
Abstract
Mice as model organisms play a vital role in translational research for developmental biology by aiding our understanding of embryonic and placental evolution in health and various disease states. The close similarity between mouse and human placentas is a great asset in reproductive translational research, making the mouse one of the more prominent model organisms in this field. Mice are routinely used to investigate mechanisms of embryonic and placental development, often through experimental manipulations involving engineered or spontaneous genetic mutations and/or xenobiotic treatments. A crucial aspect of fully appreciating the mouse as a model organism for placental research is understanding the anatomy and physiology of the major embryonic (chorion, labyrinth, junctional zone, and yolk sac) and maternal (decidua basalis and metrial gland) placenta components, including knowledge of the specific cell types active during each gestational stage. This paper discusses histochemical and immunohistochemical (IHC) methods that highlight specific tissue layers and cell types present during normal development of the mouse placenta and maternal metrial gland. Altered numbers and/or locations of these common cell types have been demonstrated in embryonic lethal phenotypes, indicating that placental assessments must examine not only the tissue organization but also affected cell populations when determining the cause of early pregnancy loss.
Introduction
Much of the existing reproductive and developmental biology research involving the mouse as a translational model for understanding human health and disease focuses on genetic alterations (engineered or spontaneous) or experimental (infectious or xenobiotic) treatments involving embryos. In comparison, the transient, albeit critical, placenta has been far less studied. The placenta proper involves both embryonic components (beginning with the trophectoderm of the blastocyst wall and advancing to the yolk sac of the primitive placenta before culminating in the chorion, labyrinth, and junctional zone in the definitive placenta) and maternal derivatives (decidua basalis and metrial gland) (Figures 1 and 2B).
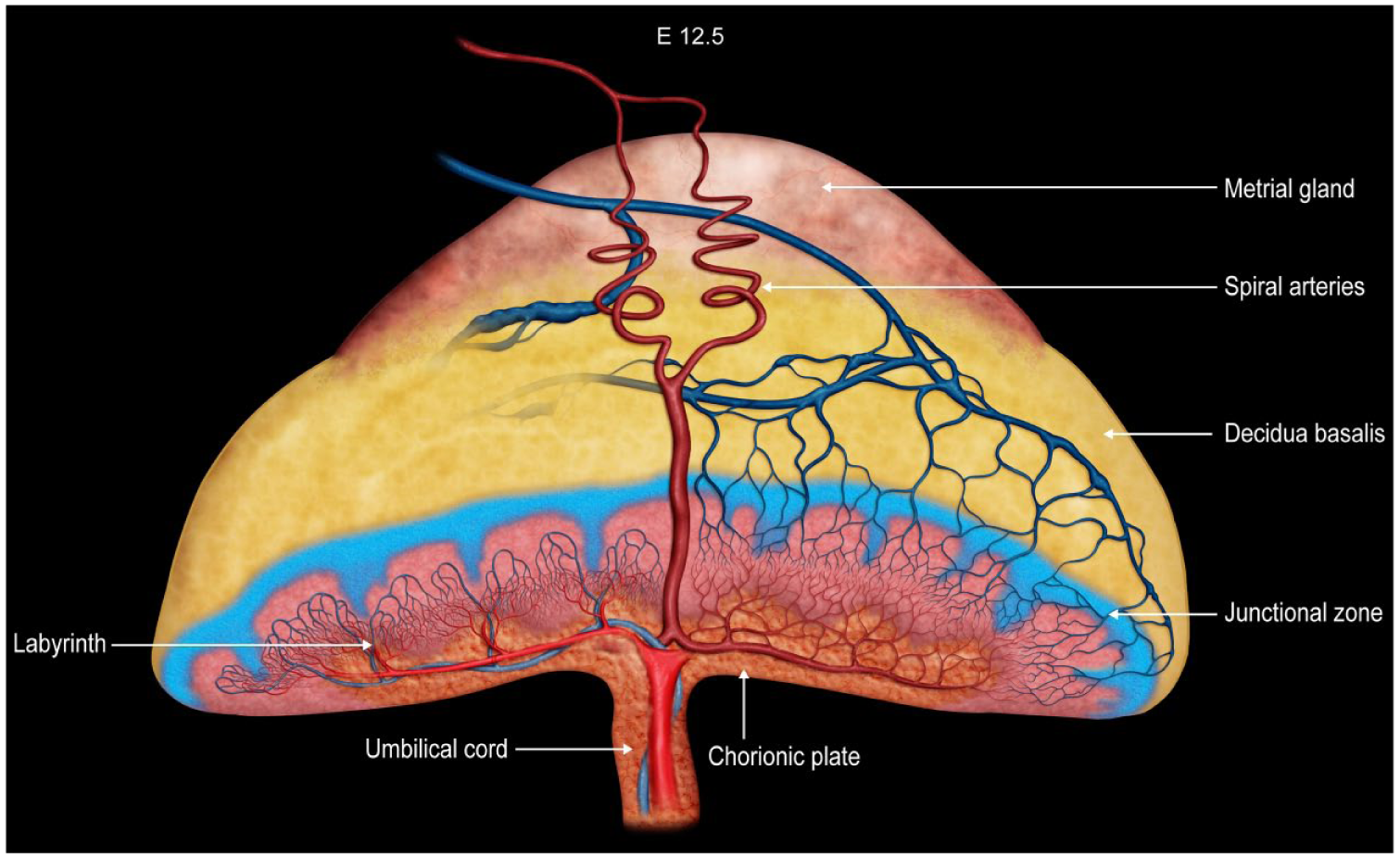
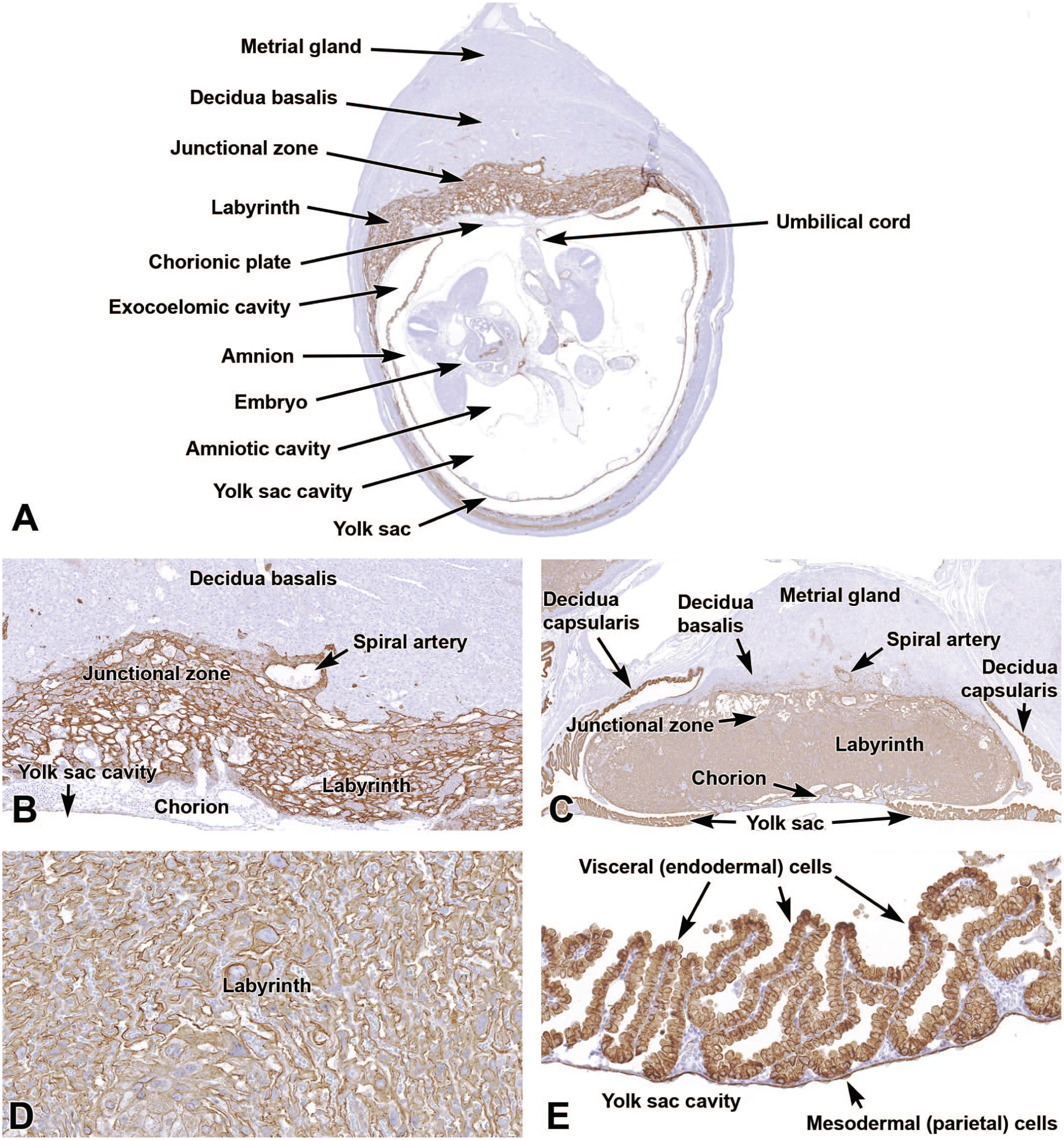
Diagrammatic representation of placental zonation at E12.5, at which point the “definitive” (mature) placenta is fully functional in mice. The color coding indicates different placental structures. The metrial gland is the pale red-orange cap that sits atop the yellow decidua basalis; both these regions are of maternal origin. The junctional zone is depicted as the blue layer with columnar projections that enter the underlying, highly vascularized reddish-pink labyrinth. The chorionic plate is the orange basal layer to which the umbilical cord is attached. Two maternal spiral arteries (red, coiled vessels) course through the metrial gland to reach the chorionic plate before branching widely throughout the labyrinth. A corresponding venous network in the labyrinth delivers blood into maternal veins (blue, straight vessels) that extend from the junctional zone through the decidua basalis and metrial gland. The labyrinth is arranged as a series of interspersed, parallel, small-caliber blood vessels (embryonic capillaries and maternal sinusoids) exhibiting countercurrent blood flow to facilitate nutrient and gas exchange. Graphic created by David Sabio. Reprinted with an adapted legend from Elmore et al, “Histology atlas of the developing mouse placenta,” Toxicol Pathol 50(1):60-117, 202249, by permission of Sage.
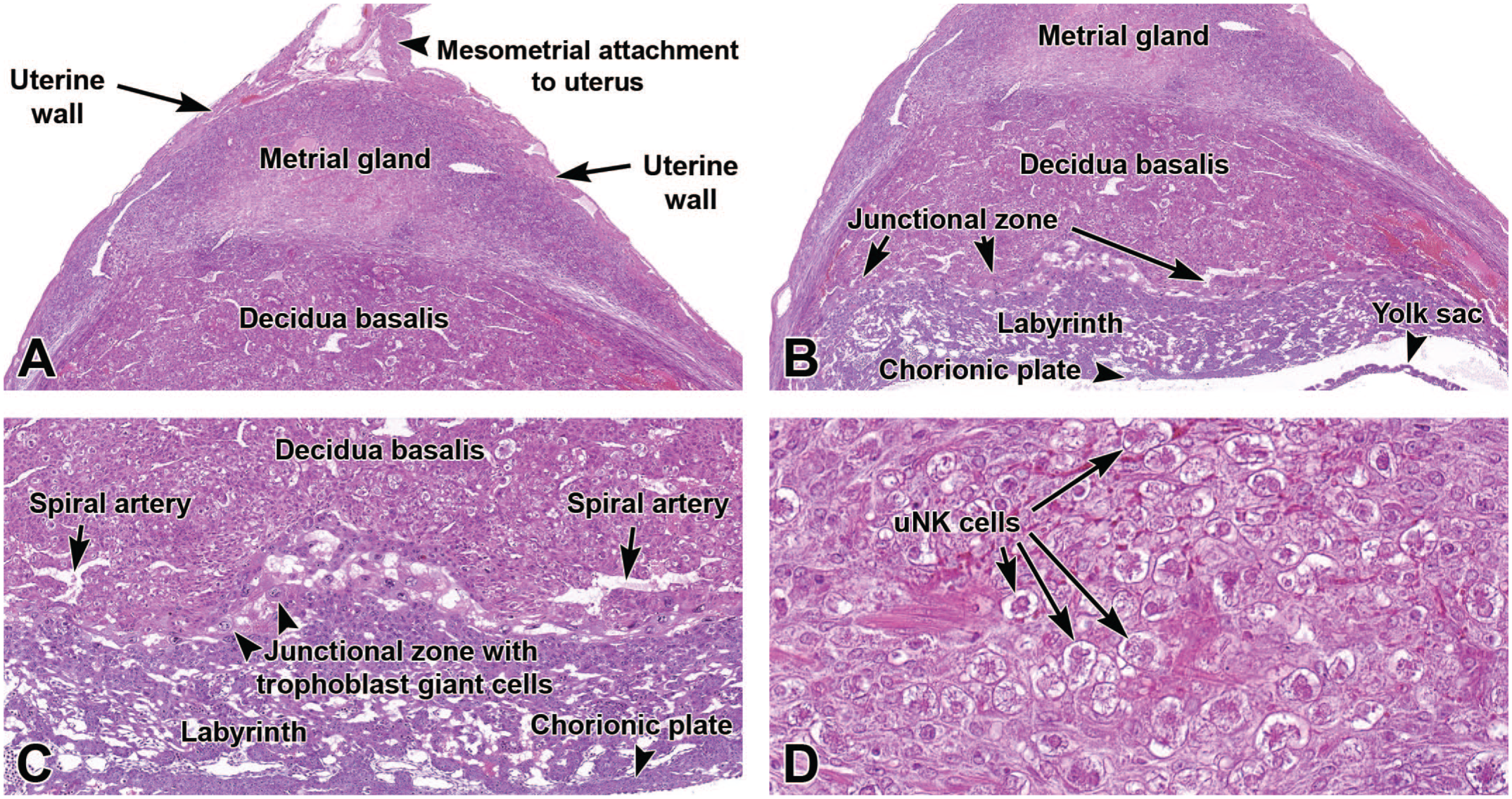
Representative images from the primordial placenta at E9.5 illustrating the location of the major placental layers (metrial gland, decidua basalis, junctional zone, labyrinth, and chorionic plate). Panel A shows the maternal-derived decidua basalis and metrial gland, as well as the uterus walls and the region of mesometrial (suspensory ligament) attachment. Panel B shows the orientation of the decidua basalis with respect to the underlying embryo-derived junctional zone, labyrinth, chorionic plate, and yolk sac. Panel C is a higher magnification view to show the spiral arteries of the decidua basalis, the trophoblast giant cells of the junctional zone, the high vascularity of the labyrinth, and the dense chorionic plate. Panel D illustrates the high number of uterine natural killer (uNK) cells with distinct cytoplasmic granules within the metrial gland. H&E. Original objective magnification: 4x (Panel A), 4x (Panel B), 8x (Panel C), or 39x (Panel D).
The placenta is initiated on approximately embryonic day (E) 4.5 by implantation of the blastocyst (multicellular embryo) in the uterine wall. Placental organization and cell populations continue to expand and evolve through the end of gestation (E18.5 to E19), when degradation occurs in preparation for birth. Throughout gestation, the placenta follows a stereotypical and strict pattern of growth-related and time-dependent events with minor disruptions having drastic effects on embryo development and survival. Interruptions to the normal functioning of any aspect of the placenta can result in embryonic abnormalities such as intrauterine growth restriction, malformations, or death. 63 Failure to properly understand the full scope of placental development and the impact on resulting embryonic phenotypes can lead to misinterpretations when assessing embryonic lethality, malformations (structural defects caused by intrinsic cell or tissue damage [i.e., abnormal development]), and deformations (structural defects produced by an extrinsic [physical] force acting on developmentally normal cells and tissues).12,63,95
Analysis of the placenta should include macroscopic and microscopic examinations of the easily distinguishable embryonic and maternal areas.95,155,156 The principal embryonic elements in the definitive placenta are arranged as a multilayered disk sandwiched between the maternal placenta (i.e., the reorganized inner uterine wall) and the thin membranes that envelop the embryo (amnion [nearest the embryo] and yolk sac). The embryonic layers of the definitive placenta are, from deep (nearest the embryo) to superficial, the chorionic plate, labyrinth, and junctional zone, which together engage in countercurrent exchange of nutrients and gases between parallel vessels of the maternal and embryonic circulations and also maintain a physical and immunological barrier between the embryo and maternal systems. The maternal placenta forms a conical cap of decidua (including the decidua basalis, a thin outer rim of decidua capsularis, and the metrial gland) that is stacked on top of the embryo-derived placental disk. The decidua broadly refers to the entire modified uterine endometrium during pregnancy, which produces growth factors and cytokines to support implantation, facilitate embryonic growth, regulate trophoblast invasion, and modulate immune reactions in the placenta. 117 The decidua basalis, a specialized part of the maternal decidua (Figures 1 and 2), works in conjunction with the embryonic placenta to provide nutrients, gas exchange, and waste removal. The decidua capsularis is a thin layer of maternal uterine tissue that is sandwiched between the decidua basalis and the uterine wall proper. The metrial gland, a specialized part of the maternal decidua, develops in the rodent myometrium during pregnancy at the mesometrial side of the uterus and functions to support decidualization, immune modulation, and placental vascularization.
Effective evaluation of placental structure often benefits from the assessment of specific embryonic and/or maternal cell populations. Such assessments may be undertaken using routine histochemical stains or immunohistochemical (IHC) methods.89,118 This paper provides detailed information on a select battery of common histochemical stains and antibodies that highlights important placental cell types for particular subanatomic locations over a range of specific time points during development. High-resolution, well-annotated color images from whole-slide scans of mouse placenta and metrial gland sections taken through the mid-axial plane illustrate the laminar organization and cell-specific distribution throughout gestation. A more comprehensive anatomic review of mouse placenta and metrial gland development over time in conventional hematoxylin and eosin (H&E)-stained mid-axial sections can be found in the companion paper Histology Atlas of the Developing Mouse Placenta. 49
Materials and Methods
Animals
Young adult, male and nulliparous female CD-1 IGS/Crl:CD1(ICR) mice (Charles River Laboratories, Raleigh, NC) were procured and quarantined for one week, after which they were paired overnight starting at the beginning of the 12-hour dark cycle. This mouse stock was chosen based on ready availability, large litter size, and the frequent use of this genetic background for developmental toxicity testing. Differences in placental/metrial gland developmental events have been distinguished among various inbred mouse strains. 10 This outbred stock follows the average (i.e., “normal”) course of mouse placental and metrial gland development.
Dams were group-housed (2-3/cage) in Green Line IVC Sealsafe PLUS Mouse cages (Tecniplast; West Chester, PA) on autoclaved Sani-Chip hardwood bedding (PJ Murphy Forest Products Corp, Montville, NJ). Animals received pelleted rodent chow (NIH-31; Envigo Corp, Indianapolis, IN) and reverse osmosis/deionized water ad libitum. Environmental conditions were held at 22°C ± 2°C and 45% ± 10% relative humidity. A constant light cycle (12-hour light and 12-hour dark) was maintained before and after breeding. Mice were supplied with Enviro-dri (Eco-bedding, Shepherd Specialty Papers, Morrisville, NC) and cotton fiber pads (nestlets) for nesting purposes.
All experimental procedures performed in this study were approved in advance by the US National Institute of Environmental Health Sciences (NIEHS) Institutional Animal Care and Use Committee (IACUC). Mice were housed in an Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC)-approved animal facility. Health surveillance reports and in-house quality assurance data confirmed that mice in this colony were free of known pathogens.
Terminology
The “embryo” classification scheme allows for standardized staging of embryos, a vital element in developmental toxicity testing since comparisons to discern treatment-related effects need to compare stage-matched individuals. By this system, an “embryo” is the in utero entity in which all organ primordia are initially forming (in humans, weeks 1-8 following fertilization), while a “fetus” is the in utero organism in which all primordia have formed and are now engaged in global expansion and remodeling (in humans, weeks 9-37 following fertilization). Since the mouse has a much shorter gestation length (19-20 days), the designation of “embryo” versus “fetus” is less important in this species. For this reason, the term “embryo” is frequently used to define all prenatal stages of murine development, with the stage of development designated by the gestational age (with conception, designated as E0, occurring at approximately 0.5 days after mating). 74 For comparison, using the human staging convention, in mice the first two trimesters of development (“embryo”) extend from E0 to E14.5 while the last trimester (“fetus”) encompasses E15.0 to approximately postnatal day (PND) 10. Thus, a substantial portion of the “third trimester” in mice actually occurs after birth. 13
Embryonic Staging
In conventional mating of mice, conception takes place during an overnight (12-hour) mating. The day following successful mating (ie, when a copulation plug can be seen) may be variously designated as E0, E0.5, or E1. The convention employed for this study is that the morning after mating is designated E0.5 (under the assumption that copulation occurred soon after the breeding pair was placed in the same receptacle.14,105 An alternative approach to embryonic staging (e.g., the Theiler system) is to base assignments on the presence of discrete morphological landmarks. 16 Given the importance of comparing stage-matched embryos, it is imperative that researchers indicate the dating system used for their experiment as slight differences (hours) in developmental age can result in substantial anatomic divergence among embryos. 105
Time Points to Evaluate
The tissue origin of the placenta is the trophectoderm that comprises the blastocyst wall, which first forms in mice at approximately E3.0 to E3.5. Without implantation in the uterus and without contact with the maternal component, this embryonic precursor tissue cannot differentiate into placental elements. E5.5 is the earliest time point for systematic evaluation of the placenta because mouse embryos typically have undergone implantation by this time (usually somewhere between E4.5-E5.0) and the egg cylinder wall that is the primordium of the placenta is well-formed by this stage (Figure 3). Just prior to birth (i.e., E18.5 or “near term”) is generally the latest time point for placental evaluation because tissue degradation accelerates thereafter in preparation for birth (~E19.0 to E19.5 in mice). All time points indicated in this article are approximations as modest variations in placental morphology are present both across mouse stocks and strains as well as among individual embryos for a particular litter.
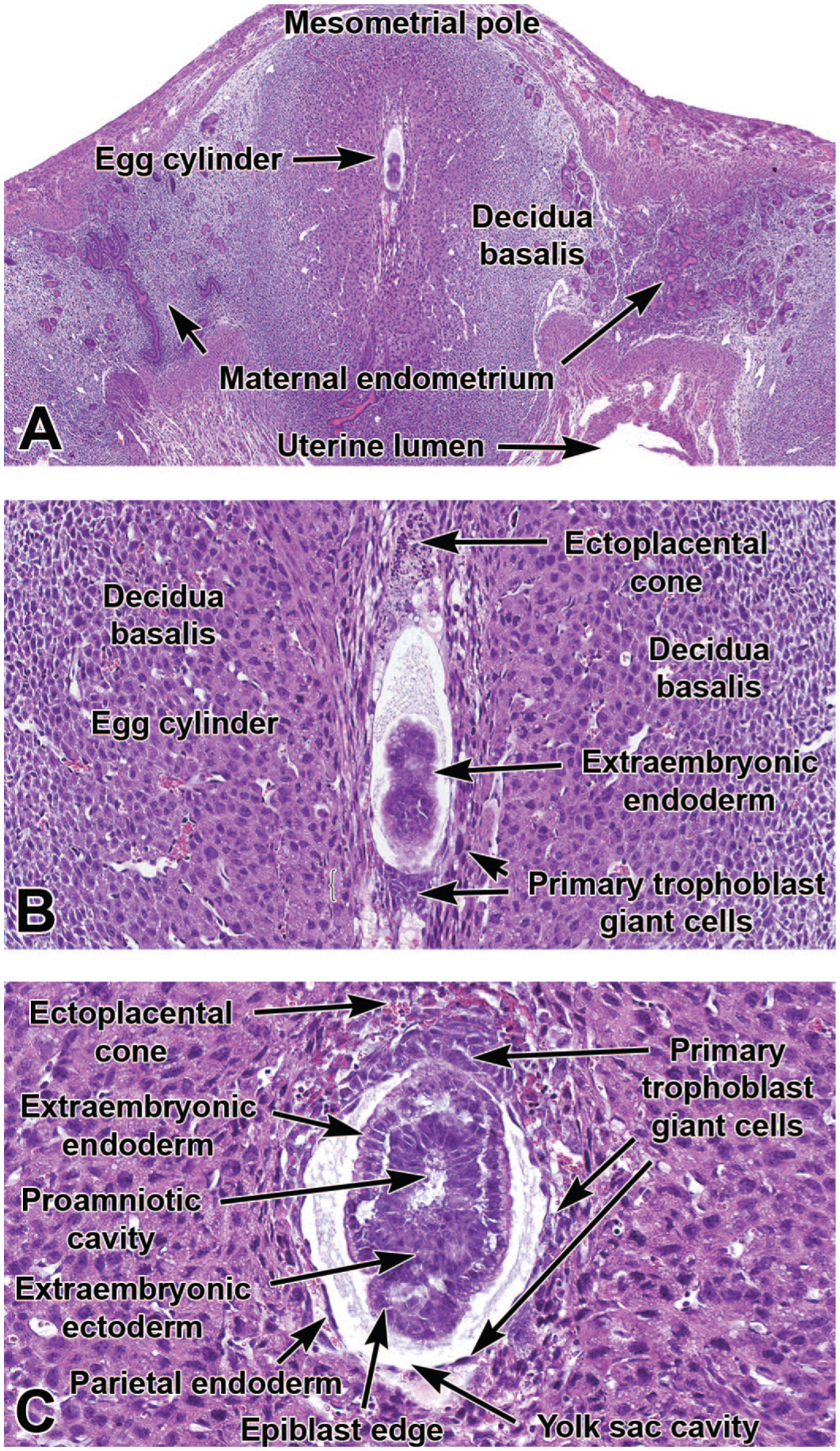
Representative images of the primordial placenta at E5.5 (shortly after implantation [Panels A and B]) and E6.5 (during gastrulation [Panel C]). The ectoplacental cone is a transient trophectoderm-derived structure that develops at the mesometrial pole of the elongated egg cylinder as the initial placental element connecting the embryo to the dam. Panel C shows the close relationship between the embryo-derived ectoplacental cone, various endoderm elements that contribute to the primitive placenta, and the epiblast (nascent embryo). H&E. Original objective magnification: 3.2x (Panel A), 16x (Panel B), and 20x (Panel C).
Tissue Collection, Handling, and Processing
Collection of placentas and metrial glands (with corresponding embryos) was carried out on the mornings of selected days between E5.5 and E18.5. Pregnant mice were euthanized by carbon dioxide inhalation according to the NIEHS standard operating procedure for euthanasia of rodents. Using a dissecting stereomicroscope (Leica MZ16; Leica, Vista, CA), the placentas with metrial glands were isolated and briefly immersed in ice-cold 0.1M phosphate-buffered saline (1X PBS, pH 7.4) to remove extraneous constituents (amniotic fluid and blood). The isolated placentas with metrial glands were then fixed by immersion at room temperature (RT) in a commercially procured solution of methanol-free 4% formaldehyde, pH 7.4 (colloquially termed 4% paraformaldehyde [PFA]; Affymetrix, Santa Clara, CA). The fixation time was 48 hours. Following fixation, placentas with metrial glands were processed routinely into paraffin. For E5.5 to E9.5 time points, conceptuses (i.e., the embryo and placenta as a combined unit) or isolated placentas were embedded while intact to avoid traumatizing the fragile placental (and embryonic) tissues. For later time points (≥E10.5), isolated placentas with attached metrial glands were placed on a hard surface, flat (chorionic) side down, and trimmed with a sharp razor blade near the mid-axial plane to produce two asymmetric halves; the larger piece—comprising approximately two-thirds of the placental mass—was embedded with the cut surface oriented “down” in the cassette to permit acquisition of full-thickness placental and metrial gland cross sections. Serial 5-μm-thick sections were acquired and placed on AutoFrost IHC (hydrophilic) clipped-corner glass microscope slides (Cancer Diagnostics, Inc., Durham, NC). Every fifth slide was stained routinely with H&E to permit histopathologic evaluation and whole-slide imaging (WSI, performed as described below). Selected serial sections were then processed by either histochemistry or indirect immunohistochemistry (IHC) to detect cell type-specific biomarkers in the placenta and metrial gland.
Briefly, sections were deparaffinized in xylene and rehydrated through a graded series of ethanol to 1x TBS wash buffer. Endogenous peroxidase activity was blocked for 15 minutes at RT with 3% H2O2. Following a rinse in 1X wash buffer, epitope retrieval was conducted by high heat and pressure in a Decloaker (Biocare Medical, Concord, CA) for 15 minutes at 110°C with either citrate buffer (pH 6.0) or EDTA buffer (pH 8.5) (Biocare Medical). After blocking with either 10% normal serum and avidin/biotin (Vector Laboratories) or Rodent Block (Biocare Medical), an appropriate primary antibody (Supplemental Table 1) or an isotype-matched negative control antibody was applied for either 30 minutes or 1 hour at RT. The primary antibody was detected with a biotinylated secondary antibody followed by Vectastain Elite ABC-HRP Reagent, Peroxidase, R.T.U. (Vector) or with Rabbit-on-Rodent HRP-Polymer (Biocare Medical). Sections for all antibodies were incubated with 3,3’-diaminobenzidine (DAB) chromogen (DakoCytomation, Carpenteria, CA) for 6 minutes at RT to visualize bound antigen-antibody complexes. Sections were counterstained with Modified Harris hematoxylin (Epredia/Fisher Scientific, Kalamazoo, MI) for 15 seconds at RT, placed in 1X PBS to blue, dehydrated through a series of graded alcohols, and coverslipped with Permount medium (Surgipath, Richmond, IL).
Whole-Slide Scanning
Bright-field whole-slide scanning was completed for all stained slides with a ScanScope AT2 instrument (Leica). Regions of interest were digitally captured as screenshots using ImageScope software (v12.4.3.5008; Leica). If an image required rotation, the selected region of interest was captured using the extraction feature in ImageScope. Global white balance correction and image resizing were completed, when warranted, using Adobe Photoshop (v2014.0.0 or later; Adobe, San Jose, CA). Image resolution was set at 300 dpi to fit the publisher’s requirements for photomicrographic illustrations.
Morphological Evaluation
Placental anatomy and background pathology were evaluated macroscopically in unfixed specimens at necropsy using a dissecting stereomicroscope. Subsequently, normal placental and metrial gland structures were assessed in tissue sections using a bright-field microscope and/or whole-slide scanned images. Representative macroscopic and microscopic images were annotated with Adobe Photoshop CC 2020 to identify salient structural features of the evolving placenta.
Placenta Collection, Processing, and Evaluation Methods
Selection of Control Specimens
Selection of appropriate control specimens (especially for time points before E15.0) is defined in one of two fashions. The preferred means is to identify “developmental stage-matched” control mice using key macroscopic features of the developing embryo rather than to choose “age-matched” controls based on the gestational day at which the animal is collected. This strategy is essential because the difference in developmental stage between the oldest and youngest embryos in a mouse litter of a given embryonic age varies from 10 to 24 hours;75,87,125,150 exposure to a toxicant may amplify the apparent difference between developmental stage and embryonic age by causing delays in the rate at which treated embryos reach particular developmental milestones. 13 This factor is especially critical if the pathology assessment will include acquisition of quantitative data (e.g., morphometric or stereological measurements). As an obvious best practice, control embryos (wild-type littermates for genetic experiments, animals from unexposed or vehicle-exposed dams for xenobiotic experiments) should share the same genetic background as the experimental animals because the average developmental stage for one mouse strain may vary by as much as 0.5 days from that of other mouse strains of the same developmental age. 150 The genotypes of the embryos are typically determined via limb snips (E11.5-E13.5) or tail snips (E14.0-E18.5); yolk sac or amniotic sac (up to E11.5) also may be used as a specimen for genotyping, but these sites should not be sampled if the placenta is the main tissue of interest. Normal developmental variation of embryos and their placentas within a litter should still be considered during histopathological evaluation.
Recommended Procedure for Placental Tissue Collection
The procedure used to collect placenta and metrial gland samples depends on the developmental stage. When these tissues are to be evaluated early in gestation (≤E9.5), the conceptuses are best collected in situ (left undisturbed within the intact uterine horn). This approach is especially effective due to the small size of the conceptus, where any manipulation of the embryo and placenta during removal can easily traumatize the delicate tissues. With this approach, the metrial gland and placenta will be anchored at the mesometrial side of the uterine wall (i.e., closest to the location where maternal blood vessels carried in the mesometrium [suspensory or “broad” ligament] enter the uterine wall, with the metrial gland located nearest the wall), providing orientation during microscopic examination. Later in gestation (E10.0 to term), the placenta with attached metrial gland can be separated from the embryo using a stereomicroscope. A pair of jeweler’s forceps is useful to separate the tough uterine wall from the more friable metrial gland and placental tissue. Careful attention and handling should be given when detaching the placenta from the uterine wall so that the metrial gland is not separated from the underlying placenta; detachment can be done by careful teasing of the metrial gland away from the uterine wall or by cutting it from the wall with micro-scissors. One forceps can then be used to grasp the yolk sac while the other forceps is used to separate and remove it. Alternatively, microscissors can be used to cut the yolk sac to expose the amniotic sac, both of which can then be removed to expose the embryo. The use of small microscissors rather than forceps permits more rapid specimen collection during mid to late gestation while also limiting potential trauma associated with tissue separation using the pair of forceps.
Choice of Fixation and Processing Protocols
The choice of fixation protocol needs to strike a balance between the degree of architectural preservation and the technical skill required for a rapid and relatively atraumatic dissection. 11 In general, soft placental tissue is well preserved by immersion fixation. Common fixatives are aldehyde-based, such as neutral buffered 10% formalin (NBF, which in commercial solutions typically contains approximately 1% methanol as a stabilizer to inhibit oxidation of formaldehyde to formic acid) or 4% PFA. If electron microscopy is of interest, modified Karnovsky’s fixative (2.5% glutaraldehyde [Glut] with 2% PFA) or similar mixtures of fast-penetrating (NBF, PFA) and slow-penetrating (Glut) aldehydes are used to provide better stabilization of lipid-rich membranes and fine cell structures. Fixatives containing aldehydes, acids, and alcohols, such as Bouin’s solution or modified Davidson’s solution, have traditionally been used in developmental toxicity testing since these mixtures provide superior fixation (through better skin penetration) and hardening of soft embryonic tissues. That said, these combination fixatives are used less often in preserving placentas as they tend to lyse red blood cells, thereby obscuring features in the richly vascularized labyrinth. 11
Investigation of Placental Phenotypes
The analysis of abnormal placental phenotypes is typically guided by the initial identification of a gross structural defect in the placenta or by in utero lethality for which a structural cause cannot be defined within the embryo. The histopathological appearance of anatomic lesions in the developing placenta and/or metrial gland may provide clues regarding their cause, timing, and the most appropriate techniques that might be used to further characterize their attributes and impact. Major placental defects in mice commonly arise during organogenesis, the period of development that encompasses the initial development of the yolk sac (~E6.0), the initial linkage of the embryonic umbilical (“vitelline”) vessels to the chorionic plate (~E8.5), and the ultimate formation of the definitive placenta at E12.5. 49 Evaluations of early placentation events, such as yolk sac formation and progressive branching of the yolk sac vasculature, are typically done in situ (i.e., by evaluating implantation sites that remain in the uterus) from E5.5 to ~E9.5, while examination of events such as expansion of the embryonic labyrinth and progressive differentiation of various cell lineages throughout the placenta are generally performed later in gestation (E10.0 to term). The embryo-derived placental precursor can be distinguished in free-floating blastocysts within the oviducts starting at about E3.5. However, mouse developmental pathology analysis at this early stage usually emphasizes evaluation of the inner cell mass (ICM [i.e., the primordial embryo]) and not the trophectoderm (i.e., the nascent embryo-derived placenta that forms the blastocyst wall).
Phenotypic Evaluation of the Placenta
A complete assessment of the maternal- and embryonic-derived sections of the placenta should include both macroscopic and microscopic examinations.95,156 The maternal region is the conical cap of the metrial gland and contiguous decidua basalis, which in turn rests on top of the embryo-derived placental disk (Figures 1 and 2). Functionally, the metrial gland aids in the modification of the mouse uterine tissue at the time of implantation and in modulating interactions of embryonic and maternal placenta components throughout gestation. The metrial gland is important as it is located in the region of the uterus through which most maternal blood flow will pass upon entry into the placenta and can play a key role in immune regulation during pregnancy as well as modulate some placental and gestational disorders.3,142 Therefore, the metrial gland should always be collected and evaluated in conjunction with the placenta.
The decidua basalis encompasses the implanted blastocyst and, like the metrial gland, is formed from the modified lining of the uterine endometrium. It provides a vascularized tissue bed that connects the dam and embryo, facilitates nutrient and gas exchange, and protects the adjacent uterine muscle from excessive invasion by trophoblast cells. The decidua capsularis is a narrow shell of endometrium that lies between the decidua basalis and the outer uterine wall.
Key embryonic derivatives of the placenta include a multilayered disk comprised (from outer to inner) of the junctional zone, labyrinth, and chorionic plate (Figures 1 and 2) and two distinct thin membranes that envelop the embryo (amnion [nearest the embryo] and yolk sac). Abnormal placental findings may be identified in one or several of these discrete structures by macroscopic and/or microscopic evaluation. When opening the yolk sac to expose the embryo, a check of the umbilical vessels should be done to ensure there is blood present as an indication that nutrient exchange is occurring. This simple assessment can provide great insight when dealing with embryos and placentas that appear pale and/or small.
Upon removal from the uterus, the placental surface should be examined for abnormalities in shape, size, and/or color. In order to ensure that the metrial gland is harvested with the placenta, it is good practice to remove the placenta and associated mesometrial uterine wall as a unit. The collection process should be carried out as quickly and atraumatically as possible due to the rate at which delicate cells and vascular-rich tissues of the placenta break down. Key macroscopic findings may be photographed, typically using a stereomicroscope. If necessary, placentas may be weighed, but in our experience this endpoint is not a standard component of routine placental phenotypic screens. If weighing is performed, the placenta is typically separated from the uterine wall and umbilical cord.
For histopathologic evaluation, the usual orientation for placental and metrial gland trimming is to prepare mid-axial cross sections. Initial histopathologic assessment of placental and metrial gland phenotypes often is limited to a qualitative (present or absent) or semiquantitative (i.e., graded) evaluation of the contours and sizes of major zones and identification of obvious cellular and tissue alterations using conventional H&E-stained sections. On occasion, the distribution and number of one or more placental cell lineages may be evaluated in serial sections using histochemical or IHC methods to detect cell type-specific biomarkers. Quantitative methods to examine the size and/or number of cells (via morphometry and/or stereology) generally are done only to evaluate specific hypotheses, 111 and as such, these special methods are not addressed here.
Overview of Normal Placenta Development and Function
The placenta is a transient support organ that is necessary to sustain the growing embryo in utero. It is responsible for all nutrient, gas, and waste exchange that takes place throughout embryonic development, beginning at approximately E4.5 when implantation occurs in the uterine endometrium. 49 The placenta changes over time, with the means of embryonic support shifting as the placental structure evolves.
Shortly after implantation, the primary (choriovitelline, or “yolk sac”) placenta takes shape as trophoblasts (the primordial placental lineage) begin evolving from trophoblast stem cells 122 to produce the yolk sac and regulate endometrial decidualization. Trophoblast stem cells are derived from the trophectoderm of a blastocyst or the extraembryonic ectoderm after implantation and are the precursors of the differentiated cells of the placenta. 122 These self-renewing and multipotent stem cells can differentiate into all types of placental cells.
Trophoblast giant cells (TGCs) form at two main stages during mouse development: the primary wave shortly after implantation and a secondary wave later from the ectoplacental cone.62,135 Primary TGCs (i.e., parietal TGCs) form after the embryo implants into the uterine wall, around E4.5, and differentiate from the mural trophectoderm. These are the first cells to invade the maternal decidua and form the initial network of blood sinuses. Secondary TGCs (i.e., spiral artery-associated TGCs and labyrinth maternal sinusoidal TGCs) appear later, around E7.5 to E9.5, and persist throughout gestation. They arise from the polar trophectoderm and the ectoplacental cone to eventually line the maternal blood spaces (“sinusoids”). The canal-associated TGCs have a mixed developmental origin.
Trophoblast cells and the yolk sac engage in histiotrophic nutrition—the process where cells of the choriovitelline placenta phagocytize maternal secretions and transfer the processed nutrients to the embryo while sending waste products from the embryo to the decidual vessels. The choriovitelline placenta will sustain embryonic growth while the embryo is small, acting from the first formation of the choriovitelline placenta until the allantois, the embryo-derived precursor of the vitelline vessels (umbilical cord), contacts and fuses with the chorion at approximately E8.5. 6
The definitive (chorioallantoic) placenta is initiated by fusion of two embryonic membranes: the allantois and the chorion.1,31,136 Thereafter, the gradual expansion of the labyrinth yields dense capillary beds needed for hemotrophic nutrition, the process whereby countercurrent vascular exchange moves nutrients and wastes along gradients between parallel maternal sinusoids and embryonic capillaries. The labyrinth forms from both embryo-derived and extraembryonic tissue, as chorionic folding that penetrates the labyrinth is accompanied by invasion of the allantoic tissue into the chorionic plate. 136 Indeed, the chorionic plate is responsible for creating much of the vascular space within the labyrinth as it continues to expand1,31,35,49 while the invading allantoic tissue wears away at the chorionic plate tissue to create space for driving vascularization of the labyrinth. The invading allantoic tissue also induces the formation of labyrinth-specific trophoblast cell types known as cytotrophoblasts and syncytiotrophoblast types I and II.31,35,70,121,136 These three trophoblast layers combine to form the interhemal membrane layer (or “selectively permeable placental barrier”) that separates the maternal sinusoids from the embryonic blood vessels (Figure 4).1,31,136 The cytotrophoblast layer becomes perforated after E12.5, and the interhemal membrane thins between E12.5 and E16.5 with a reduction of about 50% in thickness during this period. 30
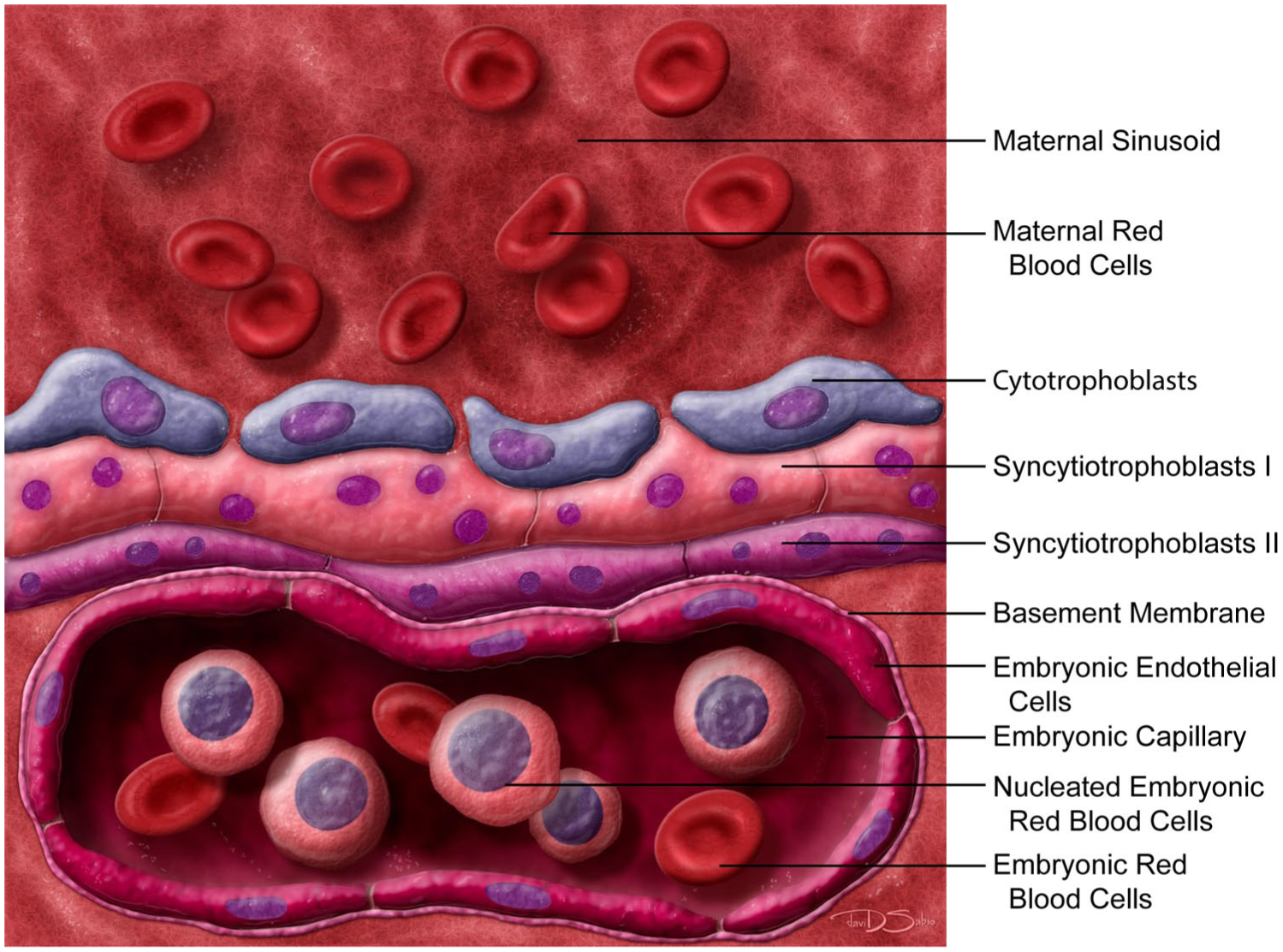
Schematic diagram of the interhemal membrane unit of the definitive mouse placenta (which is structurally mature at E12.5). In mice, a hemotrichorial barrier within the labyrinth separates the maternal and embryonic circulations (where “hemo” denotes that the potential three layers of maternal cells [maternal endothelium, connective tissue, and uterine epithelium] are missing while “trichorial” means that three embryo-derived trophoblast layers separate the maternal sinusoids from the embryonic capillaries). The three embryo-derived trophoblast layers of the membrane unit consist of a discontinuous, mononucleated cytotrophoblast layer that lines the maternal sinusoids and two continuous, multinucleated syncytiotrophoblast layers, designated as I and II, located between the cytotrophoblast and basement membranes of the embryonic capillaries. Graphic by David Sabio.
Once the labyrinth forms and becomes more vascular, the junctional zone is induced around E9.5 (Figures 2B and C). This layer separates the labyrinth from the decidua basalis and contains venous sinusoids with maternal blood that has already moved through the labyrinth and is now returning toward the maternal circulation. 31 Like the labyrinth, the junctional zone contains its own specific differentiated trophoblast cells that play key roles in placental endocrine activity by producing growth factors, hormones, and cytokines that aid the embryo throughout its remaining development. 133 A band of TGCs (Figure 2C) borders the decidua basalis (adjacent to the junctional zone), spongiotrophoblast cells line the maternal venous sinusoids in the decidua, and the glycogen-containing trophoblast cells (also known as “glycogen cells”) exist as clusters of clear vacuolated cells throughout the junctional zone. 31 The TGCs secrete hormones that regulate the development of the embryonic and maternal placental compartments.55,135 The function of the spongiotrophoblast cells is poorly understood, but they secrete a number of polypeptide hormones and may provide structural support.135,163 Glycogen cells also secrete hormones, such as prolactin, 137 and store glucose as glycogen, which is thought to provide an energy reserve for the developing embryo in late gestation. 31
The definitive placenta fully matures in terms of its multilayered structure and function by approximately E12.5. Nonetheless, both the labyrinth and junctional zone continue to undergo modest morphological changes beyond E12.5. Maximum placental volume is reached by E16.5. 30
Overview of Placental Pathology Patterns
Lesions in mouse conceptuses may lead to altered gestational outcomes at any time during gestation. Importantly, the timing of embryonic lethal events gives clues regarding when the embryo 10 and/or placenta 15 were affected during development and which embryonic organs or placental cell types might have been impacted. Studies using special histological methods to identify and characterize cell type-specific defects are an important tool for investigating such conditions. This section briefly describes common lesion patterns for key mouse placental components. 15
Lesions involving the decidua basalis may lead to embryonic death at various times during gestation, but usually before E10.5. Grossly, the placenta size is reduced. Microscopically, the decidual tissue contains fewer cells and blood vessels. The proposed defect for such phenotypes is altered decidualization (cell transformation and expansion of the uterine lining).
Lesions of the ectoplacental cone result in embryonic death between E6.5 and E8.0. Grossly and microscopically, the key feature is the diminished size of the cone. The putative mechanism of such phenotypes involves aberrant formation of the initial embryonic placenta.
Lesions of the yolk sac typically result in embryonic death between E8.5 and E10.0. Grossly and microscopically, the yolk sac has fewer and smaller blood vessels and/or hematopoietic aggregates (“blood islands”). These findings reflect altered hematopoiesis and/or abnormal placental vasculogenesis.
Lesions of the chorioallantois generally induce embryonic death between E9.5 and E11.0. Grossly and microscopically, the principal feature is the separation (nonfusion) of the allantois and the chorion. This appearance is indicative of a thwarted transition from the initial choriovitelline placenta (in which the yolk sac effectively sustains the embryo via histiotrophic nutrition) to the definitive placenta (in which chorioallantoic fusion is essential to form the umbilical cord and induce the intricate vasculature of the labyrinth needed to support hemotrophic nutrition).
Lesions of the labyrinth may cause embryonic death at many points from mid to late gestation, depending on the affected cell (trophoblast) populations. Altered initial production and expansion of trophoblasts commonly result in death between E9.0 and E11.0 while abnormal lineage-specific differentiation often incites death between E13.0 and E16.0. Grossly, trophoblast defects in the labyrinth yield an altered placental size (increased or decreased). Microscopically, the usual finding is increased (hyperplasia) or decreased (hypoplasia) numbers of one or more trophoblast lineages, typically appearing in H&E-stained sections as vascular disorganization. Examination of specific trophoblast populations using cell type-specific biomarkers is generally required to elucidate the target cell population(s) and pathogenesis.
Overview of Normal Metrial Gland Development and Function
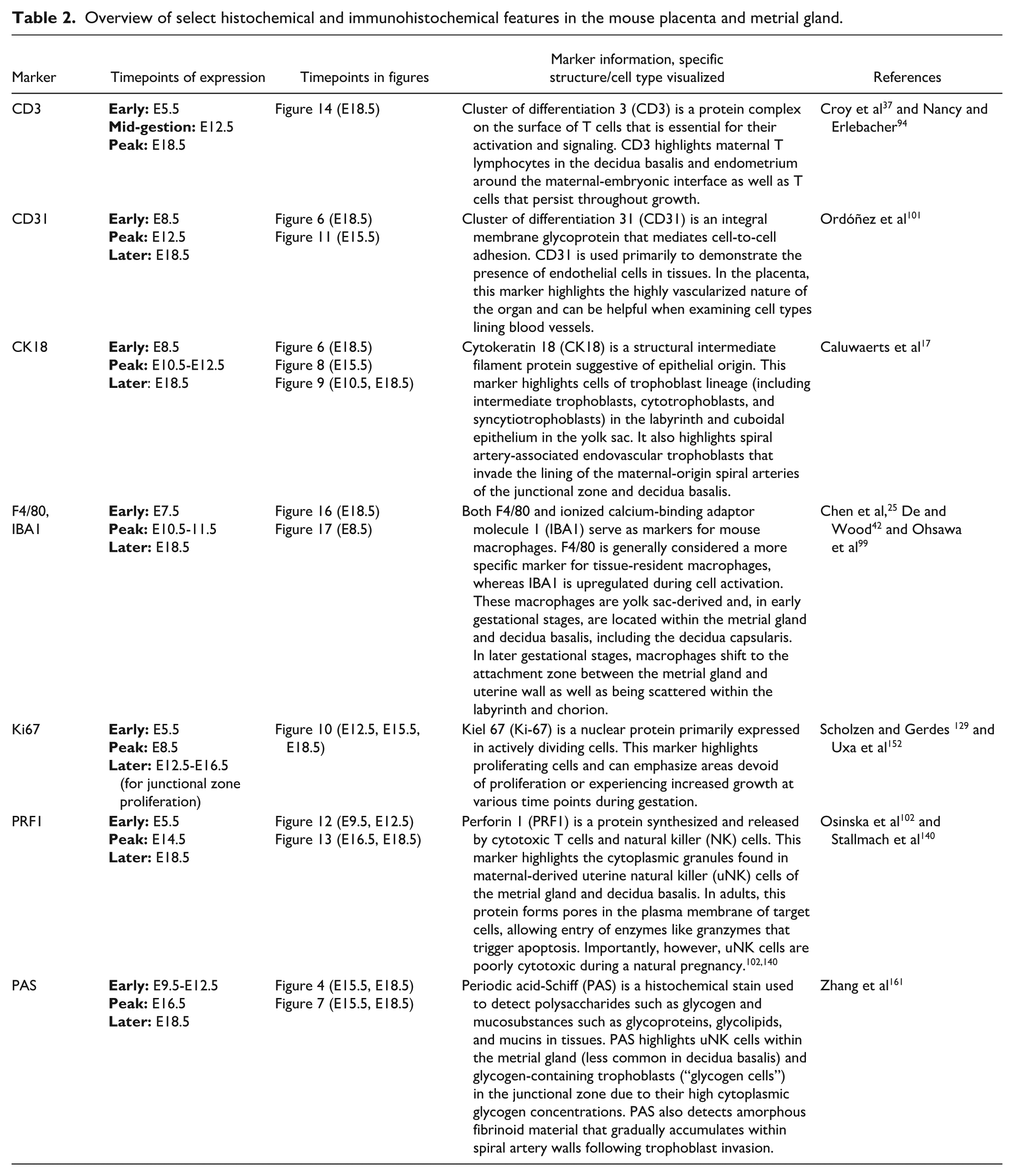
The metrial gland, also called the mesometrial lymphoid aggregate of pregnancy or the mesometrial triangle,3,36,114,143 is a maternally derived tissue that is located in the uterine wall of the pregnant mouse (Figure 2A).3,113 This tissue contains endometrial stromal cells, interstitial trophoblasts, blood vessels, fibroblasts, macrophages, and uterine natural killer (uNK) cells (Figure 2D). 113 The uNK cells (also called granulated metrial gland [GMG] cells) are large granular lymphocytes characterized by periodic acid-Schiff (PAS)-positive cytoplasmic granules that contain glycoproteins (Figure 5A-E).110,113 In mice, the metrial gland uNK cells increase in number during E13 to E15, reaching ~70% of the decidual leukocyte population at day E15. 79 By late pregnancy (E17), the uNK cells show signs of degradation due to both necrosis and apoptosis as the metrial gland regresses before parturition (compare E15.5 to E18.5 [Figure 5C]). 79 These cells are thought to regulate decidualization, downregulate maternal immune reactions to foreign antigens, and aid in decidual artery remodeling to optimize placental blood flow.49,113 Conversely, uNK cells are not thought to have a substantial cytolytic function despite containing perforin and granzymes. 107
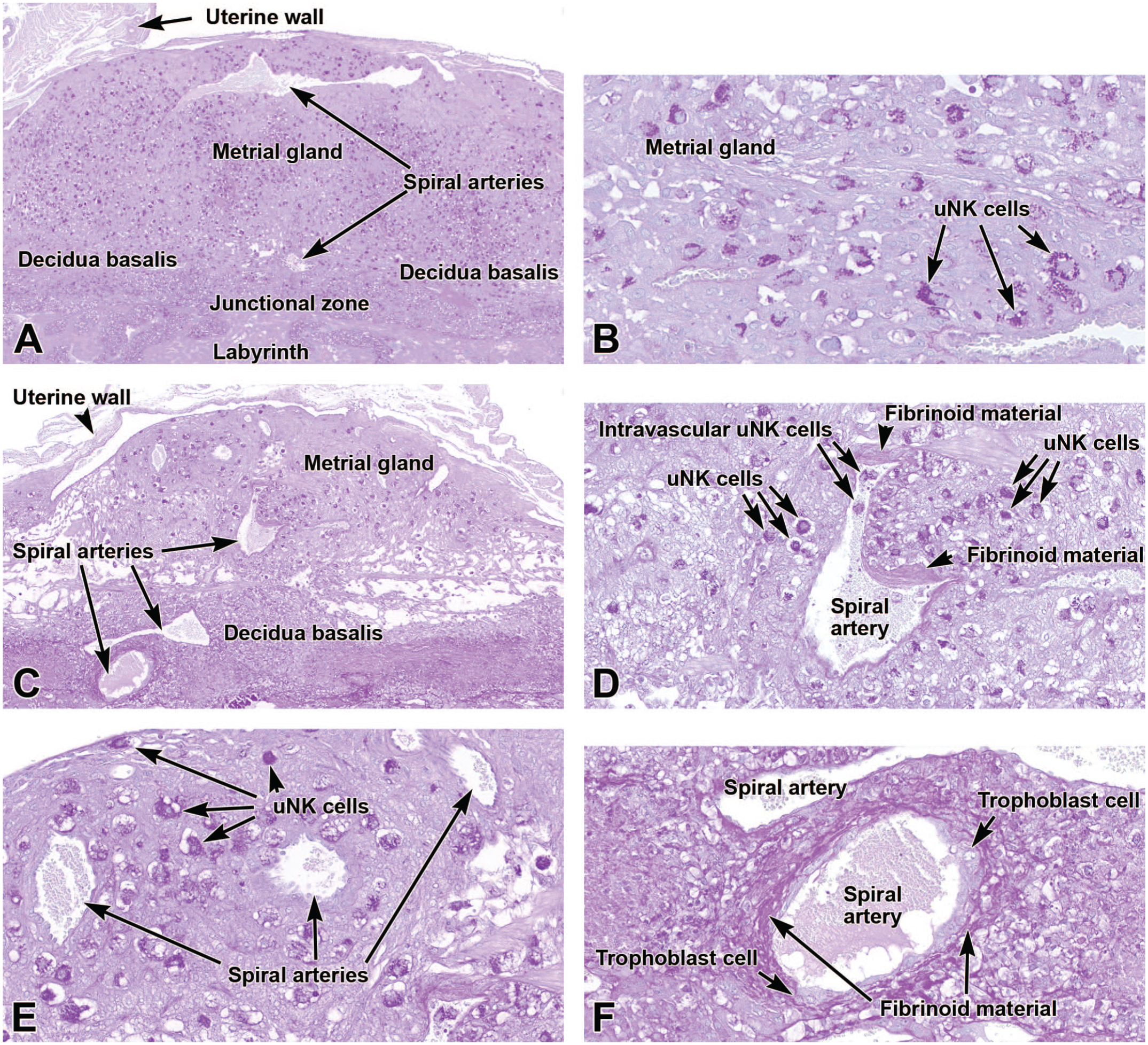
Representative images from the definitive placenta at E15.5 (Panels A-B) and E18.5 (Panels C-F) stained with periodic acid-Schiff (PAS). This histochemical method highlights the glycoprotein-laden (magenta-colored) cytoplasmic granules in uterine natural killer (uNK) cells scattered throughout the metrial gland and decidua basalis. Panels A and B (at E15.5) show the distribution of uNK cells in the metrial gland while emphasizing their relationship to spiral arteries. Panel C (at E18.5) shows that the metrial gland decreases in size while the number of uNK cells diminishes as gestation proceeds. Panel D (at E18.5) is a higher magnification view of a spiral artery depicting accumulation of acellular fibrinoid material in the vessel wall and adjacent decidua as well as uNK cells in the vessel lumen and surrounding tissue. Panel E (at E18.5) illustrates that some spiral arteries lack such fibrinoid material. In Panel F (at E18.5), invading trophoblast cells have penetrated the spiral artery walls, enlarging them and replacing their elastic tissue with PAS-positive acellular fibrinoid material. PAS. Original objective magnification: 4x (Panel A), 23x (Panel B), 5x (Panel C), 16x (Panel D), 19.5x (Panel E), and 19.5x (Panel F).
Uterine spiral arteries are present in the metrial gland (and decidua basalis), and these maternal vessels have a critical role in supplying nutrients to the placenta and embryo.22,49 These arteries are coiled, giving them this unique name. 22 During pregnancy, interstitial trophoblast cells from the embryonic placenta migrate through the decidua and uterine wall, reaching and surrounding the spiral arteries before invading the vessel walls. Subsequently, the endovascular trophoblast cells remodel spiral arteries by replacing the muscular and elastic tissues in the walls with secreted PAS-positive acellular fibrinoid material (Figure 5F) composed of fibronectin, collagen type IV, laminin, and other basement membrane-like constituents. This fibrinoid material begins to appear under the endothelium as early as E9.5 and increases through E16 to E17. The arterial remodeling process transforms the muscular arteries into large, wide, thin-walled, low-resistance vessels, thereby increasing blood flow to the placenta (and particularly the labyrinth).7,52
Core Biomarkers for Specific Placental Cell Lineages
As with complex organs of adult animals and embryos, the placenta is a complicated amalgam of many different cell populations. Most of the principal lineages originate from embryonic precursors, but cells of the decidua and metrial gland arise from maternal elements. The various cell populations in the mouse placenta may be observed in routine (H&E-stained) sections, but more detailed characterization is facilitated by localizing specific lineages using cell type-specific markers (Table 1), the expression of which fluctuates over the course of gestation (Table 2).
Useful histochemical and immunohistochemical markers for principal cell populations in the mouse placenta.
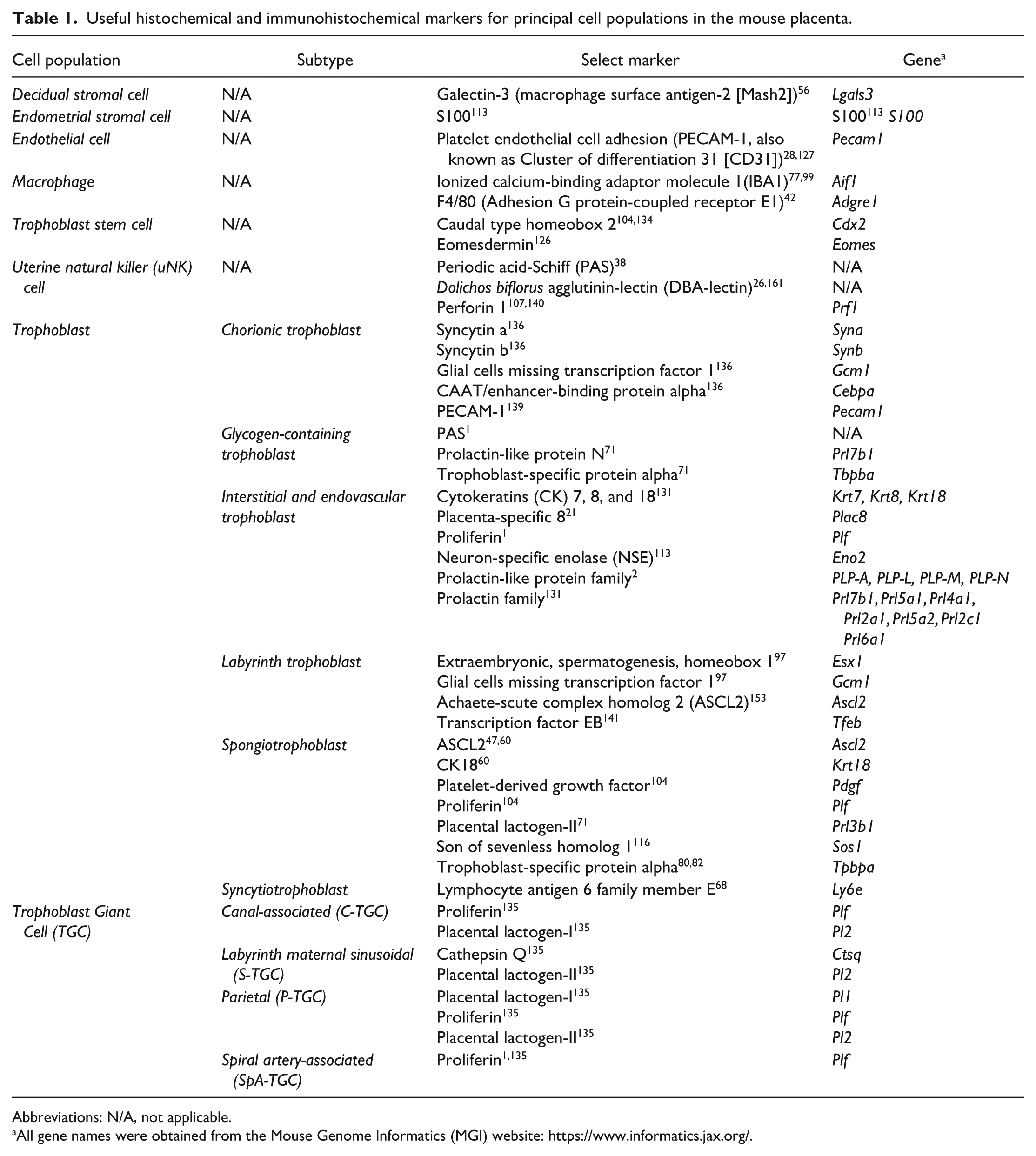
Abbreviations: N/A, not applicable.
All gene names were obtained from the Mouse Genome Informatics (MGI) website: https://www.informatics.jax.org/.
Overview of select histochemical and immunohistochemical features in the mouse placenta and metrial gland.
Trophoblast Lineages
Trophoblasts are specialized placental cells with important roles in embryo implantation, decidualization, placental immune and endocrine functions, formation of structural and biochemical barriers between the maternal and embryonic compartments, and tissue remodeling within the placenta.34,66,80,135 The initial trophoblast subtypes are primary and secondary TGCs that arise from the ectoplacental cone (Figure 3), and they mediate implantation and early placental organization. Other trophoblast subtypes arise later in development and exhibit specific phenotypes and functions depending on their locations within the placenta: spiral artery-associated endovascular trophoblasts in the major maternal vessels in the metrial gland and spanning the decidua basalis, spongiotrophoblasts and glycogen trophoblasts in the junctional zone, and cytotrophoblasts and syncytiotrophoblasts surrounding the embryonic capillaries in the labyrinth.
Trophoblast cell lineages exist prior to implantation as an epithelial cell layer of the blastocyst wall known as trophectoderm. Beginning around the time of implantation (E4.5), the trophectoderm differentiates into the polar trophoblast and the mural trophoblast. 93 The polar trophoblast borders the ICM (i.e., the crescent-shaped cell plate at one blastocyst pole that represents the nascent embryo) while the mural trophoblast forms the wall surrounding the fluid-filled cavity (blastocoel) located at the opposite blastocyst pole. Both polar and mural trophoblasts later give rise to various trophoblast subtypes throughout gestation.33,49
Primary and secondary trophoblast giant cells
Shortly after implantation, the initial wave of trophoblast differentiation produces primary TGCs,65,134 which arise from the mural trophoblast and differentiate to form a nondividing layer of enlarged cells that surround the blastocyst and separate the embryo from the endometrial surface (Figure 3).49,93,139 These cells develop by a process of endoreduplication in which DNA synthesis occurs multiple times without mitosis. The primary TGCs mediate blastocyst attachment and egg cylinder formation (Figure 3) while inciting a close relationship between invading elements of the embryonic placenta and the uterine epithelium,66,135 in particular, by regulating endometrial decidualization. 49 Concurrent with decidualization, the polar trophoblast gives rise to trophoblast stem cells that will exist as part of the ectoplacental cone and later develop into additional trophoblast subtypes. 135
Secondary TGCs develop later in gestation (~E7.5) from precursors located in the polar trophoblast-derived ectoplacental cone. 134 This second wave of TGCs is important in expanding the dimensions of the choriovitelline (yolk sac) placenta, invasion of the maternal uterine lining to allow the embryo to achieve a firmer attachment, secretion of factors that allow remodeling of the maternal vasculature, and production of various hormones and paracrine factors that regulate maternal placental function. 146
While TGCs are readily visible in routine H&E-stained sections, these initial trophoblast elements express several important marker proteins with specific spatial and temporal patterns depending on cell differentiation.2,138 Key trophoblast markers include several prolactin (PLR) family members, including placental lactogen-I (Pl-I) and II (Pl-II); PLR-like proteins (PLPs); and proliferin (PLF). 138 At least 19 different hormones/cytokines in the PLR family have been shown to be relevant in the rodent placenta. 41 Primary TGCs can be detected using Pl-I, which is expressed around E6.5 and peaks at around E9.5 before dwindling as gestation continues.41,93,96 Primary and secondary TGCs can also be visualized using heart and neural crest derivatives expressed 1 (Hand1). 135 A number of other markers are available for detecting TGCs (Table 1).
Parietal trophoblast giant cells
One subtype of migratory TGC that differentiates from the mural trophoblast of the ectoplacental cone is known as the parietal TGC. 135 At around E6.5, this cell subtype lines the entire embryo-endometrial interface, acting as a barrier separating the decidua and embryonic placenta.133,135 Primary parietal TGCs further aid decidualization while also serving as the purveyors of histiotrophic nutrition by carrying out the nutrient, gas, and waste exchange duties that are vital for embryo survival until chorioallantoic fusion occurs at around E8.5. 14 As development continues, the parietal TGCs increase in size as a result of phagocytosis and move farther apart from one another while forming cytoplasmic connections with surrounding cells. Eventually, a secondary surge of parietal TGC differentiation out of the ectoplacental cone will lead to the accumulation of secondary parietal TGCs between the junctional zone and decidua basalis. 135
Like most trophoblast lineages, parietal TGCs express hormones from the PLR family, with Pl-II serving as a principal marker. Time-dependent expression of Pl-II is upregulated beginning around E11.5, 18 although this protein is also detected in junctional zone spongiotrophoblasts and some labyrinthine trophoblasts.18,135 After E11.5, parietal TGCs continue to express Pl-II until around E17.5 when the placenta begins to degrade in preparation for birth. 18 Prior to Pl-II expression, parietal TGCs have been shown to express PLF earlier in gestation, beginning around E6.5.18,135
Spiral artery-associated endovascular trophoblast
Just after implantation, the maternal decidua contains spiral arteries that carry maternal blood to the implantation site. Highly invasive spiral artery-associated trophoblasts, a subtype of TGC, are generated in the ectoplacental cone and eventually migrate into the decidua to surround, invade, and remodel the maternal vasculature.1,64,134,135 Studies examining spiral artery-associated trophoblasts found that none of these elements are present near or in spiral arteries prior to E10.5.1,22 Thereafter, these specialized endovascular trophoblast cells intercalate between and even replace endothelial cells so that they occupy an optimal position for the actions of their angiogenic factors, vasodilators, and anticoagulants to promote greater blood flow to the embryonic placenta, especially the labyrinth. 22
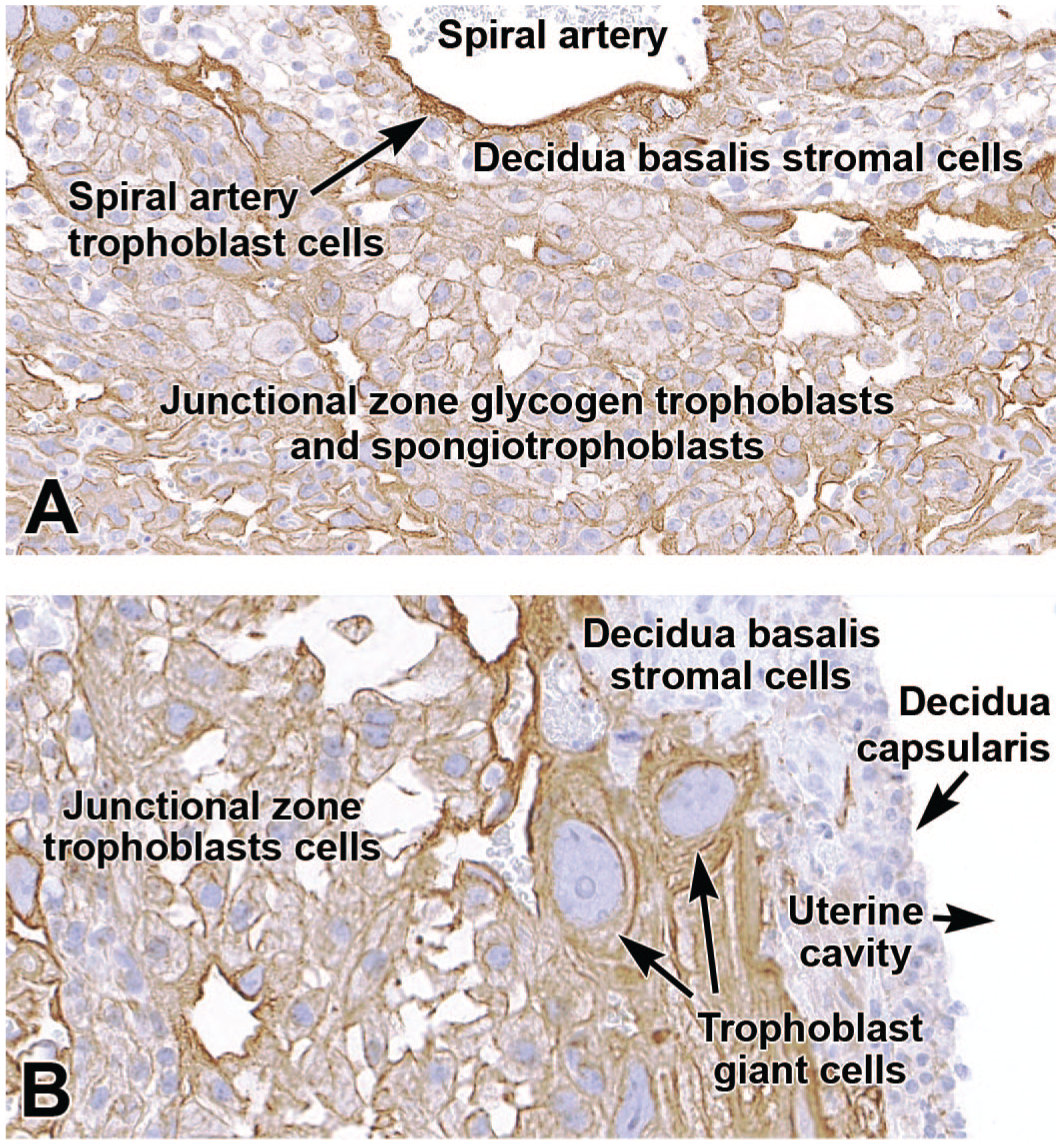
By mid-gestation (E12.5 to E14.5), spiral artery-associated trophoblasts express cytokeratin 18 (CK18) and PLF but not Pl-I, even though many other trophoblast subtypes in the surrounding placental layers strongly express Pl-I.1,54,135 Typically, spiral artery-associated trophoblasts do not line the entire vessel but instead only cover a portion of the wall while remaining juxtaposed to the initially present endothelial cells (Figure 6A and B);17,22 this intermingling can be confirmed using an endothelial marker like platelet endothelial cell adhesion molecule 1 (PECAM-1, also known as cluster of differentiation 31 [CD31]) (Figure 6C). Spiral artery-associated trophoblasts are PAS-negative, while other trophoblast derivatives in the interstitial areas around the spiral arteries, like glycogen cells in the junctional zone, are PAS-positive. 1 Additional studies attempting to differentiate among specific trophoblast subtypes have shown that spiral artery-associated trophoblasts exclusively expressed PLF and not the Pl-I or Pl-II markers found in other trophoblast subtypes. 135
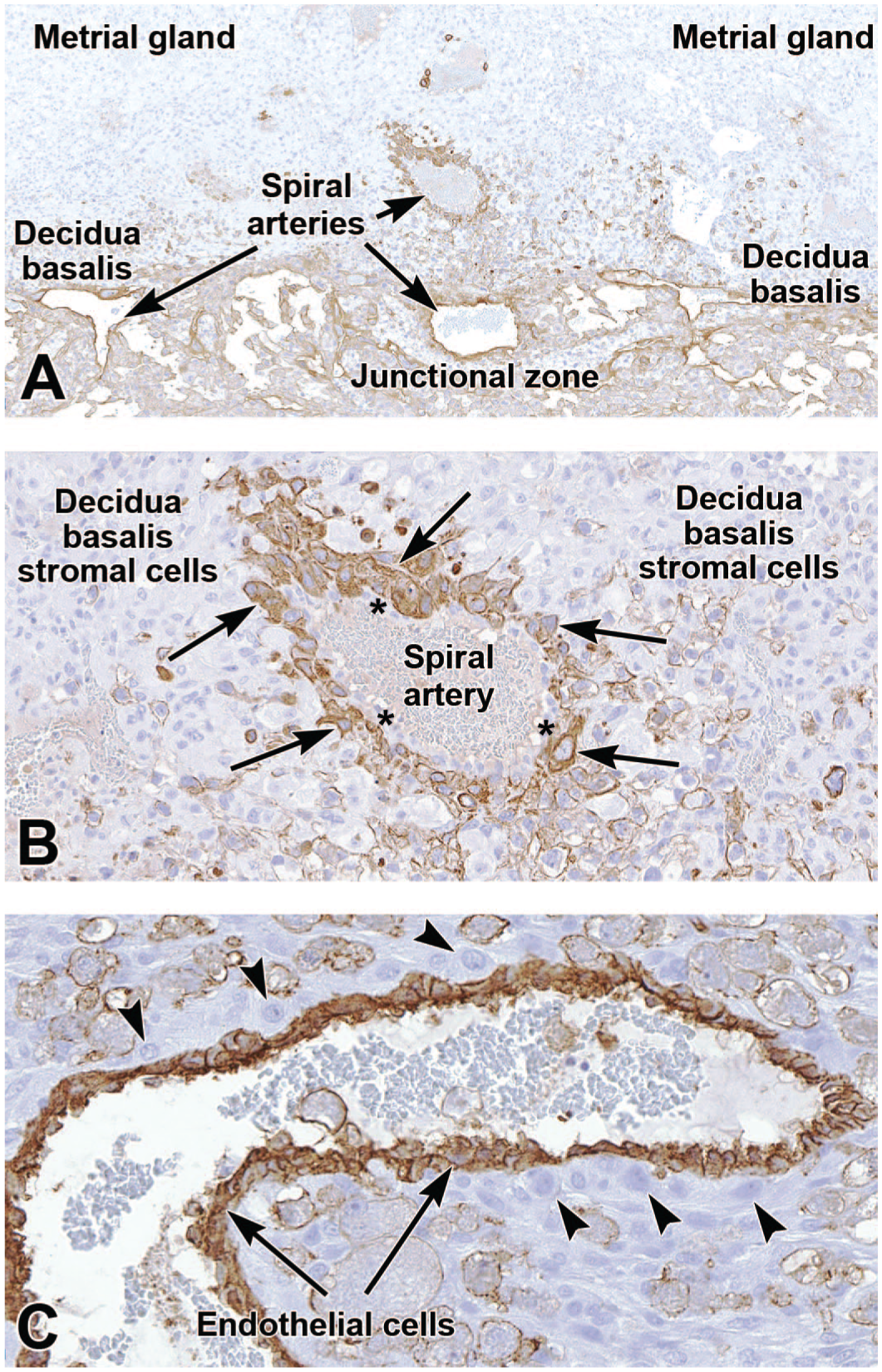
Representative images of the definitive placenta at E18.5 labeled to show cytokeratin 18 (CK18 [Panels A and B]) and cluster of differentiation 31 (CD31 [Panel C]). CK18 is an intermediate filament protein expressed in the cytoplasm of epithelial cells, including spiral artery-associated endovascular trophoblasts (arrows in Panel B). These trophoblasts do not form the entire artery wall since some intact endovascular cells do not express CK18 (asterisks in Panel B). CD31 (alternatively platelet endothelial cell adhesion molecule 1 [PECAM-1]) is an integral membrane glycoprotein in endothelial cells that mediates cell-to-cell adhesion and is expressed at high levels in spiral artery walls. Panel C illustrates the close association of spiral artery-associated trophoblast cells (arrowheads) and endothelial cells. Immunohistochemistry using 3,3’-diaminobenzidine (DAB [brown color]) as the chromogen and hematoxylin as the counterstain. Original objective magnification: 7x (Panel A), 23x (Panel B), and 36x (Panel C).
Spongiotrophoblasts and glycogen trophoblasts
Spongiotrophoblasts and glycogen trophoblasts (glycogen cells) are both located in the junctional zone. Spongiotrophoblasts likely originate from an ectoplacental cone precursor, while glycogen cells arise from spongiotrophoblasts. 134 During gestation, the junctional zone has a significant level of cellular proliferation, with the number of nonmigratory spongiotrophoblast cells increasing 4-fold while the migratory glycogen cells increase 250-fold to facilitate invasion into the decidua. 133 Differentiation of spongiotrophoblast cells and glycogen cells occurs by E9.5.
Spongiotrophoblasts arranged in sheets are the predominant cells in the junctional zone at E10.5. These cells are identified readily in H&E-stained sections by their characteristic clear, colorless, vacuolated cytoplasm. Spongiotrophoblasts remain the principal cell type in the junctional zone at E12.5, double their numbers by E16.5, and decline by about 50% by E18.5. 29 These elements function to produce hormones, such as prolactin and placental lactogen-II, and other factors like cytokines needed to sustain pregnancy. Sometime after E12.5, they begin to migrate into the decidua basalis. 1
Glycogen cells occur as islands within the spongiotrophoblast fields of the junctional zone and are distinguished by their extensive PAS-positive cytoplasmic glycogen accumulation (Figure 7). Their population substantially increases between E12.5 and E16.5, declining thereafter. 29 Glycogen cells are thought to metabolize their glycogen stores to release the energetically favorable substrate glucose-6-phosphate for the developing placenta and embryo, and they are the main source of insulin-like growth factor 2 (IGF2) within the placenta in the second half of gestation. 119 Placental IGF2 is a paternally-imprinted growth factor that regulates embryonic cell division and metabolism.
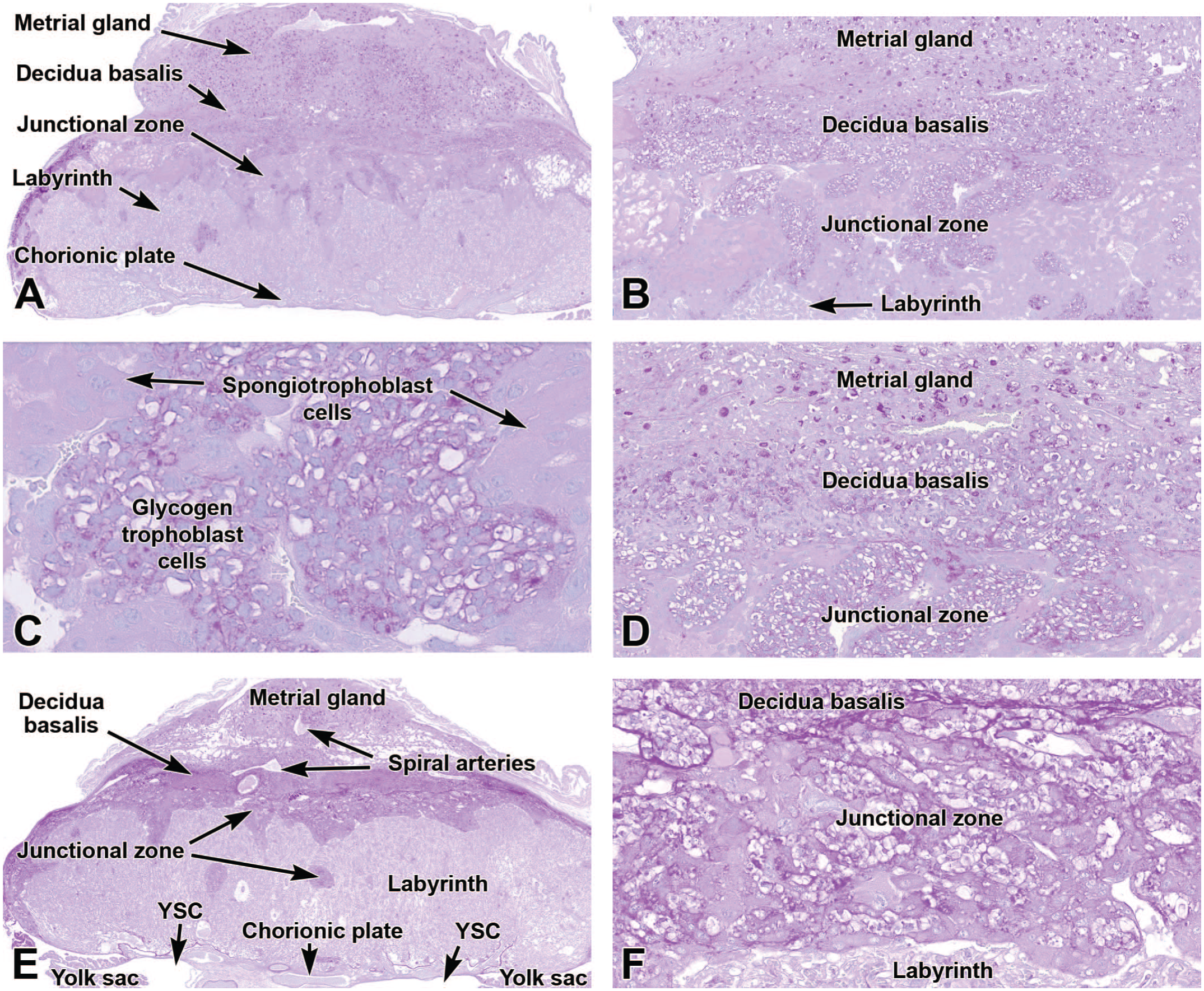
Representative images of the junctional zones of the definitive placenta at E15.5 (Panels A-D) and E18.5 (Panels E-F) stained with periodic acid-Schiff (PAS) to highlight embryo-derived glycogen-containing trophoblast cells (“glycogen cells”). PAS is a histochemical stain that binds polysaccharides (such as glycogen), mucosubstances, and basement membranes. Panels A (E15.5) and E (E18.5) show the relationship of the junctional zone to other placental areas, while Panels B, C, D, and F demonstrate various placental regions at higher magnifications. Glycogen trophoblast cells are widespread in the junctional zone (Panels C, D, and F) and are dispersed in lesser numbers in the decidua basalis (Panel D). Unstained spongiotrophoblast cells surround clusters of glycogen trophoblast cells in the junctional zone (Panels B, C, and D). In Panel D, the more vibrantly PAS-positive cells in the metrial gland are uterine natural killer (uNK) cells of maternal origin. YSC = yolk sac cavity. PAS. Original objective magnification: 2x (Panel A), 7.5x (Panel B), 40x (Panel C), 14x (Panel D), 1.6x (Panel E), 19.5x (Panel F).
Trophoblasts of the junctional zone can be identified using various markers. Both spongiotrophoblasts and glycogen cells express CK18 (Figure 8). 149 As noted above, glycogen cells are readily distinguished by their high content of PAS-positive glycogen, while spongiotrophoblasts are PAS-negative. A common marker for spongiotrophoblasts is the basic helix-loop-helix transcription factor achaete-scute complex homolog 2 (ASCL2, formerly known as mammalian ASCL2 [Mash2]), which occurs in the ectoplacental cone, spongiotrophoblast layer, and chorion. 47 Expression of ASCL2 is evident from E8.5 to E12.5 in trophoblasts of various placental regions, but after E12.5 this marker is mainly expressed in spongiotrophoblasts.47,60 This exclusivity explains the phenotype of Ascl2 knockout mice, where placentas of null mutants have an overabundance of TGCs, a poorly developed labyrinth layer, reduced chorionic ectoderm, and an absent spongiotrophoblast layer.47,60,130 These mutant conceptuses fail to progress properly after E10.5, ultimately resulting in embryonic death.
Representative images of the definitive placenta at E15.5 demonstrating cytokeratin 18 (CK18 [brown labeling]) in spiral artery trophoblasts, spongiotrophoblasts, and trophoblast giant cells in or near the junctional zone. While CK18 is expressed in trophoblast cells, it is not a specific marker for any one type of trophoblast. Panel A illustrates how the CK18-positive spongiotrophoblasts in the junctional zone can be readily differentiated from the unlabeled stromal cells of the decidua basalis. Panel B shows two large trophoblast giant cells with CK18-positive cytoplasm located at the border between the junctional zone and decidua basalis. Note the lack of CK18 expression in the decidua basalis and decidua capsularis, which are derivatives of the endometrial stroma. Immunohistochemistry using 3,3’-diaminobenzidine (DAB [brown]) as the chromogen and hematoxylin as the counterstain. Original objective magnification: 23x (Panel A) and 40x (Panel B).
Cytotrophoblasts and syncytiotrophoblasts
The chorionic trophoblast is a layer of cells from the embryonic chorion that differentiate into specialized cells, the cytotrophoblasts and the syncytiotrophoblasts, that form the maternal-fetal interface, including the labyrinthine and junctional zones. 72 The labyrinth layer of the placenta is induced at the time of chorioallantoic fusion (E8.5). Upon contacting the chorionic plate, the allantoic mesoderm erodes the chorionic tissue and branches into it to promote vascularization of the labyrinth. This process (termed “branching morphogenesis”) encourages the formation of multiple differentiated trophoblast cell types in organized, densely packed columns that contribute to regulating the countercurrent exchange between parallel networks of maternal sinusoids and embryonic capillaries.
The principal trophoblast populations within the labyrinth of the hemotrichorial mouse placenta are cytotrophoblasts (also known as sinusoidal TGCs) and syncytiotrophoblast types I and II (Figure 4).49,133,136 The term “hemotrichorial” denotes that the expected three layers of maternal cells (maternal endothelium, connective tissue, and uterine epithelium) are missing while the trophoblast layers abutting the embryonic capillaries are formed by three trophoblast layers; by comparison, the primate (including human) placenta has a hemomonochorial (one trophoblast layer) configuration. 12 The cytotrophoblasts are mononuclear cells that arise from chorionic trophoblast progenitors and form a discontinuous monolayer that lines the walls of maternal sinusoids. The syncytiotrophoblasts are multinucleated cells that originate by fusion of adjacent cytotrophoblasts, after which the syncytial cells form two continuous layers sandwiched between the cytotrophoblasts and the walls of embryonic capillaries.49,133,136 Together, these three labyrinth trophoblast subtypes and the endothelial cells of embryonic capillaries form the interhemal membrane unit that separates the maternal sinusoidal and embryonic capillary circulations (Figure 4).31,133
All trophoblasts within the labyrinth express CK18, an epithelial cytoskeletal protein (Figure 9). Syncytiotrophoblast type II cells can be highlighted via the marker glial cells missing transcription factor 1 (Gcm1). 139 The Gcm1 protein is initially present in the chorion prior to labyrinth induction and thereafter is involved in branching morphogenesis and cell-cell fusion, thereby aiding in the formation of syncytiotrophoblasts. 139 After completion of branching morphogenesis (around E12.5), Gcm1 is coexpressed with CCAAT/enhancer-binding protein alpha (Cebpa) and syncytin B (Synb) specifically in syncytiotrophoblast type II cells.136,139 At the same time, syncytiotrophoblast type I cells express syncitin A (Syna) while cytotrophoblasts express cathepsin Q (Ctsq) and Hand1. 136 Cytotrophoblasts also express platelet-derived growth factor B (PdgfB). 98 Indeed, Pdgfb-deficient mice have placentas with a reduced labyrinth trophoblast population, leading to a reduced maternal-embryonic vascular surface area and thus disrupted labyrinth organization, leading to decreased countercurrent exchange.8,27,98
Representative images from the primitive placenta at E10.5 (Panels A-B) and the definitive placenta at E18.5 (Panels C-E) demonstrating expression of cytokeratin 18 (CK18 [brown labeling]) to highlight various trophoblast types. Panel A at E10.5 shows the overall organization of the placenta and the restricted CK18 expression in trophoblasts, chiefly localized in regions of embryo-derived tissue (junctional zone, labyrinth, and yolk sac). Panel B at E10.5 is a higher magnification of the junctional zone and labyrinth showing CK18-positive cytotrophoblasts and syncytiotrophoblasts. Panels C, D, and E at E18.5 show portions of placenta from three adjacent embryos at E18.5 to demonstrate CK18 expression in the junctional zone, labyrinth, and yolk sac as well as around large blood vessels in the decidua basalis (due to invading endovascular trophoblast cells migrating from the junctional zone) and the decidua capsularis. Note the labyrinth and metrial gland in Panel C from adjoining embryos (upper left and right corners of the image). Panel D shows cytoplasmic CK18 expression in cytotrophoblast and syncytiotrophoblast cells of the labyrinth that separate the maternal blood sinusoids from embryonic capillaries. Panel E shows CK18 expression in various yolk sac components. Immunohistochemistry using 3,3’-diaminobenzidine (DAB [brown]) as the chromogen and hematoxylin as the counterstain. Original objective magnification: 1.3x (Panel A), 7.5x (Panel B), 2x (Panel C), 20x (Panel D), 22x (Panel E).
Proliferating trophoblasts
Placental cell proliferation can be visualized with the marker Ki67,51,151 a nuclear protein expressed only in the G1, S, and G2 phases of the cell cycle but not in cells during the quiescent G0 phase.51,129,145 Ki67 expression continues after the G2 phase into mitosis (M phase), allowing this marker to delineate cells that are actively moving through the cell cycle from those that are arrested in their progression.152,159
Following implantation at E4.5, the most prominent dividing cells in the placenta are the primary TGCs and their progenitors in the ectoplacental cone. Later in gestation, further proliferation in the ectoplacental cone gives rise to secondary TGCs and thereafter to the trophoblast glycogen and spongiotrophoblast cells that proliferate and move throughout the decidua and later into the labyrinth and junctional zone of the chorioallantoic (definitive) placenta. 49 Ki67 can be used to detect distinct areas of placental growth at different times during gestation, with the key timepoints for cell proliferation occurring between E4.5 and E12.5 (Figure 10). Proliferation of placental cells generally decreases as gestation continues, although the extent of the decline depends on the placental layer.
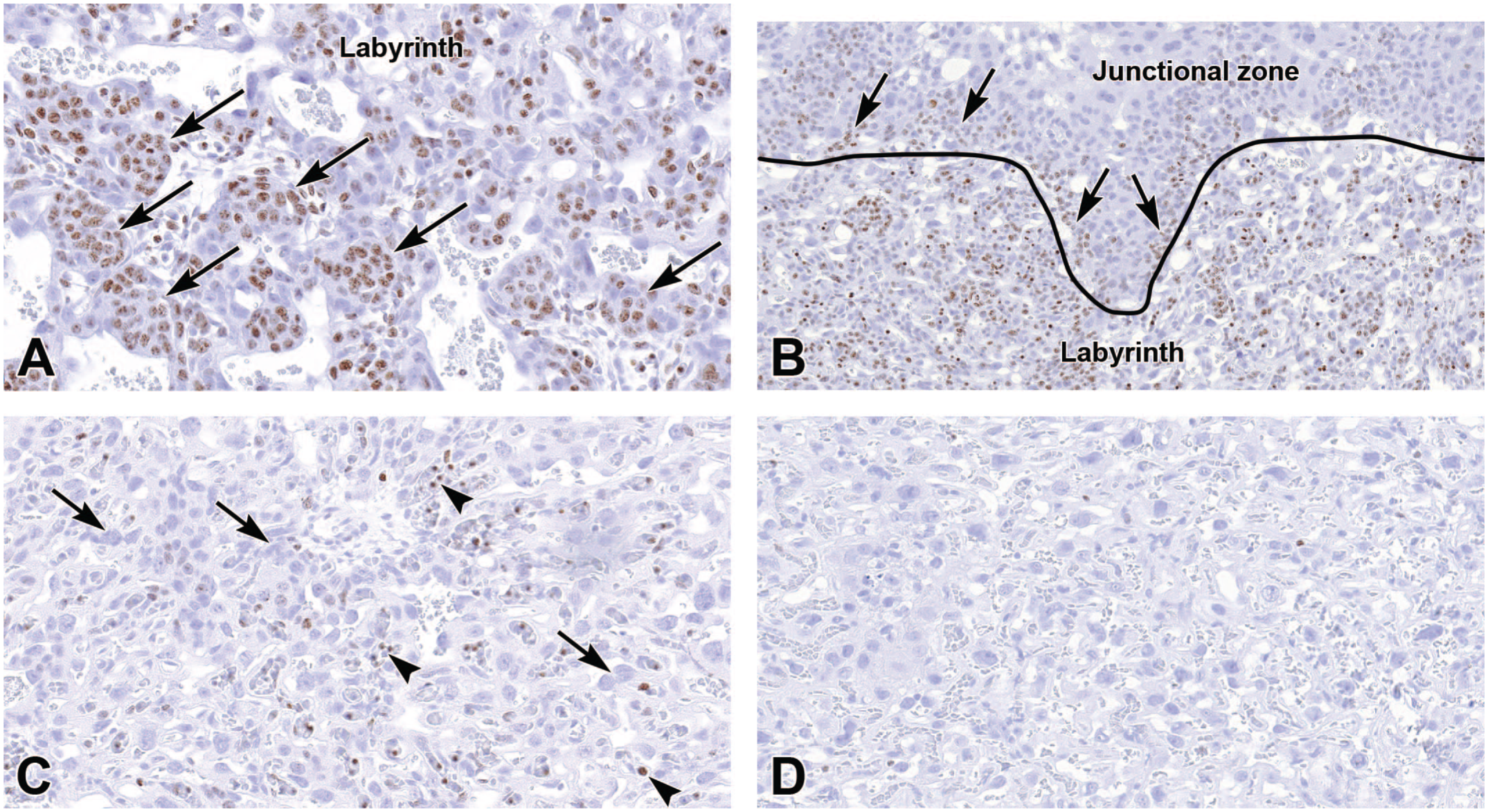
Representative images of the labyrinth of the definitive placenta at E12.5 (Panels A-B), E15.5 (Panel C), and E18.5 (Panel D) demonstrate cell proliferation (brown labeling) using the marker Ki67, a nuclear protein expressed during the G1, S, and G2 phases of the cell cycle and into mitosis (M phase). Panels A and B (at E12.5) show proliferating trophoblast cells (arrows) of the labyrinth and junctional zone. Panel C (at E15.5) emphasizes the lack of proliferative trophoblast cells (long arrows) in the labyrinth later in gestation while the embryonic circulation contains scattered Ki67-positive nucleated blood cells (arrowheads). Panel D (at E18.5) demonstrates the lack of proliferative cells in the labyrinth near term, including the lack of nucleated blood cells in embryonic vessels. Immunohistochemistry using 3,3’-diaminobenzidine (DAB [brown color]) as the chromogen and hematoxylin as the counterstain. Original objective magnification: 28x (Panel A), 16x (Panel B), 29.5x (Panel C), 29.5x (Panel D).
Endothelial Cells
The endothelial marker PECAM-1 (CD31) can be used to detect angiogenesis throughout the placenta and metrial gland (Figure 11). This integral membrane glycoprotein plays a prominent role in cell-cell adhesion and maintains endothelial cell monolayers that line blood vessels, especially for the embryonic capillaries of the labyrinth. 101 The presence of PECAM-1 is essential for vascular integrity as Pecam1 knockout mice exhibit enhanced vascular permeability.19,86,157
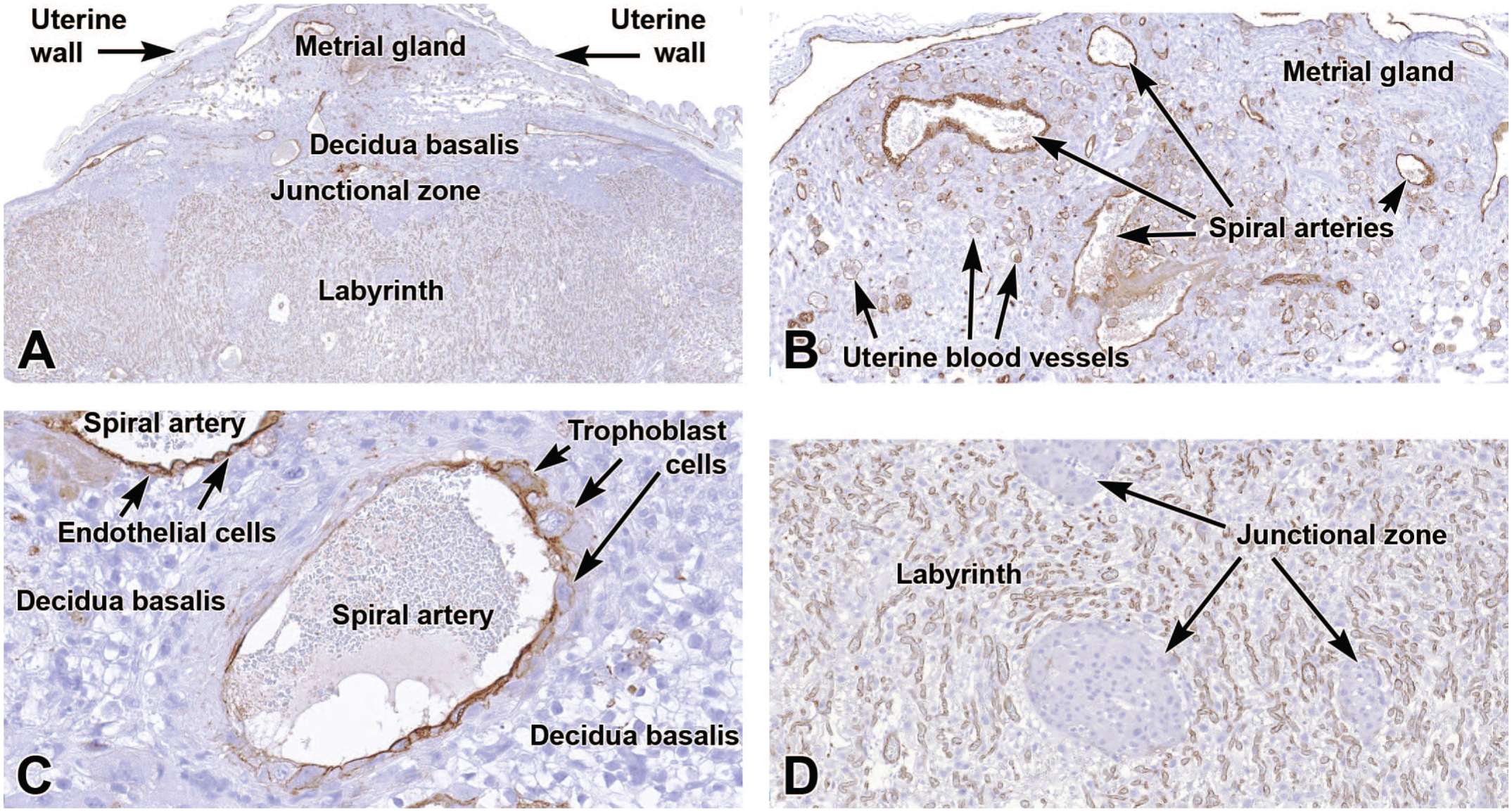
Representative images of the definitive placenta at E15.5 showing robust expression of cluster of differentiation 31 (CD31 [brown labeling]) by endothelial cells throughout the placenta. Panel A shows the overall pattern of CD31 expression in the placenta. Panels B and C demonstrate labeling of maternal-derived spiral arteries and smaller arterial branches within the metrial gland (Panel B) and decidua basalis (Panel C). The spiral artery (Panel C) shows trophoblast invasion (evident as interruption of the CD31-positive endothelial cell lining) that remodels the small uterine blood vessels into wider, low-resistance vessels. Panel D shows widespread CD31 expression of embryo-derived endothelial cells in the highly vascular labyrinth but no CD31 expression in the nonvascular penetrating columns of junctional zone tissue. Immunohistochemistry using 3,3’-diaminobenzidine (DAB [brown color]) as the chromogen and hematoxylin as the counterstain. Original objective magnification: 2.5x (Panel A), 10x (Panels B and D), and 28x (Panel C).
As the allantois contacts the chorion, its branches penetrate to initiate the dense vascular beds of the labyrinth layer. A multitude of new blood vessels form in this layer beginning around E8.5, and they exist continuously until birth at approximately E19.5. 103 In addition, PECAM-1 is also expressed in endothelial cells in the mature chorionic plate, junctional zone, metrial gland, and in the spiral arteries of the decidua (Figure 11).
Importantly, PECAM-1 is also expressed in other placental cell populations such as platelets and most leukocytes, albeit at reduced levels compared to endothelial cells. 101 Due to this broader and variable expression pattern, careful examination of all tissues and cells is warranted when utilizing PECAM-1 to examine functions of specific vessel-associated cell types.
The placenta does not have lymphatic vessels. Nonetheless, lymphatic markers have been reported in human 154 and mouse 109 placentas, including differential expression in various human placental structures based on the particular location within the organ. Expression of lymphatic-related genes in the mouse placenta occurs in remodeled spiral arteries between E11.5 and E13.5 and includes such proteins as lymphatic vessel endothelial hyaluron receptor 1 (Lyve-1), Prospero-related homeobox 1 (Prox1), and vascular endothelial growth factor receptor 3 (VEGFR3).37,109 The uterine wall does contain a complement of lymphatic vessels. Therefore, in typical cases, the only reason for using lymphatic markers in placental investigations would be to investigate potential alterations in lymphatic vessels within the decidua basalis or vicinity of the metrial gland.
Uterine Natural Killer Cells
Maternally derived uNK cells are a specialized and transient leukocyte population found in the placental and uterine tissues during pregnancy. These cells are the most abundant innate lymphoid cells in the uterus, congregating at the maternal-embryo interface where their principal functions include modulation of spiral artery remodeling (as they produce angiogenic and vasoactive factors67,73), regulation of angiogenesis in the decidua, and control of trophoblast invasion into maternal tissue. 53 The conceptus is a semi-allogenic (genetically different [“foreign”]) structure in that it harbors both maternal and paternal antigens. Accordingly, the placenta depends on leukocytes (lymphocytes and macrophages) to prevent rejection of the conceptus by a maternal immune response. As cells of maternal lymphocyte lineage, uNK cells regulate the immunological barrier between maternal and embryonic tissues.88,140
Though present in modest numbers beginning at the time near implantation (E4.5), uNK cells begin to congregate in the uterine tissue surrounding the ectoplacental cone at about E5.5, 160 where they and trophoblasts will cooperate to control decidualization. 83 During decidualization, uNK cells modify the vascular arrangement of the uterine tissue to increase blood flow, but these changes reverse after birth. 40 Mice lacking uNK cells have reduced decidua basalis and metrial gland development as well as diminished vascular remodeling that is evident by approximately E8.5.57,59 The number of uNK cells continue to increase throughout gestation until approximately E14.5, at which time their numbers stabilize until they begin to decrease near birth (compare Figures 12 and 13).39,49,83
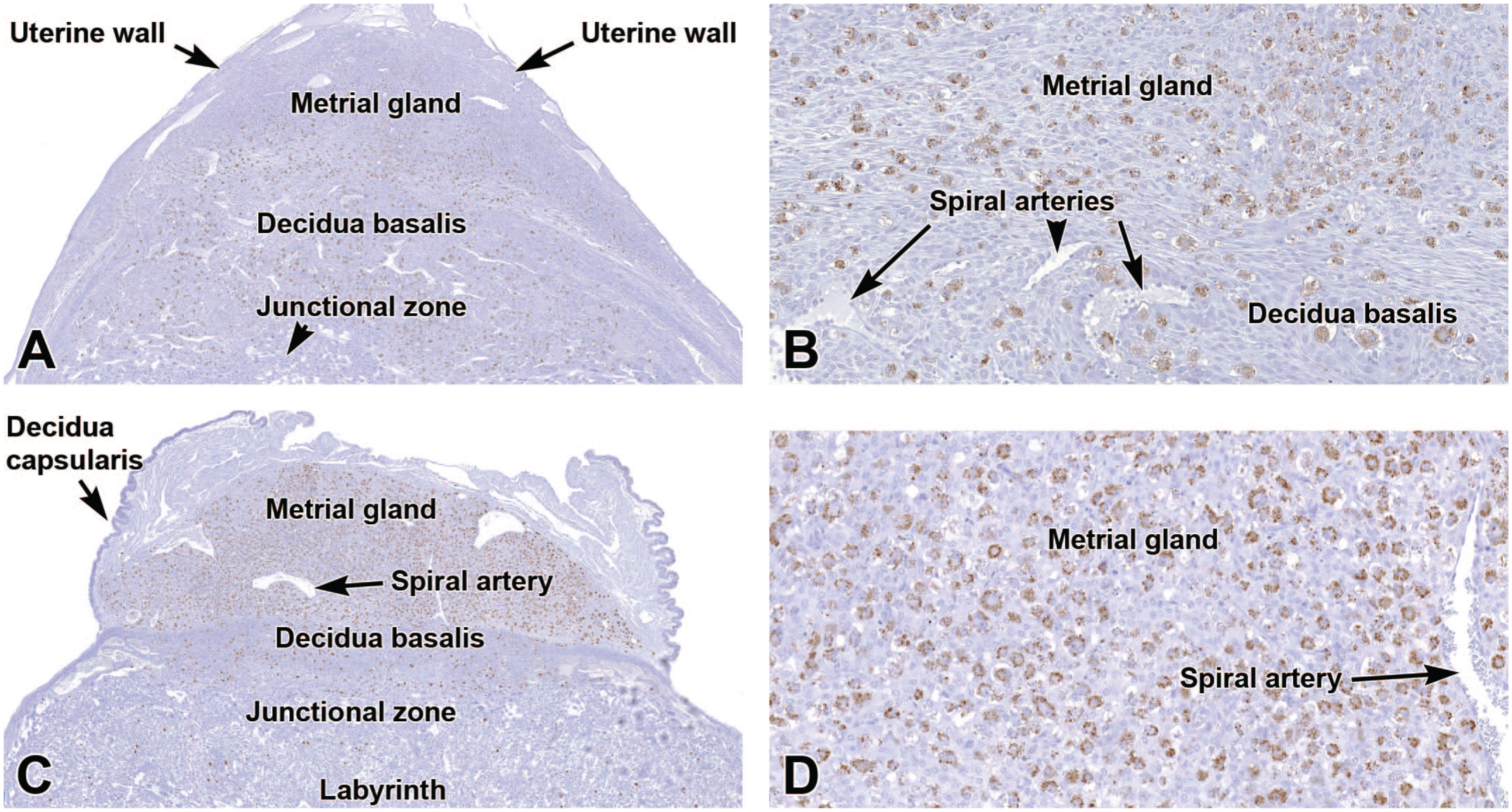
Representative images of the primordial placenta at E9.5 (Panels A-B) and definitive placenta at E12.5 (Panels C-D) demonstrating the distribution of perforin 1 (PRF1 [brown labeling]), a glycoprotein concentrated in the cytoplasmic granules of uterine natural killer (uNK) cells. The uNK cells are numerous in the metrial gland, less common in the decidua basalis, and essentially absent in the embryo-derived placental layers (junctional zone and labyrinth). Immunohistochemistry using 3,3’-diaminobenzidine (DAB [brown color]) as the chromogen and hematoxylin as the counterstain. Original objective magnification: 4x (Panel A), 18x (Panel B), 3x (Panel C), 18x (Panel D).
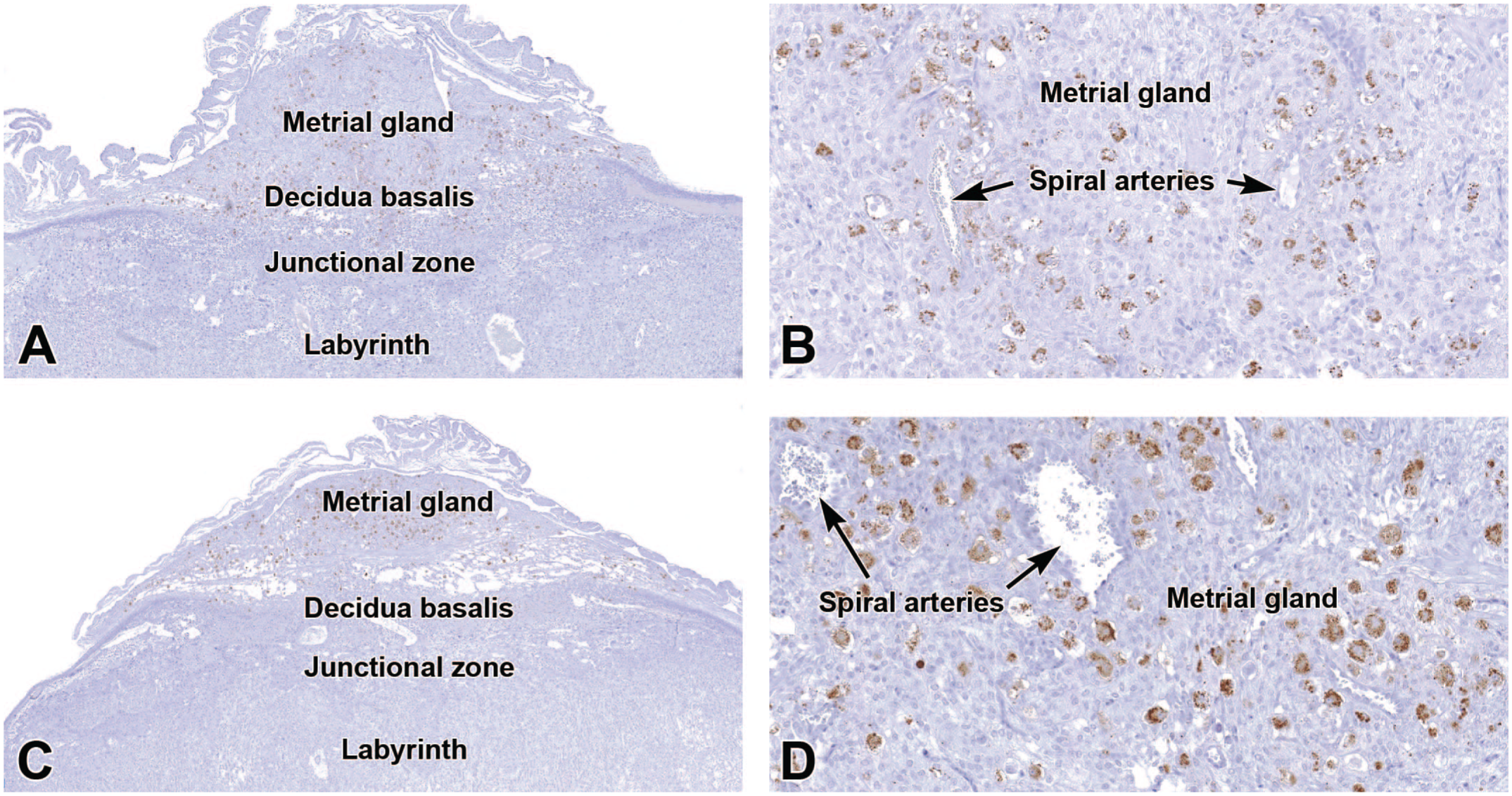
Representative images of the definitive placenta at E16.5 (Panels A-B) and E18.5 (Panels C-D) demonstrating perforin 1 (PRF1) in cytoplasmic granules of uterine natural killer (uNK) cells in the metrial gland and decidua basalis. Note the decreased numbers of uNK cells compared to the initial maturation of the definitive placenta (E12.5 [see Figure 12D]) as well as the smaller size of the metrial gland and decidua basalis in the near-term placenta compared to E9.5 (see Figure 2A). In Panel D (at E18.5), the increased uNK cell density compared to E16.5 (Panel B) is due to the decreased metrial gland size resulting from a decline in the amount of metrial gland stroma. Immunohistochemistry using 3,3’-diaminobenzidine (DAB [brown color]) as the chromogen and hematoxylin as the counterstain. Original objective magnification: 3.5x (Panel A), 18x (Panel B), 3x (Panel C), 18x (Panel D).
Several markers are expressed by uNK cells. The presence of large cytoplasmic granules makes visualization of uNK cells easy by H&E (Figure 2D). Various glycoproteins are present on their surface and within their granules, including perforin, which can be used to demonstrate that uNK cells are primarily localized to the metrial gland and, to a lesser extent, the decidua basalis and embryonic placenta (Figures 12 and 13).6,7,49,140 While perforin is also secreted by CD8+ and CD4+ T lymphocytes, the large size of uNK cells (typically 40-100 micrometers) compared to the smaller size of conventional lymphocytes (8-10 micrometers) allows these cell populations to be discerned with ease. 102 Other useful uNK markers include Dolichos biflorus agglutinin (DBA) lectin, a cell surface glycoprotein, and PAS, which stains cytoplasmic granules (Figure 5A-E). 161 While uNK cells have intracellular granules that contain perforin, granzymes, and granulysin, their general cytotoxic activity appears to be reduced compared with peripheral blood NK cells. 70
T Lymphocytes
Few studies have assessed the T cell complement of the mouse placenta, but T cells have been reported to comprise approximately 3% of mouse decidual leukocytes (Figure 14).37,94 Placental T cells are primarily of maternal origin. 90
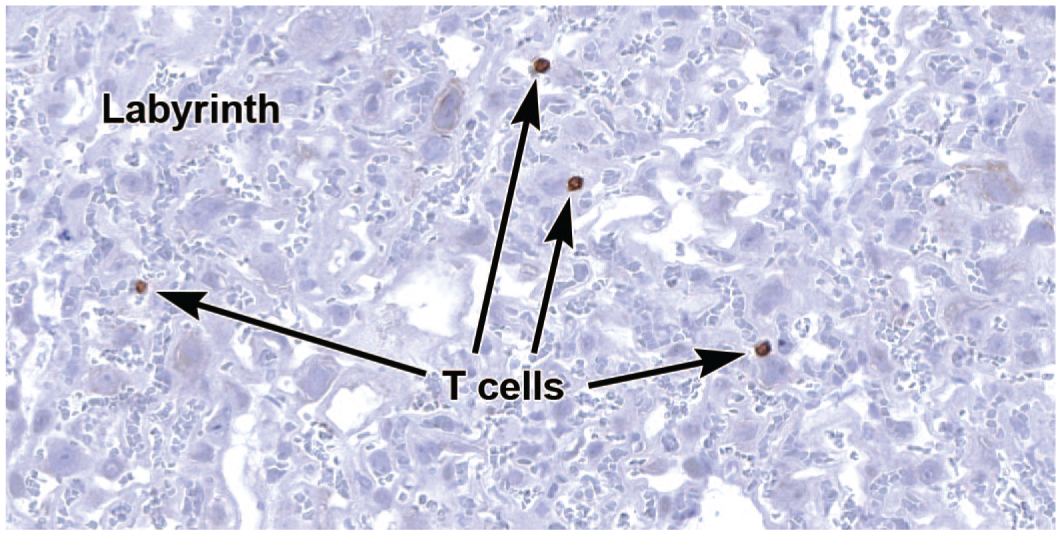
Representative image of the labyrinth of the definitive placenta at E18.5 to demonstrate rare, maternally derived T lymphocytes expressing cluster of differentiation 3 (CD3). This cell surface protein complex is made up of integral membrane glycoproteins that associate with the T cell receptor (TCR). Immunohistochemistry using 3,3’-diaminobenzidine (DAB [brown color]) as the chromogen and hematoxylin as the counterstain. Original objective magnification: 32.5x.
Similar to uNK cells, subsets of regulatory T (Treg) cells in the placenta control maternal immune function to prevent rejection of the semi-allogenic conceptus.78,132 Even before conception, Treg cell induction within the endometrium is initiated by exposure to seminal fluid.78-80 During gestation, Treg cell activity plays a prominent role in immune tolerance at two maternal-embryo interfaces: one that forms early in gestation and a second that develops later when maternal blood flow to the labyrinth commences. The initial interface is where maternal decidual cells first contact embryonic trophoblasts. The later labyrinthine interface is where maternal blood percolates through a mesh of sinusoids lined entirely by embryonic trophoblasts (Figure 4). Similar to uNK cells, properly regulated maternal-origin Treg cells do not recognize the embryonic trophoblast cells as foreign.
In addition to their function in maintaining a viable pregnancy, Treg cells are also thought to have a role in endometrial remodeling in preparation for implantation of the blastocyst. 94 Pregnant mice deficient in Treg cells exhibit disrupted implantation, and even when successful implantation occurs, the subsequent pregnancy is fraught with difficulties that often culminate in embryonic death and conceptus resorption.123,132 The prevention of early conceptus rejection is most important from implantation throughout the initial function of the choriovitelline placenta (about E4.5 to E8.5), and during this span, Treg cells are recruited in mounting numbers to the maternal-embryonic interface. Moreover, Treg cells persist within the placenta throughout pregnancy and endure in uterine tissues while maintaining functionality beyond birth.5,124 The expansion of these cells in placental tissues also correlates with decreased fetal resorption during subsequent pregnancies that involve the same paternal background. 124
Cluster of differentiation 3 (CD3) is the quintessential T cell marker. This cell surface protein complex is composed of 4 distinct subunits (delta, epsilon, gamma, and zeta) of integral membrane glycoproteins that associate with the T cell receptor (TCR), which is required for signal transduction and cell activation. Markers for additional T cell subpopulations (e.g., CD4 for T-helper cells) are seldom used when screening mouse placental samples for conventional phenotypic analysis and toxicity testing.
Macrophages
The majority of murine macrophages in the placenta are derived from hematopoietic precursors produced in the yolk sac and are therefore embryonic in origin. 25 These newly formed macrophages can be detected in the placenta at approximately E7.5, primarily in the perivascular mesenchyme.20,58,77,91,120,164 Although these macrophages constitutively express major histocompatibility complex II, they show a decreased ability to present antigen. 20 However, these cells can perform Fc-receptor-mediated phagocytosis. 20 Therefore, these cells likely supplement the activities of the uterine macrophages by providing additional defense against microbial invasion. There may be additional functions of murine placental macrophages that have yet to be elucidated.
Early in development (e.g., E9.5 [Figure 15]), macrophages are predominantly present in the metrial gland, decidua basalis, and decidua capsularis. As gestation progresses (e.g., E18.5 [Figure 16]), fewer macrophages populate the metrial gland and decidua basalis while more macrophages concentrate in the connective tissue where the metrial gland attaches to the uterine wall as well as in the labyrinth and chorion. Macrophages are present in large numbers in the decidua capsularis that forms over the placenta and embryo (Figure 16). 91 This tissue layer gradually degenerates and disappears as the conceptus grows. 43 The macrophages in the decidua capsularis play crucial roles, including immune tolerance, tissue remodeling, angiogenesis, and defense of the conceptus against infection.84,144,162 Because the decidua capsularis is easily torn during sampling, it may be missing in the final specimen unless the placenta and uterine wall are processed as an intact unit.
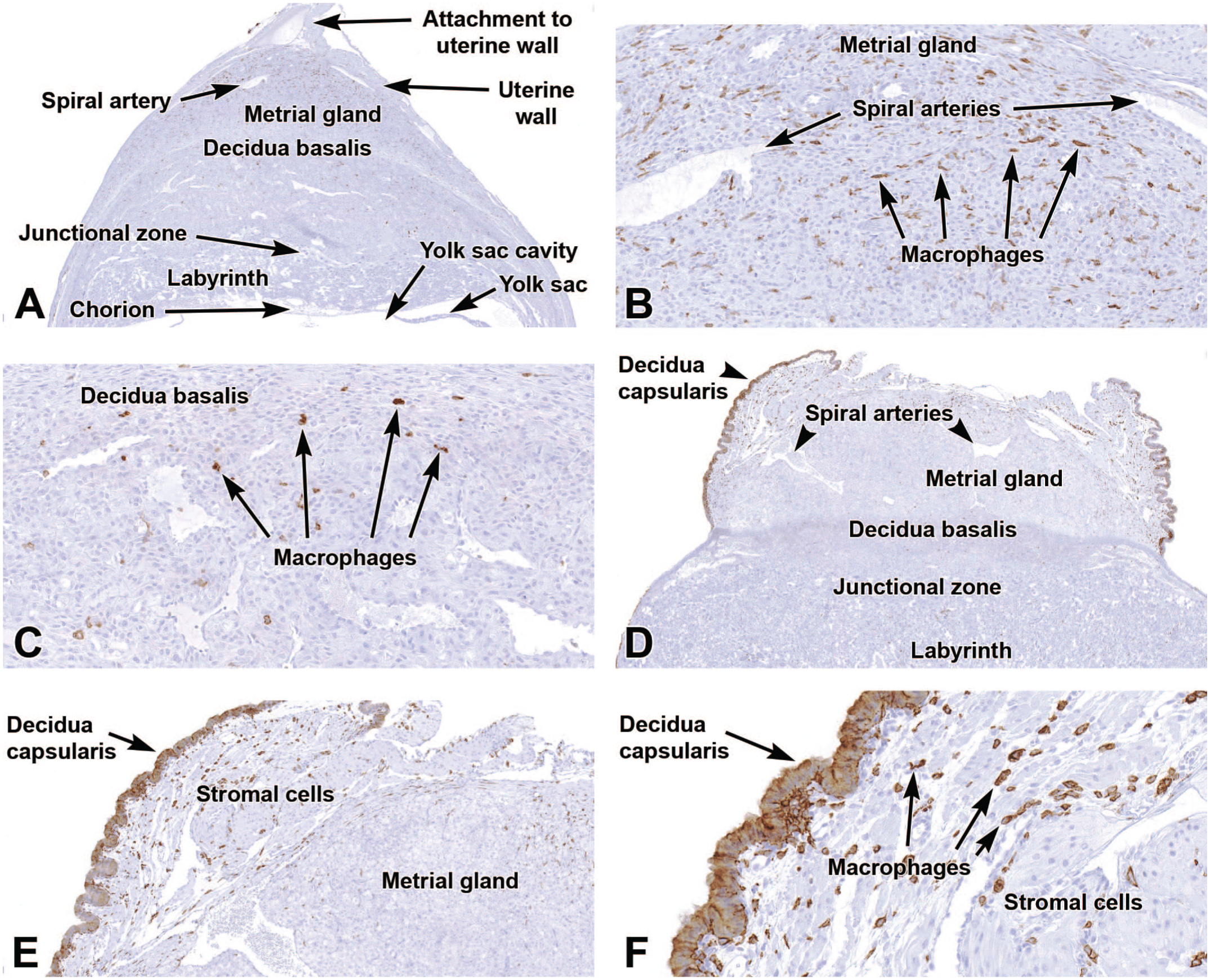
Representative images of the primordial placenta at E9.5 (Panels A-C) and the definitive placenta at E12.5 (Panels D-F) to show expression of F4/80, a transmembrane protein expressed by cells of the macrophage lineage. At E9.5, Panel A shows limited labeling in the metrial gland (see also Panel B) and decidua basalis (see also Panel C) with virtually no expression in the junctional zone, labyrinth, chorion, and yolk sac. At E12.5, Panel D demonstrates macrophages scattered mainly in the decidua capsularis and underlying fields of undifferentiated stromal cells (see also Panels E and F). Immunohistochemistry using 3,3’-diaminobenzidine (DAB [brown color]) as the chromogen and hematoxylin as the counterstain. Original objective magnification: 3x (Panel A), 18x (Panel B), 18x (Panel C), 2.5x (Panel D), 7x (Panel E), and 23x (Panel F).
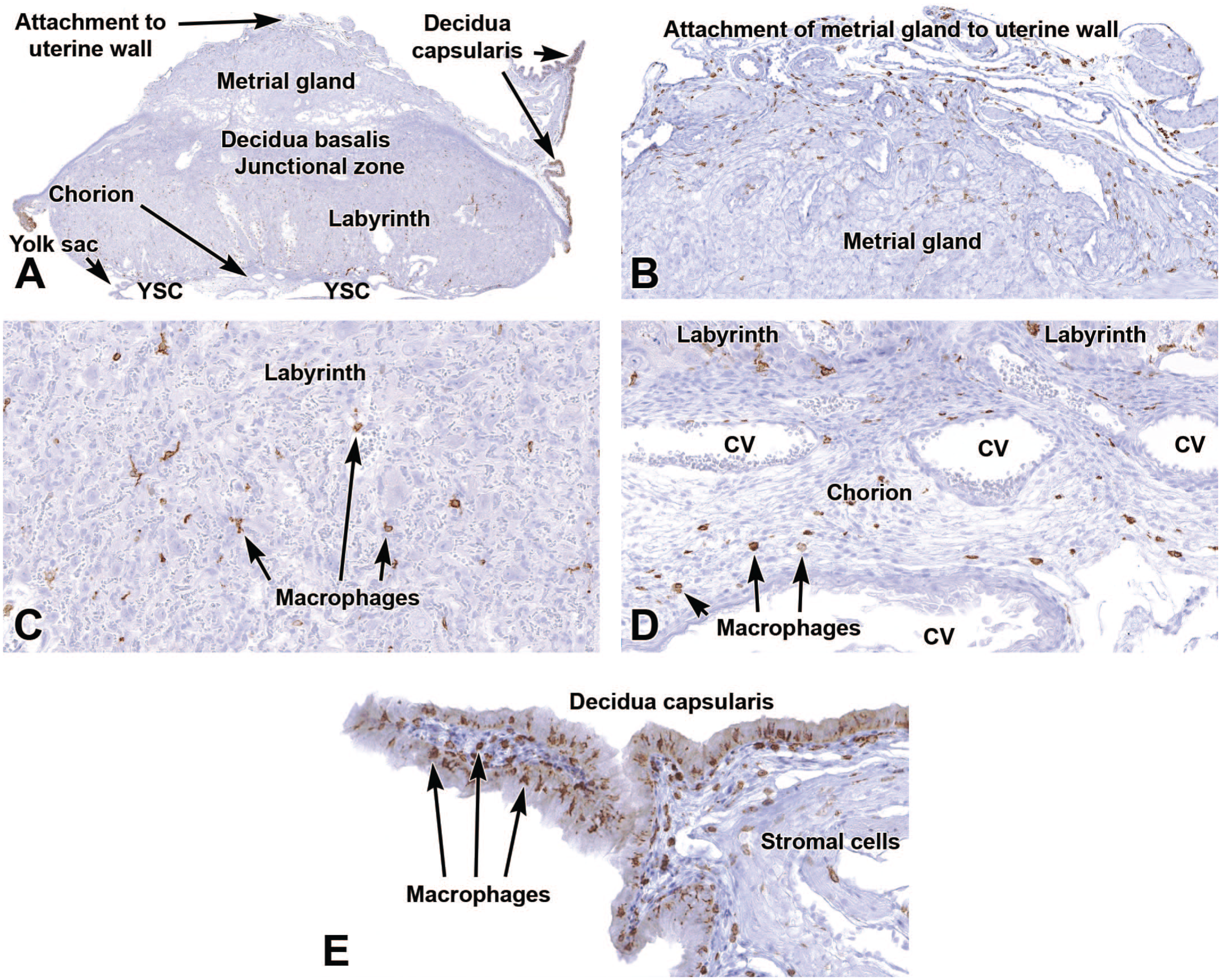
Representative images of the regressing definitive placenta near term (E18.5) showing a shift in F4/80 distribution as gestation progresses. Compared to E9.5 (see Figure 15), F4/80 expression is highest in the decidua capsularis (Panels A and E) while labeled cells are also evident at the interface between the metrial gland and uterine wall (Panel B) as well as in the labyrinth and chorion (Panels C and D). Panel E shows that the macrophages in the decidua capsularis are present within both the epithelial lining and the subjacent stroma. CV = chorionic blood vessels, YSC = yolk sac cavity. Immunohistochemistry using 3,3’-diaminobenzidine (DAB [brown color]) as the chromogen and hematoxylin as the counterstain. Original objective magnification: 2x (Panel A), 10x (Panel B), 20x (Panel C), 20x (Panel D), 19.5 (Panel E).
In mice, the main markers for placental macrophages are F4/80 (Figures 15 and 16) and ionized calcium-binding adaptor molecule 1 (IBA1, also known as allograft inflammatory factor-1 [AIF-1]; Figure 17). 77 Both markers can be co-expressed in the same cells, particularly in activated macrophages. F4/80 is a cell surface glycoprotein of the epidermal growth factor-seven transmembrane (EGF-TM7) family used to detect mature macrophages. 69 IBA1 is a “pan-macrophage marker” (except for alveolar macrophages 77 ) localized in both the cytoplasm and nucleus, where it is thought to control cell motility and phagocytic activity. 99 While both markers are reliable, F4/80 is considered to be the more specific marker for tissue-resident macrophages, while IBA1 is preferred for detecting activated cells. 99
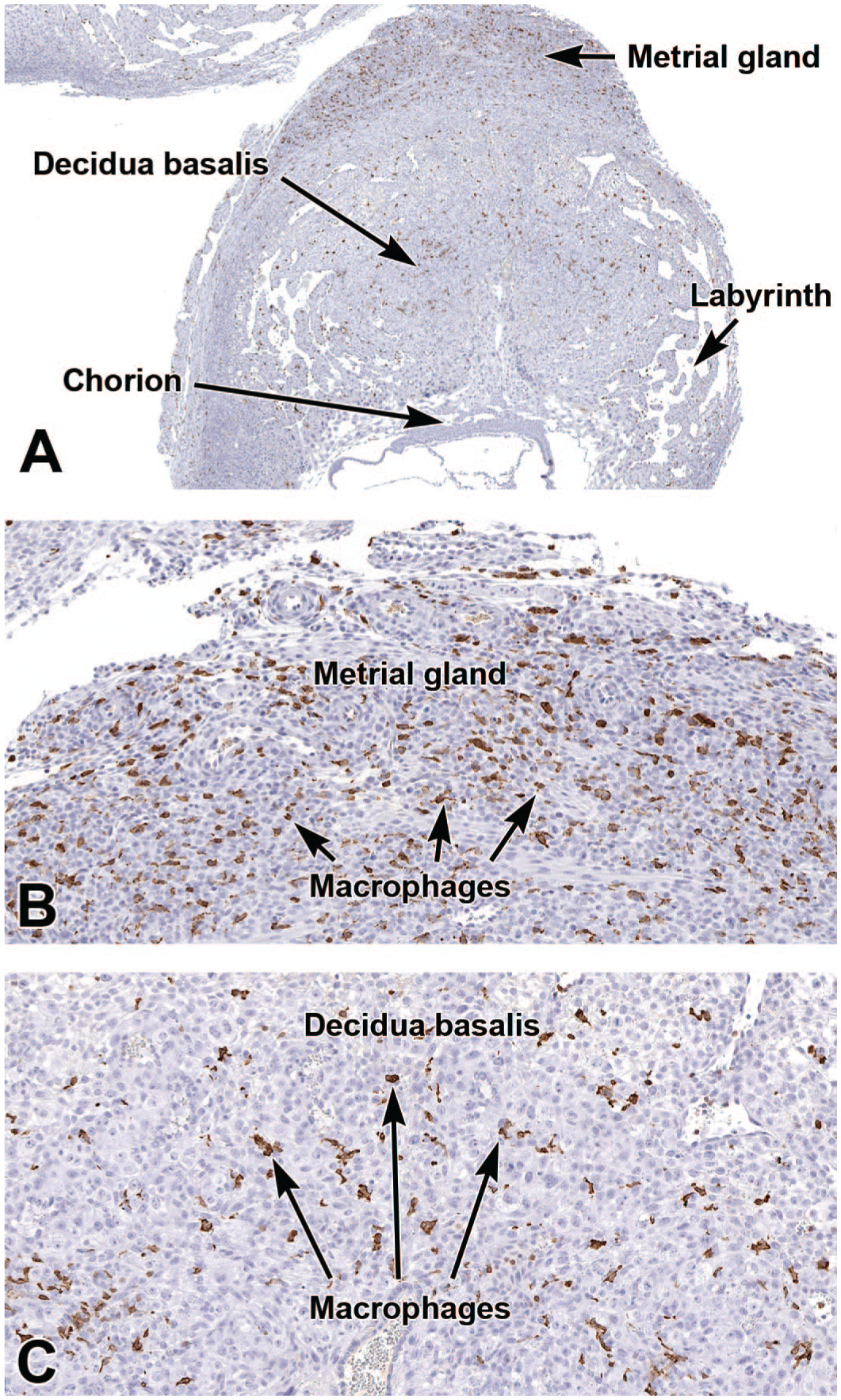
Representative images of an early placenta (E8.5) showing expression of the macrophage marker ionized calcium-binding adaptor molecule 1 (IBA1). The IBA1-positive macrophages are concentrated in the metrial gland (Panels A and B) and are widespread in the decidua basalis, but with fewer numbers (Panel C). Immunohistochemistry using 3,3’-diaminobenzidine (DAB [brown color]) as the chromogen and hematoxylin as the counterstain. Original objective magnification: 3x (Panel A), 15x (Panel B), and 17x (Panel C).
Additional Markers for Placental Cell Populations and Processes
Other markers may also be utilized to investigate placental cells and processes. In general, these options are not specific to particular placental cell populations but are antigens expressed more widely (including in embryonic and adult tissues). These biomarkers employ IHC to localize the cell and tissue distribution of distinct antigens involved in particular cell processes.
Caspase 3
Caspase 3 is one of many cysteine proteases involved in a signaling cascade that mediates apoptosis (programmed cell death), which is an essential process in remodeling embryonic organs throughout development.48,92,115 Apoptosis can occur via multiple pathways, all of which depend on caspase 3 as the main “executioner” enzyme to drive the final steps culminating in cell death.48,73 The central placement of caspase 3 in the cell death signaling cascade accounts for its common use as an apoptosis marker.
In placental development, apoptosis is involved in remodeling multiple layers throughout gestation. Apoptosis during decidualization strikes many endometrial cells following blastocyst implantation.4,23,55 These apoptotic decidual cells are phagocytized by trophoblasts to clear room for placental expansion and embryonic growth. 106 Endothelial cell apoptosis is essential in spiral artery remodeling within the decidua basalis and metrial gland. 9 Caspase 3 is expressed in the uterine luminal epithelium at and around the implantation site between E5.5 and E6.5. 73 From the time of implantation to around E9.5, the rate of apoptosis is less than that of proliferation in the decidua, thus leading to some growth of this region. However, after E9.5, decidual proliferation declines, which results in a gradual reduction in decidual volume as gestation proceeds due to continued cell death. 55 At E18.5, caspase 3 is most prominent in the maternal decidua, with some minute presence in the embryonic placenta and amnion. 92 This near-term caspase 3 expression pattern indicates that placental apoptosis is not a critical process in parturition. That said, aberrant embryonic and/or placental apoptosis may result in complicated or unsuccessful pregnancies. 46
Galectin-3
Galectin-3 (previously referred to as macrophage surface antigen-2 [Mac-2]) is a marker of decidual tissue near the time of implantation and throughout decidualization. 112 This molecule is a β-galactoside-binding lectin with speculated functions related to cell-cell interactions during implantation due to its increased expression in the endometrium and trophoblasts.81,100,158 However, the exact mechanisms by which galectin-3 modulates such interactions have yet to be defined.76,81,100,112,158
Galectin-3 is expressed chiefly in the endometrial lining,9,112 in minute amounts prior to implantation, but in modestly increased quantities by E5.5 in some endometrial cells abutting the implantation site and newly forming placenta. 112 Within the endometrium, galectin-3 expression occurs in a structure- and time-dependent manner, peaking in the glandular epithelium at E2.0 to E4.0 and in luminal epithelium at E6.0 to E8.0. 40 By E8.5, decidual tissue more prominently expresses galectin-3 while the outer (nondecidualized) endometrium and embryonic tissues (e.g., the ectoplacental cone [nascent placenta] and embryo) do not. 112 At E12.5, galectin-3 expression continues in the decidua but is expanded to include extensive distribution, marking the majority of trophoblasts in the junctional zone and labyrinth. 112 This marker is greatly reduced in the endometrium after birth. 112
Galectin-3 has also been demonstrated in uNK cells later in gestation, specifically around E15.5.76,81 Due to the presence of uNK cells in the metrial gland and decidua basalis, galectin-3 is visible in these locations. 76
Achaete-Scute Complex Homolog 2
The transcription factor Achaete-Scute Complex Homolog (ASCL2) is critical for regulating differentiation of multiple cell populations in the hemochorial placenta, especially various trophoblast lineages. As discussed above, this marker occurs in the ectoplacental cone, spongiotrophoblast layer, and chorion from E8.5 to E12.567,148 but thereafter is primarily expressed in spongiotrophoblast within the junctional zone.47,60 ASCL2 in the ectoplacental cone is required for regulating TGC differentiation, but its overexpression inhibits further giant cell differentiation.67,130 The need for ASCL2 in early placental development is highlighted by the progressive degeneration of Ascl2 null mutant embryos during early gestation: at E7.5, the morphology of the ectoplacental cone and chorion shows no distinction when compared to controls, but by E8.5 the ectoplacental cone no longer supports trophoblast proliferation. 60 The pan-placental deficit in trophoblast production results in chorion compaction and reduced labyrinth vascularity, 60 which together produce death of the embryo by ~E9.5.
Vascular Cell Adhesion Protein-1
Vascular Cell Adhesion Protein-1 (VCAM-1) is a member of the immunoglobulin superfamily that, among other functions, has an essential role in chorioallantoic fusion (E8.5) and evolution of the definitive placenta. Contact of the allantois (the embryo-derived umbilical cord precursor) with the chorion is essential in promoting placental vasculogenesis (the process of forming new blood vessels de novo from pluripotent mesenchymal stem cells), angiogenesis (the process of forming new blood vessels from existing ones), and vascular remodeling. Expression of VCAM-1 in the allantoic mesothelium occurs at low levels when this tissue is initiated, but expression peaks between E7.5 and completion of chorioallantoic fusion at E8.5.44,61 VCAM-1 is mainly located at the distal allantoic end,44,70 primarily within the core of the tissue. 45 Interaction of VCAM-1 on the allantois with an α4ß1 integrin receptor (also known as very late antigen-4 [VLA-4]) that is constitutively expressed on the chorion surface 70 mediates the cell adhesion that allows fusion to take place. Expression of both VCAM-1 and this 4-integrin is time-dependent, and as such so is the process of chorioallantoic fusion. 35 Interestingly, in Vcam-1 knockout mouse embryos, chorioallantoic attachment still occurs normally approximately 30% of the time, indicating that other cell adhesion molecules are capable of facilitating this process, albeit by less efficient means. 35
Conclusions
The placenta undergoes extensive reorganization throughout the course of gestation, reflecting successive waves of cell differentiation necessary to produce the stereotypical form and function of the maternal and embryonic placental tissues. Disturbances in cell differentiation or numbers of seemingly minor importance may nonetheless have catastrophic effects on placental growth, which in turn may lead to restricted intrauterine growth, malformations, or even death of the embryo. Determining the placental contribution to such adverse pregnancy outcomes may begin with conventional macroscopic observation and microscopic evaluation of routinely prepared (H&E-stained) tissue sections, but in many cases a more detailed investigation to examine one or multiple biomarkers of particular placental cell lineages will be necessary to generate a more definitive diagnosis.
The current paper provides an overview of recognized histochemical and IHC methods that have utility in performing such cell-type-specific placental assessments. Appropriate implementation of these methods for particular subanatomic locations and for one or usually several time points during development can substantially enhance the likelihood that the target cell population(s) will be identified. This information, in turn, may provide clues regarding the potential pathogenesis of placenta-related pathology contributing to altered embryo viability. The authors trust that this primer will provide pathologists faced with developmental biology and developmental toxicology problems with a suitable starting point for formulating a rational study design and performing a flexible but rigorous pathology evaluation.
Supplemental Material
sj-docx-1-tpx-10.1177_01926233261436343 – Supplemental material for Select Histochemical and Immunohistochemical Methods to Investigate the Cell and Tissue Composition of the Mouse Placenta and Metrial Gland
Supplemental material, sj-docx-1-tpx-10.1177_01926233261436343 for Select Histochemical and Immunohistochemical Methods to Investigate the Cell and Tissue Composition of the Mouse Placenta and Metrial Gland by Susan A. Elmore, Robert Z. Cochran, Charan Ganta, Heather Jensen, Beth A. Lubeck, Beth Mahler, David Sabio, Greg Stamper and Brad Bolon in Toxicologic Pathology
Footnotes
Acknowledgements
We would like to give special thanks to Drs. Jerrold Ward (Global VetPathology), Priyanka Thakur (NIEHS), and Sonika Patial (NIEHS) for their careful reviews and edits.
Dedication
This paper is dedicated to David Sabio, our regular coauthor and friend, whose remarkable talent, creativity, and collaborative spirit helped shape the multiple papers in the Histology Atlas of the Developing Mouse Embryo series published in Toxicologic Pathology over the last decade. His contributions were invaluable and have left a lasting mark on the developmental pathology field. He will be greatly missed. David Sabio (1972-2026).
Authors’ Note
This manuscript is one of a series of mouse developmental atlases that include (in order of publication) the heart, 128 hepatobiliary system, 32 hepatobiliary hemolymphatic vascular system, 147 central nervous system, 24 urinary system, 50 respiratory system, 108 placenta, 49 and digestive system. 85
Author Contributions
Declaration of Conflicting Interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article. Toxicologic Pathology: BM serves as the journal’s Illustrations Editor, and BB serves as a senior advisor for the journal’s Editor. Neither BB nor BM took part in the peer review or decision-making processes for this paper.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported (in part) by the US National Institutes of Health (NIH), specifically the US NIEHS, via NIH Intramural Research project ZIC ES103376-03.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.