
Abstract
Introduction
Infertility is a growing global health concern, affecting approximately 15% of couples of reproductive age (Cox et al., 2022). Male factors contribute to nearly half of cases, primarily through abnormalities in sperm count, motility, morphology, and viability (Takeshima et al., 2021). These parameters are critical determinants of successful conception (Houston et al., 2022). Over recent decades, the prevalence of male infertility has risen, driven by environmental pollutants, lifestyle factors, dietary habits, and increased exposure to oxidative stress and chronic inflammation (Minhas et al., 2021).
Conventional treatments, including hormonal therapies, antioxidants, and lifestyle modifications, often show limited efficacy and accessibility (Asadi et al., 2021). This has prompted interest in novel, cost-effective interventions, including probiotics—live microorganisms that, when administered in adequate amounts, confer a health benefit on the host (Hill et al., 2014; Hotel and Cordoba, 2001), which have gained increasing attention for their role in modulating human health, including reproductive health (Del Giudice et al., 2021; Morshedzadeh et al., 2021).
Note that, throughout this review, probiotic nomenclature follows the updated taxonomy from the 2020 reclassification of the genus Lactobacillus (Zheng et al., 2020). For example, species formerly classified as Lactobacillus rhamnosus are now designated Lacticaseibacillus rhamnosus, and Lactobacillus plantarum as Lactiplantibacillus plantarum.
Traditionally recognized for improving gastrointestinal health (Bakhtiary et al., 2021; Oliveira et al., 2024), probiotics also exert systemic effects by reducing oxidative stress and inflammation, modulating immune responses, and enhancing gut–testis axis communication (Helli et al., 2022; Magill and MacDonald, 2023). Emerging evidence suggests that certain species may improve sperm motility, morphology, and DNA integrity while lowering oxidative damage (Akram et al., 2023).
Despite growing interest, evidence on probiotic effects on sperm parameters remains fragmented, with variability in study designs, species, dosages, durations, and outcome measures. Moreover, the underlying mechanisms are incompletely understood. This review builds upon recent syntheses (Oliveira et al., 2024) by incorporating animal models to provide mechanistic depth and a more comprehensive evidence base. Also, this systematic review aims to comprehensively synthesize available evidence on probiotic interventions for sperm parameters, identify effective species and protocols, elucidate potential mechanisms, and highlight gaps to guide future research and clinical application.
Methods
Study design
The protocol for this systematic review was not prospectively registered in a public registry such as PROSPERO. This decision was made because the review uniquely integrates substantial preclinical evidence from animal models with human studies, a mixed design that may not fully align with PROSPERO's primary focus on systematic reviews of interventions with direct health-related outcomes in humans. Nonetheless, methodological rigor and transparency were ensured through strict adherence to the Preferred Reporting Items for Systematic reviews and Meta-Analyses (PRISMA) 2020 guidelines (Page et al., 2021), including comprehensive reporting of the search strategy, eligibility criteria, study selection process, data extraction, and risk of bias (RoB) assessments. No restrictions on publication year were applied, and the review encompassed studies published in English.
Eligibility criteria
Both human and animal studies were included to provide a comprehensive evaluation: human studies offer direct clinical insights, while animal models allow investigation of mechanisms (e.g., gut–testis axis modulation, hormonal changes, and oxidative stress reduction) that are often unethical or infeasible in humans. This combined approach facilitates translational interpretations, especially given the limited human evidence.
Eligible studies involved oral probiotic interventions with clearly identified species in adult human males (≥18 years, with or without infertility) or mammalian animal models. Comparisons included placebo, no intervention, standard treatment, or alternative approaches. Primary outcomes were changes in sperm parameters (count, motility, morphology, viability, and DNA integrity). Secondary outcomes included mechanistic markers (antioxidant levels, inflammatory markers, and gut–testis axis modulation).
A range of designs was considered: randomized controlled trials, cohort, case-control, or quasi-experimental studies for humans; experimental studies for animals. Exclusions comprised in vitro-only studies, non-probiotic primary interventions, reviews, meta-analyses, editorials, conference abstracts, unpublished manuscripts, non-English publications, or studies with insufficient/isolation-unable data on sperm outcomes.
Search strategy
A systematic search was conducted with librarian assistance across PubMed/MEDLINE, Web of Science, Scopus, Embase, and Cochrane Library from inception to 21 November 2024. Additional searches included Google Scholar and gray literature (conference proceedings, dissertations, preprints) to minimize publication bias.
Search terms combined probiotics-related keywords (e.g., “probiotics,” “Lactobacillus,” “Bifidobacterium,” “gut microbiota”) with sperm parameter terms (e.g., “sperm,” “sperm count,” “sperm motility,” “sperm morphology,” “male infertility,” “seminal parameters”) using Boolean operators, truncation, and wildcards. No restrictions were applied on study design or population to capture both human and animal studies. Reference lists of included studies and relevant reviews were hand-searched. Records were imported into EndNote (version 10), and duplicates removed.
Study selection
Two independent reviewers (AShG and MRM) screened titles/abstracts and full texts against eligibility criteria. Disagreements were resolved by discussion or a third reviewer. Reasons for full-text exclusions were documented. The process is summarized in the PRISMA flow diagram (Figure 1).
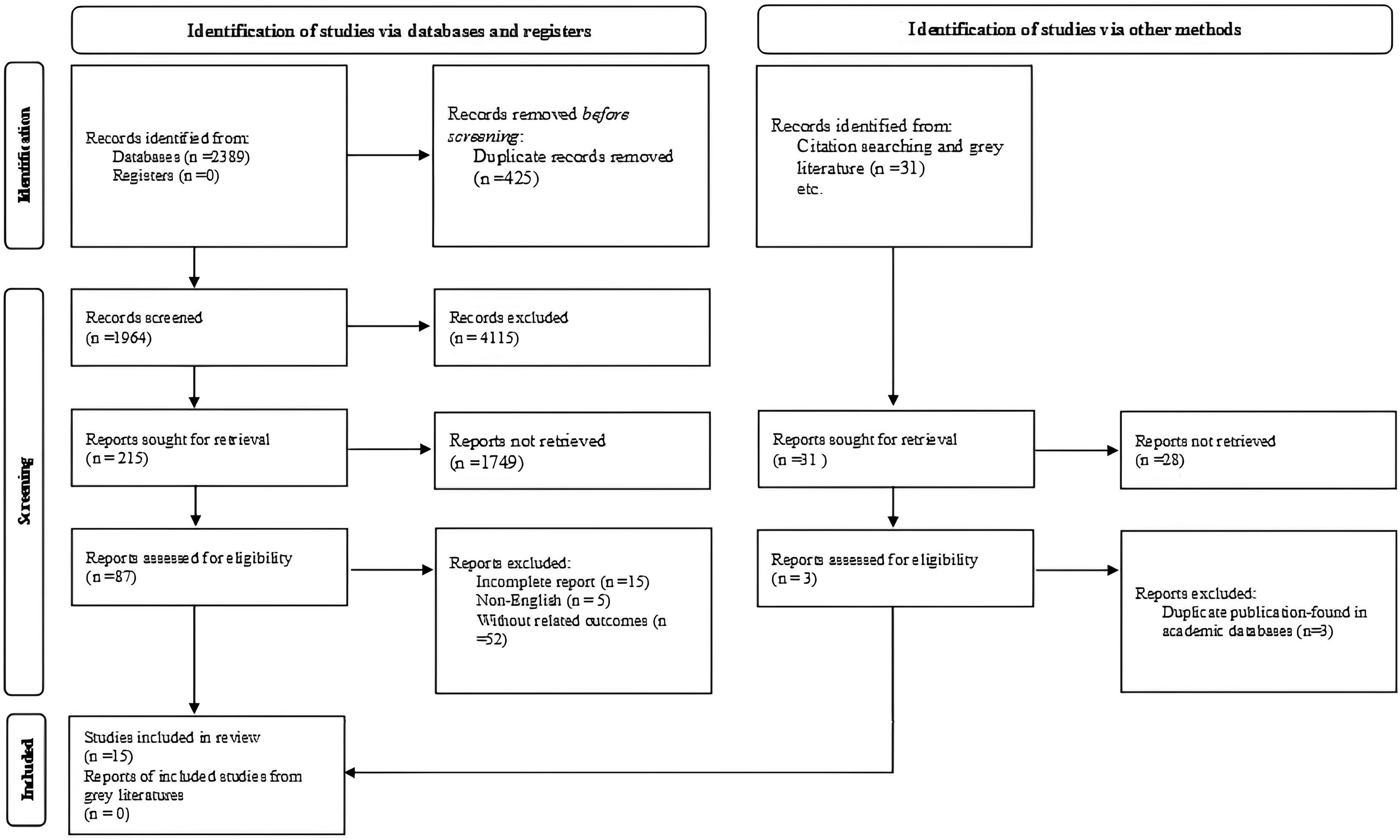
Preferred reporting items for systematic reviews flow diagram (2020) of the search process.
Data extraction
Two reviewers (AShG and RR) independently extracted data using a piloted standardized form, with disagreements resolved by consensus or a third reviewer. Extracted items included: study characteristics (author, year, country, design, sample size, population/model details); intervention details (species, dose, formulation, administration, duration); comparators; primary and secondary outcomes; results (including statistical significance); and additional information (funding, conflicts, limitations).
Quality assessment
The quality of the included animal studies was assessed using SYRCLE's RoB tool, which is specifically designed to evaluate methodological rigor in animal research. This tool is an adaptation of the Cochrane RoB tool, tailored to address features unique to animal experiments. Two independent reviewers conducted the quality assessment, and any discrepancies in judgments were resolved through discussion or by consulting a third reviewer.
The SYRCLE's RoB tool evaluates the RoB across ten domains, grouped into six categories: selection bias, performance bias, detection bias, attrition bias, reporting bias, and other biases. Each domain was rated as having a low, high, or unclear RoB based on the information provided in the study.
For human studies, the Cochrane RoB 1 tool was applied. This tool evaluates seven domains: random sequence generation, allocation concealment, blinding of participants and personnel, blinding of outcome assessment, incomplete outcome data, selective reporting, and other sources of bias. Each domain was rated as having a low, high, or unclear RoB. The same process of independent evaluation by two reviewers was followed, with disagreements resolved through consensus or by consulting a third reviewer.
The findings from both quality assessment tools were summarized to inform the reliability of the included studies. These assessments contributed to the interpretation of the results and the overall strength of the evidence synthesized in this review.
Data synthesis
Data were synthesized narratively, with results presented separately for human and animal studies to account for inherent differences in models and outcomes. A quantitative meta-analysis was deemed infeasible due to substantial clinical, methodological, and statistical heterogeneity across studies. Key sources of heterogeneity included variations in probiotic species (single- vs. multi-specie formulations), dosages, intervention durations, outcome definitions and measurement methods, participant/animal characteristics (e.g., idiopathic infertility vs. induced models of stress, diabetes, or toxicity), and study designs. The inclusion of both human clinical trials and preclinical animal studies further precluded direct statistical pooling, as outcomes are not directly comparable across species. Heterogeneity was therefore assessed qualitatively through stratified reporting and discussion of differences in interventions and findings, rather than formal statistical tests (e.g., I2 statistic or subgroup analyses).
Results
Literature search and study characteristics
The database searches yielded 2389 results. In total, 425 duplicates were removed, leaving 1964 for review. After title and abstract scanning, 87 articles were considered relevant and their full manuscripts were reviewed for inclusion. A total of 15 studies met the inclusion criteria, consisting of 5 human trials and 10 animal experiments (5 human studies: (Abbasi et al., 2021; Asadi et al., 2023; Dardmeh et al., 2017; Helli et al., 2022; Maretti and Cavallini, 2017; Valcarce et al., 2017)) and 10 animal studies (Akram et al., 2023; Belgacem et al., 2022; Chen et al., 2013; Cooke et al., 2024; Lee et al., 2024; Liu et al., 2024; Rahimiyan-Heravan et al., 2020; Sanchez-Rodriguez et al., 2024; Tian et al., 2019). Figure 1 summarized the studies selection process. Most interventions involved multi-species probiotics, with species from the genera Lacticaseibacillus, Lactiplantibacillus, Limosilactobacillus, and Bifidobacterium being predominant. Specific species were identified in several animal studies (e.g., Lacticaseibacillus rhamnosus GG, Lactiplantibacillus plantarum MON03, TW1-1).
Risk of bias assessment
In terms of animal studies RoB assessment, we used SYRCLE Guidelines (Table 1). Four studies reported on the method used for random sequence generation (Akram et al., 2023; Lee et al., 2024; Sanchez-Rodriguez et al., 2024; Tian et al., 2019). Method used for allocation concealment was unclear or with high RoB except one study (Lee et al., 2024). Only 3/10 studies didn’t report that baseline characteristics were similar in the probiotic and no probiotic groups (Chen et al., 2013; Lee et al., 2024; Rahimiyan-Heravan et al., 2020). None of the studies reported on measures that were used to house the animals randomly within the animal room. In terms of the blinding process of researchers and outcome assessors, most of the studies did not follow these approaches in a good way. Also, all studies reported complete follow-up data. In terms of the domain of selective reporting, all of the studies obtained low RoB.
Quality Assessment of the Included Animal Studies.
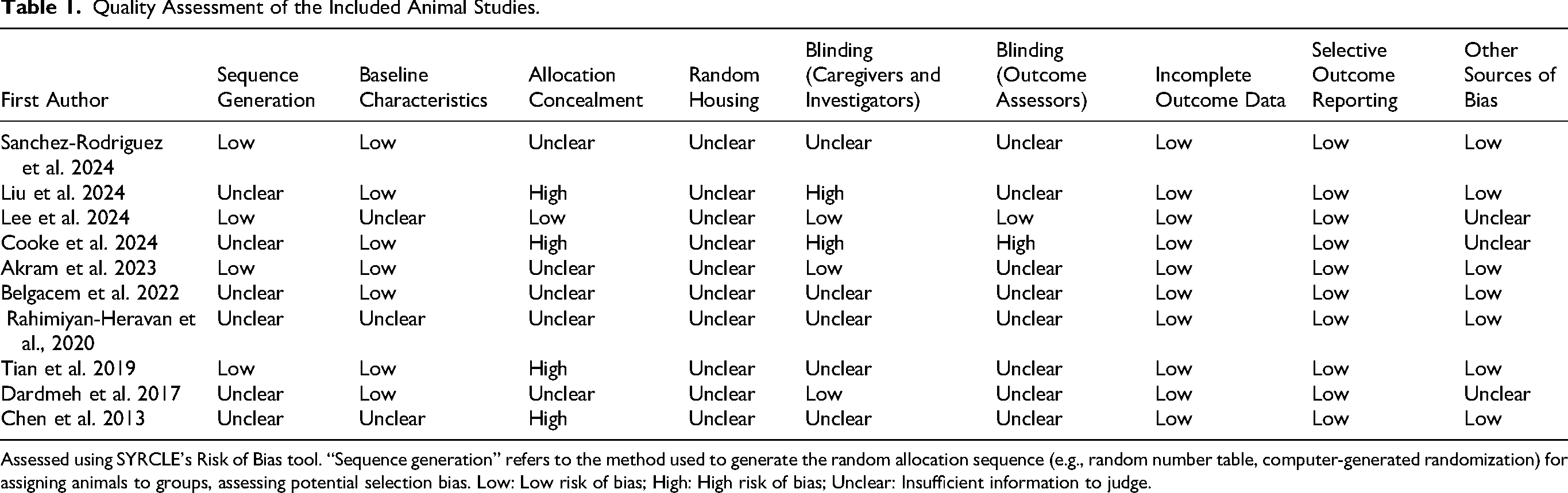
Assessed using SYRCLE's Risk of Bias tool. “Sequence generation” refers to the method used to generate the random allocation sequence (e.g., random number table, computer-generated randomization) for assigning animals to groups, assessing potential selection bias. Low: Low risk of bias; High: High risk of bias; Unclear: Insufficient information to judge.
In terms of RoB assessment among the human studies and based on random-sequence generation, three studies obtained low RoB (Abbasi et al., 2021; Helli et al., 2022; Maretti and Cavallini, 2017). None of the studies reported the allocation concealment approaches and in terms of blinding, three studies obtained low RoB (Abbasi et al., 2021; Asadi et al., 2023; Helli et al., 2022). Also, we found a good approaches in outcome assessors blinding only in two studies (Abbasi et al., 2021; Asadi et al., 2023). All of the studies had low RoB regarding incomplete outcome data, and selective reporting (Table 2).
Quality Assessment of the Included Human Studies.
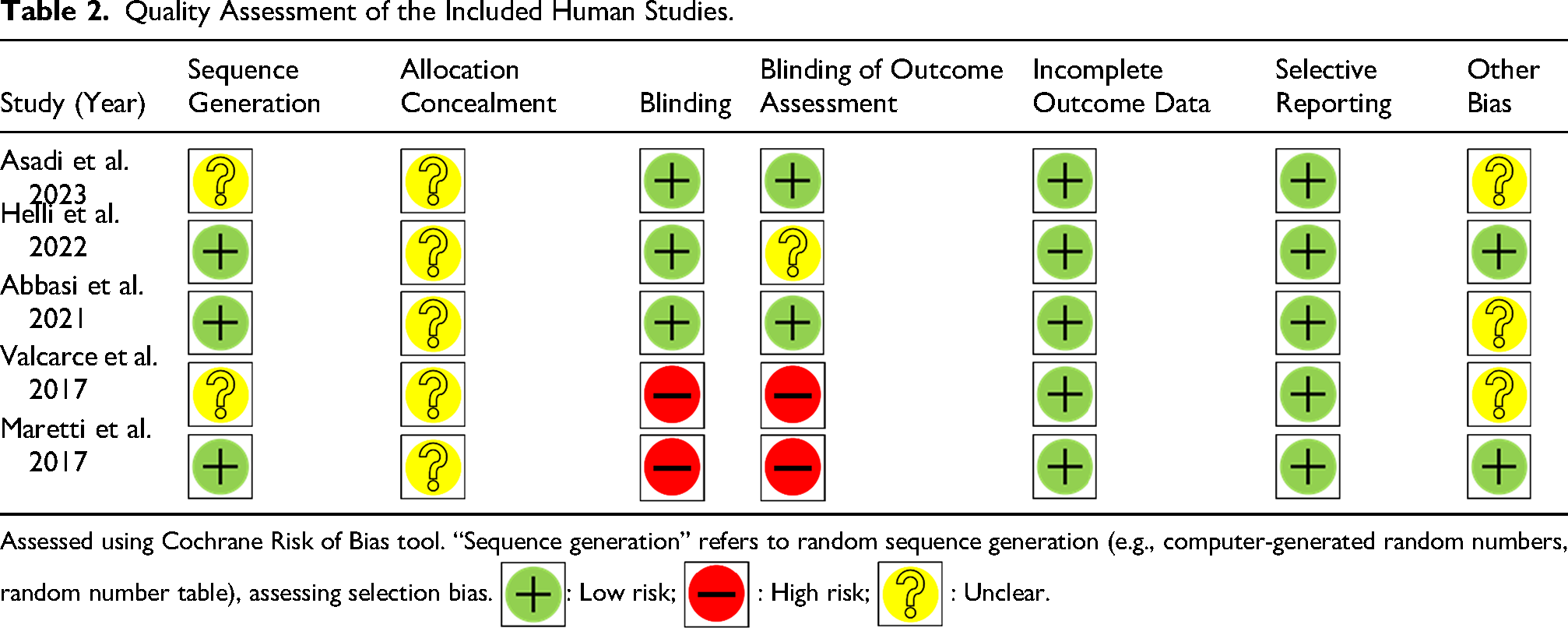
Assessed using Cochrane Risk of Bias tool. “Sequence generation” refers to random sequence generation (e.g., computer-generated random numbers, random number table), assessing selection bias.  : Low risk;
: Low risk;  : High risk;
: High risk;  : Unclear.
: Unclear.
The RoB assessment revealed several methodological limitations that may influence confidence in the synthesized evidence. In animal studies, the majority exhibited unclear or high RoB in allocation concealment and blinding of personnel and outcome assessors (Table 1). These domains are critical, as inadequate blinding can lead to performance and detection bias, potentially overestimating treatment effects. Only a minority of studies reported adequate random sequence generation and similar baseline characteristics, further contributing to potential selection bias (Table 1). Nevertheless, all animal studies had a low risk of selective reporting and complete outcome data, which strengthens confidence in the reported results. In human studies, while three out of five trials demonstrated low RoB in random sequence generation and blinding, allocation concealment was not reported in any study (Table 2). The absence of reported allocation concealment was a primary driver of the moderate to high overall RoB assessment in human studies. Blinding of outcome assessors was adequate in only two trials (Table 2).
These limitations suggest a moderate to high overall RoB across the included human studies. Collectively, the predominant risks in blinding and allocation concealment across both animal and human studies reduce confidence in the magnitude of the observed beneficial effects of probiotics on sperm parameters. While the consistency of positive findings across studies supports a potential true effect, the methodological shortcomings indicate that the current evidence should be interpreted with caution. These biases highlight the need for future high-quality randomized controlled trials with rigorous methodological reporting to confirm the efficacy of probiotic interventions in improving male reproductive health.
Experimental studies
Ten animal studies were included in this systematic review. These studies were conducted in China (Chen et al., 2013; Liu et al., 2024; Tian et al., 2019), Spain (Sanchez-Rodriguez et al., 2024), Republic of Korea (Lee et al., 2024), Australia (Cooke et al., 2024), India (Akram et al., 2023), Tunisia (Belgacem et al., 2022), Iran (Rahimiyan-Heravan et al., 2020) and Denmark (Dardmeh et al., 2017). Mouse models were predominant (n = 8), with additional rat (n = 1) and miniature pony stallion (n = 1) models (Table 3). Lacticaseibacillus rhamnosus was the most investigated species (used in five studies, either alone or in combination), followed by Lactiplantibacillus plantarum (n = 3). Intervention durations ranged from 5 days to 20 weeks. Formulations included both single-species and multi-species probiotics, with specific species identified in several studies (e.g., GG, MON03, TW1-1, NCDC-610, and NCDC-400).
Characteristics and Main Results of Experimental Studies.
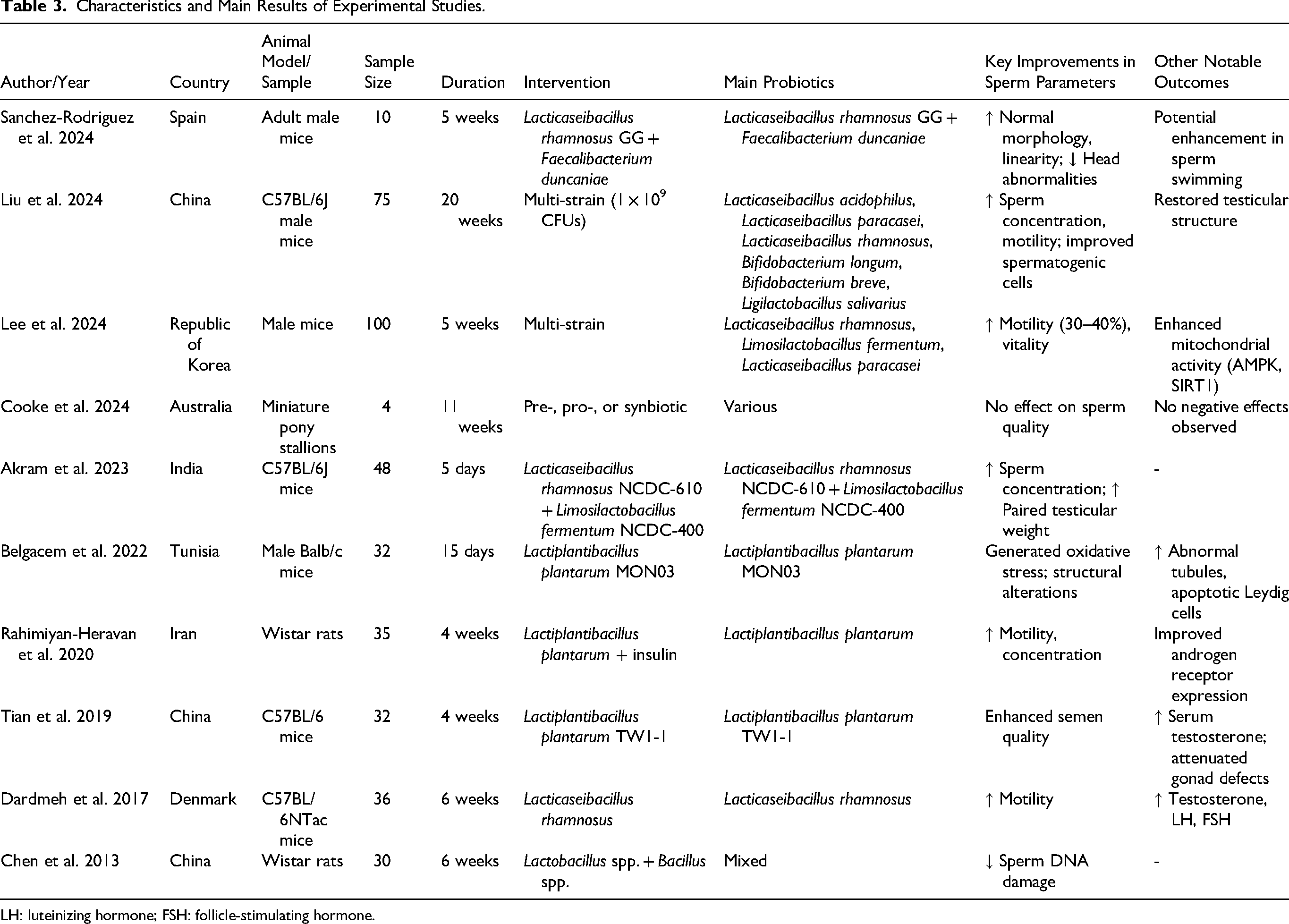
LH: luteinizing hormone; FSH: follicle-stimulating hormone.
As summarized in Table 3, probiotic supplementation led to improvements in sperm parameters in eight of the ten studies (Akram et al., 2023; Chen et al., 2013; Dardmeh et al., 2017; Lee et al., 2024; Liu et al., 2024; Rahimiyan-Heravan et al., 2020; Sanchez-Rodriguez et al., 2024; Tian et al., 2019). Commonly reported benefits included increased sperm concentration (n = 4 studies (Akram et al., 2023; Liu et al., 2024; Rahimiyan-Heravan et al., 2020; Tian et al., 2019)), enhanced motility (n = 5 studies (Dardmeh et al., 2017; Lee et al., 2024; Liu et al., 2024; Rahimiyan-Heravan et al., 2020; Sanchez-Rodriguez et al., 2024)), improved morphology (n = 3 studies (Akram et al., 2023; Liu et al., 2024; Sanchez-Rodriguez et al., 2024)), and increased vitality (n = 1 study (Lee et al., 2024)).
Additional positive outcomes in some studies involved higher testicular weight and elevated hormone levels (testosterone, luteinizing hormone (LH), follicle-stimulating hormone (FSH)). Two studies reported no significant effects on sperm quality (Cooke et al., 2024) or potential lack of improvement with structural changes (Belgacem et al., 2022). No adverse effects on sperm parameters were noted in the majority of studies.
In an experimental study, Sanchez-Rodriguez et al. (2024) assessed the impact of a probiotic mixture containing Lacticaseibacillus rhamnosus GG and Faecalibacterium duncaniae A2-165 on sperm quality in a mouse model. Over a 5-week period, probiotics were administered twice weekly to adult mice. Liu et al. (2024) evaluated the effects of a multi-species probiotic formula containing Lacticaseibacillus acidophilus, Lacticaseibacillus paracasei, Lacticaseibacillus rhamnosus, Bifidobacterium longum, Bifidobacterium breve, and Ligilactobacillus salivarius (1 × 109 CFUs) in mice over 20 weeks. Probiotic treatment, either alone or combined with metformin, was found to restore sperm quality and improve testicular structure. Similarly, Lee et al. (2024) investigated a multi-species probiotic mixture containing Lacticaseibacillus paracasei, Lacticaseibacillus rhamnosus, and Limosilactobacillus fermentum in mice. The probiotics significantly enhanced sperm motility and vitality, with motility increasing by 30–40% compared to untreated sperm.
In another study, Akram et al. (2023) explored the effects of Lacticaseibacillus rhamnosus NCDC-610 and Limosilactobacillus fermentum NCDC-400, administered alone or with the prebiotic fructooligosaccharides (FOSs), on restraint stress-induced reproductive deficits in mice. Probiotics improved sperm count, motility, and morphology, restored testicular histopathology, and enhanced intestinal health. Additionally, they upregulated steroidogenic proteins, reduced oxidative stress markers, and lowered inflammatory cytokines, further confirming their protective effects against stress-induced reproductive impairments. In terms of Lacticaseibacillus rhamnosus, Dardmeh et al. (2017) reported that supplementation with Lacticaseibacillus rhamnosus significantly increased serum testosterone, LH, and FSH levels, as well as sperm motility in mice.
Human studies results
The results of probiotic supplementation on sperm parameters among the human studies are summarized in Table 4. Among the five studies included in the present study, three were conducted in Iran (Abbasi et al., 2021; Asadi et al., 2023; Helli et al., 2022) and the rest in Italy (Maretti and Cavallini, 2017) and Spain (Valcarce et al., 2017). The sample size of included studies varied between 9 and 78. Also, intervention duration varied between 10 weeks and 6 months.
Characteristics and Main Results of Human Studies.
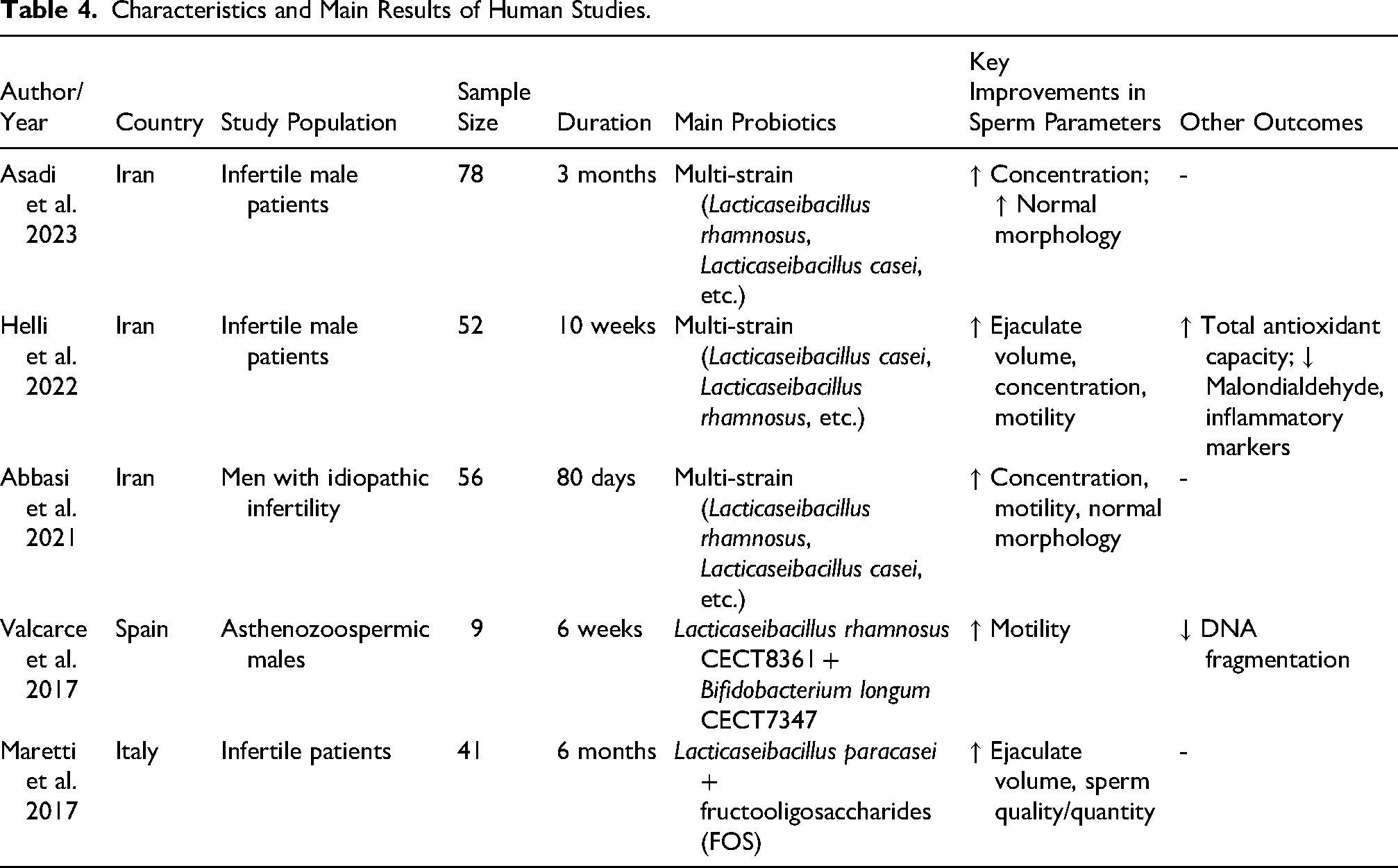
Asadi et al. (2023) in a double-blind randomized clinical trial examined the effects of multi-species probiotic supplement (including Lacticaseibacillus rhamnosus, Lacticaseibacillus casei, Lactobacillus delbrueckii subsp. bulgaricus, Lactobacillus acidophilus, Bifidobacterium breve, Bifidobacterium longum, and Streptococcus thermophilus) on sperm concentration, and the percentage of the sperms with normal morphology. They found that probiotic supplementation for 3 months led to a significant improvement in the sperm concentration and the percentage of the sperms with normal morphology in the intervention group. In line with this study, Abbasi et al. (2021) in a triple-blinded randomized clinical trial that used a similar probiotic product like in the study of Asadi et al. and they observed a considerable improvement in the mean sperm concentration, motility, and normal morphology. Moreover, in another study among infertile men, researchers prescribed a probiotic supplement to 52 infertile male patients for 10 weeks and they observed a significant improvement in ejaculate volume, number, concentration, percentage of motile sperm, and total antioxidant capacity of plasma and considerable reduction in the concentration of plasma malondialdehyde and inflammatory markers (Helli et al., 2022).
In another study among the Spanish infertile patients, Valcarce et al. (2017) evaluated the efficacy of a probiotic formula including Lacticaseibacillus rhamnosus and Bifidobacterium longum among the 9 asthenozoospermic males and the results of this study revealed a significant improvement in sperm motility and decrease in DNA fragmentation. Finally, Maretti and Cavallini (2017) examined the effects of Lacticaseibacillus paracasei plus FOS on sperm parameters among the infertile patients and they found a significant improvement in the volume of the ejaculate and the quality/quantity of spermatozoa.
Discussion
Male infertility is a growing concern, and it's becoming increasingly clear that environmental factors and lifestyle choices can significantly impact a man's reproductive health. One crucial factor is the delicate balance of the male reproductive system's microbiome (Katz et al., 2017). This systematic review synthesizes evidence from 15 studies (5 human RCTs and 10 animal studies), suggesting potential beneficial effects of probiotic supplementation on sperm count, motility, and morphology, particularly in individuals with baseline impairments. However, given the limited number of human trials, considerable heterogeneity in species, dosages, durations, and study designs, and the moderate to high RoB, these findings should be interpreted with caution.
The intricate community of microorganisms residing within the human gut, collectively known as the intestinal microbiota, plays a crucial role in promoting overall host health (Kataoka, 2016). This beneficial impact is achieved through a multifaceted approach, including: strengthening the protective lining of the gut, outcompeting harmful bacteria for attachment sites, modulating the immune response, and producing essential nutrients such as vitamins and short-chain fatty acids (SCFAs) (El-Sayed et al., 2021; Vajpeyee et al., 2021). Gut microbial dysbiosis, characterized by an imbalance in the composition of the gut microbiota, disrupts the beneficial interactions between the host and its microbial community, consequently leading to disruptions in the host's internal balance. By colonizing the gastrointestinal tract, probiotics exert a numerical advantage over pathogenic microorganisms, ultimately facilitating improvements in epithelial barrier function, immune system regulation, and the production of SCFAs and vitamins (Weiss and Hennet, 2017).
Probiotic microorganisms may exert beneficial influences beyond the confines of the gastrointestinal system. This broader impact is likely facilitated by intricate interactions that modulate the immune response and by the synthesis of neurotransmitters, suggesting a more systemic role for these beneficial bacteria (Tomaiuolo et al., 2020). In most of the included studies in this systematic review, researchers found a significant improvement in the sperm parameters following different probiotic species. The most common bacteria used in the included studies were species from the genera Lacticaseibacillus, Lactiplantibacillus, Limosilactobacillus, and Bifidobacterium, whose antioxidant properties were shown in various experiments (Saez-Lara et al., 2015). The systemic therapeutic potential of these specific strains is further supported by evidence in other clinical areas. For instance, a recent systematic review concluded that Lacticaseibacillus rhamnosus and Lactiplantibacillus plantarum are effective in modulating local inflammatory environments and microbial dysbiosis in oral health, including caries and periodontal diseases (D’Agostino et al., 2024). This suggests that the immunomodulatory mechanisms of these probiotics are robust across different physiological systems, reinforcing their potential utility in treating inflammation-driven conditions such as male infertility.
In line with these results, Almeida Gomes et al. (2023) in a systematic review that evaluated the association between microbiota and semen quality was reported a positive association between the presence of Lactobacillus species and normal semen parameters in the male population studied. In contrast, individuals exhibiting semen quality deviations demonstrated a higher prevalence of Prevotella species.
Disruptions to the delicate equilibrium of the male reproductive tract's microbial ecosystem can have profound consequences (Oghbaei et al., 2020). These alterations can pave the way for the development of infections, both those with overt symptoms and those that remain undetected. Such infections can exert a detrimental influence on spermatogenesis, the intricate process of sperm formation, or may even stimulate the production of harmful toxins (Pasqualotto, 2007). Furthermore, these microbial imbalances can contribute to a state of oxidative stress within the reproductive system (Jendraszak et al., 2024). Oxidative stress, characterized by an overabundance of reactive oxygen species, can inflict significant damage upon cellular structures and DNA, thereby compromising sperm quality and potentially diminishing fertility (Bhattacharya et al., 2023). Notably, the intricate interplay within a couple's shared microbial environment underscores the potential for these male reproductive tract disturbances to exert an indirect yet significant impact upon the female partner's reproductive health, potentially contributing to infertility in her (Asadi et al., 2021).
The normal microbiota of the male reproductive tract, particularly in semen, plays a pivotal role in maintaining reproductive homeostasis and protecting sperm function. In healthy, normospermic men, the seminal microbiome is typically characterized by a predominance of Lactobacillus species, often alongside genera such as Corynebacterium, Streptococcus, Staphylococcus, and Bifidobacterium. These commensal bacteria contribute to a protective environment by producing antimicrobial substances (e.g., lactic acid, bacteriocins), maintaining an acidic pH, and competing with potential pathogens. This composition is associated with normal sperm parameters and reduced oxidative stress. In contrast, dysbiosis—marked by enrichment of genera such as Prevotella, Anaerococcus, or certain Enterobacteriaceae—is frequently observed in men with impaired semen quality, potentially exacerbating inflammation, reactive oxygen species production, and sperm DNA damage (Chatzokou et al., 2025; Farahani et al., 2021; Gomes et al., 2023).
Probiotics from genera such as Lacticaseibacillus (formerly Lactobacillus), Limosilactobacillus, and Bifidobacterium have been shown to improve intestinal barrier integrity, preventing the translocation of endotoxins such as lipopolysaccharides (LPS) into systemic circulation (Hajian et al., 2024). LPS are endotoxins found in the outer membrane of gram-negative bacteria. When LPS enter systemic circulation, they trigger a strong inflammatory response by activating Toll-like receptor 4 signaling pathways (Bhandari et al., 2016). This results in the release of pro-inflammatory cytokines, such as tumor necrosis factor-alpha and interleukins (e.g., IL-1β, IL-6). Chronic inflammation and oxidative stress caused by elevated LPS levels are detrimental to sperm cells, as they can lead to DNA fragmentation, reduced motility, and impaired morphology (Guo et al., 2020; Rahimlou et al., 2022). Probiotics mitigate these effects by decreasing gut-derived LPS levels and reducing inflammation, indirectly improving sperm quality. In line with these mechanical insights, in a cross-sectional study, Voroshilina et al. (2021) analyzed 227 semen samples from men with normal sperm parameters (normospermia). A significant finding was that approximately half of these samples exhibited a predominance of three bacterial genera: Lactobacillus, Enterobacteriaceae, and Bifidobacterium.
The Hypothalamic–Pituitary–Gonadal (HPG) Axis is a hormonal system that regulates reproductive function. Probiotics have been shown to influence this axis by interacting with the gut–brain axis, a bidirectional communication network between the gut and the central nervous system (Rafati et al., 2023; Rani et al., 2023). Probiotic metabolites, such as SCFAs, can stimulate the production of gonadotropin-releasing hormone (GnRH) in the hypothalamus. GnRH, in turn, triggers the pituitary to release LH and FSH, which are essential for testosterone synthesis and spermatogenesis in the testes (Akram et al., 2023; Asadi et al., 2020; Eghbali et al., 2023).
Although the present systematic review focused exclusively on in vivo human and animal studies, complementary evidence from in vitro experiments highlights potential direct effects of probiotic bacteria on semen parameters. Co-incubation of human or animal sperm with Lacticaseibacillus strains (or their metabolites/postbiotics) has been shown to improve progressive motility, viability, and kinematic parameters while reducing DNA fragmentation and oxidative damage, without adverse effects at moderate concentrations. These findings suggest direct protective mechanisms—such as antioxidant activity and pathogen inhibition—independent of systemic gut-mediated pathways, providing additional support for the therapeutic potential of probiotics in male reproductive health (Kotarska, 2024; Mousavi et al., 2022; Rahimlou et al., 2024; Wang et al., 2020).
This study offers several strengths that contribute to its novelty and reliability. Firstly, the systematic inclusion of diverse primary studies (15 studies: 5 human and 10 animal) allows for a comprehensive evaluation of the association between probiotics, gut health, and male reproductive outcomes, extending beyond prior reviews. Notably, compared to a recent systematic review by Oliveira et al.(2024), which focused exclusively on four human randomized clinical trials and provided limited mechanistic insights, the present review uniquely incorporates substantial preclinical evidence from animal models. This enables deeper exploration of underlying mechanisms, such as gut–testis axis communication, HPG axis modulation, and systemic reductions in oxidative stress and inflammation. Furthermore, our literature search was updated to November 2024, capturing additional recent studies (Amirkhizi et al., 2023; Hasanloei et al., 2021). By incorporating insights into mechanistic pathways such as intestinal barrier integrity and modulation of the HPG axis, alongside rigorous quality assessments and stratified analyses (human vs. animal), the discussion advances the understanding of the multifaceted role of probiotics in reproductive health, enhances the credibility and generalizability of the findings across different models, and provides a stronger foundation for future translational research.
Despite these strengths, important limitations must be acknowledged. The small number of human RCTs (only five, with sample sizes ranging from 9 to 78 participants) limits statistical power and generalizability. High heterogeneity in probiotic species, dosages, durations, populations, and study designs prevented quantitative meta-analysis and complicates direct comparisons. Many human studies originated from a single region (predominantly Iran), introducing potential geographic bias. The moderate to high RoB (particularly in allocation concealment and blinding) may have overestimated effects. Additionally, the lack of long-term data, standardized protocols, and direct mechanistic evidence in human further tempers confidence in the findings.
Furthermore, the search was restricted to English-language publications (with five non-English reports excluded during screening), potentially introducing language bias and missing relevant studies published in other languages. Publication bias is another concern, as studies with positive or significant results are more likely to be published than those with null or negative findings; however, formal assessment (e.g., via funnel plots or Egger's test) was not feasible due to the small number of included studies and absence of meta-analysis. Furthermore, a substantial limitation is the incomplete reporting of probiotic strain details in most included studies. While a few investigations identified specific strains (e.g., Lacticaseibacillus rhamnosus GG, Lactobacillus plantarum MON03, or Bifidobacterium longum CECT7347), the majority provided only species-level information without strain acronyms. Given that the health benefits of probiotics are frequently strain dependent, this reduces the ability to accurately compare outcomes across studies, assess reproducibility, and formulate precise therapeutic guidelines.
A key limitation is the reliance on surrogate biomarkers—such as sperm parameters (count, motility, morphology, DNA integrity) and biochemical markers (e.g., total antioxidant capacity, inflammatory markers) rather than direct clinical fertility outcomes, such as pregnancy rates or live birth rates. While improvements in sperm parameters are relevant proxies for male reproductive health, they do not necessarily translate to enhanced fertility or successful conception in clinical practice.
Conclusion
This systematic review provides preliminary evidence from a limited number of studies suggesting that probiotic interventions may improve sperm parameters, potentially through mechanisms involving reduced oxidative stress, inflammation, and gut–testis axis modulation. However, the evidence—particularly from human trials—is constrained by the small number of RCTs, high heterogeneity in species and protocols, methodological limitations (including RoB), and reliance on surrogate outcomes rather than direct fertility measures.
At present, the available data are insufficient to support the use of probiotics as a clinical intervention for male infertility or to inform practice guidelines. Large-scale, multicenter, standardized, high-quality randomized controlled trials are essential to confirm any potential benefits, establish optimal species and dosages, evaluate long-term safety, and assess translational impact on clinical fertility outcomes (e.g., pregnancy and live birth rates) in diverse populations.
Footnotes
Ethical approval
Not applicable.
Consent for publication
Not applicable
Authors’ contributions
AShG, RR, and MRM designed the research. AShG, MShJ, SA, and MRM conducted the search and screening process. AShG, MRM, and RR extracted the data. MRM and SA wrote the article. RR had primary responsibility for the final content. All authors read and approved the final manuscript.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Availability of data and material
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.