the variables into the model in this way allowed for the examination of mediating effects. Specifically, the degree to which the Genetic Risk Scale is mediated by the neuropsychological deficits measures will be identified by a reduction in the genetic risk coefficient from Model 1 through the subsequent models.
The second step to the analysis observed whether the parental influence scales and the Neighborhood Disadvantage Scale increased the odds of being identified as an LCP offender after controlling for genetic risk (Hypothesis 2). These models explored the main effects of each measure to determine the relative magnitude of the effects among the various items. Additionally, interaction terms were analyzed to test Moffitt’s (1993) interactional hypotheses (Hypothesis 3). The interaction terms were calculated by mean centering the constituent variables and then generating a new variable that was the product of the two items.
The third step to the analysis examined the link between each of the covariates (except for the neighborhood disadvantage measure because it did not vary between siblings; see, generally, Caspi, Moffitt, & Plomin, 2000) and LCP offending, but this time the covariates were included as measures of the nonshared environment (Hypothesis 4). Prior research has identified a substantial nonshared environmental component to LCP offending, but it is unclear what these nonshared environments might be. For capturing nonshared environmental effects, each of the neuropsychological deficits measures and the parental influence measures was entered as between-sibling difference scores (Beaver, 2008), and a logistic DeFries-Fulker (DF) equation was estimated (Rodgers, Rowe, & Li, 1994). The logistic DF equation can be expressed algebraically as follows:
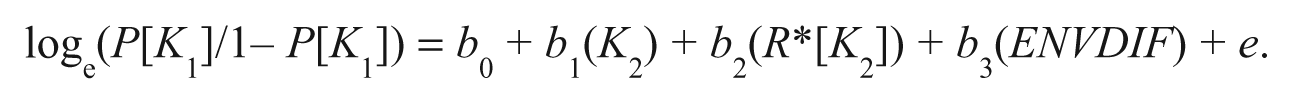
The logistic DF equation has two features that deserve mentioning. First, genetic influences on the outcome variable (i.e., persistent offender) are captured by the heritability coefficient (i.e., b2). Second, the nonshared environmental influences of the covariates will be captured by the ENVDIF coefficient estimate (i.e., b3).
Results
The first step to the analysis was to estimate the ACE model. Recall that the ACE model decomposes variance in an outcome measure into genetic (A), shared environmental (C), and nonshared environmental (E) influences. Behavior geneticists typically estimate the full ACE model and then estimate several supplemental models to determine whether equivalent model fit can be reached with a more parsimonious model; the procedure is often referred to as hierarchical model fitting in the structural equation modeling literature. The full ACE model was estimated first, and results (not reported in a table) suggested that a combination of genetic (A = .51) and nonshared environmental factors (E = .49) explained variance in the persistent offending variable. Next, an AE model was estimated. In this model, the C parameter was fixed to zero. The AE model produced the best fit to the data and revealed that genetic (A = .51) and nonshared environmental factors (E = .49) explained the totality of the variance in the persistent offending variable. As compared to the full ACE model, the AE model produced an equivalent chi-square value and a reduced Akaike information criterion (AIC) value (because of the fixed C parameter) (∆χ2 = .00, p > .05, ∆AIC = −2.00). The CE model produced a nonsignificant chi-square value, but the AIC was larger than the AIC gleaned from the AE model (∆χ2 = 1.87, p > .05, ∆AIC = −.14). Finally, the E model was estimated. This model also produced a nonsignificant chi-square statistic, but when compared to the ACE model, the AIC indicated a worsening of fit (∆χ2 = 4.50, p > .05, ∆AIC = .50). In all, the findings from the ACE model-fitting analysis suggested that genetic factors influenced the persister status variable. This analysis, therefore, supported the utility of the Genetic Risk Scale by revealing that genetic factors explain a portion of the variance in the persistent offending variable.
Presented in Table 2 are the findings from seven separate logistic regression models. 1 The first four models analyzed the link between the Genetic Risk Scale and the various measures of neuropsychological deficits. Model 1 revealed that the Genetic Risk Scale was positively and significantly associated with LCP offending. The odds ratio (OR) is reported and indicates that each one-unit increase on the Genetic Risk Scale was associated with a 148% increase in the odds of being identified as an LCP offender. This finding offers support for Hypothesis 1 and is an important first step in the examination of the remaining three hypotheses.
Logistic Regression of Persistent Offending on Genetic Risk Scale, Neuropsychological Deficits, Parental Influences, and Neighborhood Disadvantage
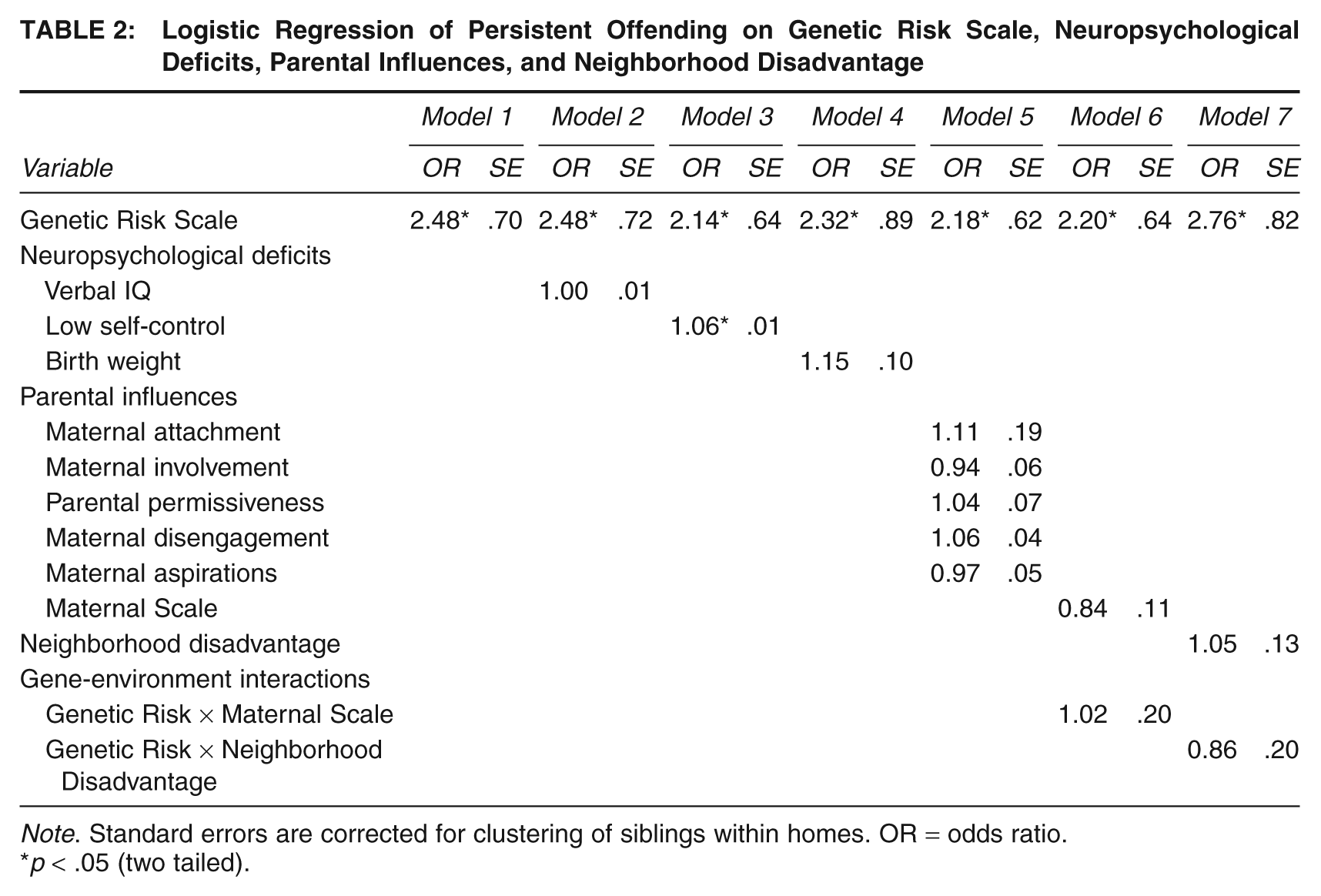
Note. Standard errors are corrected for clustering of siblings within homes. OR = odds ratio.
p < .05 (two tailed).
To explore the link between the Genetic Risk Scale and LCP offending more closely, predicted probabilities were generated using the model estimates from Model 1 in Table 2. These predicted probabilities are presented graphically in Figure 2, which shows that the probability of a respondent’s being identified as an LCP offender was strongly tied to his or her genetic risk score. Respondents with the lowest genetic risk (i.e., MZ twins whose cotwin was not identified as LCP) had a predicted probability of .02 for being an LCP offender. Note that this predicted probability is lower than the population incidence of LCP offending within this sample (see Table 1). Participants with the highest genetic risk (i.e., MZ twins whose cotwin was identified as LCP) had a predicted probability of .25, a probability that is more than 11 times greater than those with the lowest genetic risk.
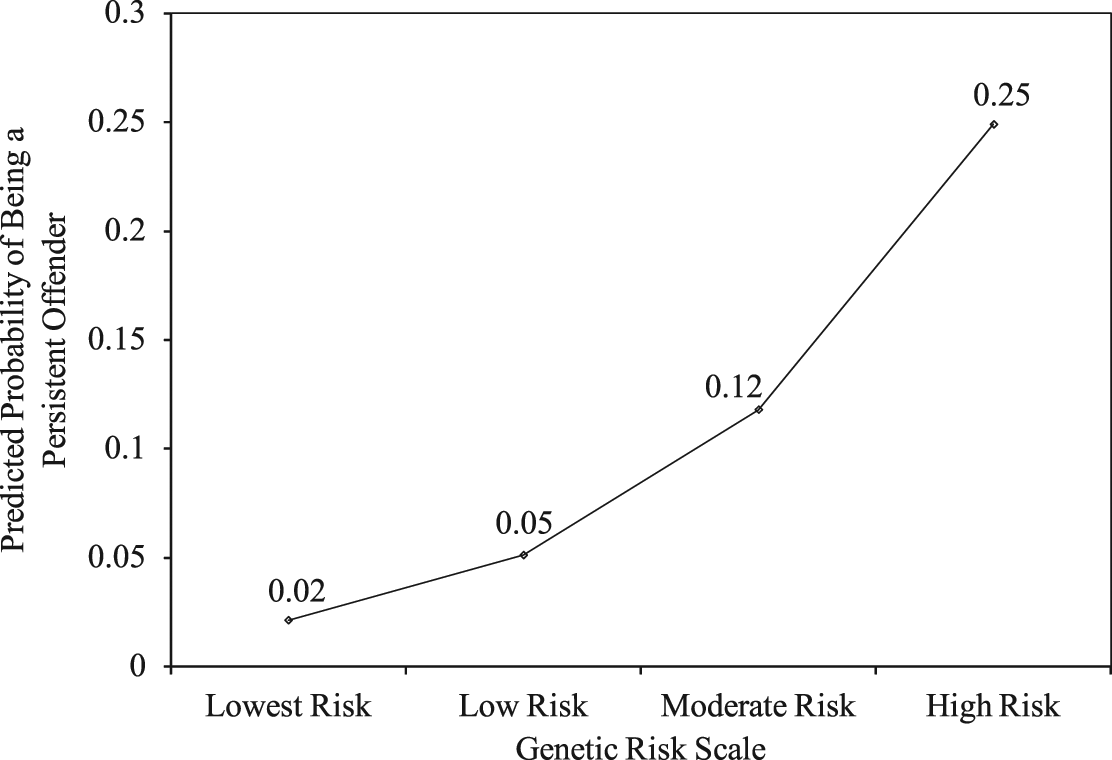
Predicted Probability of Being a Persistent Offender as a Function of Genetic Risk
Models 2 through 4 (Table 2) test for mediation of the genetic effect via neuropsychological deficits (Hypothesis 1). 2 In broad strokes, one of the three neuropsychological deficit measures substantially mediated the genetic effect. Indeed, the genetic effect was unchanged from Model 1 to Model 2 (when verbal IQ was entered into the equation) and was only marginally decreased in Model 4 when birth weight was entered. The only measure of neuropsychological deficits that appeared to mediate the genetic effect was the Low Self-Control Scale (Model 3), where roughly 13% of the genetic effect was mediated (b = .87 for Model 1; b = .76 for Model 2). A Sobel mediation test suggested that this effect was statistically significant (z = 3.59, p < .05). To explore the influence of low self-control on LCP offending, predicted probabilities were plotted and are presented in Figure 3. As shown in the figure, probabilities of LCP offending are strongly correlated with low self-control. Respondents with the highest level of self-control (coded as low numbers on the scale) had a predicted probability of .01, whereas respondents with the lowest level of self-control (coded as high numbers on the scale) had a predicted probability of .23.
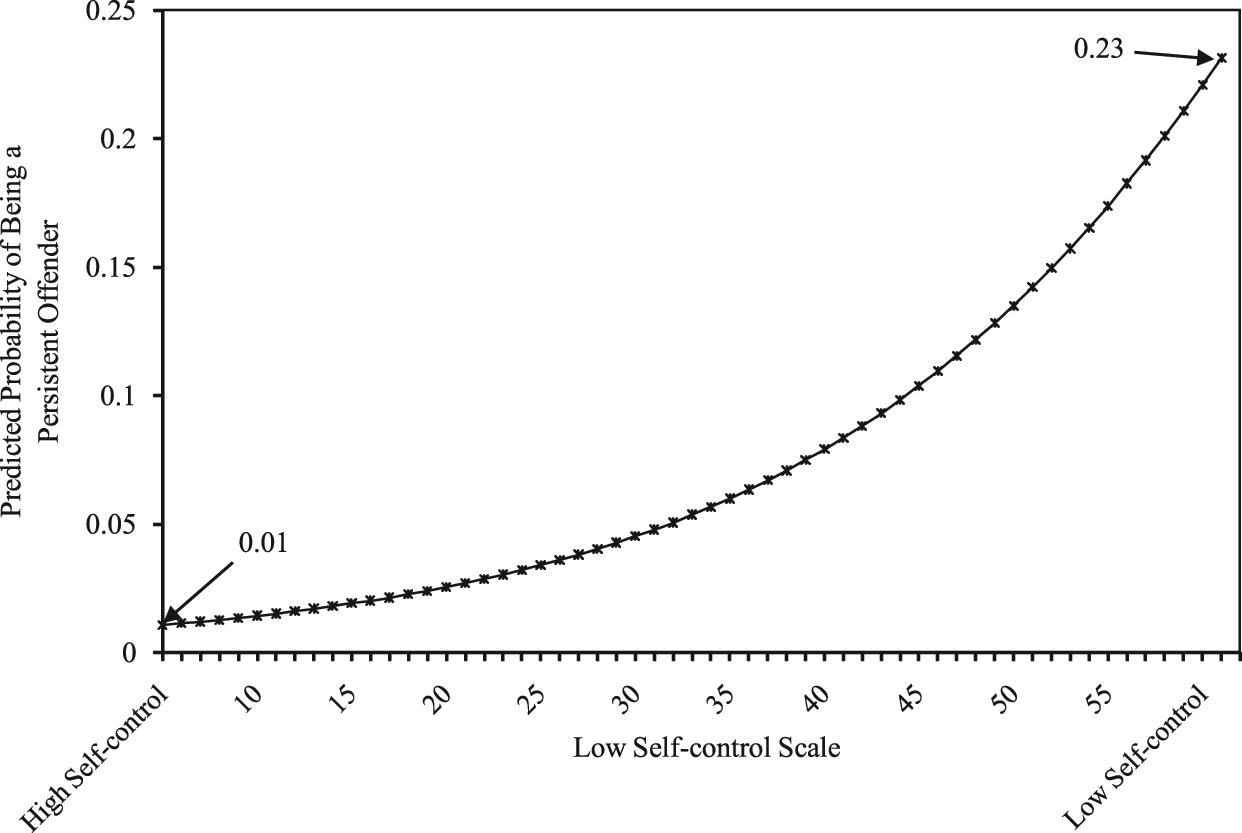
Predicted Probability of Being a Persistent Offender as a Function of Low Self-Control
Returning to Table 2, Model 5 analyzed the parental influence variables, and the results revealed that none of the parenting scales attained statistical significance (Hypothesis 2). The lack of statistical significance was not attributable to multicollinearity between the scales—none of the scales attained statistical significance when entered into the model individually. Model 6 enters the Maternal Scale as a predictor of LCP offending, along with the Genetic Risk × Maternal Scale interaction (Hypothesis 3). Neither the main effect for the Maternal Scale nor the interaction with genetic risk attained statistical significance. The lack of a main effect was not attributable to multicollinearity—the Maternal Scale did not significantly predict the persister status variable in a bivariate model (OR = .84, p > .05). The final model presented in Table 2 (Model 7) explored the link between genetic risk, neighborhood disadvantage, and an interaction between the two on the probability of persistent offending. As shown in the table, the main effect for neighborhood disadvantage did not attain significance (p = .80 when only the main effect was included), nor did the effect of the Gene × Environment interaction. 3
Thus far, the results have revealed that the genetic risk toward LCP offending is partially mediated by levels of self-control. This finding offers some support for Hypothesis 1. Hypothesis 2 was examined by entering the parental influence variables as covariates. As was shown, none of the parenting scales predicted LCP offending. These findings do not offer support for Hypothesis 2 (but see the possible limitations in the Discussion section). Finally, the results did not provide support for Hypothesis 3, which expected an interaction between genetic risk and the environmental variables (i.e., Maternal Scale and neighborhood disadvantage). Hypothesis 4 has yet to be examined. Recall that Hypothesis 4 stated that each of the covariates would operate as nonshared environmental influences on LCP offending. These analyses are considered in Table 3.
Logistic DeFries-Fulker (DF) Model Results
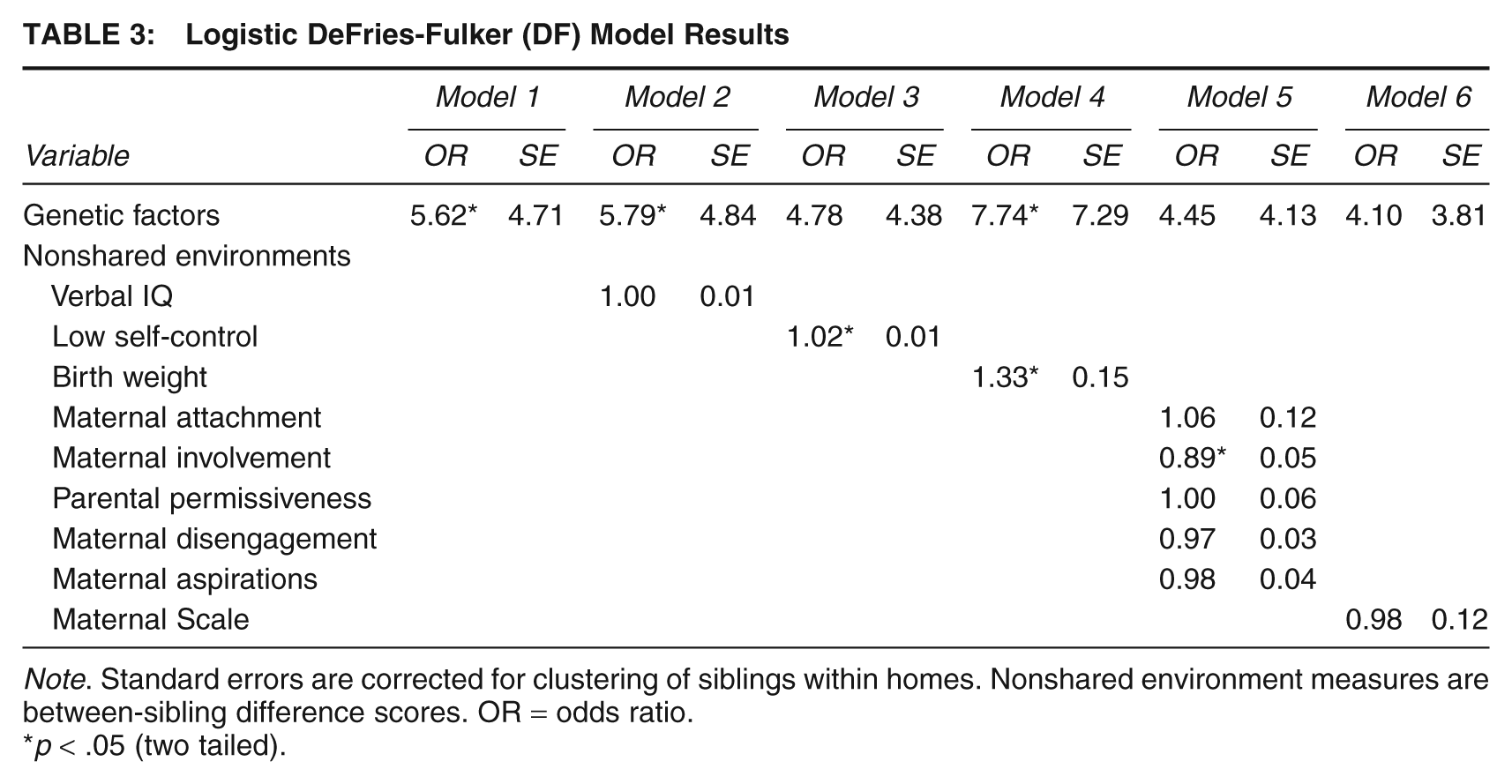
Note. Standard errors are corrected for clustering of siblings within homes. Nonshared environment measures are between-sibling difference scores. OR = odds ratio.
p < .05 (two tailed).
The logistic DF equation described earlier was estimated, and the results can be found in Table 3. 4 Before discussing the findings, it is important to reiterate that the DF model estimated environmental influences as sibling differences after controlling for genetic factors. In this way, any environmental effect that is identified in Table 3 is free of genetic confounding. As shown in Model 1 (Table 3), genetic factors accounted for a statistically significant portion of the variance in LCP offending. This finding is consistent with the regression models presented in Table 2 and the ACE model results discussed above. Model 2 entered the nonshared environmental influence of verbal IQ, and the effect did not reach statistical significance. Model 3 entered the Low Self-Control Scale as a nonshared environmental factor. The effect of low self-control was positive and statistically significant, indicating that the sibling with lower self-control was more likely to be identified as a persistent offender. Model 4 indicated that birth weight operated as a source of nonshared variance. The coefficient was positive, however (opposite to predictions), indicating that the sibling who was born of higher birth weight was more likely to be identified as a persistent offender.
Model 5 entered the parental influence variables as nonshared environments. As can be seen, only the Maternal Involvement scale emerged as a statistically significant predictor. The effect was negative, indicating that the sibling who received more maternal involvement was less likely to be identified as an LCP offender. Only the Maternal Involvement scale attained statistical significance when the parenting scales were analyzed individually. Model 6 analyzed the Maternal Scale and statistical significance was not reached.
Discussion
Moffitt’s (1993) developmental taxonomy has been extremely influential to criminology, and it has sparked a line of research that has vastly expanded knowledge about long-term criminal careers (DeLisi & Piquero, 2011). Although a number of studies have tested portions of the theory (Moffitt, 2006), no study has simultaneously examined the interconnections between genetic risk, neuropsychological deficits, adverse rearing environments, and LCP offending. The current effort sought to fill this gap in the literature by testing four hypotheses drawn or developed directly from Moffitt’s (1993) theoretical statements.
Hypothesis 1 suggested that genetic risk factors for LCP offending would operate indirectly through neuropsychological deficits. Three statistical models were analyzed and one revealed evidence to support the hypothesis. Specifically, when a measure of the respondent’s level of self-control was entered into the equation, the effect of genetic risk for LCP offending was weakened. To the extent that self-control reflects neuropsychological deficit, this finding offers support for Hypothesis 1. Neuroscience has revealed that self-control and other executive functions are housed in the prefrontal cortex of the brain (Beaver et al., 2007), suggesting that deficiencies in self-control may be indicative of neuropsychological deficits. Also, Moffitt (1993) anticipated that low self-control reflected neuropsychological deficit:
Longitudinal studies suggest that neuropsychological dysfunctions that manifest themselves as poor scores on tests of language and self-control—and as the inattentive, overactive, and impulsive symptoms of ADHD—are linked with the early childhood emergence of aggressive antisocial behavior and with its subsequent persistence. (p. 681).
In all, the finding that genetic risk factors for LCP offending are mediated by levels of self-control provides support for Hypothesis 1 and is consistent with Moffitt’s theory.
Finding a link between genetic risk for LCP offending and self-control suggests two avenues for future researchers to consider. First, scholars should prioritize research that seeks to identify the other neurological pathways that mediate genetic risk for LCP offending. Indeed, research has shown that persistent or psychopathic offenders have functional brain differences as compared to controls (Raine et al., 2003; Weber, Habel, Amunts, & Schneider, 2008; Yang et al., 2005). The second avenue for future research involves the identification and examination of different proxies for neuropsychological deficits (Moffitt, 1990; Piquero, 2001). The current study employed three measures that have been used previously to index the presence of neuropsychological deficits. Certainly, these are not the only measures that can serve as proxies, and therefore, Hypothesis 1 must be considered in future work.
Hypothesis 2 examined the effect of parental influences on LCP offending. Early criminological theorizing (e.g., Hirschi, 1969) exalted parenting as a key component to antisocial behavior. More recent analyses suggest that parenting may not be as consequential for offending (or to correlates of offending) as was once believed (J. Harris, 1998; Pinker, 2002; Wright & Beaver, 2005). To add to this literature, the current study examined the influence of parenting on LCP offending in two ways. First, the effects of six parenting scales were analyzed after controlling for genetic risk factors toward LCP offending. The results from these tests revealed that none of the parenting measures predicted LCP offending (Table 2).
The second way that parenting was analyzed was as a nonshared environmental factor. Although parenting is typically conceptualized as a shared environmental influence (i.e., it is typically argued that parents treat their children the same, thus falling under shared environmental influences; J. Harris, 1998), some have argued that parenting may operate as a nonshared environmental factor to the extent that siblings interpret parenting differently (Turkheimer & Waldron, 2000). To explore this possibility, each of the six parenting scales was entered as a nonshared environmental influence. The results from these models provided little evidence to suggest that parenting operated as a nonshared environmental influence. In fact, only one of the scales reached statistical significance (Maternal Involvement).
Collectively, the results provided very weak support for Hypothesis 2 (i.e., parental influences affect LCP offending after controlling for genetic factors). Indeed, only 1 out of 12 tests revealed a significant influence. Note also that the correlation between the parenting scales and persister status could reflect child-driven effects (i.e., parenting strategies are a reaction to the child’s behavior; see Burt, McGue, Krueger, & Iacono, 2005; Larsson, Viding, Rijsdijk, & Plomin, 2008). If so, then the limited relationships found here are even more suspect because they may reflect child-driven effects or, at a minimum, bidirectional effects. It is also worth noting that the parenting variables used in the current study were drawn from Wave 1 of the Add Health and, therefore, captured parenting as it occurred during adolescence. Some evidence suggests that parenting effects are identifiable in the first years of life but fade over time (Ferguson, 2010). The pattern of results presented herein is certainly compatible with this reasoning; shared environmental factors may matter in childhood and, to a lesser extent, in adolescence and adulthood (see, generally, J. Harris, 1998; Sampson & Laub, 1993). In summary, the current findings suggest that criminologists should begin to look elsewhere when trying to identify the environmental (especially, nonshared environmental) influences for LCP offending. To offer one suggestion, it may turn out that parenting styles and strategies (as analyzed here) are inconsequential for LCP offending as measured during adolescence or adulthood, but this says nothing about other parental risk factors, such as intergenerational transmission effects (Boutwell & Beaver, 2010; Rowe & Farrington, 1997). Criminology will do well to expand the environmental focus to factors beyond parental strategies (Wright & Beaver, 2005).
The third hypothesis examined by the current analysis is sometimes referred to as Moffitt’s (1993) interactional hypothesis (Tibbetts & Piquero, 1999). Specifically, Hypothesis 3 argued that the effect of genetic risk on LCP offending would be strengthened in the presence of environmental risk factors, such as poor parenting or a disadvantaged neighborhood. This hypothesis was examined in two models, and neither produced results consistent with the hypothesis. Neither the Maternal Scale nor the Neighborhood Disadvantage Scale interacted with the Genetic Risk Scale to predict LCP offending. This finding stands in contrast to a sizable body of research (Caspi et al., 2002; Moffitt, 2006). For this reason, it is only appropriate to speculate as to why the findings turned out the way they did. Two explanations are possible. First, it may be the case that the Maternal Scale and Neighborhood Disadvantage Scale do not adequately tap the “adverse rearing environment” described by Moffitt (1993). Although this is certainly a possibility, it should be noted that both scales were developed out of prior research that has shown robust environmental effects on offending (Wright & Beaver, 2005; Sampson et al., 1997).
The second explanation for the lack of supportive evidence for Hypothesis 3 has roots in the differences between shared and nonshared environmental influences. Recall that the nonshared environment has consistently been found to explain the largest portion of environmental variance on offending (Ferguson, 2010; Moffitt, 2005). The measures employed by the current study may, however, tap shared environmental influences. If this is the case, and the Maternal Scale and the Neighborhood Disadvantage Scale are actually shared environments, then the current results are less surprising (J. Harris, 1998). Nonetheless, the fact that the current findings are inconsistent with a large body of research should compel future studies to reanalyze these relationships using different data sets and different operationalizations of the environmental measures.
The fourth and final hypothesis examined was generated as a by-product of Moffitt’s (1993) theory being blended with recent behavioral genetic research (Ferguson, 2010; Moffitt, 2005). Specifically, Hypothesis 4 stated that the neuropsychological deficits measures and the parental influence measures would operate as nonshared environmental influences on LCP offending. Some limited support was found for Hypothesis 4. Indeed, two variables emerged as a statistically significant (and in the predicted direction) predictor of LCP offending: self-control and maternal involvement. These results indicated that siblings with lower levels of self-control and siblings with lower levels of maternal involvement were more likely to be identified as persistent offenders. The limited support received for Hypothesis 4 should be interpreted with caution because the current analysis was exploratory in the sense that it is unclear whether (and to what degree) each of the environmental variables reflects nonshared environmental influences. Take, for instance, the Maternal Scale. Relying on criminological theorizing (e.g., Sampson & Laub, 1993) would suggest that parenting variables should act as shared environmental factors. This may be the case, but scholars have also argued that many ostensible shared environmental influences will have nonshared environmental effects to the extent that two individuals have different perceptions or interpretations of the event (Turkheimer & Waldron, 2000). Thus, for this reason, the current analysis sought to uncover whether any of the included variables operated as nonshared environmental influences. Overall, the results from Hypothesis 4 are encouraging for criminological research because they reveal that much work is left to be done in terms of uncovering the nonshared environmental factors that affect persistent offending.
Limitations of the current study must be acknowledged. First and foremost, the measure of LCP offending may include measurement error. Although others have taken a similar approach to identifying LCP offenders (Barnes et al., 2011; Turner et al., 2007), scholars have struggled with properly identifying the different offending patterns (Laub & Sampson, 2003; Moffitt, 2006) and a consensus has yet to be reached. As such, the current results must be viewed as tentative until they are replicated with alternative measurement strategies. The second primary limitation also hinges on measurement. Specifically, the identification and measurement of neuropsychological deficits poses a difficult task for social science researchers. At its core, the term neuropsychological deficit refers to an abnormality (however small) in brain formation or function (Wright, Tibbetts, et al., 2008). This means that a proper measurement of neuropsychological deficits requires access to brain scans or analyses of brain functioning. These technologies have only recently begun to make their way onto the criminological landscape (Moffitt, Ross, & Raine, 2011) and will require time before they are readily available to a majority of researchers. Nonetheless, priority should be given to studies that replicate the findings here by using more direct measures of neuropsychological deficit.
In summary, the current study offers one of the first glimpses into the interconnections between genetic risk, neuropsychological deficit, parental influences, neighborhood influences, and LCP offending. No study has sought to examine the link between various environmental influences and LCP offending after accounting for genetic factors. In this way, the present research moves the body of developmental and life course research forward to a clearer understanding of the mechanisms that play a role into the etiology of LCP offending.
Footnotes
Author’s Note
This research uses data from Add Health, a program project directed by Kathleen Mullan Harris and designed by J. Richard Udry, Peter S. Bearman, and Kathleen Mullan Harris at the University of North Carolina at Chapel Hill and funded by Grant P01-HD31921 from the Eunice Kennedy Shriver National Institute of Child Health and Human Development, with cooperative funding from 23 other federal agencies and foundations. Special acknowledgment is due Ronald R. Rindfuss and Barbara Entwisle for assistance in the original design. Information on how to obtain the Add Health data files is available on the Add Health website: ![]() . No direct support was received from Grant P01-HD31921 for this analysis.
. No direct support was received from Grant P01-HD31921 for this analysis.