
Abstract
Objective
Allergic fungal rhinosinusitis (AFRS) is a clinical subtype of chronic rhinosinusitis with nasal polyps (CRSwNP), characterized by eosinophilic mucin, evidence of fungal elements within the mucin, fungal-specific type I hypersensitivity, and characteristic computed tomography findings. It remains controversial whether AFRS represents a disease with a unique pathophysiology from chronic rhinosinusitis or is merely a severe form of CRSwNP. The goal of this study was to identify molecular features unique to AFRS.
Study Design
Cross-sectional case-control.
Setting
Single academic tertiary referral institution.
Subjects and Methods
Subjects included 86 patients undergoing endoscopic sinus surgery: CRSwNP (n = 34), AFRS (n = 37), and healthy controls (n = 15). Pathway and correlation analyses were performed with whole-genome microarray data for study patients undergoing surgery for recalcitrant chronic rhinosinusitis. Our findings were confirmed with quantitative polymerase chain reaction and immunohistochemical studies.
Results
AFRS was uniquely characterized by a pronounced association with adaptive T helper 2–associated immune gene expression. AFRS exhibited altered expression of proteins associated with secretory salivary peptides—namely, histatin, a peptide with known antifungal activity in the oral cavity. Furthermore, the expression of histatins correlated negatively with that of type 2 inflammatory mediators. We confirm the decreased expression of histatins in AFRS when compared with CRSwNP by quantitative polymerase chain reaction and localized its expression to a submucosal cell population.
Conclusion
There exist clear molecular profiles that distinguish AFRS from CRSwNP. This divergence translates into an altered ability to control fungal growth and may in part explain some of the phenotypical differences between CRSwNP and AFRS.
Keywords
Chronic rhinosinusitis (CRS) is an inflammatory condition of the paranasal sinuses comprising several heterogeneous subtypes, and it still represents a challenge for the treating physician. Indeed, our understanding of the molecular pathophysiology that drives the expression of disease phenotypes in CRS is limited, hindering efforts aimed at developing treatment strategies that are deliberate and effective. More research is needed that highlights different molecular pathways driving disease phenotypes in CRS, thereby supporting the development of a classification scheme that employs endotypes.1,2 Endotyping has advanced the development of biological therapies that target disease-specific pathways in asthma subtypes. The same biological therapies are now being evaluated in clinical trials of patients with CRS; however, endotyping in CRS remains incomplete, and the application of novel therapies in CRS without regard to disease endotypes may obscure outcomes. Ideally, endotyping should enable a scenario where molecular data can be leveraged to deliver more effective, personalized medicine to patients with CRS.
Disease endotyping in asthma and CRS employs canonical type 2 inflammatory markers as indicators of disease burden in many instances. Studies by Tyler et al demonstrated that the allergic fungal rhinosinusitis (AFRS) phenotype represents an exaggerated type 2 inflammatory disease state. 2 AFRS represents a clinical subtype of CRS, accounting for 15% of all CRS cases, and it is differentiated by criteria initially proposed by Bent and Kuhn over 20 years ago.3,4 The AFRS subtype thus represents an ideal disease model to study molecular alterations that may contribute to disease burden. Unfortunately, few studies have attempted to systematically describe molecular features that differentiate AFRS from other subtypes, and thus far, the available studies have been limited in sample size and scope. In this study, we employ a top-down approach using large-scale microarray-based gene analysis to elucidate molecular differences among subtypes. To the best of our knowledge, we present the largest molecular profiling study to date that highlights unique gene changes occurring in the AFRS subtype.
Methods
Patients
Patients undergoing medically indicated functional endoscopic sinus surgery consented to having sinonasal tissue, which was removed as a standard of their surgery, collected and analyzed for gene expression. The Institutional Review Board at the University of Texas Health Science Center at Houston approved the study protocol.
Patients were grouped into CRS with nasal polyps (CRSwNP), aspirin-exacerbated respiratory disease (AERD), CRS without nasal polyps (CRSsNP), or AFRS according to criteria set forth in the European position paper on rhinosinusitis and nasal polyps. 4 Polyps documented by an ear, nose, and throat physician on nasal endoscopy at any time categorized a patient as a CRSwNP subtype. Patients with CRS and nasal polyps were diagnosed with AFRS if there was evidence of eosinophil-rich mucus with noninvasive fungal hyphae, hypersensitivity to fungi, and characteristic radiographic findings. 3 AERD was characterized by the presence of asthma, the presence or documented history of nasal polyps, and history of intake of aspirin or nonsteroid anti-inflammatory drugs inciting worsening respiratory symptoms. Patients classified as asthmatic had a prior diagnosis of asthma by a pulmonologist, allergist, and/or positive pulmonary function testing. In this study, CRSwNP excludes patients with AERD and AFRS. Healthy controls (HCs) had no history of atopy or asthma symptoms and were undergoing sinus surgery as a component of a surgical approach for removal of a benign pituitary lesion or repair of anterior skull base cerebrospinal fluid leak. Table 1 delineates the number of patients enrolled according to each clinical subtype with relevant clinical factors (see also, Supplemental Table S1, available in the online version of the article).
Clinical Characteristics of Enrolled Participants. a
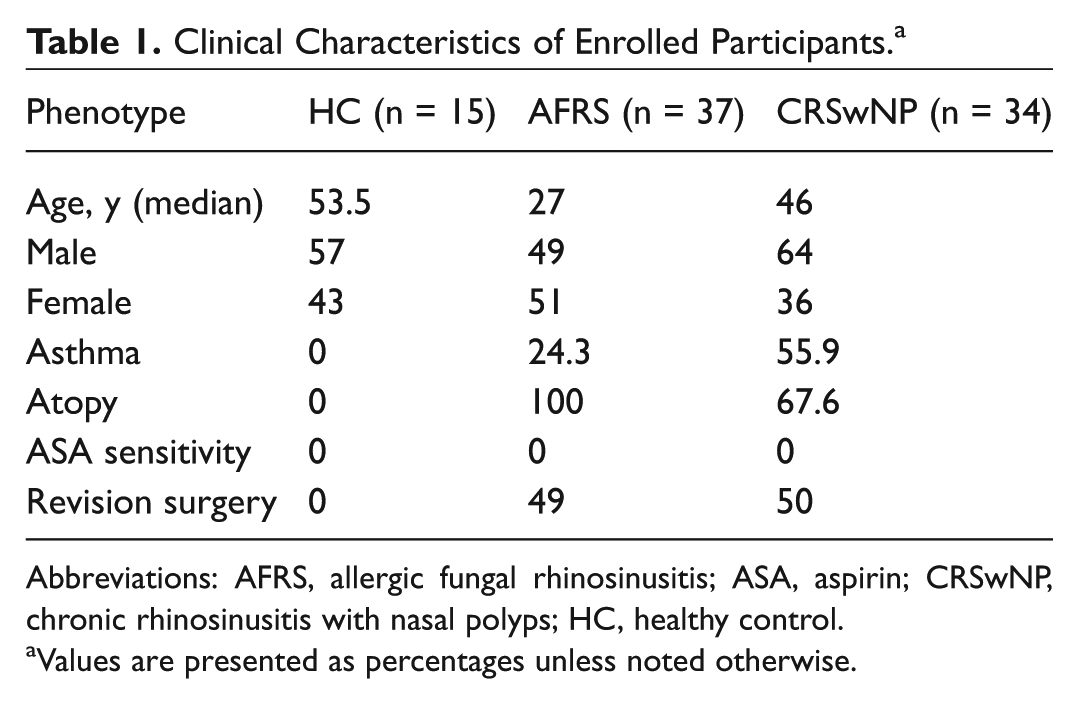
Abbreviations: AFRS, allergic fungal rhinosinusitis; ASA, aspirin; CRSwNP, chronic rhinosinusitis with nasal polyps; HC, healthy control.
Values are presented as percentages unless noted otherwise.
No clinical subtype was treated in an exclusive manner in that patients did not receive any specific medical or surgical therapy that deviated from the standard of care delivered for other subtypes. All topical and systemic corticosteroids were withheld at least 4 weeks prior to surgery, when tissue was harvested.
RNA Extraction and Analysis
A fragment of ethmoid mucosa was removed during endoscopic sinus surgery, immediately wrapped in aluminum foil, and flash frozen in liquid nitrogen. Frozen biopsies were placed in lysis buffer and homogenized with a Qiagen TissueRupter (Qiagen, Valencia, California). RNA was prepared with the mirVana miRNA Isolation Kit (Applied Biosystems, Carlsbad, California), modified to include an on-column treatment with RNase-free DNase (Qiagen). RNA quality was assessed by RIN values from the BioAnalyzer 2100 (Agilent, Palo Alto, California). Total RNA (50 ng) was amplified with the Ovation RNA Amplification System V2 and WB Reagent (Nugen, Inc, San Carlos, California). Of the amplified cDNA, 4.4 µg was labeled with the FL Ovation cDNA Biotin Module V2 (Nugen, Inc) according to the manufacturer’s recommendations. The labeled cDNA was hybridized onto Affymetrix human genome HG-U133_Plus_2 arrays (Affymetrix, Santa Clara, California) and processed according to Affymetrix technical protocols. The average intensity of each array was scaled to a target intensity of 500.
Microarray Data Analysis
Raw Affymetrix data were imported into BRB-Array Tools 4.4.1 (Dr Richard Simon and BRB-Array Tools Development Team). Genes whose log intensity variance was in the bottom 75th percentile were filtered out, and differentially expressed genes were selected with an 80% confidence level and a false discovery rate of 0.1, an acceptable exclusion parameter for a study of this size.5,6 Class comparisons were then used to intersect gene lists and generate unique and common differentially expressed genes in CRSwNP and AFRS. These lists were then analyzed with the curated gene sets from Ingenuity Pathway Analysis (IPA; Qiagen) to generate top canonical pathways for each disease subtype. A right-tailed Fisher’s exact test was used to generate P values, with P < .01 considered significant. The overlap, presented as a ratio, represents the number of differentially expressed genes in a disease subtype over the total number of genes in a designated pathway reference set.
To test the relationship between the level of histatin probe expression (expressed as log rank intensity) and proinflammatory mediators, a Spearman’s correlation coefficient was calculated with microarray gene expression data from 115 patients with various subtypes of CRS (CRSwNP, AFRS, CRSsNP, AERD) or HCs (Supplemental Table S1).
Semiquantitative Polymerase Chain Reaction
Semiquantitative real-time polymerase chain reaction (PCR) was performed as previously described with an ABI Prism 7500 Sequence Detection System 7 (Applied Biosystems, Foster City, CA). The threshold cycle (Ct) was determined in duplicate for HTN1, HTN3, and β-actin, and these values were used to calculate relative Ct based on the DDCT method with β-actin as a reference gene according to the following formula: relative Ct = [1.8(Ctβ-actin– CtUnknown)] × 1,000,000. The following primer sequences were used: HTN1—FW: 5′ CATCATTCACATCGAGAATTTCC-3′, RV: 5′- TGCCAGTCAAACCTCTATAATC-3′; HTN3—FW: 5′-GCAAAGAGACATCATGGGTATAA-3′, RV: 5′-GAAGGGAAGTATCCTGAAACAC-3′; actin—FW: 5′- GGATGCAGAAGGAGATCACTG-3′, RV: 5′- CGATCCACACGGAGTACTTG-3′. Data were analyzed with Prism software (GraphPad, La Jolla, California). Outliers were identified and removed via the ROUT method. Data are represented as mean ± SEM. Data with nonnormal distributions were analyzed by 1-tailed Mann-Whitney U test (P ≤ .05 was considered significant).
Immunohistochemistry
Inflamed sinonasal mucosa was obtained at the time of surgery, placed into formalin, and processed for immunohistochemical staining according to the standard protocol of the University of Texas McGovern Medical School Department of Pathology and Laboratory Medicine. Polyclonal rabbit anti-human histatin 1 antibody (Cloud-Clone Corp, Katy, Texas) was used at a dilution of 1:1000. Monoclonal mouse anti-human histatin 3 antibody (Millipore Sigma, Billerica, Massachusetts) was used at a dilution of 1:500. Counterstaining was performed with hematoxylin. A Dako Omnis (Santa Clara, California) automated slide staining system was used. Images were obtained with a Motic EasyScan digital slide scanner (Richmond, British Columbia, Canada). Images were obtained at 40× magnification.
Results
Patient Demographics
Table 1 outlines the clinical features of the patients enrolled in this study. In total, there were 86 participants: 15 healthy controls, 37 AFRS, and 34 CRSwNP. Patients with AFRS had a lower median age (27 years) when compared with HCs (53.5 years) and the CRSwNP group (46 years). The prevalence of asthma in patients with AFRS was less than half (24.3%) than the CRSwNP group (55.9% with comorbid asthma), which is consistent with previous investigations showing decreased asthma prevalence among patients with AFRS. 8 As expected, all patients with AFRS tested positive for allergy to mold. For our correlation analysis, we included patients with CRSsNP and patients with AERD. The demographic data for these patients are included in Supplemental Table S1.
Differential Gene Expression in AFRS and CRSwNP
We performed a class comparison microarray analysis to identify the number of unique and shared gene expression variations in AFRS and CRSwNP ( Figure 1 , Venn diagram). Our initial analysis revealed 3331 differentially expressed genes in AFRS when compared with HCs, while there were 435 differentially expressed genes in CRSwNP versus HCs. We also performed a class comparison analysis to identify variations in gene expression that were unique and shared between CRSwNP and AFRS. We found that AFRS and CRSwNP shared 405 gene expression variations, AFRS exhibited 2923 unique gene expression variations, and CRSwNP demonstrated 30 unique gene expression variations.
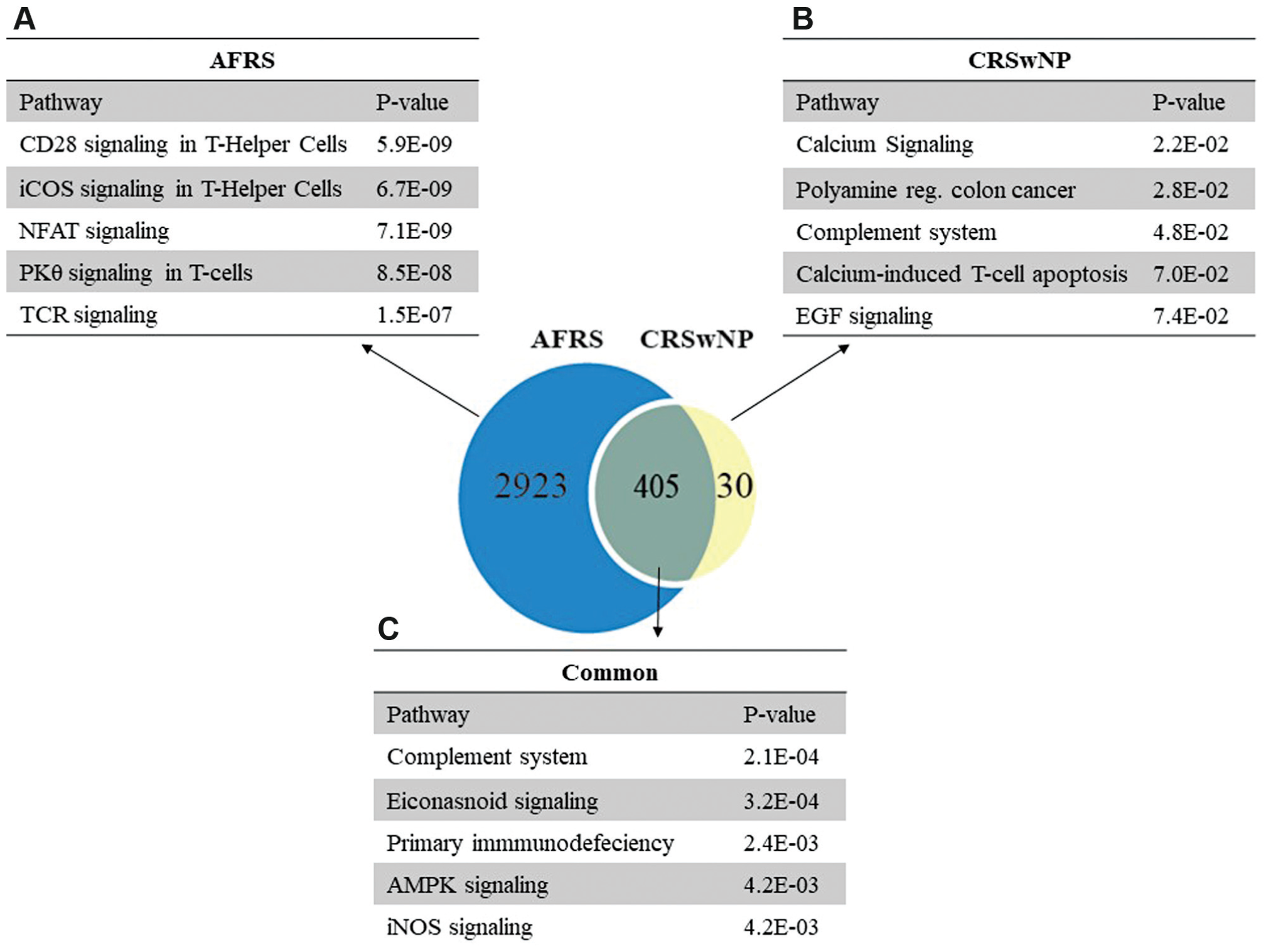
Ingenuity Pathway Analysis modeling of unique and shared gene expression variation in chronic rhinosinusitis with nasal polyps (CRSwNP) and allergic fungal rhinosinusitis (AFRS). Pathways unique to (a) AFRS subtype and (b) CRSwNP subtype. (c) Pathways common to both subtypes.
Pathway Analysis
We next modeled our microarray findings using IPA software and identified the top canonical pathways that were unique or shared between AFRS and CRSwNP ( Figure 1 ). AFRS exhibited markedly robust T helper 2 (Th2)–associated pathway signaling ( Figure 1a ), as demonstrated by its strong association with costimulatory signaling, including the ICOS, CD28, and PKCθ pathways, which play pivotal roles in Th2 effector function, including B-cell activation, isotype switching, T regulatory suppression, and CD4+ polarization.9,10 Few unique gene sequences were identified with differential expression in the CRSwNP group but not in the AFRS group. The pathway associations in CRSwNP are, by virtue of differential expression of type 3 inositol 1,4,5-triphosphate receptor and tropomyosin 2 beta (calcium signaling) and proteasome inhibitor subunit 1 (PI31), a modulator of proteasome formation and antigen processing ( Figure 1b ). 11
We next modeled our whole-genome microarray data in IPA to identify molecules with the greatest difference in relative expression in AFRS ( Table 2 ). This analysis was performed by comparing whole-genome microarray gene expression data between AFRS and CRSwNP. Many of the top 10 molecules with the greatest degree of upregulation in patients with AFRS were inflammatory mediators associated with type 2 inflammation, including the IgE immunoglobulin heavy locus (IGHE: 7.23-fold increase, P < 1.0E-07), and others associated with eosinophil chemotaxis (CCL11: 3.080 increase, P = 2.6E-04), eosinophil activation (IL5RA: 2.780 increase, P = 1.4E-04), B-cell differentiation (MS4A1: 2.910 increase, P = 5.3E-04), and Th2 development (CD28: 2.440 increase, P = 3.9E-05). Interestingly, the gene with the greatest decrease in expression was that encoding an antifungal peptide, histatin 1 (HTN1: 2.04-fold decrease, P = 3.1E-03).
Top Differentially Expressed Genes in AFRS.

Abbreviation: AFRS, allergic fungal rhinosinusitis.
HTN1 and HTN3 Expression Correlates Negatively with Type 2 Proinflammatory Gene Expression
The histatins are a family of peptides encoded by 2 genes: HTN1 and HTN3. Posttranslational processing generates a variety of histatin peptides—the most commonly studied are histatin 1, 3, and 5. 12 Histatins have currently been described exclusively in the oral cavity, where they exhibit antifungal, antibacterial, wound-healing, and immune-modulating properties.13-17 To date, no study has demonstrated expression of histatins in sinonasal mucosa, let alone within the context of CRS. Alterations in levels of such peptides may constitute a potent factor driving the exaggerated inflammatory response observed in AFRS.
We wished to further investigate the relationship between histatin expression and other genes of interest. We employed a Spearman’s rank correlation test to identify the relationship between histatin gene expression and that of type 2 proinflammatory mediators, using microarray data from 115 patients with all subtypes of CRS (CRSwNP, AFRS, CRSsNP, AERD; see Supplemental Table S1 for demographic data) and HCs to increase the strength of our test results ( Figure 2 ). HTN1 and HTN3 both demonstrated moderate to strong negative correlations with several markers of type 2 inflammation, including IL13RA1 (HTN1: r = −0.58, HTN3: r = −0.61), IL4R (HTN1: r = −0.55, HTN3: r = −0.59), POSTN (HTN1: r = −0.49, HTN3: r = −0.55), and MS4A2 (HTN1: r = − 0.47, HTN3 = −0.5) (for all correlations, P < .001).
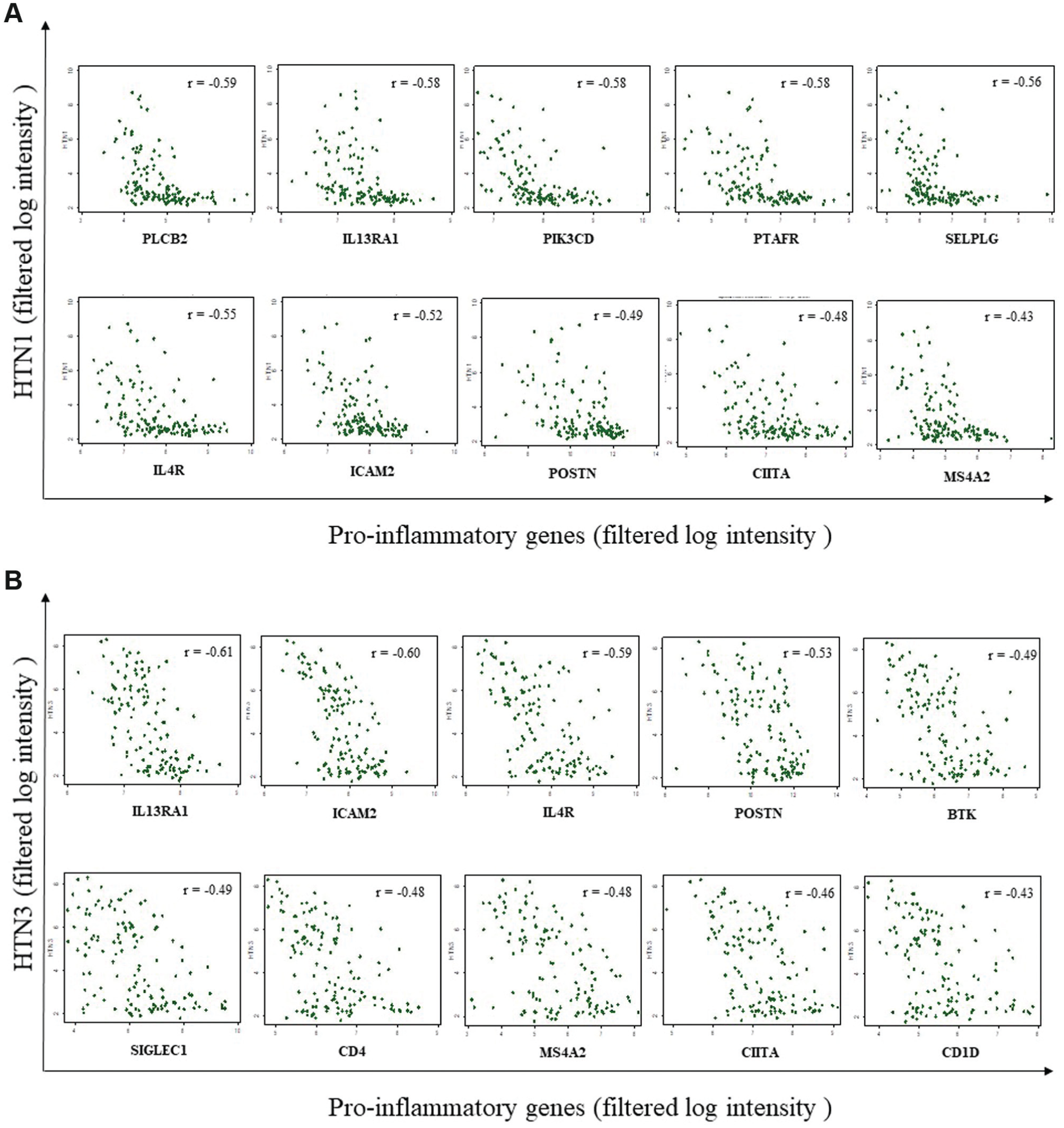
Negative correlation between histatins and type 2 proinflammatory mediators. Each patient’s gene expression is represented by a dot. (a) HTN1 and (b) HTN3. r = correlation coefficient. For all correlations, P < .001.
Quantitative PCR Confirmation of Altered Histatin Expression and Cell Localization
As HTN1 and HTN3 mRNA levels in many AFRS samples were at the lower limits of detection in the microarray data, thereby limiting the fold change measurement, we confirmed the differential expression between CRSwNP and AFRS by performing quantitative PCR in independent primary sinonasal tissue obtained from patients with AFRS and CRSwNP. Our results from dissected components of inflamed sinonasal mucosa in patients with AFRS and CRSwNP ( Figure 3 ) confirm our microarray findings indicating that HTN1 and HTN3 expression is significantly decreased in patients with AFRS when compared with CRSwNP (HTN1: P < .01; HTN3: P < .05). Of note, there was a substantial difference in the degree of histatin gene expression between our microarray results for histatin expression ( Table 2 ) and confirmatory quantitative PCR results ( Figure 3 ). We attribute this largely to differences in technique and sample size. Table 2 represents results from microarray-based gene analysis, and Figure 3 represents a quantitative PCR technique. The microarray-based gene analysis was conducted in a larger sample size (AFRS, n = 37; CRSwNP, n = 34); thus, differences in gene expression between AFRS and CRSwNP measured by microarray analysis may have been tempered by larger variations in expression within AFRS and CRSwNP.
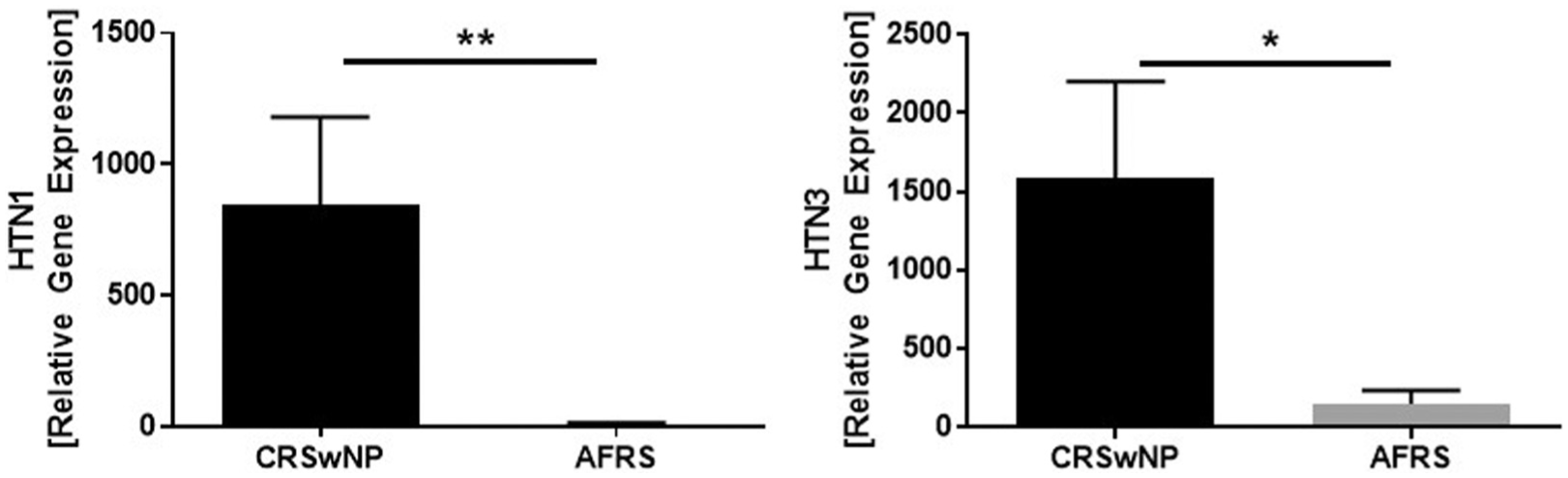
HTN1 and HTN3 gene expression in allergic fungal rhinosinusitis (AFRS) and chronic rhinosinusitis with nasal polyps (CRSwNP). Quantitative polymerase chain reaction was performed in whole inflamed sinonasal mucosal tissue obtained from patients with AFRS (n = 9) and CRSwNP (n = 9). Values are presented as mean ± SEM. *P < .05. **P < .01.
Histatin expression is localized to the serous acinar and serous demilune cells in the major salivary glands; however, its cell of origin in sinonasal mucosa is unknown, and so we wished to localize its expression within inflamed mucosa. 18 Preliminary studies in different fractions of cells isolated from inflamed sinonasal mucosa localized HTN1 and HTN3 transcript expression within a fraction that included macrophages, fibroblasts, and seromucinous glands (data not shown). To more definitively determine the localization of HTN1 and HTN3 within the inflamed mucosa from patients with CRSwNP, immunohistochemistry studies were performed. HTN1 protein expression was detected in the seromucinous glands ( Figure 4a ) and fibrovascular stromal cells ( Figure 4b ). HTN3 protein expression was localized to the seromucinous glands ( Figure 4c ) and respiratory epithelium ( Figure 4d ).
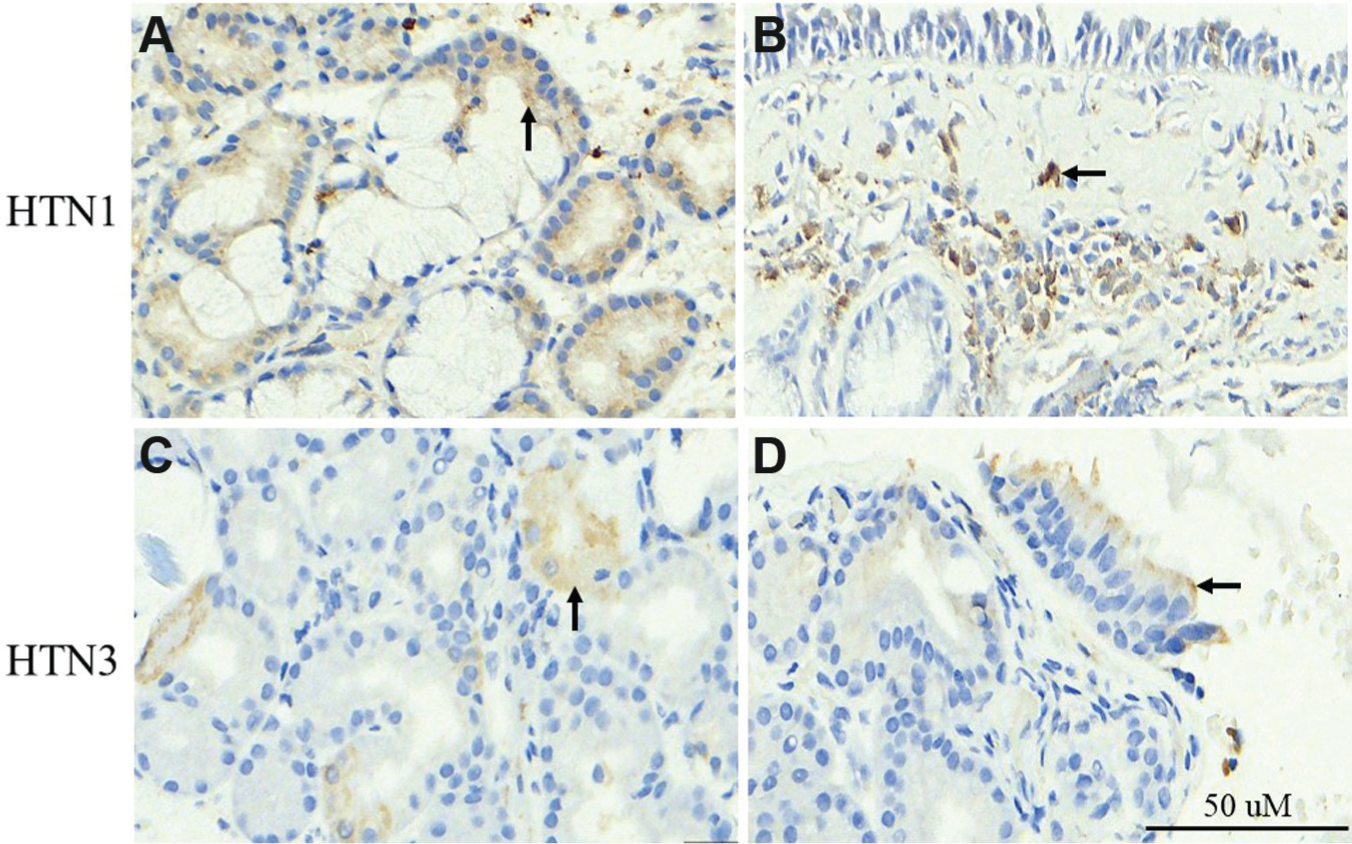
Localization of HTN1 and HTN3 in chronic rhinosinusitis with nasal polyps. HTN1 expression in seromucinous cells (a; arrow) and fibrovascular stromal cells (b; arrow). HTN3 expression in seromucinous cells (c; arrow) and respiratory epithelial cells (d; arrow).
Discussion
Successful disease classification in CRS will incorporate clinical features and molecular endotypes to promote more effective, personalized disease management. Proving endotypes in CRS is a major research endeavor that will result from (1) characterizing the molecular biology that drives the expression of specific phenotypes and (2) determining the impact of these molecular variations on clinically relevant outcomes.1,2,19 In this study, we attempted to characterize molecular expression patterns that differentiate AFRS and CRSwNP. To this end, we employed a top-down, microarray-based study design in a large population of clinically well-defined subtypes of CRS. In doing so, we present novel findings that differentiate AFRS from CRSwNP as a subtype demonstrating an exaggerated type 2 adaptive immune pathology, and we are the first to show altered expression of antifungal peptides, the histatins, in AFRS.
The expression of canonical type 2 inflammatory biomarkers, such as local eosinophilia, IgE, IL-4, IL-5, and IL-13, is linked with increased burden of disease and a recalcitrant course and, as such, represents a platform for endotype classification in CRS and asthma. We demonstrated that many subtypes of CRS, including those without polyps, express varying degrees of type 2 inflammatory markers, with AFRS representing an extreme of type 2 inflammation. 2 When we modeled the differentially expressed genes that were unique to AFRS, our pathway analysis revealed a strong association with Th2-mediated immune signaling, as evidenced by AFRS’s association with canonical costimulatory and T-cell receptor signaling pathways. These findings provide insight into the biological basis behind the observed “allergic type” inflammatory signature characterized by excessive mucin production, fungal hypersensitivity, tissue eosinophilia, and robust local and serum IgE production. Indeed, IL-4, IL-5, and IL-13 expression can originate from T cells, mast cells, and innate lymphoid cells in response to diverse signals originating from innate and adaptive immune arms. Our findings implicate the adaptive immune arm as a distinguishing molecular feature in AFRS and a potent source for type 2 inflammation. The inciting factor responsible for the observed exaggerated adaptive immune response in AFRS remains uncertain, and theories include subtype-specific differences in antigen-presenting cell populations, superantigens, pathogenic microbiota, and innate immune/barrier dysfunction, to name a few.20-28
Innate barrier proteins in the sinonasal mucosa constitute an important first line of defense against pathogens and mucosal injury in normal healthy mucosa. Studied examples in CRS include epithelial-derived proteins, such as defensins, cathelicidins, lysozyme, and SPLUNC1.20,28-30 As an example, Wei et al recently showed that SPLUNC1 is suppressed in eosinophilic CRSwNP when compared with noneosinophilic CRSwNP and that its expression can be decreased by stimulation with Th2 cytokines IL-4 and IL-13. 29 At present, the drivers of the exaggerated type 2 inflammatory response and fungal overgrowth in AFRS are unclear; however, loss of antimicrobial defense peptides (eg, histatin) may support fungal/antimicrobial growth and promote a skewed Th2 immune profile, such as the one observed in AFRS. This disease mechanism forms the basis of the “immune barrier” hypothesis, and it is believed to contribute to inflammation propagation in CRS. 20 This concept, however, has never been applied in a model of AFRS. Furthermore, histatins have never been described as being expressed in sinonasal mucosa.
Our microarray analysis highlighted significant negative correlations between the level of histatin gene expression and that of several known type 2 inflammatory mediators, including IL4R, IL13RA1, and POSTN. Future studies will elucidate the mechanisms surrounding decreased histatin expression in AFRS, and whether this is the result of specific cytokine-mediated suppression, represents an inciting factor in Th2 inflammation, or both. Our localization studies suggest that the source of histatins in patients with CRSwNP is the seromucinous and stromal cells of the sinonasal mucosa for HTN1 and seromucinous and epithelial cells for HTN3. Admittedly, we cannot rule out the possibility that histatin gene expression in AFRS is downregulated at the time of our analyses, nor do our experiments indicate whether the downregulation of histatins in AFRS is due to decreased gene expression in cells that normally express histatins or to the decrease in an entire population of those specific cells.
While this study revealed several significant findings, we are mindful of some inherent limitations. First, participants represent a category of CRS not controlled with medical therapies. As such, the conclusions may not be readily applicable to patients whose disease is controlled with medical therapies alone. Second, this study represents a cross-sectional design. The expression patterns observed correspond only to a single point in time and do not represent a continuum of gene expression. Last, our study is hypothesis generating—we wished to provide a molecular basis for the clinical differences between AFRS and CRSwNP on a large scale. Thus, we did not seek to provide a mechanistic explanation of our observations; however, our findings provide a foundation for the development of future mechanistic studies.
In summary, we present findings that clearly distinguish AFRS from CRSwNP on a molecular level, and we provide evidence that may explain many of the observed clinical features that are characteristic of AFRS. In our analysis, AFRS can be differentiated from non-AFRS CRSwNP by its association with pronounced adaptive immune signaling and the loss of expression of antimicrobial peptides, including the histatins. Our findings add to a growing body of literature in CRS that advances disease endotypes and the development of novel treatment strategies aimed at targeting these associated pathways.
Author Contributions
Disclosures
Supplemental Material
OTO764349_suppl_mat – Supplemental material for Distinguishing Molecular Features of Allergic Fungal Rhinosinusitis
Supplemental material, OTO764349_suppl_mat for Distinguishing Molecular Features of Allergic Fungal Rhinosinusitis by Matthew A. Tyler, Caroline J. Padro Dietz, Chris B. Russell, Martin J. Citardi, Shervin Assassi, Jun Ying, and Amber U. Luong in Otolaryngology–Head and Neck Surgery
Footnotes
Acknowledgements
The authors would like to thank Melissa Stephens of the McGovern Medical School immunohistochemistry core facility for her assistance with IHC studies, and Robyn Reed, MS for technical assistance.
Sponsorships or competing interests that may be relevant to content are disclosed at the end of this article.
This article was presented at the 2017 AAO-HNSF Annual Meeting & OTO Experience; September 10-13, 2017; Chicago, Illinois.
Supplemental Material
Additional supporting information is available in the online version of the article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.