
Abstract
Objective
Mannose-binding lectin is an important component of innate immunity; it initiates the lectin pathway of complement activation critical for innate immunity. Failure of local innate defenses may result in defective responses that lead to the persistent carriage of microorganisms or ongoing inflammation. This study investigated the role of mannose-binding lectin levels and the frequency of the 6 functional mannose-binding lectin polymorphisms in Turkish individuals with nasal polyposis.
Study Design
A case-control study.
Setting
University hospital.
Subjects and Methods
Fifty-one patients with nasal polyposis and 53 healthy controls were enrolled. Serum mannose-binding lectin levels were obtained by enzyme-linked immunosorbent assay (ELISA) using the mannose-binding lectin oligomer ELISA kit. Mannose-binding lectin 2 genotyping was performed by isolating the genomic DNA from leukocytes.
Results
Mean mannose-binding lectin levels were 1693.2 and 1887.8 in the patient and control group, respectively. Although mannose-binding lectin levels were lower in the patient group, the difference was not statistically significant (P > .05). No overall association was observed between the mannose-binding lectin genotype and susceptibility to nasal polyposis (95% confidence interval = 0.716–4.389, odds ratio = 1.773). The mutant allele frequencies of the 3 structural polymorphisms did not differ significantly between the nasal polyposis patients and the controls (P = .659).
Conclusions
Mannose-binding lectins are not involved in the pathogenesis of nasal polyposis in adult Turkish patients, but additional research is needed for further comment.
Chronic inflammation of the nasal cavity and paranasal sinuses may result in the formation of nasal polyposis. Given that this is a recalcitrant disease, medical therapy is usually not enough to manage nasal polyposis, and multiple surgical procedures are required. Although staphylococcal superantigens and hypersensitivity to ubiquitous fungi are important in the pathogenesis of the inflammation, 1 the exact etiopathogenesis has not been fully elucidated. In the literature, studies have shown the linkage of certain phenotypes of nasal polyps to candidate gene polymorphisms. A significant association was identified between human leukocyte antigens (HLA) and nasal polyps. Patients with certain HLA haplotypes have shown an increased risk of developing nasal polyps.2,3 However, ethnic background may have an influence on these results. Also, there have been several reports indicating that advocate inflammation in nasal polyps may be related to certain genes and single-nucleotide polymorphisms. 3
Mannose-binding lectin (MBL) is a serum protein produced in the liver but also found at sites of inflammation, albeit at levels substantially less than in the blood. MBL (also known as mannan-binding lectin) is involved in the first line of defense. MBL initiates the lectin pathway of complement activation, which is a critical component of innate immunity against a number of clinically relevant pathogens. MBL is important in opsonophagocytosis, inflammation, and complement activation. 4
MBL levels among populations have been shown to be quite variable. Low levels of MBL are associated with susceptibility to recurrent childhood infections. 5
MBL is encoded by 2 separate genes: MBL1 and MBL2. In humans, MBL1 is a pseudogene. It is known now that the mean serum MBL level in an individual is genetically determined by 3 single-nucleotide substitutions in exon-1 of the human MBL2 gene, 6 located at codons 52, 54, and 57, also referred as D, B, and C mutations; the wild-type allele is referred to as A. These mutations result in amino acid substitutions in the collagen-like domain, leading to significant decreases in the functional serum MBL level. 7 In addition, 3 pairs of allelic dimorphisms can occur in the downstream promoter and the 5′-untranslated region of the MBL2 gene at positions −550, −221, and +4. Of these, the −221 polymorphism has the greatest effect on MBL serum concentrations; the allele that gives high MBL expression is termed Y, and the allele giving low expression is termed X. In contrast to the structural (A/B, C, D) and −221 (Y/X) polymorphisms, the functional effects of −550 (L/H) and −221 (P/Q) polymorphisms appear to be relatively minor.
This exploratory study evaluated the correlation of nasal polyposis with MBL polymorphisms and MBL levels. We investigated 3 promoter-region mutations (–550G>C, −221G>C, and +4C>T) and mutations in codons 52, 54, and 57 in the MBL2 gene. We hypothesized that low levels of MBL and specific polymorphisms might be associated with a defective immune response to Staphylococcus aureus and certain fungi and that a defective immune response may trigger the cascade that leads to nasal polyposis.
Materials and Methods
Patient Selection
Fifty-one patients with nasal polyposis and 53 volunteers were enrolled. The groups were matched for ethnic background. The patient group was recruited from the Otolaryngology Department, Ege University Hospital, Turkey, during 2006 and 2007. Patients diagnosed with bilateral nasal polyposis by biopsy who had no upper respiratory system infection and no history of allergic rhinitis or asthma were selected. All control patients were examined by nasal endoscopy to exclude any sinus disease after obtaining a detailed anamnesis. Two separate blood samples were collected to measure MBL blood levels and to detect MBL polymorphisms. Because MBL is an acute-phase reactant, blood samples were not collected from patients with an infectious disease. All blood samples were collected prior to medication (eg, oral corticosteroids) so as not to affect the immune system.
Fasting peripheral venous blood samples were collected from the antecubital vein using Vacutainer tubes containing EDTA. They were stored at −80°C until analysis. MBL serum levels were determined by enzyme-linked immunosorbent assay (ELISA) using the MBL oligomer ELISA kit (Antibodyshop, Gentofte, Denmark) according to the manufacturer’s protocol.
Informed consent was obtained from all participants. The study was approved by Ege University Local Ethics Committee.
MBL2 Genotyping
Genomic DNA was isolated from leukocytes. Genotyping was conducted using the INNO-LiPA MBL2 assay (Innogenetics, Gent, Belgium). In total, 51 patients and 54 control samples were tested; for all samples, both haplotypes were identified.
The polymorphisms are localized in the promoter and 5′ untranslated region (–550G>C, −221G>C, and +4C>T) and affect the expression of the MBL2 gene.
The INNO-LiPA MBL2 assay is a multiparameter assay allowing simultaneous detection of the 6 relevant DNA variants in the MBL2 gene (–550G>C, −221G>C, +4C>T, and codons 52, 54, and 57) using reverse hybridization technology. DNA was isolated from anticoagulated blood (InVitek Blood Kit, Berlin, Germany). The entire duration from blood sampling to the identification of mutations required less than 6 h, and hybridization/detection was carried out manually.
SPSS 16 software was used for statistical analysis. MBL allele/genotype frequencies were compared by contingency table analysis using the χ2 test. The Fisher exact test was used when results were <5. Results were considered statistically significant if P < .05. The univariate analysis of variance test was used to examine relationships between MBL plasma concentrations and MBL2 genotypes in the patient and control groups. When a significant difference was found, individual means were compared using Tukey test. The MBL levels of the patient and control groups were compared with the Student t test. Differences were considered significant when P < .05, and the 95% confidence intervals (CIs) are given. A compromise power analysis for χ2 contingency tables was used (G*Power 3.1.3). The power of the χ2 contingency tables was 0.832 (effect size w = 0.3, df: 5).
Results
Patient age, sex, and MBL levels are presented in Table 1 . In the patient group, the mean age was 46.3 years, and in the control group, the mean age was 33.7 years. The patient group consisted of 29 men and 22 women, and the control group included 33 men and 20 women.
Mean Age and Mannose-Binding Lectin (MBL) Levels in the Patient and Control Groups

The mean MBL levels were 1693.2 and 1887.8 in the patient and the control group, respectively ( Table 1 ). Although MBL levels were lower in the patient group, the difference was not statistically significant (P > .05). The correlation between MBL concentration and MBL2 genotype is shown in Figure 1 (P > .05).
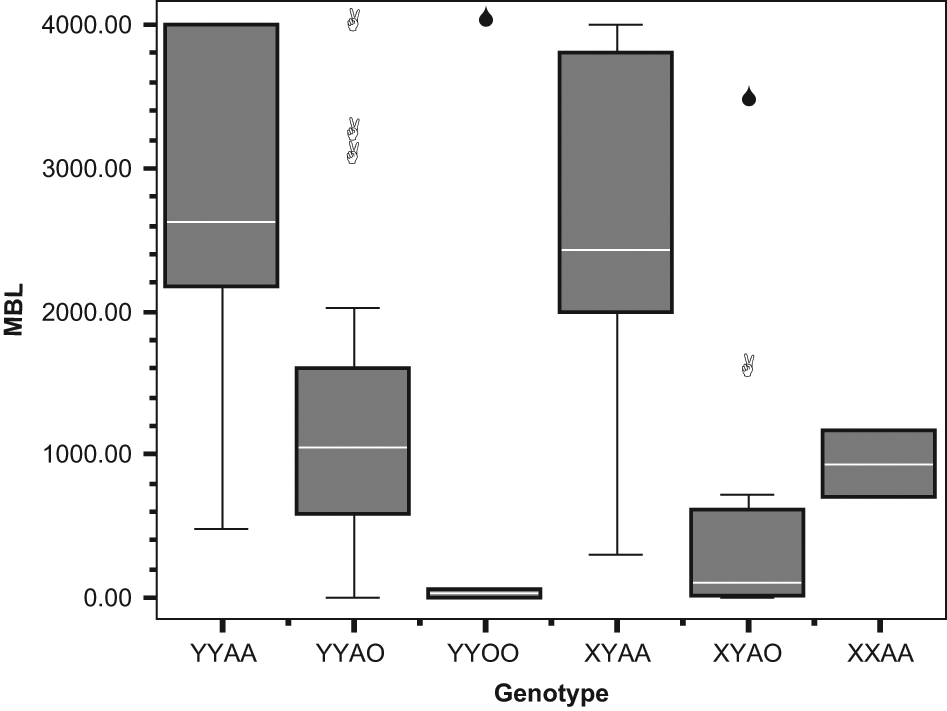
A box plot graph showing concentration of circulating mannose-binding lectin (MBL; ng/µL) and corresponding MBL2 genotypes.
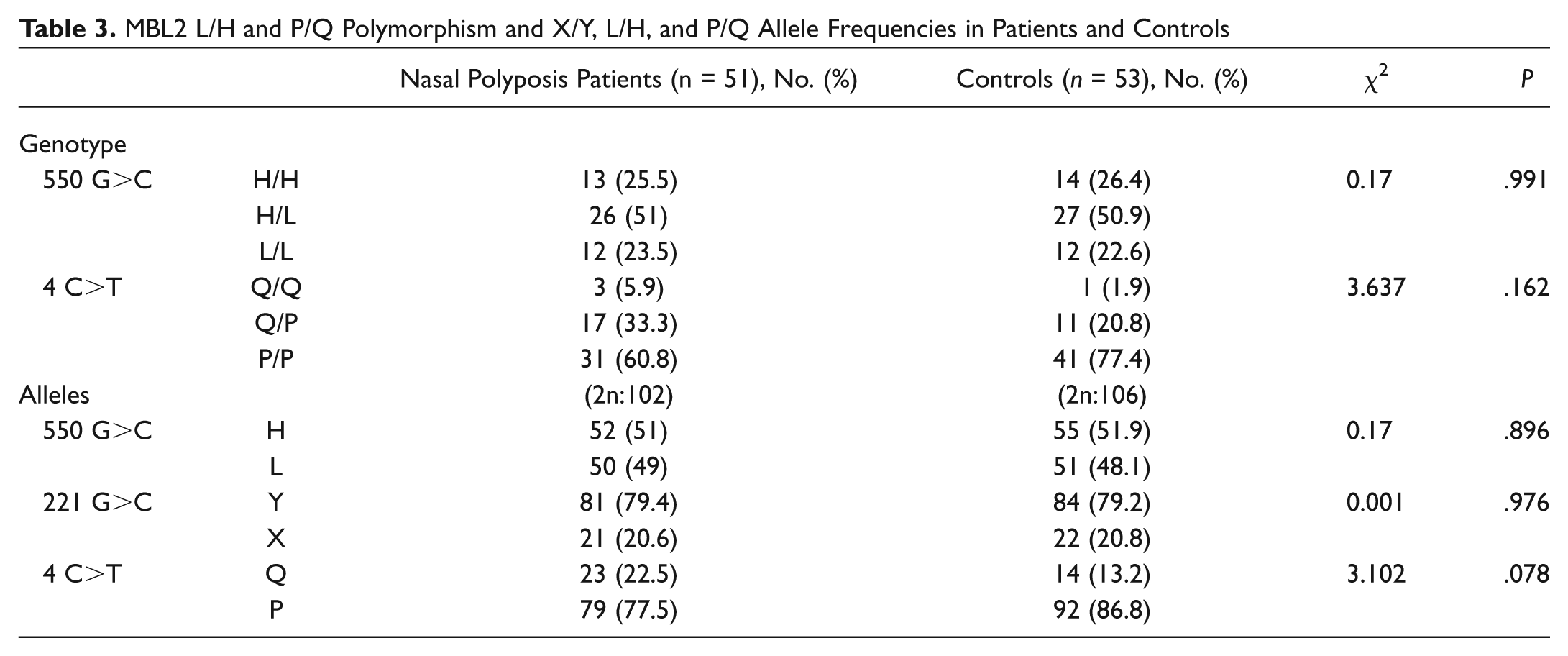
None of the MBL2 polymorphisms were individually associated with susceptibility to nasal polyposis. In particular, the mutant allele frequencies of the 3 structural polymorphisms did not significantly differ between nasal polyp cases and the controls (D allele frequency 0.088 in cases and 0.113 in controls, 2 × 2 χ2 = 0.357, P = .550, odds ratio [OR] = 1.319, CI = 0.531-3.279; B allele frequency 0.157 in cases and 0.189 in controls, 2 × 2 χ2 = 0.368, P = .544, OR = 1.250, CI = 0.607-2.574; C allele frequency 0.01 in cases and 0.009 in controls, 2 × 2 Fisher exact test, P = 1, OR = 0.962, CI = 0.059-15.586). The functional level of MBL is not determined by individual polymorphisms but by the combinations of different alleles. In particular, the structural polymorphisms (A/O) and the −221 promoter polymorphism (X/Y) exert the strongest effects on MBL function. Individuals who are A/A (ie, do not have MBL structural variant alleles) or YA/O (heterozygotes for a single structural variant allele, ie, A/B, A/C, or A/D) are functionally normal, whereas individuals who have the genotypes O/O (homozygotes for variant alleles B/B, C/C, or D/D, or compound heterozygotes, ie, B/C) or XA/O (heterozygous for the X promoter allele and a structural variant allele) are functionally deficient. The functional consequences of XA homozygosity are controversial, although some studies have considered this to be a deficient state. Taking this approach, the results of MBL functional status in the nasal polyposis and control groups are presented in Table 2 . No overall association was observed between MBL genotypic deficiency (defined on the basis of B, C, D, and X) and susceptibility to nasal polyposis (2 × 2 χ2 = 1.552, P = .213, OR = 1.773, CI = 0.716-4.389; Table 2 ). The functional consequences of the L/H and P/Q polymorphisms are less well defined, and there was no apparent association with susceptibility to nasal polyposis ( Table 3 ).
Mannose-Binding Lectin (MBL) Functional Status (–221, 52, 54, and 57 Polymorphisms) in Patients and Controls a
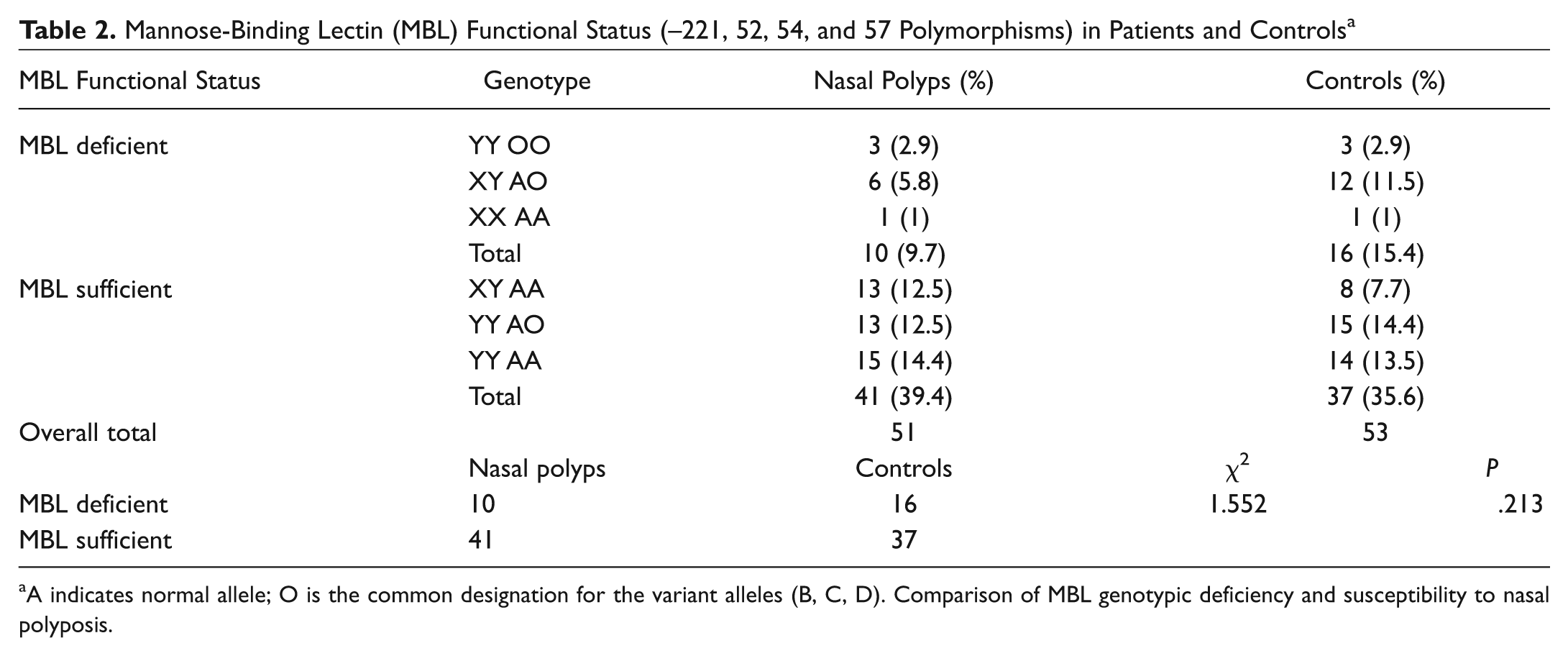
A indicates normal allele; O is the common designation for the variant alleles (B, C, D). Comparison of MBL genotypic deficiency and susceptibility to nasal polyposis.
MBL2 L/H and P/Q Polymorphism and X/Y, L/H, and P/Q Allele Frequencies in Patients and Controls
Discussion
Innate immunity may hide clues to elucidate the underlying pathology of nasal polyposis. The actions of the innate immune system and the cross-talk with the slower but longer lasting adaptive immune system enable pathogen clearance, restoration of mucosal homeostasis, and the prevention of ongoing inflammation; the failure to restore homeostasis is believed to result in chronic inflammation. 8 Improper activity of some components of nasal innate immunity have been implicated in chronic sinusitis with or without nasal polyposis. Components of innate immunity can organize adaptive immunity through T- and B-cell activity. 9 Defective responses lead to persistent carriage of some microorganisms or ongoing inflammation.
Toll-like receptors (TLRs), a component of innate immunity, have been investigated for their possible role in nasal polyps, and several authors have reported a possible relationship. 10 TLRs may act as early sensors for the presence of viral, bacterial, or fungal pathogens in the nasal airway and may play a sentinel role in the activation of local inflammation through the expression of innate immune mediators such as the complement cascade. Ip et al 11 demonstrated that MBL can cooperate with TLR2 in sensing S aureus and suggested that the cooperation between the receptors may be important for coordinating the appropriate host response. MBL is one of the important components of the main pathways of the complement activation cascade. Failure of local innate defenses may result in microbial colonization, leading to recurrent infections. 12 Low MBL levels and certain MBL polymorphisms that result in low levels may be responsible for misdirected immune activity against certain pathogens, including S aureus and fungus, and this misdirected reaction leads to a series of events that trigger nasal polyp formation.
Superantigen-producing strains of S aureus activate T-helper cells, leading to severe inflammation, including polyclonal activation of B cells and recruitment of eosinophils. An increased colonization rate of S aureus in nasal polyps, but not in chronic sinusitis without nasal polyps, was recently reported. 13 Genetic polymorphisms in the MBL gene are known to be associated with susceptibility to infectious disease. The lectin-dependent complement-activation pathway contributes significantly to the opsonophagocytosis of S aureus. 14 MBL haplotypes may be responsible for persistent carriage of S aureus in the nasal cavity. 15
MBL is also important in the immune response directed against fungus. Low levels of MBL in patients who have had allergenic stem cell transplantations leads to invasive fungal infections. 16 Kaur et al 17 reported that nucleotide mutations associated with high MBL serum levels may have a role in increasing disease activity in patients with bronchial asthma with allergic rhinitis and allergic bronchopulmonary aspergillosis.
Asthma sufferers are predisposed to develop nasal polyposis. 18 Although the exact interaction is not fully understood, they may share particular pathophysiological mechanisms. A recent study by Aittoniemi et al 19 showed that the carriage of a promoter region variant, allele X, which causes low MBL expression, is a significant risk factor for asthma in males.
In this study, we have investigated the role of certain mutations that affect the blood levels of MBL, located in exon 1, and promoter regions and their possible correlation in nasal polyposis. As low MBL levels and MBL haplotypes may be responsible for the persistent carriage of S aureus in the nasal cavity and increased nasal fungal infection, we expected to find lower MBL levels in patients with nasal polyposis and an increase in the allelic/genotypic frequencies of MBL-lowering mutations.
The mean MBL blood levels of the patient group were found to be lower than those of the control group (1692.29/1887.8), but this difference was not statistically significant (P > .05). However, a recent study conducted in adult Chinese patients reported increased MBL levels in chronic rhinosinusitis patients both with and without nasal polyps. Moreover, MBL levels were found to be significantly higher in chronic rhinosinusitis (CRS) patients with nasal polyps than in CRS patients without nasal polyps, and MBL levels were positively correlated with the extent of the disease seen on computed tomography, via endoscopy, and through peripheral eosinophil count. 20 The authors did not, however, investigate MBL mutations.
In certain populations, a functional MBL deficiency may be advantageous. Given that MBL levels differ among populations, 21 the discrepancy in the MBL between our results and those of Cui et al 20 may indicate genetic differences in MBL expression between Chinese and Turkish populations.
Polymorphisms in codons 52, 54, and 57 and dimorphisms in promoter regions −221, −550, and +4 were studied. To date, no study has investigated the possible role of these 6 polymorphisms in nasal polyposis; their effect on MBL level is variable. Mutations in exon 1 tend to lower MBL levels more than do mutations in promoter regions.
In this study, we did not identify a correlation between the serum levels of MBL in CRS with nasal polyps and healthy controls. The most likely reason is that the serum levels do not reflect the MBL levels in the nasal mucosa. It may be necessary to investigate the MBL levels of the nasal polyp biopsies together with the serum of the patients and correlate the results according to the persistence of fungus and/or S aureus in the nasal mucosa. In addition, because of the relatively small sample size, a type 2 error should be always kept in mind.
Allele frequencies of codons 52, 54, and 57 and promoter regions −221, −550, and +4 were not found to be statistically different between patient and control groups, and the genotypic deficiency in MBL was not associated with susceptibility to nasal polyposis. However, these results should be interpreted with caution because of the relatively small sample size of the study group and the possibility of a type 2 error.
Our study was an exploratory study of the pathogenesis of nasal polyposis. Given the relatively small sample size, we might have missed clinically significant findings. In addition, given that CRS with nasal polyps is not a homogeneous disease, studies involving patients with allergic fungal sinusitis with Samter triad are needed. Additional research with a larger sample size is needed for further comment.
Conclusion
Serum MBL levels and MBL2 gene mutations (codons 52, 54, 57 and promoter regions −221, −550, and +4) do not play a role in nasal polyposis pathophysiology in adult Turkish patients.
Author Contributions
Disclosures
Footnotes
Acknowledgements
This article was presented at the 4th Turkish National Rhinology Congress 2008 in Antalya, Turkey, and was selected as the winner in the oral presentation.
Sponsorships or competing interests that may be relevant to content are disclosed at the end of this article.