
Abstract
Background:
Finding ways to a healthier ageing are increasingly becoming the focus of geriatric research. One way to accomplish this could be calorie restriction, as this is known to positively influence the ageing of model organisms.
Aim:
The aim of this study was to investigate the influence of calorie reduction (F. X. Mayr therapy) and of the calorie restriction mimetics resveratrol and spermidine on the expression of ageing-associated genes.
Methods:
mRNA expression in peripheral blood mononuclear cells (PBMCs) of 18 participants taking part in an F. X. Mayr therapy was analysed. The PBMCs of one additional participant were treated ex vivo with spermidine or resveratrol. mRNA expression of SIRT1, SIRT3, FOXO3 and SOD2 was determined for these two calorie restriction mimetics. For the F. X. Mayr therapy samples, mRNA of XPA was analysed additionally.
Results:
mRNA expression of the ageing-associated genes showed a distinct donor variation during F. X. Mayr therapy, with a significant increase in mRNA expression of SIRT1. Expression of XPA was similar to SIRT1, with a significant correlation at the last time point tested. Spermidine treatment of PBMCs resulted in a significantly increased expression of all genes tested, whereas resveratrol treatment caused a significant increase of SIRT3, FOXO3 and SOD2 mRNA expression.
Conclusions:
By increasing SIRT1 and XPA mRNA expression, calorie reduction in the form of F. X. Mayr therapy could contribute to a healthier ageing; however, the donor variability observed showed that not everyone benefited from this. Calorie restriction mimetics may be an option for promote healthier ageing for those who do not benefit from calorie reduction.
Keywords
Introduction
In an increasingly ageing society, identifying approaches to a healthier ageing and interventions counteracting the ageing process have become the focus of increasing research. In model organisms, calorie reduction (CR) has been shown to have a positive impact on ageing and age-related diseases (Berg and Simms, 1965; Colman et al., 2009; Jiang et al., 2000; Weindruch et al., 1986). Although humans have similar physiological and biochemical changes as for example monkeys and rodents on comparable diets (Walford et al., 2002), the studies are only partially transferable to humans. Nevertheless, it was found that in humans, CR reduced the risk of cardiovascular diseases (Fontana et al., 2007; Most et al., 2018), caused a reduction of oxidative damage to nucleic acids (Hofer et al., 2008) and improved DNA repair capacity (Matt et al., 2016).
In order to increase adherence to caloric restrictive diets, the addition of calorie restriction mimetics (CRMs) appears to be a promising intervention to support CR. Two known CRMs are the polyamine spermidine found for example in dried soybeans, wheat germ, and matured cheddar (Atiya Ali et al., 2011) and the polyphenol resveratrol, which is contained in red grapes, raspberries and peanuts. With increasing age the spermidine content in different rat tissues decreases (Jaenne et al., 1964), and it has been shown that spermidine treatment prolongs the lifespan of model organisms and human immune cells (Eisenberg et al., 2009). Spermidine also induces autophagy (Eisenberg et al., 2009), which may be the mechanism underlying its anti-ageing effect (Bergamini et al., 2007; Hansen et al., 2008) and may thus contribute to a healthier ageing. Similar to spermidine, resveratrol extends the lifespan of different model organisms (Howitz et al., 2003; Wood et al., 2004) and improves the health of mice (Berg and Simms, 1965). In humans, Timmers et al. reported an improved mitochondrial metabolism involving Sirtuin 1 (SIRT1) and reduced inflammation markers in obese humans after resveratrol treatment (Timmers et al., 2011). In addition, resveratrol leads to increased insulin sensitivity as well as decreased oxidative stress (Brasnyó et al., 2011). Spermidine has recently become the focus of ageing research. With increasing age the spermidine content in different rat tissues decreases (Jaenne et al., 1964). Moreover, spermidine treatment prolongs the lifespan of model organisms and human immune cells and also induces autophagy (Eisenberg et al., 2009). Provided that autophagy plays an important role in ageing, spermidine, as an inducer of autophagy, may thus contribute to a healthier ageing.
In the context of CR, sirtuins are of major interest. mRNA expression of SIRT1, a NAD-dependent deacetylase, is induced in human blood cells after a CR-period of 8 weeks (Crujeiras et al., 2008) and in human monocytes after treatment with resveratrol (Tsuchiya et al., 2017). SIRT1 also plays an important role in nucleotide excision repair by deacetylating Xeroderma pigmentosum complementation group A (XPA) (Fan and Luo, 2010). Sirtuin 3 (SIRT3), a mitochondrial deacetylase, possibly acts as a tumour suppressor by maintaining mitochondrial integrity (Kim et al., 2010). SIRT3 also regulates pathways leading to age-related diseases such as cardiovascular diseases (Hafner et al., 2010), insulin resistance (Lantier et al., 2015) and can reduce oxidative stress by an activation of superoxide dismutase 2 (SOD2) (Qiu et al., 2010). Oxidative stress, resulting from an increase in reactive oxygen species (ROS) generated during respiration, has been shown to be a major cause of ageing. As a potent antioxidant, SOD2 has been shown to play an important role in these processes (Qiu et al., 2010) Both, SIRT3 and SOD2 can be induced by intermittent fasting (Wegman et al., 2015). Another gene of interest, Forkhead Box O3 (FOXO3), has been associated with longevity in humans (Flachsbart et al., 2009). FOXO3 also triggers autophagy (Mammucari et al., 2007), which decreases with ageing (Uddin et al., 2012).
We investigated the impact of F. X. Mayr therapy as a form of CR, as well as the influence of two CRMs, namely resveratrol and spermidine, on the expression levels of the ageing-associated genes SIRT1, SIRT3, XPA, FOXO3 and SOD2 in human peripheral blood mononuclear cells (PBMCs). There were 18 participants in the study. Participants lost an average of 5 kg of body weight during the 6-week period of CR. The results on changes in the DNA repair capacity of these participants have been published previously (Matt et al., 2016). Based on these results, the influence of CR on mRNA expression in the 18 participants was investigated and the results are presented here.
Materials and methods
Ex vivo gene expression during calorie reduction
mRNA expression in the PBMCs of 18 human participants between 25 and 63 years of age was analysed after a period of CR in the frame of an F. X. Mayr therapy. Thirteen of the individuals were female and five were male. These participants were previously tested for their DNA repair capacity (Matt et al., 2016). In brief, the participants underwent a 6-week therapy consisting of a 1-week pretreatment (eating no indigestive food), followed by 3 weeks of diet food intake and a 2-week convalescence period (re-adjusting the body to regular food). Assorted diet food was tailored for each person and consisted of eupeptic food like spelt buns, sheep yogurt, potatoes, root vegetables, protein-containing side-dishes and omega-3 fatty acids (Rupprecht, 2016). Crucial to the success of this intervention was that the participants ate very slowly, chewed the food thoroughly and stopped eating when satiety was reached. During this time, calorie intake was reduced and participants lost 3–8 kg of body weight (5 kg on average), therefore this intervention qualified as CR. Venous blood was drawn before CR, on the eighth day of therapy and during convalescence. For more detailed information see Matt et al. (2016). Isolated PBMCs of the 18 participants were stored pelleted and frozen (–86°C) until RNA isolation.
Ex vivo treatment of PBMCs with calorie restriction mimetics
Spermidine (Sigma-Aldrich, Taufkirchen, Germany) and resveratrol (Sigma-Aldrich, Taufkirchen, Germany) were used to test the influence of CRMs on mRNA expression of ageing-associated genes. For ex vivo treatments of PBMCs 9 ml venous blood was withdrawn using K-EDTA Monovettes® (Sarstedt AG & Co., Nuembrecht, Germany). PBMCs of a healthy female were isolated using Leucosep™ tubes (Greiner Bio-One GmbH, Frickenhausen, Germany) according to the manufacturer’s instructions. This healthy volunteer did not take part in the F. X. Mayr therapy previously mentioned or any other kind of CR. Cell count was determined using a Luna-FL™ automated cell counter (Logos Biosystems, Inc.). For each concentration of the CRMs, as well as for the untreated control, 3 × 106 PBMCs were used. Since resveratrol was dissolved in dimethyl sulfoxide (DMSO), a solvent control containing 3 × 106 cells and 40 µl of DMSO (the same amount that was used for 100 µM resveratrol) was performed. Spermidine was dissolved in water. The PBMCs were seeded and incubated in six-well plates (Sarstedt AG & Co., Nuembrecht, Germany) containing 3 ml RPMI1640 medium without phenol red and 20% FBS (both Gibco® by Life Technologies). Spermidine and resveratrol concentrations were initially chosen following Morselli et al. (2011) but the final concentrations were determined experimentally. Final concentrations of spermidine were 10 µM, 20 µM and 30 µM and resveratrol was used at concentrations of 75 µM and 100 µM. For both CRMs incubation times were 24 h, 48 h and 72 h at 37°C and 5% CO2. The medium was changed every 24 h. At indicated time points, RNA was isolated. Treatment with spermidine or resveratrol was carried out in two independent experiments each.
Gene expression analyses
Isolation of RNA was performed using the High Pure RNA Isolation Kit (Roche Diagnostics GmbH, Mannheim, Germany). Some 106 PBMCs from the CR samples and 3 × 106 cells from CRM samples were used per isolation. Amplification of cDNA was conducted with the Transcriptor First Strand cDNA Synthesis Kit (Roche Diagnostics GmbH, Mannheim, Germany), using random hexamer primers for CR samples and random hexamer and anchored-oligo(dT)18 primers for CRM samples. Both kits were utilized according to the manufacturer’s instructions.
Quantitative polymerase chain reaction (qPCR) was carried out on LightCycler® 480 instrument (Roche Diagnostics GmbH, Mannheim, Germany) with LightCycler® 480 SYBR Green I Master (Roche Diagnostics GmbH, Mannheim, Germany). Each qPCR consisted of 1 µl (0.5 pmol/µl) of each primer (biomers.net GmbH, Ulm, Germany), 10 µl SYBR Green I Master and 8 µl of cDNA. qPCR analyses were carried out in triplicate for each sample. The following PCR programme was used: 5 min pre-incubation (95°C), followed by 95°C 10 s, 62°C 10 s and 72°C 10 s with fluorescence measurements at the end of the elongation phase. After 45 cycles, melting curve analysis was performed. Ribosomal protein lateral stalk subunit P0 (RPLP0), glyceraldehyde-3-phosphate dehydrogenase (G3PDH), β-Actin and glucuronidase beta (GUSB) were used as housekeeping genes for normalization of the target genes for the CR samples. For the CRM samples, RPLP0, G3PDH and β-Actin were used for normalization. N-fold mRNA expression was calculated with the 2-ΔΔCt method. Table 1 shows the primer sequences; primers for RPLP0 were designed by Gebhard et al. (2014).
Sequences of the utilized primers.

Data analyses
In addition to the analysis including all 18 participants, individuals were also grouped into the lower 50% (‘low’) and the higher 50% (‘high’) according to their DNA repair capacity before pretreatment. The first normalization step of the qPCR data was subtracting Ct values of housekeeping genes from the Ct values of the genes of interest, resulting in ΔCt values. ΔCt values were used to analyse the difference in the initial SIRT1 mRNA expression of the F. X. Mayr participants. Graph Pad Prism 7.04 for Windows (GraphPad software, inc., San Diego, CA) was used for statistical analyses and graph generation. Statistical analyses were conducted using one-way analysis of variance (ANOVA) with Tukey’s multiple comparison test (CR samples) or as a two-way ANOVA with Dunnett’s multiple comparison test (CRM samples). For the evaluation of possible correlations, linear regression analyses were carried out and Pearson’s coefficient of correlation was calculated.
Results
Influence of CR on mRNA expression of ageing-associated genes
There were 18 participants in our CR (F. X. Mayr therapy) study. Participants lost an average of 5 kg of body weight during this 6-week programme. In 2016, we published the first results of this study (Matt et al., 2016). Here, the influence of CR on mRNA expression in the 18 participants was investigated. We focused on the ageing-associated genes SIRT1, XPA, SIRT3, FOXO3 and SOD2, hypothesizing a positive effect of CR on their expression. When analysing all 18 participants together, only SIRT1 expression was affected by CR, with a slight increase after 8 days of CR and a significant increase in expression during the study period (Figure 1). A similar, although not significant, pattern of mRNA expression was observed for XPA.
Since SIRT1 and XPA are particularly related to DNA repair we grouped the participants according to their DNA repair capacity prior to CR (Matt et al., 2016). As previously published, the 18 participants were stratified into a lower (low) and a higher (high) DNA repair capacity at onset of CR.
Participants in the low pre-existing DNA repair capacity group had a significant increase in mRNA expression of SIRT1 on the eighth day of fasting, which was maintained throughout the convalescence period (Figure 1). When divided into high and low pre-existing DNA repair capacity, it became apparent that the significant increase in mRNA expression of SIRT1 was exclusively in the participants with low pre-existing DNA repair capacity. In this group, mRNA expression of XPA also increased on the eighth day of therapy and again during convalescence, although not significantly. Interestingly, expression of SOD2 mRNA was also affected by CR. While there was a tendency for increased expression on the eighth day, as well as a subsequent reduction of SOD2 mRNA expression during the convalescence period for all participants, this transient increase became significant in the group with higher DNA repair capacity at onset of the F. X. Mayr therapy.
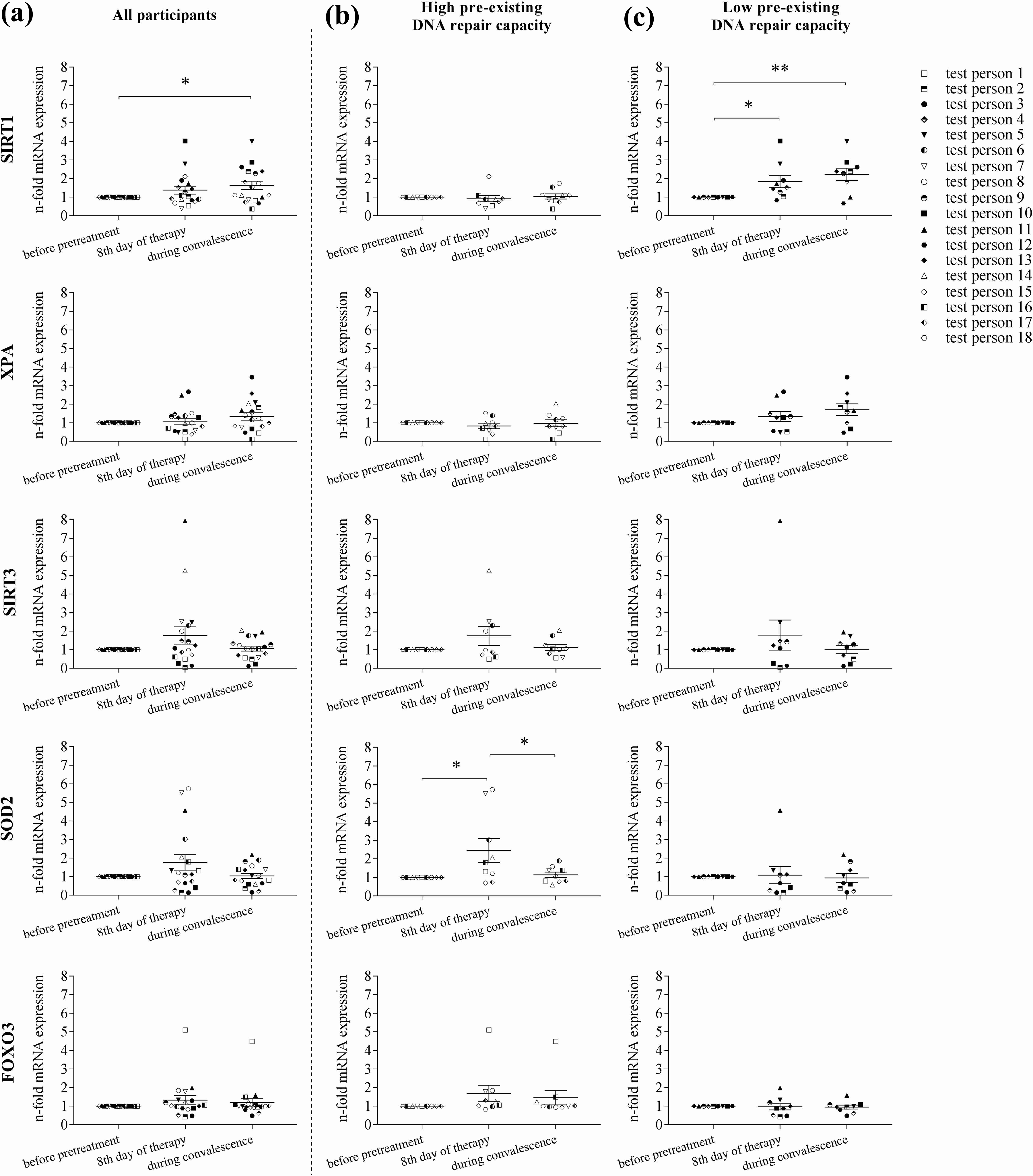
Significant increase of SIRT1 mRNA expression during calorie reduction. PBMCs from all 18 participants were analysed for mRNA expression by quantitative PCR and gene expression was stratified into three groups: (a) all participants analysed together, (b) stratified into participants with higher (‘high’) DNA repair capacity and (c) with lower (‘low’) DNA repair capacity at onset of therapy (****p < 0.0001, ***p < 0.0002, **p < 0.0021, *p < 0.0332).
Due to the similar increases in mRNA expression of SIRT1 and XPA, we additionally looked for a direct correlation in expression of the two genes by plotting their expression mutually (Figure 2). On the eighth day of the therapy, a weak correlation became apparent, but was not significant. During convalescence, however, a significant correlation between SIRT1 and XPA mRNA expression became evident (Figure 2(b)).
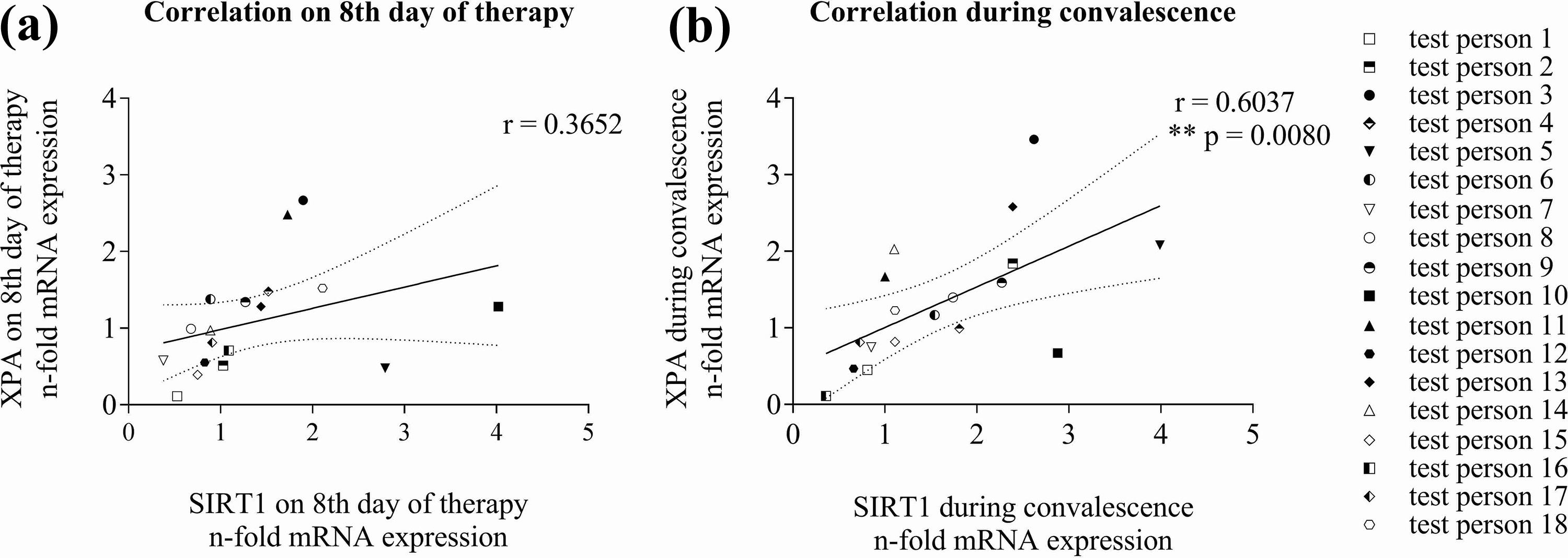
Correlation of SIRT1 and XPA gene expression during calorie reduction. (a) Expression on Day 8 of therapy of XPA mRNA was plotted against expression of SIRT1, as well as (b) during the convalescence period (****p < 0.0001, ***p < 0.0002, **p < 0.0021, *p < 0.0332. Pearson’s coefficient of correlation r = 0.3652 for the eighth day of therapy and r = 0.6037; **p = 0.0080 for the last time point).
When analysing the PCR results independently by normalizing the gene expression levels only to the internal housekeeping genes, that is, using the Ct values, it became apparent that expression levels of SIRT1 were associated with DNA repair capacity. In the participants with low DNA repair capacity, SIRT1 expression was significantly lower prior to CR and increased through CR and further during the convalescence period (Figure 3). When directly correlating SIRT1 expression levels and DNA repair capacity prior to CR, a higher pre-existing DNA repair capacity correlated with the slightly lower ΔCt values (i.e. higher mRNA expression) in this group. Furthermore, the slightly higher ΔCt values correlated with a lower pre-existing DNA repair capacity prior to CR (r = 0.5138), again indicating a strong link between DNA repair and SIRT1.
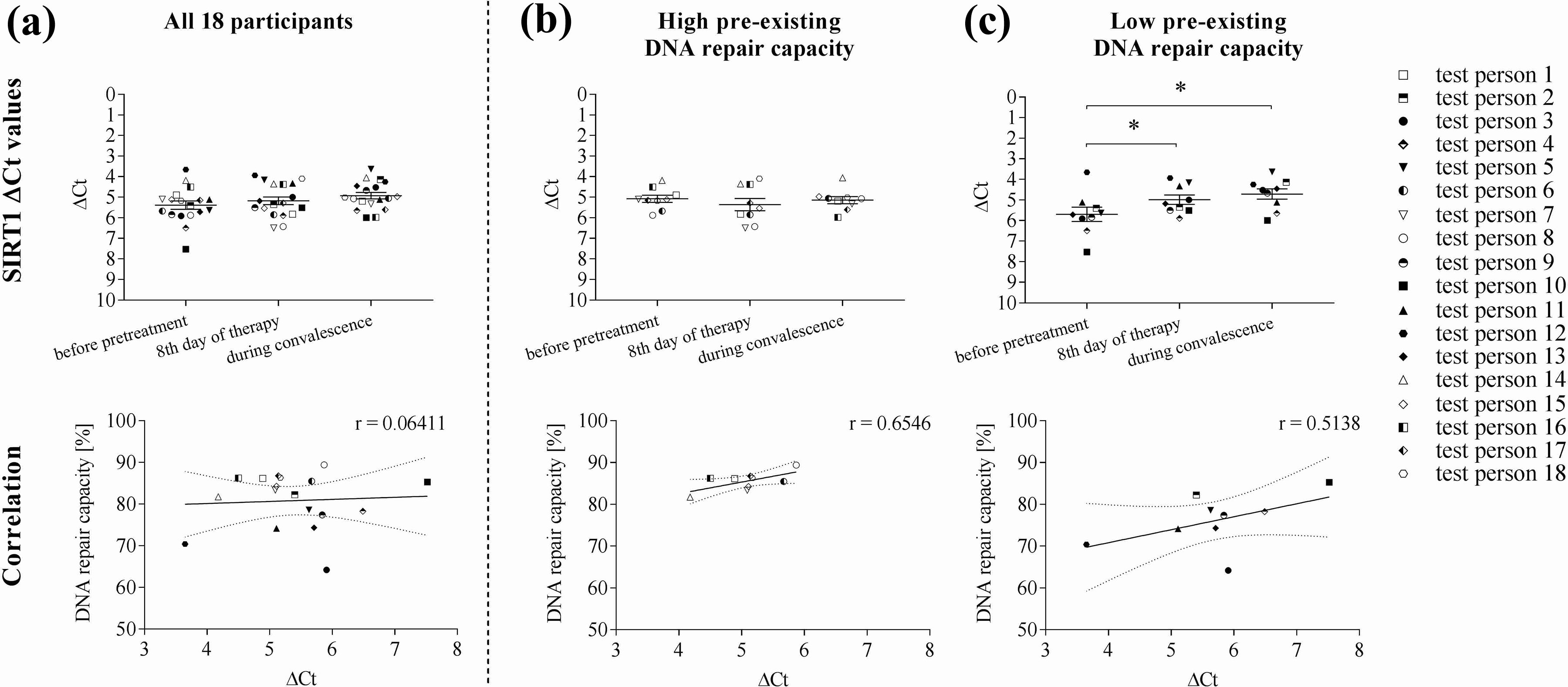
Correlation of DNA repair capacity and expression levels: (a) all participants, (b) high pre-existing DNA repair capacity and (c) low pre-existing DNA repair capacity. For a better understanding, Y-axes were inverted (****p < 0.0001; ***p < 0.0002; **p < 0.0021; *p < 0.0332; Pearson’s coefficient of correlation r = 0.06411 for all participants; r = 0.6546 for the ‘high’ group; r = 0.5138 for the ‘low’ group).
Influence of CRMs on mRNA expression of ageing-associated genes
In order to test the influence of CRMs on the mRNA expression of ageing-associated genes, ex vivo treatment of human PBMCs with CRMs was performed. For this purpose, the PBMCs of a healthy volunteer were treated with either spermidine or resveratrol. This volunteer was not subjected to the F. X. Mayr therapy or any other fasting intervention. Results of mRNA expression analyses are displayed in Figure 4 as mean n-fold mRNA expressions of two independently conducted experiments.
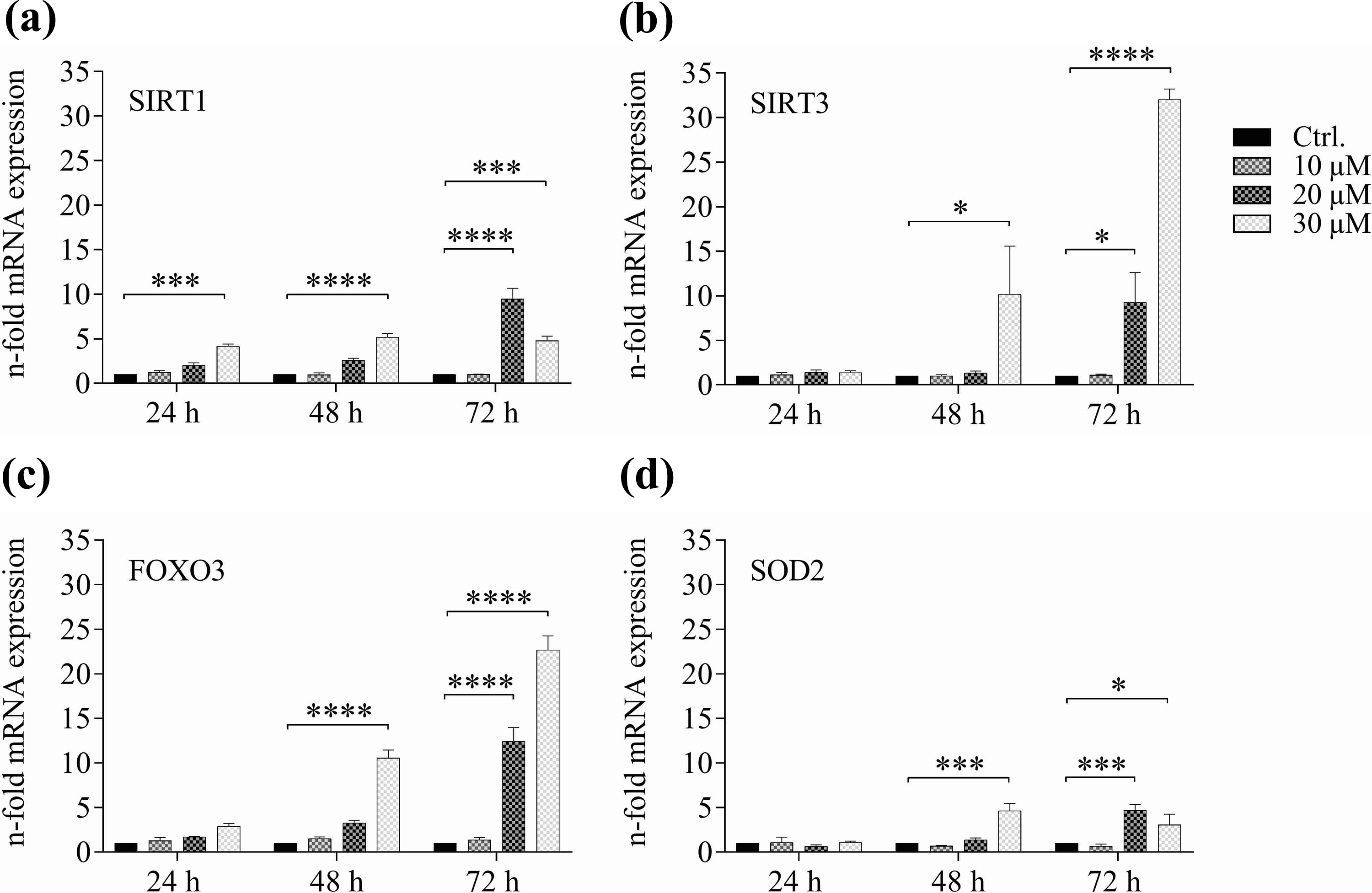
The CRM spermidine induced mRNA upregulation of the ageing-associated genes: (a) SIRT1, (b) SIRT3, (c) FOXO3 and (d) SOD2 in vitro in human PBMCs. Donor PBMCs were treated with spermidine for indicated time points with increasing concentrations of spermidine, mRNA extracted and analysed by qPCR (****p < 0.0001, ***p < 0.0002, **p < 0.0021, *p < 0.0332).
After 24 h and 48 h of treatment, there was a significant increase in SIRT1 mRNA expression with 30 µM of spermidine, and after 72 h with 20 µM of spermidine. With 30 µM the increase in SIRT1 levels was lower but still significant. When looking at SIRT3 expression, significant increases in gene expression were observed after 48 h, and were further increased at 72 h, with an upregulation of over thirtyfold for the highest concentration of spermidine. Furthermore, a significantly elevated mRNA expression could be seen for 20 µM of spermidine. Similar results were obtained with FOXO3 mRNA expression, which also started increasing after 48 h at the highest concentration of spermidine, as well as after 72 h at 20 µM and 30 µM. Of all four ageing-associate genes, SOD2 mRNA levels increased the least, but did change significantly after 48 h of treatment with 30 µM spermidine and stayed high. At 72 h of incubation with 20 µM and 30 µM spermidine, SOD2 mRNA expression also increased significantly.
When treating PBMCs with resveratrol, we did not observe significant increases in SIRT1 mRNA expression (Figure 5). However, expression levels of SIRT3 were affected with a significant increase in mRNA expression after 48 h at 100 µM resveratrol, and after 72 h already starting at the lower concentration. Significant changes of FOXO3 mRNA expression were observed after 48 h at a concentration of 75 µM and of 100 µM. After an incubation time of 72 h, a significant increase of FOXO3 mRNA expression was observed, starting at the lower concentration of 75 µM. Interestingly, mRNA expression of SOD2 was only regulated at the earlier time point of 24 h of treatment, albeit only at the lower concentration of 75 µM resveratrol. Neither 48 h nor 72 h of treatment resulted in an induction of SOD2 mRNA expression.
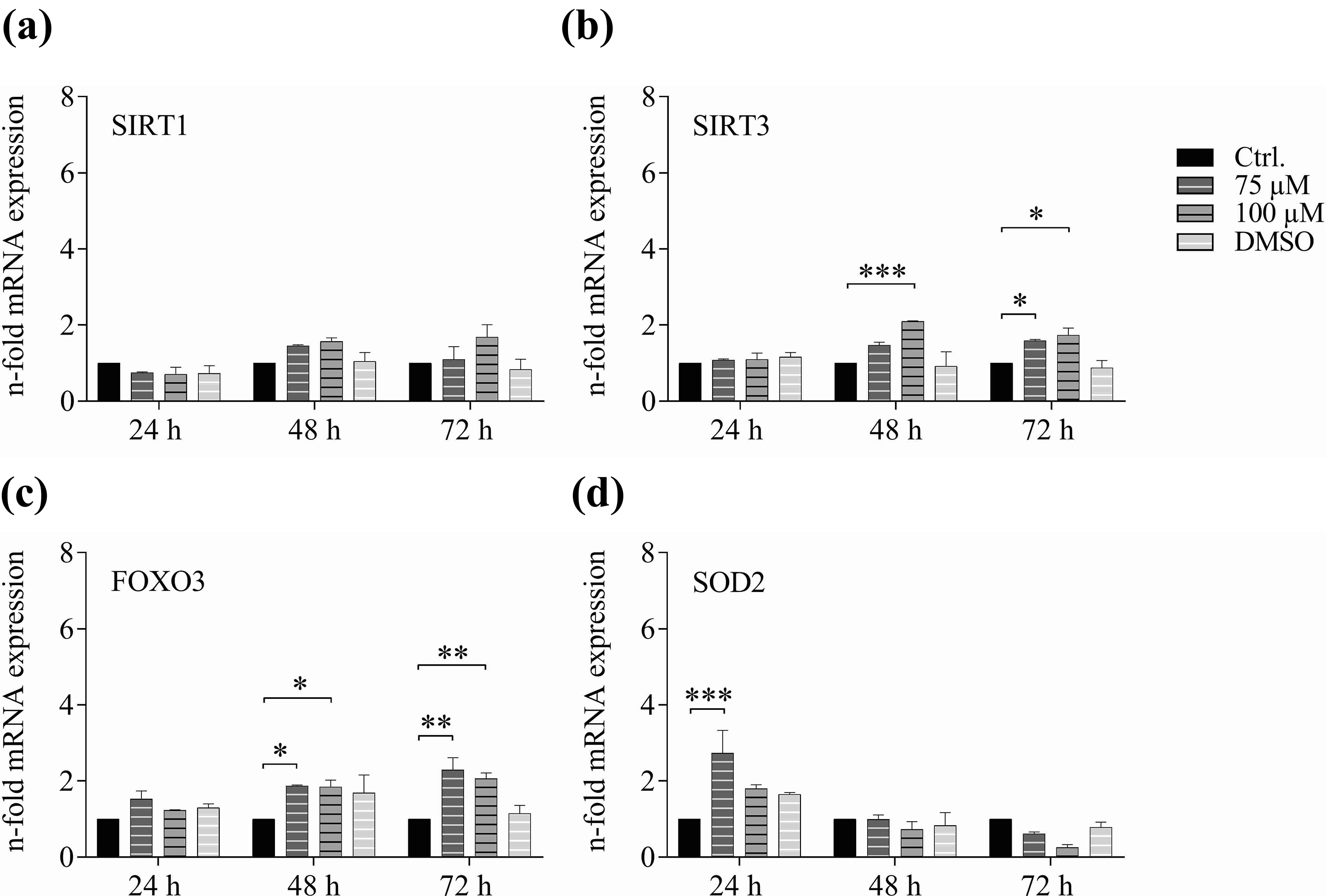
Resveratrol induced mRNA expression of the ageing-associated genes: (a) SIRT1, (b) SIRT3, (c) FOXO3 and (d) SOD2 in vitro in human PBMCs. Donor PBMCs were treated with resveratrol for indicated time points with increasing concentrations of resveratrol, mRNA was extracted and analysed by qPCR (****p < 0.0001, ***p < 0.0002, **p < 0.0021, *p < 0.0332).
Discussion
Here, we investigated the influence of CR on mRNA expression of ageing-associated genes in the PBMCs of 18 participants undergoing CR. We also tested the effects of two CRMs (spermidine and resveratrol) on the PBMCs of one additional volunteer ex vivo. This additional individual did not participate in F. X. Mayr therapy or any other intervention involving CR. Subsequently, mRNA expression of the ageing-associated genes SIRT1, SIRT3, FOXO3 and SOD2 was examined for both treatments. The CR samples were additionally analysed for XPA mRNA expression.
In a previous study, the same 18 participants had been analysed for their DNA repair capacity before, during and after CR. When stratifying the participants into lower (low) and higher (high) DNA repair capacity at onset of F. X. Mayr therapy, we were able to show a significant CR-induced benefit in DNA repair capacity in the group with lower capacity at onset (Matt et al., 2016). Again, when grouping the participants into low and high DNA repair capacity prior to CR, it became apparent that the participants of the low group had a significant increase in SIRT1 mRNA expression, compared with the group with higher DNA repair capacity at onset of CR. SIRT1 is involved in the regulation of DNA repair by modulating the acetylation status of XPA (Fan and Luo, 2010) and XPA in turn is an essential factor for nucleotide excision repair (Sugitani et al., 2016). Increased mRNA expression of SIRT1 may thus lead to an increased deacetylation of XPA and possibly to an increased DNA repair rate. The observed correlation of DNA repair capacity and ΔCt values of XPA support this. Mercken et al. showed a shift from growth to repair due to CR in humans (Mercken et al., 2013). The here described increase in mRNA expression of XPA suggests an improvement in DNA repair capacity triggered by CR. This in turn may contribute to maintaining genome integrity and thus contribute to prevent ageing. For the other genes studied, we also observed inter-individual differences regarding the induction of mRNA expression due to CR. An important function of SIRT3 is to control the amount of ROS in the cell by deacetylation and activation of SOD2. This process leads to an elimination of ROS and consequently protects the cell (Qiu et al., 2010). For participants with increased SIRT3 and SOD2 mRNA expression this means increased protection against oxidative damage. FOXO3 in turn has already been associated with longevity in various human populations (Anselmi et al., 2009; Flachsbart et al., 2009; Willcox et al., 2008). As a transcription factor, FOXO3 also regulates ROS signalling by inducing SOD2 (Kops et al., 2002) and thus protects cells from oxidative damage. Furthermore, oxidative stress stimulates SIRT3 to deacetylate FOXO3, thereby affecting mitochondrial biogenesis and finally resulting in an increased protection against oxidative stress (Tseng et al., 2013). However, the here reported increase in mRNA expression of SOD2 can also be caused indirectly since the transcription factor FOXO3 activates SOD2 transcription, resulting in increased levels of mRNA (Kops et al., 2002). In summary, CR affected the expression of ageing-associated genes differently in individual participants, suggesting donor variability. Individuals displaying an increase in mRNA expression of ageing-associated genes could benefit from CR via an improved DNA repair rate and increased protection against oxidative damage. This in turn could lead to a slowing of the ageing process.
The results of the ex vivo treatments with CRMs showed that spermidine and resveratrol affected mRNA expression in human PBMCs, possibly to an even greater extent than the F. X. Mayr therapy did. Various studies have already demonstrated the influence of resveratrol on the mRNA and protein expression of ageing-associated genes in humans (Mathieu et al., 2016; Zhou et al., 2014). Mansur et al. for example found that 500 mg of resveratrol and caloric restriction (1000 calories/day) similarly increased the plasma level of SIRT1 (Mansur et al., 2017). Our findings underscore the potential of resveratrol as a CRM, since it influences ageing-associated genes in human PBMCs in a similar manner as CR does. Due to the increase in mRNA expression of SOD2, PBMCs may be better protected from oxidative stress and might have elevated levels of autophagy due to increased levels of FOXO3. Increased oxidative stress as well as decreased autophagy are associated with ageing. By counteracting this, the process of ageing could be slowed down.
So far, few studies on spermidine and its influence on the expression of ageing-associated genes exist. In cell culture experiments, Chen et al. showed that nutrient starvation induces SIRT3 mRNA expression (Chen et al., 2011). This in turn leads to an activation of SOD2 (Chen et al., 2011), resulting in an increased protection from oxidative damage. In 2018, a population-based study focused on mortality in humans and associated a higher spermidine intake (>79.8 µmol/day) with a lower mortality (Kiechl et al., 2018). These results point to the CR-mimicking effects of spermidine in terms of reducing mortality in whole organisms. Our own results point to CR-mimicking effects of spermidine and resveratrol, as they also affected mRNA expression in human PBMCs. Although these are promising initial results, CRMs cannot replace the holistic approach of CR in the form of the F. X. Mayr therapy, as CR is, among other treatments (e.g. manual abdominal treatments), only part of F. X. Mayr therapy. They may however have a supportive effect in individuals who experience no, or only a slightly positive effect on the expression of ageing-associated genes. Accordingly, further studies involving more participants and additional therapies in combination with CRMs are required to support this hypothesis.
In conclusion, we observed a positive effect of CR and CRMs on ageing-associated genes. Furthermore, we showed similar effects of CR and CRMs on the genes investigated. Perhaps CRMs could be used as a tool to boost the effects of CR on gene expression in this context. Despite the small number of participants, our results contribute to a better understanding of the mechanisms with which CR and CRMs affect human cells and could point to new approaches to a healthier ageing process.
Footnotes
Acknowledgements
The authors thank Dr med. Karin Rupprecht (Centre for Traditional Chinese and Naturopathic Medicine, Sigmaringen), Dr med. Adrian Schulte (F. X. Mayr Centre, Bodensee) for drawing of the blood samples and helpful discussion on calorie reduction and Prof Dr Suzanne Kadereit for editing this work.
Authors’ contributions
Design and conception of the study on CR: KM, JB; mRNA analyses of the CR samples, data interpretation, drafting of manuscript: KM; statistical analyses: KM, ASM; design and conception of the study on CRMs: KM, BH, JB; gene expression analyses of CRM samples, data analysis: BH; critical revision of the manuscript: KM, BH, ASM, JB. All authors approved the manuscript.
Availability of data and materials
Data analysed during these studies are available from the corresponding author upon reasonable request.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
All experiments described in the following sections were conducted in accordance with the declaration of Helsinki and were approved by the ethics committee of the Landesärztekammer Baden-Württemberg (State Medical Association of Baden Württemberg). Participants gave their informed consent before the drawing of blood samples.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Baden-Württemberg Ministry of Science, Research and Art, funding line ‘Innovative Projekte/Kooperationsprojekte’. ASM holds a scholarship of the ‘Cooperative Graduate School InViTe’, funded by the Baden-Württemberg Ministry of Science, Research and Art. JB was supported by the International Society of Mayr Physicians and mse Pharmazeutika GmbH.