
Abstract
The calculation of doses to organs and tissues of interest due to internally emitting radionuclides requires knowledge of, amongst other quantities, the fraction of energy emitted from a source region which is absorbed per mass of a target region. This quantity is the specific absorbed fraction (SAF). Publication 133 contains SAF values for the reference adults, and the Publication 155 contains the same for the reference paediatric individuals along with explanations for their derivation and use. Computation of SAF values involves simulating radiation transport in computational models which represent the geometry of the reference individuals. The reference voxel phantoms of Publications 110 and 143 were used for photon and neutron transport and most of the electron transport. Additional computational models were used for charged particles in small, overlapping, or interlaced geometries. SAF values developed for the next general recommendations will benefit from the development of mesh phantoms including those in Publication 145. Finally, additional opportunities for improvements to SAF values will be discussed.
INTERNAL DOSIMETRY AND THE SPECIFIC ABSORBED FRACTION
ICRP Committee 2 oversees the computation of internal dose coefficients. These coefficients give the effective dose to a reference individual per unit activity of intake. In this section, the quantities used to obtain the internal effective dose coefficient are reviewed with an emphasis on the role of the specific absorbed fraction (SAF).
Brief review of ICRP internal dosimetry
An intermediate step in calculating an effective dose involves the determination of the equivalent dose rate,
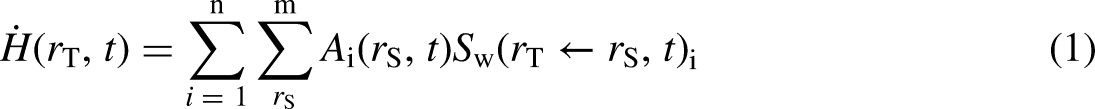
The S-coefficient is obtained as described in Eq. (2). It gives, for a given radionuclide, the equivalent dose to a target region per nuclear transformation that takes place in a source region, or correspondingly, the equivalent dose rate per unit activity in a source region. The summations are over the radiation type,
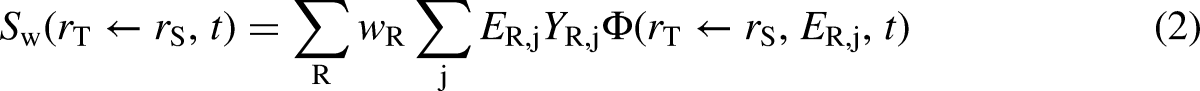
The SAF is the fraction of energy emitted from a source region which is absorbed in a target region (the absorbed fraction) per mass of the target region,
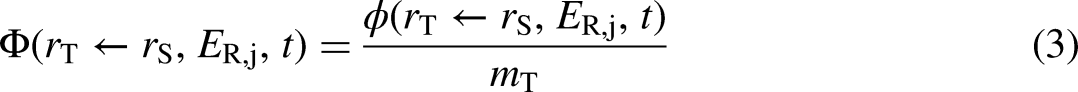
Reference individuals
The ICRP reference individuals consist of a set of idealised males and females with defined anatomical and physiological characteristics for the purpose of radiological protection (ICRP 2007). Thus, the reference individuals are theoretical constructions designed for the purpose of computing the effective dose. Reference individuals are not reference phantoms, although phantoms can be designed based on the parameters of the reference individuals. The SAF values provided in Publications 133 and 155 (ICRP 2016a, 2023) are based on the reference individuals defined in Publication 89 (ICRP 2002). Publications 133 and 155 provide additional reference mass data not included in Publication 89 as well as improvements for a few tissue regions.
For the first time, ICRP has modelled a whole-body blood source where that blood is distributed throughout tissues and vasculature in the body (ICRP 2002; Wayson et al. 2018). Understandable confusion has resulted in differences between the masses of organs in the voxel phantoms of Publications 110 and 143 (ICRP 2009, 2020b) and the target masses of the same organs in Publications 133 and 155. It was the intention of phantom developers to include appropriate blood content in the organs. Regrettably, the misinterpretation of Table 2.8 in Publication 89 led to the Publication 110 and 143 voxel phantoms not appropriately assigning the mass of systemic blood. Consequently, for geometries where the target and source regions are the same or overlapping (self-irradiation geometries), the SAF computed in the phantoms must be adjusted to account for the missing organ blood mass. Publication 155 describes approaches for making this adjustment and when it is or is not desirable.
The set of reference individuals is defined at six ages (newborn; 1, 5, 10, 15 years; and adult) with two sexes at each age. The biokinetic models define 79 source regions, and the definition of effective dose along with inclusion of other targets of interest (e.g. lens of the eye) results in 43 target regions. When considering four radiation types (alpha, electron, photon, and spontaneous fission neutrons) and the energy grids associated with each, the full set of age- and sex-dependent SAF values consists of more than three million data points.
Supporting models
Tissue geometries
As described in the previous section, the mass of tissue regions is defined in Publication 89, but with only a few exceptions, additional information is needed to describe the shape and relative proximity of tissue regions to one another. The latest SAF values represent a significant improvement in that they are informed by image-based models of the tissue geometries rather than solely supported by stylised, mathematical models. These image-based models include the previously mentioned voxel phantoms of Publications 110 and 143 but also include models of the skeletal microstructure for determining SAF values for intraskeletal geometries (Pafundi et al. 2010; Hough et al. 2011; O'Reilly et al. 2016). Stylised models are used for geometries interior to the alimentary and respiratory tracts (ICRP 1994, 2006).
Radiation transport
Simulations of radiation transport through the modelled tissue geometries can be performed using an established Monte Carlo radiation transport code. In addition to finding descriptions of the methods for computing SAF values in the ICRP publications, separate publications (Schwarz et al. 2021b, 2021a; Griffin et al. 2022) describe greater detail for computations in the voxel phantoms.
Example SAF values
The full set of SAF values is provided as an electronic download accompanying Publication 155. Multiple example plots are provided in the text of Publication 155 as well as explanations of theoretical checkpoints and their basis. In this article, one such example is provided.
Fig. 1 displays a set of SAF values versus electron energy from Publications 133 and 155. Values for all six reference ages are provided for the left colon wall irradiating the left colon stem cell region. This geometry is an example of the source region fully encompassing the target region. For such regions a theoretical limiting value of the SAF can be computed for electrons of energy approaching zero. This limit is simply the volume fraction of the stem cell region to the wall divided by the mass of the target region. In this case the SAF does not fall away from this limiting value until electrons of about 200 keV or more are considered.
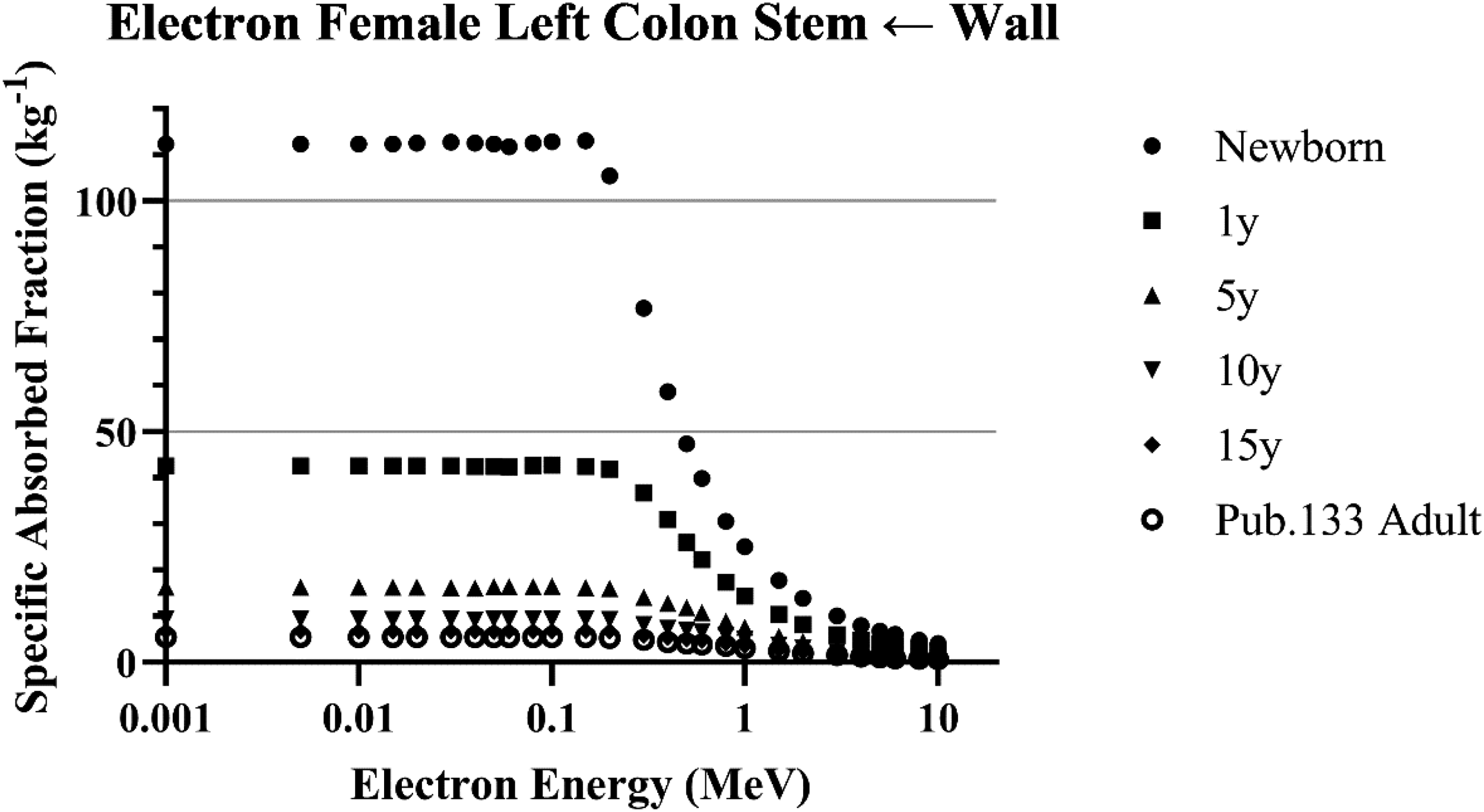
Specific absorbed fractions for electrons emitted from the left colon wall irradiating the left colon stem cell target region for females at all reference ages (ICRP 2023).
The SAF values in Publication 133 were used to support the computation of internal dose coefficients for the occupational intakes of radionuclides (OIR series) (ICRP 2015, 2016b, 2017, 2019, 2022). Similarly, the age-dependent SAF values in Publication 155, which includes the adult values from Publication 133, support the computation of internal dose coefficients for members of the public exposed to radionuclides in the environment (ICRP 2024a). The age-dependent SAF values are also supporting work in progress from ICRP Task Group 36 on radiation dose to patients in diagnostic nuclear medicine and ICRP Task Group 112 on emergency dosimetry.
POTENTIAL IMPROVEMENTS IN INTERNAL DOSIMETRY FOR USE IN THE NEXT SYSTEM
New target and source geometries
The first improvement which may be needed to support dosimetry associated with the next system of radiological protection involves modelling new target regions. The science is still evolving on which tissues are sensitive to ionising radiation. For example, if cardiovascular disease is deemed to be a disease associated with exposure to ionising radiation (Zanzonico et al. 2016; Boice Jr et al. 2022; Little et al. 2023), the target tissue will need to be identified and modelled appropriately.
New target models may also be of benefit as new knowledge is gained about where within a volume the sensitive tissues or cells reside. For example, Eckerman and Veinot (2019) have developed a new model for the region where at-risk cells reside in the urinary bladder wall.
And in some cases, new knowledge on the non-uniformity of the distribution of material within an organ may drive a need for new models even if the target remains the same. For example, NCRP Commentary 31 proposes new biokinetic models for certain elements in subregions of the brain where data show non-uniformity of activity (Leggett et al. 2022; NCRP 2022).
Model improvements
Even where there are not new definitions of source or target regions, improvements to models can be made which are more representative of the specific body region. At the whole-body level, the recently developed ICRP mesh phantoms of Publications 145 and 156 (ICRP 2020a, 2024b) represent improvements to the surfaces of regions over their voxel ancestors. A future task will involve determining for which source–target combinations and radiation types the mesh phantoms will represent significant improvements to internal dosimetry applications. For many of the important geometries, the SAF value will not be sensitive to a mesh versus a voxel surface.
Alimentary tract
Choi et al. (2022) recently published mesh versions of the alimentary tract which include delineation of the small source and target regions in various regions lining the tract. While the definitions of these regions are based on information in Publication 100 (ICRP 2006), the inclusion inside the whole-body mesh phantom allows for radiation crossfire across regions of the alimentary tract organs and into other tissues in the body. A significant improvement contained within this work is a realistic anatomy of the oral cavity and surrounding tissues.
Respiratory tract
Similar to the work on the alimentary tract, Choi et al. (2023) have published work on a mesh version of the respiratory tract which fits inside the whole-body mesh phantoms. Based primarily on the definitions provided in Publication 66 (ICRP 1994), this model will allow for radiation crossfire through regions of the lung which could not previously be handled with the stylised cylindrical models.
Skeleton
To date, the internal microstructure of the skeleton has required separate modelling from the whole-body phantoms. Preliminary work (Choi et al. 2021; Shin et al. 2022) has demonstrated the ability to fold the microstructure into the whole-body phantoms. This will allow for radiation crossfire across skeletal sites into surrounding tissue (e.g. cranium and brain), as well as for the study of the accuracy of photon and neutron skeletal dose response functions since such response functions will not be required in such a model.
Another potential improvement to skeletal dosimetry modelling could involve revisiting the definition of source regions inside the skeleton. The separation of bone source regions into planar and volume sources was necessary in the past given practical modelling considerations. But its intent, to model a non-uniform activity distribution in bone mineral which varies with depth, can now be modelled without requiring approximation as an unrealistic planar surface region.
Vasculature
As mentioned earlier, the latest models used to compute dose coefficients take advantage of a whole-body blood distribution model and treat blood as an explicit source region. This is an improvement, but with only a few exceptions, the energy deposition models do not distinguish the parenchyma tissue from blood in the vasculature perfusing the organ. Xing et al. (2022) and Correa-Alfonso et al. (2022) have published work which explicitly models the vasculature interior to the liver. Such work will allow for consideration of energy losses from short-range radiations (alpha particles and low-energy electrons) separately considering dose to the parenchyma tissue, blood, and vessel walls.
Sex-specific biokinetic modelling
Finally, in reference to Eq. (1), it is helpful to remember that while the energy absorption term (S-coefficient) is sex dependent, the biokinetic models which inform the source term are not sex dependent. This dichotomy presents an inconsistency which has the potential to increase or decrease a dose coefficient. Note that frequently there may be no need for sex-dependent biokinetic models. That is, many elements behave with similar kinetics in females and males. But where the kinetics does vary with sex and there is sufficient data available to support it, sex-specific biokinetic modelling should be developed to bring consistency with the sex specificity in the energy absorption models.
Footnotes
ACKNOWLEDGEMENTS
This article has been authored by UT-Battelle, LLC, under contract DE-AC05-00OR22725 with the US Department of Energy (DOE). The US government retains, and the publisher, by accepting the article for publication, acknowledges that the US government retains a non-exclusive, paid-up, irrevocable, worldwide licence to publish or reproduce the published form of this manuscript, or allow others to do so, for US government purposes. DOE will provide public access to these results of federally sponsored research in accordance with the DOE Public Access Plan (![]() ).
).
FUNDING
The author disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work described in this article was sponsored by the Office of Radiation and Indoor Air, US Environmental Protection Agency (EPA), under Interagency Agreement DOE No. 182-Z614-23, under contract No. DE-AC05-00OR22725 with UT-Battelle.