
Abstract
Previous studies show that newborn infants and adults orient their attention preferentially toward human faces. However, the developmental changes of visual attention captured by face stimuli remain unclear, especially when an explicit top-down process is involved. We capitalized on a visual search paradigm to assess how the relative strength of visual attention captured by a non-target face stimulus and explicit attentional control on a target stimulus evolve as search progresses and how this process changes during development. Ninety children aged 5–14 years searched for a target within an array of distractors, which occasionally contained an upright face. To assess the precise picture of developmental changes, we measured: (1) manual responses, such as reaction time and accuracy; and (2) eye movements such as the location of the first fixation, which reflect the attentional profile at the initial stage, and looking times, which reflect the attentional profile at the later period of searching. Both reaction time and accuracy were affected by the presence of the target-unrelated face, though the interference effect was observed consistently across ages. However, developmental changes were captured by the first fixation proportion, suggesting that initial attention was preferentially directed towards the target-unrelated face before 6.9 years of age. Furthermore, prior to 12.8 years of age, the first fixation towards face stimuli was significantly more frequent than for object stimuli. In contrast, the looking time proportion for the face stimuli was significantly higher than that for the objects across all ages. These findings suggest that developmental changes do not influence the later search periods during a trial, but that they influence the initial orienting indexed by the first fixation. Moreover, the manual responses are tightly linked to eye movement behaviors.
Introduction
The face conveys an abundance of socially relevant visual information associated with personality traits (Willis & Todorov, 2006) and identity (Bruce & Young, 1986) that are important for social interaction. The developmental changes in face processing follow a somewhat different trajectory than those associated with domain-general functions such as object processing (McKone, Crookes, Jeffery, & Dilks, 2012). The theoretical framework for the development of face processing suggests the involvement of two independent cooperative systems (Johnson, 2005; Morton & Johnson, 1991). A subcortical face detection system involving regions such as the superior colliculus, pulvinar nucleus, and amygdala is responsible for the rapid orientation of attention towards faces, while a cortical system involving regions such as the fusiform gyrus, inferior occipital gyrus, and superior temporal sulcus is responsible for facial identification (Haxby, Hoffman, & Gobbini, 2000; Kanwisher, McDermott, & Chun, 1997).
Most previous developmental studies on face perception/recognition have focused on the configural aspects (i.e., spacing of facial features) of face processing and the cortical representation of faces during childhood (Lee, Anzures, Quinn, Pascalis, & Slater, 2011). A number of these studies have demonstrated that changes in such face processing abilities follow nonlinear developmental trajectories. Several neuroimaging studies have also noted the nonlinear developmental trajectories of face processing using functional magnetic resonance imaging and electroencephalography, revealing that the neural correlates related to face processing typically mature around adolescence (Itier & Taylor, 2004; Scherf, Behrmann, Humphreys, & Luna, 2007; Taylor, Batty, & Itier, 2004). For example, researchers have revealed that mature levels of activation in face-selective cortical regions emerge around the age of 11 years (Scherf et al., 2007) and that the face-selective N170 event-related potential (ERP) component at the age of 11 years is also comparable to that of adults (Itier & Taylor, 2004; Taylor et al., 2004).
However, the developmental trajectory of the subcortical face-orienting system may differ from that of the cortical face-processing system. This subcortical face detection system seems to mature in the early stages of development, as newborns exhibit preferences for facial and face-like pattern stimuli (Johnson, Dziurawiec, Ellis, & Morton, 1991). Moreover, orienting attention towards faces dramatically changes during the first year of life (Amso, Haas, & Markant, 2014; Di Giorgio, Leo, Pascalis, & Simion, 2012; Frank, Vul, & Johnson, 2009; Gliga, Elsabbagh, Andravizou, & Johnson, 2009; Gluckman & Johnson, 2013). For example, several studies have confirmed that 6-month old infants orient spontaneously toward upright faces when compared with other objects or body parts (Frank et al., 2009; Gliga et al., 2009; Gluckman & Johnson, 2013), and that 3-month-old infants are less consistent in where they look than older infants. Moreover, younger infants are more likely to fixate on stimuli with low-level image salience, rather than on the locations of faces; between 3 and 9 months of age, infants gradually begin to focus their attention on faces. These findings suggest that newborn infants have the capacity for spontaneous orientation of visual attention towards faces, but that the manner in which such attention is oriented towards faces dramatically changes during the first year of life.
A number of studies utilizing face-like (protoface) stimuli have revealed that both 5- to 6-year-old children (Shah, Happe, Sowden, Cook, & Bird, 2015) and adults (Tomalski, Csibra, & Johnson, 2009) exhibit rapid orientation towards facial stimuli. Moreover, participants ranging in age from 4-month-old infants through adulthood spontaneously shift their visual orientation towards faces in a natural scene, although changes in the influence of visual salience occur across development (Amso et al., 2014). These findings suggest that orienting towards face/face-like stimuli can be observed even during the early stages of development. Indeed, converging evidence suggests that this subcortical route may provide a developmental foundation for the adult “social brain” network in the cortex, and that dysfunction of this pathway may be associated with certain developmental disorders such as autism spectrum disorders (ASD) (Dalrymple et al., 2018; Di Giorgio et al., 2016).
Although research supports the notion that differential developmental patterns underlie the processes of face detection and face identification, the neural mechanisms underlying the ability to attend to faces presented in competition with other non-face stimuli during childhood—as well as the developmental changes associated with this ability—remain to be elucidated. One important factor influencing the ability of target-unrelated faces to capture visual attention is the presence of configural face information. Indeed, a previous behavioral study revealed that inverted faces do not capture visual attention (Langton, Law, Burton, & Schweinberger, 2008). Other studies have indicated that developmental changes in the configural processing of faces occur by age 11 (Itier & Taylor, 2004; Scherf et al., 2007; Taylor et al., 2004), suggesting that the orienting mechanisms also undergo modulation in accordance with the development of face processing (i.e., by the age of 11). Moreover, Shah, Happe, Sowden, Cook, and Bird (2015) have suggested that orientation to face stimuli may be stronger during critical periods of development, when input into the developing visual system may facilitate the emergence of perceptual expertise. Therefore, the strength of visual attention captured by faces may change during childhood due to the development and refinement of the visual system, including aspects related to the configural processing of faces.
In order to elucidate the developmental trajectory of orientation towards faces during childhood, previous studies from our research group utilized a task originally developed by Langton, Law, Burton, and Schweinberger (2008). In this task, participants were asked to search for a target butterfly among six objects that were placed in a circular array. This arrangement ensured objects were equidistant from the center of the screen. In some trials, a distractor was replaced with a face, which allowed for a comparison of the relative strength of goal-directed searching and stimulus-driven attention. Both manual responses such as reaction time and eye movements were measured in order to formulate more complete attentional profiles capturing different periods of the visual search (Hirai et al., 2016a, 2016b). These studies demonstrated that behavioral measures such as reaction time and accuracy are not sensitive enough to capture the atypical pattern of orientation towards faces in cases of the developmental disorder known as Williams syndrome (Hirai et al., 2016a, 2016b). More precisely, the location of the first fixation and the duration of subsequent fixations were analyzed in order to investigate the temporal dynamics of the relative dominant ability of a target-unrelated face to capture visual attention. Atypical looking time for target-unrelated faces, but not atypical first fixation, was observed. Because the initial fixation is rapid, it reflects the attentional profile during the early search period. Looking time, in contrast, indicates how much time participants gazed at each item during the search, reflecting the attentional profile at later periods. In summary, the atypical attentional characteristics captured by target-unrelated faces in individuals with Williams syndrome are not observed during the initial search period but become dominant as the search progresses. It is worth emphasizing the potential for these two eye-movement measures, first fixation and looking time, to assess the relative strength of visual attention capture by a target-unrelated face and explicit attentional orientation towards the target, respectively.
In the present study, we predicted several outcomes from the viewpoint of manual responses, eye movement behaviors, and developmental changes. For the manual responses, the reaction time would be longer in face-present trials than in face-absent trials for children when compared to previously reported results for adults (Langton et al., 2008), as the face identification system seems to mature during adolescence. For the eye movements, if visual attention captured by a target-unrelated face is relatively stronger than the explicit attentional orientation towards the target (i.e., butterfly) in the early search period, the first fixation should frequently fall on the face rather than on the target object. Similarly, in the later search periods, if visual attention captured by a target-unrelated face is relatively stronger than the explicit attentional orientation towards the target object, looking time on faces should be longer than those on the target object. Regarding the relationship between gaze behaviors and manual responses, if gaze behaviors at different periods of a visual search are tightly linked to reaction time, reaction time would be significantly affected by eye movement indices. More precisely, we predicted that explicit attentional control is dominant during the initial search period. As a result, reaction time should negatively correlate with the frequency of initial butterfly-fixations. However, if visual attention towards faces is stronger than explicit attentional control during the initial search period, then reaction time should positively correlate with the frequency of the first fixation towards faces. Additionally, if the visual attention captured by face stimuli is stronger during the later periods of searching, reaction time should positively correlate with the duration of face fixations; otherwise, it should negatively correlate with the duration of butterfly fixations. Regarding developmental changes, the proportion of trials during which the first fixation corresponded to faces would not change across development, as previous studies have revealed that the face orienting system matures in the early stages of development. Moreover, if the face orienting system indeed matures during the early stages of development, then the looking time on target-unrelated faces would be similar after the early stages.
Methods
Participants
Ninety children, free of neurological disorders, ranging from 5 to 14 years of age (mean = 9.8 years; standard deviation = 2.3) were recruited from nearby elementary and junior high schools; participants from our previous eye-tracking studies were excluded. Written informed consent for the children to participate in this study, approved by the institutional ethics committee in accordance with the Declaration of Helsinki on ethical principles for medical research, was provided by the parents of all children.
Stimuli and Experimental Apparatus
Experiments were conducted using a computer equipped with Tobii Studio and E-prime 2.0 software (Psychology Software Tools, Inc., PA, USA) with the E-prime extension for Tobii (Tobii, Inc., Stockholm, Sweden). Stimuli were presented on a 24-inch liquid crystal display color monitor (PLE2407HDS; Iiyama, Tokyo, Japan), placed approximately 60 cm from the observer. As in the study by Langton and colleagues (2008), six images of objects (fruits, flowers, leaves, trees, and houseplants) including a face, were displayed in a circular array (Figure 1). All images were randomly taken from a copyright-free database.
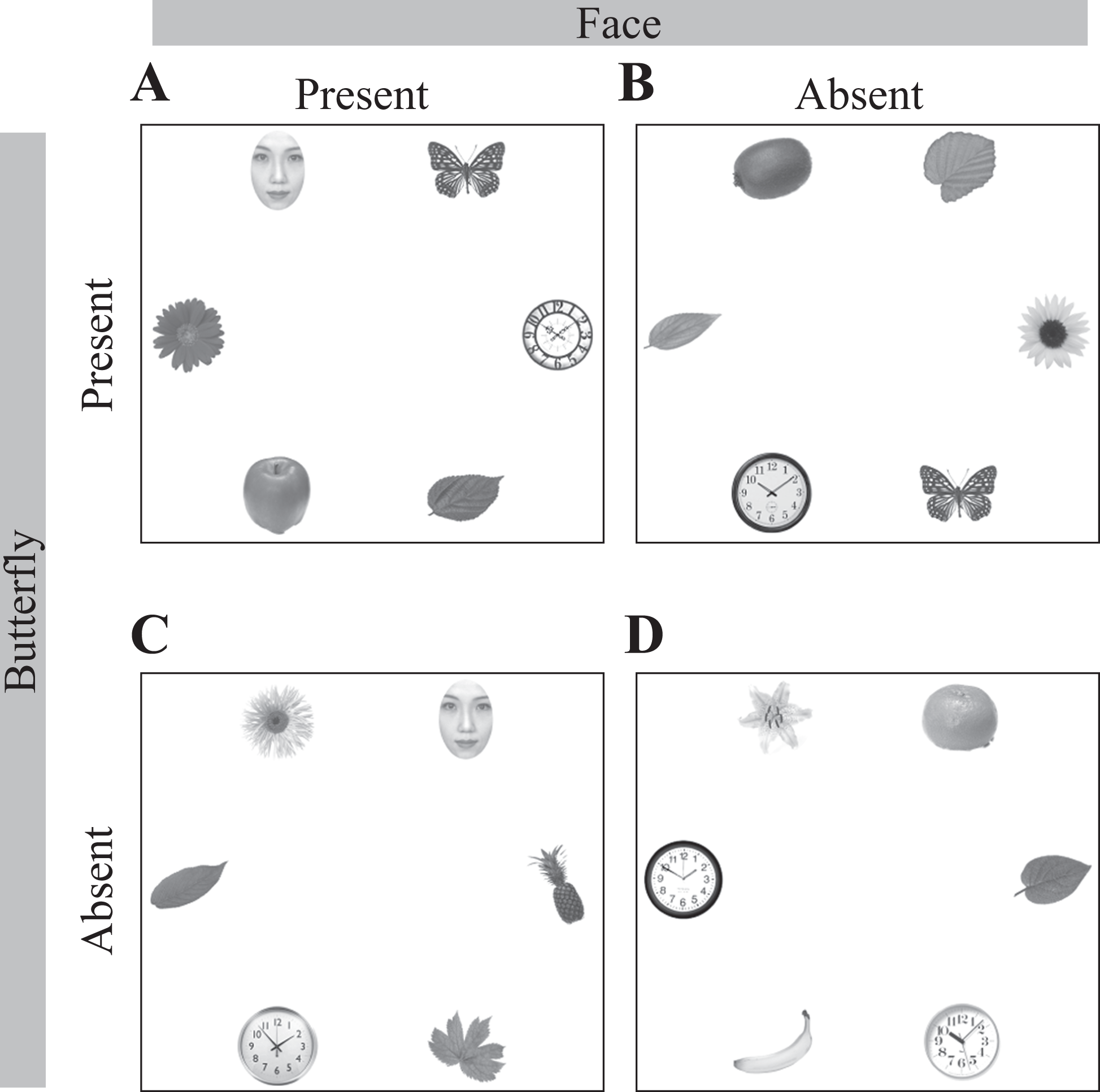
A schematic of the four experimental conditions: (A) face-present, butterfly-present; (B) face-absent, butterfly-present; (C) face-present, butterfly-absent; and (D) face-absent, butterfly-absent.
These objects were gray-scale shapes that fit within a 3.5° × 3.5° area. The average luminance was equated for all objects using the SHINE toolbox (Willenbockel et al., 2010). Each object was centered with a visual angle of approximately 15° from the central fixation point.
Because we aimed to determine how a target-unrelated face stimulus may modulate gaze behavior when both a target and a face are present, we included four conditions based on the presence and absence of two factors: target and face. Thirty arrays were created for each experimental condition, yielding a total of 120 trials. The target objects were six different pictures of butterflies. The face distractors, obtained from a commercially available database (ATR, Kyoto, Japan), comprised eight different upright faces (gender balanced); all faces had neutral expressions and were cropped to remove external features such as hair and ears. The other distractors were varied exemplars from the categories of fruits, flowers, leaves, clocks, and houseplants. Non-face distractor locations in each array were filled with randomly selected objects from these five object categories. In the butterfly-present and face-present conditions (Figure 1A), a target butterfly appeared at one of the six locations and a face appeared at one of the remaining five. Each trial had a unique butterfly–face pairing. Target-present and face-absent arrays (Figure 1B) were created by replacing the face in each array with an object that was randomly selected from an object category without using two identical pictures.
For the target-absent and face-present conditions (Figure 1C), a face was located at one of the six locations and the remaining distractor locations in each array were filled with randomly selected objects from the five categories. For the target-absent and face-absent conditions (Figure 1D), the array was created by selecting six different objects from the five categories.
Behavioral responses were reported via a custom-made response box with two large buttons. Eye movements were recorded using the Tobii X60 eye-tracking system. The system tracked both eyes with an accuracy of 0.5° and a sampling rate of 60 Hz. The eye-tracker was calibrated for each participant using a five-point calibration for each eye. All participants underwent a successful five-point calibration prior to testing.
Task Procedure
To record reliable eye-movement data during each trial, the stimulus array was not presented until participants had fixated upon a cross at the center of the screen for 1 second (Figure 2).
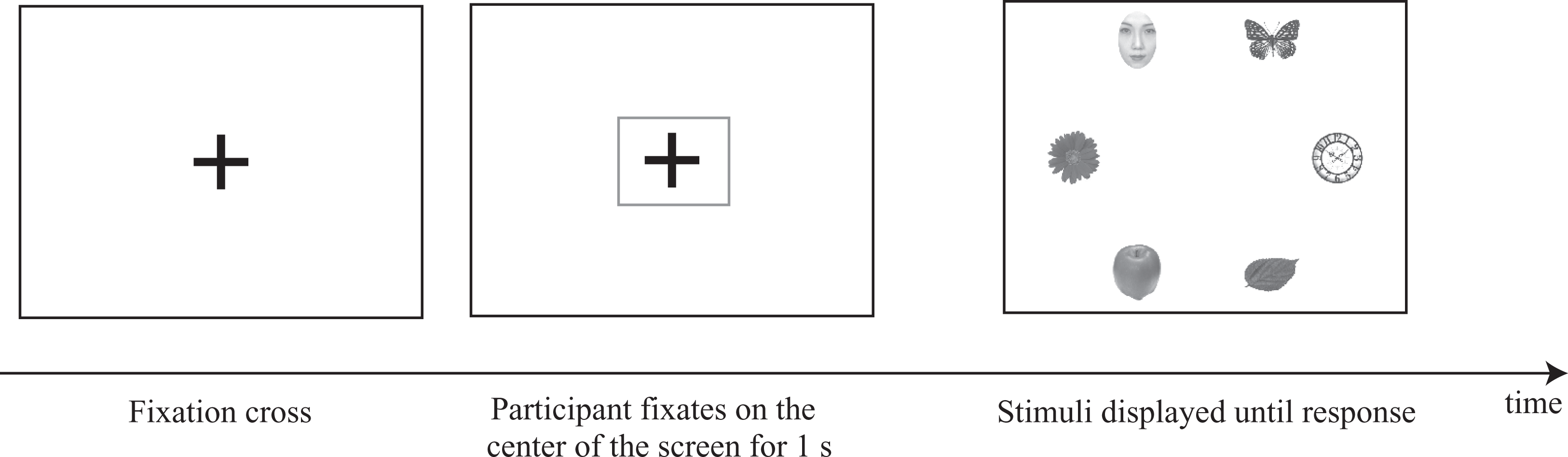
The experimental procedure. Stimuli were displayed after the participant had fixated on the central cross for 1 second. After the participant had pressed a response button, the stimuli disappeared.
Participants were instructed to focus on finding the butterfly and were not told that faces would be appearing on the screen. They judged whether a butterfly was present in each array as quickly and accurately as possible and recorded their responses by pressing one of the two response box buttons. No feedback was given to participants. Half of the participants were asked to use their left hand to press the left button if they saw the butterfly and the right button if they did not. The other half were given the opposite instructions. Before testing, participants performed three trials for each condition which led to 12 practice trials in total.
The experiment consisted of three sessions, each consisting of 40 trials (10 per condition). Trials within a session were randomized, as was the session order. Each trial lasted approximately 2 seconds, depending on the participant’s performance, and ended after the participant responded.
Data Analysis
We analyzed manual responses and eye movements utilizing a previously reported developmental trajectory approach (Annaz, Karmiloff-Smith, Johnson, & Thomas, 2009). For this, we constructed cross-sectional trajectories for manual and eye movement measures. The analysis was performed in two phases. The first phase involved covariate omission to explore within-subject effects. In the second phase, we added age as a covariate and performed an analysis of covariance (ANCOVA) using the procedure described on the website of the Developmental Neurocognition Laboratory at Birkbeck College (http://www.psyc.bbk.ac.uk/research/DNL/stats/Thomas_trajectories.html). The point at which two trajectories converged was defined as the point where the two confidence interval lines crossed.
For manual responses, we analyzed reaction times and accuracy. The presence or absence of targets and faces yielded four trajectories linking reaction time and accuracy with age. A two-way analysis of variance (ANOVA) was applied in the first phase to both reaction time and accuracy data. Face (present vs. absent) and target (present vs. absent) were used as within-subject factors. In the second analysis phase, we added age as a covariate and conducted an ANCOVA.
Eye movements were recorded and analyzed using Tobii Studio 2.3.1. The inbuilt Velocity-Threshold Identification filter was applied for fixation detection. Eye movements were classified as saccades when the velocity exceeded 30°/second, and the minimum fixation duration was 60 milliseconds. One area of interest (AOI) was assigned to each item in each array; each area comprised a polygon encompassing the whole image. The size of the image was similar across items, and the distances of the AOIs from the center of the screen were identical across items (5.0°).
We evaluated two measures of gaze behavior: first fixation (Fletcher-Watson, Leekam, Benson, Frank, & Findlay, 2009); and looking time (Riby & Hancock, 2008). These measures were obtained, for the face-present and target-present conditions only, to gauge the relative attention strength captured by target-unrelated faces and the explicit attentional control toward target butterflies during the initial and later periods of directed searches across development. The first fixation corresponds to the frequency with which participants fixated on an item (face, butterfly, or objects) at the end of the first saccade. Therefore, this measure can seize the bottom-up visual attention captured by faces and the explicit top-down attentional control (toward butterfly) at the initial stage of searching. The looking time indicates how long a participant gazed at each item during the experiment. Therefore, it can represent the relative dominance of visual attention captured by faces and the explicit attentional control toward a target at the later stages of searching.
Since the time spent searching for a target varied across participants, it was necessary to compute normalized measures based on the total numbers of first fixations and looking times. The first fixation proportion was defined as the number of first fixations on an item (butterfly, face, or object) divided by the total number of trials. This measure can be used to assess which items received attentional priority in a scene. The looking time proportion was defined as the amount of time spent looking at each type of stimulus during a trial divided by the total duration of fixations for that trial. Both proportions were further modified for the face-present and butterfly-present conditions by dividing their total by the number of non-face distractors (i.e., four items). We analyzed only trials in which the percentage of recorded eye movements was above 80% of the trial duration.
In the first analysis phase, a one-way repeated measures ANOVA was performed on the eye movement measurements (first fixation proportion, looking time, and looking time proportion). Items (face, butterfly, and remaining objects) were used as within-subject factors. In the second analysis phase, we added age as a covariate and conducted an ANCOVA. We used the Greenhouse–Geisser epsilon coefficient to correct the degrees of freedom if a violation of sphericity was assessed via the Mauchly’s sphericity test. Tukey’s honest significant difference tests were performed for post-hoc pairwise comparisons. The threshold for statistical significance was set to p < 0.05.
Results
Manual Responses (Reaction Times)
In the first phase of analysis, significant main effects of Target [F(1, 89) = 238.1, p < 0.01, ηp 2 = 0.73] and Face [F(1, 89) = 48.0, p < 0.01, ηp 2 = 0.35] were observed (Figure 3A); however, the two-way interaction of Target and Face was not significant [F(1, 89) = 2.40, p = 0.12, ηp 2 = 0.03]. These results suggest that the reaction time in the face-present condition was significantly longer than in the face-absent condition (1.34 seconds vs. 1.29 seconds). Furthermore, reaction time in the target-absent condition was significantly longer than that in the target-present condition (1.15 seconds vs. 1.48 seconds).
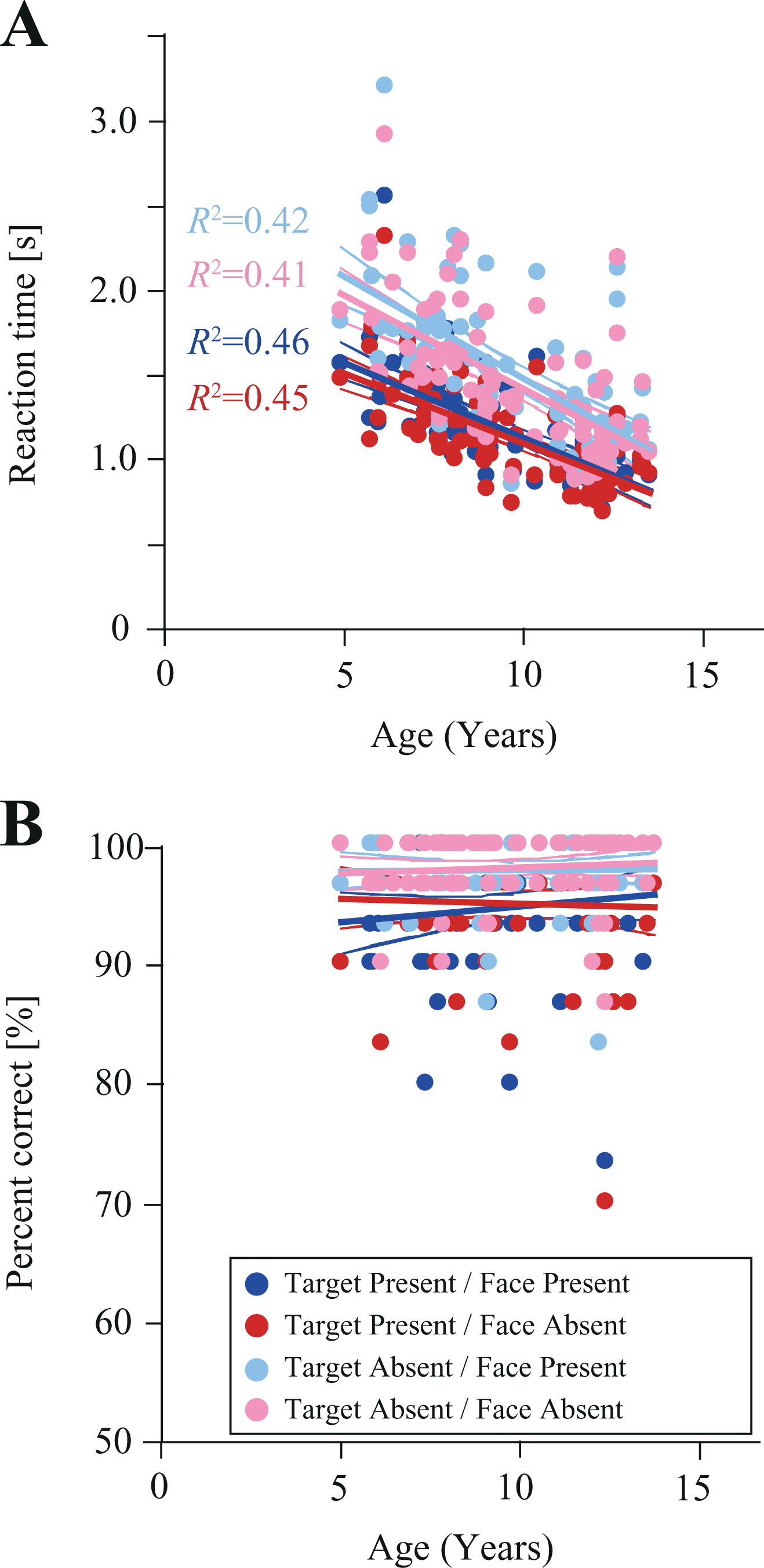
The developmental trajectories for reaction times (A) and accuracy (B) (N = 90). Red, blue, pink, and aqua lines indicate 95% confidence intervals for each experimental condition. R2 values indicate the proportion of variance explained by each trajectory.
The second phase of analysis, which included age as a covariate, revealed that performance significantly improved with age [F(1, 88) = 78.4, p < 0.01, ηp 2 = 0.47]. Moreover, main effects of Target [F(1, 88) = 98.3, p < 0.01, ηp 2 = 0.53] and Face [F(1, 88) = 21.1, p < 0.01, ηp 2 = 0.19] were observed. Again, these results suggest that reaction time was significantly longer in the face-present condition than in the face-absent condition, and that reaction time in the target-absent condition was significantly longer than in the target-present condition. Furthermore, a significant interaction of Target × Age [F(1, 88) = 11.6, p < 0.01, ηp 2 = 0.12] was observed. This interaction suggests that both the reaction times for target-present and target-absent conditions significantly decreased according to participant age, and the reaction time for the target-absent condition was significantly longer than that for the target-present condition. However, other interactions such as Face × Age [F(1, 88) = 3.4, p = 0.07, ηp 2 = 0.04], Target × Face [F(1, 88) = 0.47, p = 0.50, ηp 2 = 0.005], and Target × Face × Age [F(1, 88) = 0.002, p = 0.97, ηp 2 = 0.00] were not significant.
Accuracy
In the first phase of analysis, a significant main effect of Target [F(1, 89) = 54.8, p < 0.01, ηp 2 = 0.38] was observed on accuracy (Figure 3B). However, the main effect of Face and the interaction of Target × Face (ps > 0.96) were not. These results suggest that accuracy was significantly higher in the target-absent condition than in the target-present condition (98.0 vs. 94.9%).
In the second phase of analysis, the significant main effect of Target [F(1, 88) = 10.6, p < 0.01, ηp 2 = 0.11] was preserved when adding age as a covariate. Other effects were not significant [Fs < 2.46, ps > 0.12].
Gaze Behavior
As the aim of the current study was to gauge the relative bottom-up/top-down attention during the initial and later periods of directed searches across development, we focused on only the face-present and target-present condition. Overall, all participants fixated on the initial item in more than 90% of trials (average available trials: 27.9 for the target-present/face-present conditions). No effect of age on the available number of trials was observed (r = -0.05).
First Fixation Proportion
The developmental trajectory for the first fixation proportion is shown in Figure 4A. A significant main effect of Item was observed with regard to first fixation proportion [F(2, 178) = 96.3, p < 0.01, ηp 2 = 0.73]. The first fixation proportion for butterfly stimuli was significantly higher than that for face (34.3% vs. 18.3%, p < 0.01) and object (34.3% vs. 11.8%, p < 0.01) stimuli. Furthermore, the first fixation proportion for face stimuli was significantly higher than that for object stimuli (18.3% vs. 11.8%, p < 0.01).
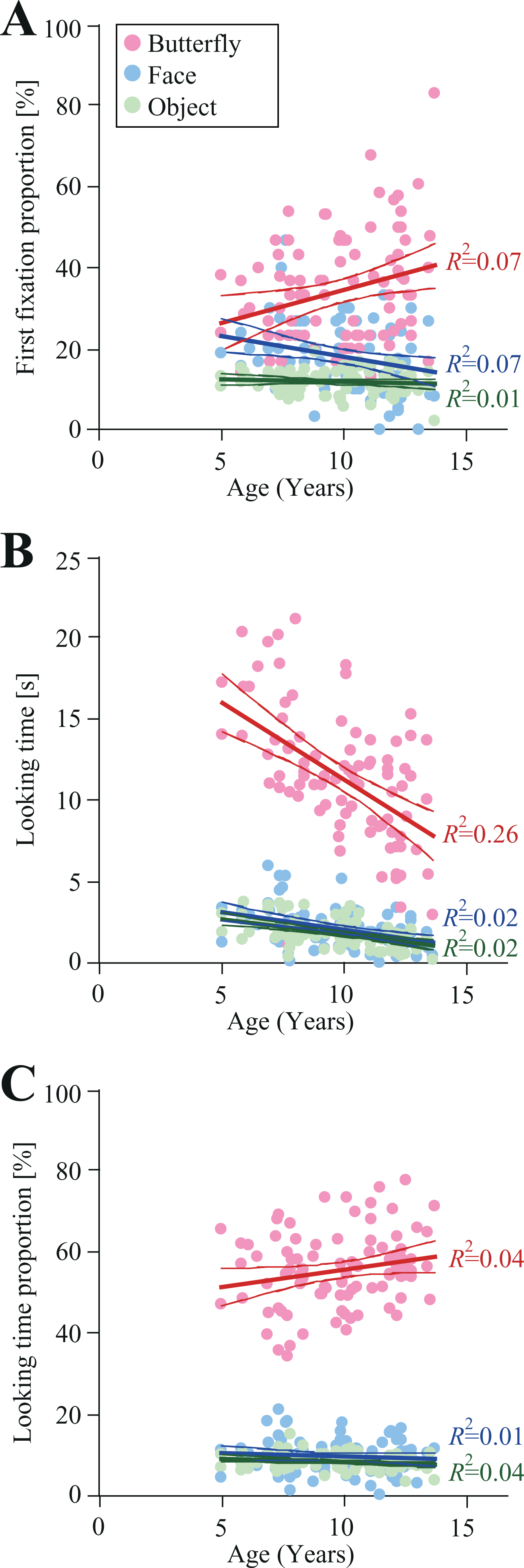
The developmental trajectories for the first fixation proportion (A), looking time (B), and looking time proportion (C) (N = 90). Pink, aqua, and green lines indicate 95% confidence intervals for gaze behavior toward butterflies, faces, and objects, respectively. R2 values indicate the proportion of variance explained by each trajectory.
In the second phase of analysis, when including age as a covariate, a significant main effect of Item [F(1.4, 176) = 7.1, p < 0.01, ηp 2 = 0.08] was still observed, while the main effect of Age [F(1, 88) = 1.1, p = 0.29, ηp 2 = 0.13] was not significant. A significant two-way interaction of Item × Age [F(1.4, 176) = 7.1, p < 0.01, ηp 2 = 0.07] was observed. The trajectories for face and butterfly converged at the age of 6.9 years. After the age of 6.9 years, the first fixation towards butterfly stimuli was significantly more frequent compared to that towards face stimuli. Both face and object trajectories converged at the age of 12.8 years. This result suggests that, prior to the age of 12.8 years, the first fixation towards face stimuli was significantly more frequent compared to that for object stimuli.
Looking Time
The developmental trajectory for looking time is shown in Figure 4B. A significant main effect of Item was observed with regard to looking time in the first phase of analysis [F(1, 89) = 9.08, p < 0.01, ηp 2 = 0.09]. Subsequent post-hoc analysis revealed that the looking times for butterfly stimuli were significantly longer than those for faces (11.5 seconds vs. 2.08 seconds, p < 0.01) and objects (11.5 seconds vs. 1.81 seconds, p < 0.01). Moreover, the looking times for faces were significantly longer compared to those for objects (2.08 seconds vs. 1.81 seconds, p = 0.01).
In the second phase of analysis, which included age as a covariate, significant main effects of Item [F(1.1, 96.7) = 237.1, p < 0.01, ηp 2 = 0.73] and Age [F(1, 88) = 40.1, p < 0.01, ηp 2 = 0. 31] were observed. Moreover, a significant two-way interaction of Item × Age [F(1.1, 96.7) = 21.6, p < 0.01, ηp 2 = 0.20] was observed, indicating that trajectories for faces and objects—but not for butterfly stimuli—converged prior to the age of 5.0 years.
Looking Time Proportion
The developmental trajectory for looking time proportion is shown in Figure 4C. A significant main effect of Item was observed with regard to looking time proportion in the first phase of analysis [F(2, 178) = 1348.0, p < 0.01, ηp 2 = 0.94]. This result suggests that the looking time proportion for butterfly stimuli was significantly higher than that for face (56.0% vs. 9.8%, p < 0.01) and object (56.0% vs. 8.5%, p < 0.01) stimuli. Furthermore, the looking time proportion for face stimuli was significantly higher than that for object stimuli (9.8% vs. 8.5%, p < 0.01).
In the second phase of analysis, when including the age as a covariate, a significant main effect of Item [F(1.2, 104.9) = 192.2, p < 0.01, ηp 2 = 0.69] was observed, although the main effect of Age [F(1, 88) = 3.79, p = 0.06, ηp 2 = 0.04] was not significant. Furthermore, the two-way interaction of Item × Age [F(1.2, 104.9) = 3.35, p = 0.06, ηp 2 = 0.04] was not significant. Subsequent post-hoc analysis revealed that the looking time proportion for butterfly stimuli was significantly longer compared to those for face (56.0% vs. 9.8%, p < 0.01) and object (56.0% vs. 8.5%, p < 0.01) stimuli. Moreover, the looking time proportion of face stimuli was significantly higher than that of object stimuli (9.8% vs. 8.5%, p = 0.02).
Association between Behavioral Responses and Gaze Behavior
The association between reaction time and first fixation proportion was also assessed in the present study (Figure 5A). We observed a significant positive correlation between reaction time and first fixation proportion for faces (r = 0.40, p < 0.01) and objects (r = 0.22, p < 0.05), while a significant negative correlation was observed between reaction time and first fixation proportion for butterfly stimuli (r = -0.43, p < 0.01). Furthermore, the association between reaction time and looking time for each item was assessed (Figure 5B). We found a significant positive correlation between reaction time and looking time for all items (rs > 0.63, ps < 0.01). Finally, the association between reaction time and looking time proportion for each item was assessed (Figure 5C). We observed significant positive correlations between reaction time and looking time proportion for object stimuli (r = 0.35, p < 0.01), but opposite correlations for butterfly stimuli (r = −0.38, p < 0.01). However, no significant association was observed between reaction time and face stimuli (r = 0.19).
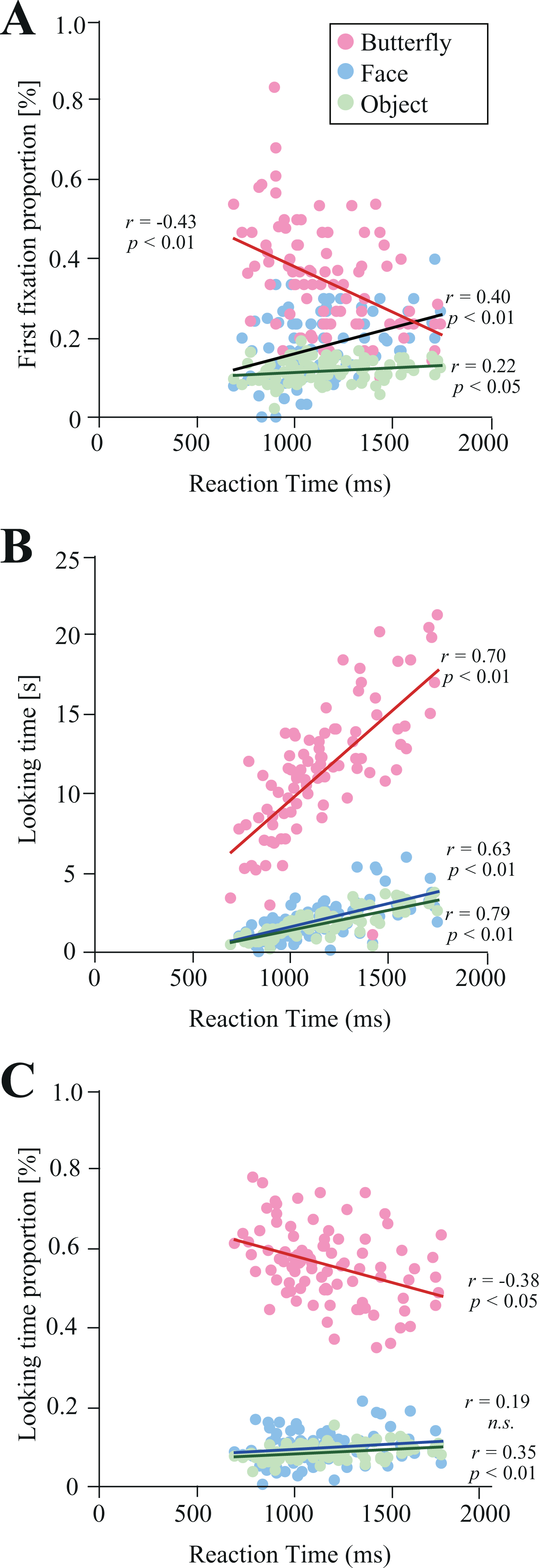
The association between reaction time and gaze behavior (N = 90). Associations between reaction time and first fixation proportion (A), reaction time and looking time (B), and reaction time and looking time proportion (C) for butterfly (pink), face (aqua), and object (green) stimuli.
Discussion
The present study aimed to elucidate developmental changes in visual attention to target-unrelated faces versus target objects during childhood by measuring reaction time, accuracy, and eye movement parameters. Overall, both reaction time and accuracy were affected by the presence of the target-unrelated face, but the interference effect was observed consistently across ages. However, developmental changes were observed when considering eye movement parameters, such as first fixation proportion. These results suggest that initial attention was significantly directed towards the target-unrelated face compared to the target butterfly before the age of 6.9 years. Furthermore, prior to the age of 12.8 years, the first fixation towards target-unrelated face stimuli was significantly more frequent compared to that for object stimuli. In contrast, the looking time proportion for face stimuli was significantly higher than that for objects across all ages. This result suggests that no developmental changes influence the search period during a trial, except for the initial orienting phase indexed by the first fixation.
In line with the findings of Langton and colleagues (2008), we observed significantly prolonged reaction times when the target-unrelated face was present compared to when the target-unrelated face was absent, across all ages. These findings suggest that, although manual responses can adequately capture the interference effect of faces, they are not sensitive enough to capture subtler developmental changes, as we did not observe a significant effect of age. However, the use of an eye-tracking system allowed us to observe subtle developmental changes in the first fixation towards target-unrelated faces and objects, differentiating the initial orienting from the later fixation phases.
We also observed that the looking time proportion for face stimuli was significantly longer than that for objects across all ages, consistent with previous studies that utilized an identical experimental paradigm (Hirai, et al., 2016a, 2016b). Therefore, the looking time proportion reflected the attentional profile at the later period of searching and did not differ across groups. In particular, looking time reflected attentional disengagement resulting from a combination of visual attention captured by the target-unrelated face and explicit attentional control toward the butterfly. This was consistent with the findings of previous infant behavioral studies, which have reported a strong preference for faces revealed by looking time measures (Di Giorgio, Turati, Altoe, & Simion, 2012; Frank et al., 2009; Gliga et al., 2009; Gluckman & Johnson, 2013).
However, contrary to our initial prediction, we observed developmental changes in the orientation of attention towards faces during the initial search period, which was defined based on the first fixation proportion. This result is somewhat inconsistent with previous theoretical and experimental implications, as even newborn human infants direct their attention towards face-like stimuli (Johnson, 2005; Johnson et al., 1991) as rapidly as adults (Tomalski et al., 2009). Using a naturalistic scene with embedded human faces, an eye-tracking study demonstrated that orienting attention toward faces improved after the first year (Amso et al., 2014). Moreover, regarding the spatial location processing of faces, the inferior parietal region activation in children between 10 and 12 years of age was comparable to that recorded in adults (Passarotti et al., 2003). It is possible that, unlike experimental paradigms in previous infant studies, the current visual search task required top-down explicit attentional control (directing attention toward the butterfly) compared to free-viewing paradigms, which do not require explicit top-down attention.
Several behavioral studies have shown that the orientation of visual attention reaches mature levels in the early stages of development (Donnelly et al., 2007; Hommel, Li, & Li, 2004; Rueda et al., 2004). For instance, performance on a visual orienting task has been reported to not significantly differ across participants aged 6 years and older, suggesting that the attentional orienting network matures by that stage of development (Rueda et al., 2004), and that selective attention on simple search tasks matures around the age of 6 years (Hommel et al., 2004). Using a visual search task, Donnelly et al. (2007) reported no significant difference in search speed between adults and 9- to 10-year-old children. Furthermore, recent neuroimaging evidence suggested that the neural networks involved in the orienting of attention undergo changes between 4 and 7 years of age, and that selective attention is significantly associated with functional connectivity between the inferior parietal sulcus and frontal eye field regions after controlling for age (Rohr et al., 2016). In light of these behavioral and neuroimaging studies, our finding that top-down attentional control of visual orientation away from the target-unrelated face reached mature levels by the age of 6.9 years may reflect the development of visual selective attention.
We also observed that, before the age of 12.8 years, the first fixation was directed towards the target-unrelated face more often than towards an object—a somewhat unexpected finding. Although we did not directly measure individual abilities for configural face processing in the current study, developmental changes in the configural processing of faces may also occur during childhood. Around the age of 11, distinct patterns of activity are clearly observed in the fusiform face area (Scherf et al., 2007), and the shape of the waveform of the face-selective ERP component (N170) resembles that of adults (Itier & Taylor, 2004; Scherf et al., 2007; Taylor et al., 2004). As we have shown in a similar experimental paradigm that configural face information is critical for attracting visual attention (Hirai et al., 2016a, 2016b), developing configural face processing, particularly in response to upright faces, can be responsible for the initial orienting of attention towards target-unrelated faces.
As predicted, we observed a significant positive correlation between reaction time and first fixation proportion for both faces and objects, while a significant negative correlation was observed for butterfly stimuli. These results suggest that children whose attention is captured by the target-unrelated face during the initial search period exhibit prolonged reaction times, while children whose attention is captured by the target exhibit shortened reaction times. Regarding later periods, we observed significant negative associations between reaction time and looking time proportion for both butterfly and object stimuli. Therefore, measures generated by different effectors such as eyes and hands appear to be tightly linked, and specific eye movement patterns geared to orienting attention in the initial search period can correlate with manual responses. Our current technique may allow us to explore the developmental mechanisms underlying the orienting of attention towards task-unrelated faces in both typically and atypically developing children.
Several limitations should be acknowledged. In our current study, we have traced developmental changes in both manual responses and eye movement measures. However, interindividual variability was observed in both manual responses and eye movements, possibly influencing the tracing of the developmental trajectory. At present, we are unable to identify the factors that can potentially affect the interindividual variability. However, a body of literature on individual differences in orienting emphasizes underlying interindividual variations of the attention network white matter fiber tracts (Niogi, Mukherjee, Ghajar, & McCandliss, 2010). Further studies are needed to identify the reasons for the observed interindividual variance. A related limitation of the current study is that, although we measured both manual responses and eye movements during the task and revealed fine developmental changes of visual attention orienting towards target-unrelated faces, the neural mechanisms underlying such developmental changes remain unclear. Though the theoretical framework proposed by Johnson (2005), suggests that subcortical regions may be responsible for this preferential orientation towards faces, further studies are needed to confirm this hypothesis and elucidate the potential changes that occur in visual orientation patterns across development. Furthermore, previous studies support the occurrence of changes in the cortical representation of face processing during childhood. However, the direct relationship between the mechanisms underlying the orienting of attention towards faces and the configural processing of faces remains to be elucidated. Furthermore, in general, the relationship between developmental changes that occur in the various dimensions of face processing during childhood should be further investigated. Finally, one might argue that the looking time measure might not directly reflect the searching process; for instance, one might search first and then attend to an item for a longer time. As we did not look into the relationship between the detailed eye movement patterns and the manual responses for each trial for each participant, further studies are needed to reveal the precise relationship between different types of eye movements and manual responses.
It is of paramount importance to expand the current research towards atypical development. We previously employed the current experimental paradigm to reveal atypical looking time, but normal first fixation towards target-unrelated faces, in developmental disorders (Hirai, et al., 2016a, 2016b). This line of investigation could prove useful for characterizing the relative intermingling of top-down control and bottom-up attention induced by target-unrelated faces. Several studies have emphasized that not only atypical face processing (Rutishauser et al., 2013; Webb, Dawson, Bernier, & Panagiotides, 2006), but also atypical orientation towards faces (Dalrymple et al., 2018), are observed in ASD. However, the relative strength of top-down control and bottom-up attention elicited by task-unrelated faces in ASD remains unknown. Therefore, it would be beneficial to capitalize on the current experimental settings to clarify open issues in ASD and other developmental disorders.
Conclusions
A consistent body of research has highlighted the preference for orienting towards face and face-like stimuli in infancy as well as adulthood. However, relatively little is known regarding the mechanisms underlying orienting behaviors towards faces in childhood due to the lack of developmentally appropriate measures. In order to elucidate the developmental trajectory for orienting attention towards face stimuli during childhood, we utilized a visual search paradigm designed to measure both manual responses and eye movements during a single task. Although no developmental changes were observed with regard to the face interference effect for manual responses, significant changes were noted with regard to first fixation proportion—but not looking time proportion—for both face and target stimuli. These findings suggest that the mechanisms underlying the orienting of attention towards faces change during childhood, and such changes can be observed during the initial search period using a measure of first fixation. The procedure, experimental paradigm, and findings described in the present study further enhance the current understanding of the mechanisms underlying the preference for orienting towards faces in both typically and atypically developing populations.
Footnotes
Acknowledgments
We thank all the children, as well as their caregivers, for their participation.
Funding
The authors declared receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Japan Society for the Promotion of Science (JSPS KAKENHI Grant Number 23830127), a grant-in-aid for Scientific Research on Innovative Areas (15H01585) for Constructive Developmental Science from the Ministry of Education, Culture, Sports, Science and Technology, a grant from the Daiko Foundation to M.H., and a grant from JSPS KAKENHI (grant numbers 23119733 and 21119529) to M.N.