
Abstract
The complement system consists of at least 50 proteins that serve as one of the first lines of defence against foreign, or damaged, cells and invading microorganisms. Its dysregulation underlies the pathophysiology of many different diseases, which makes functional assays of complement activity crucial; they are, however, underutilised. Standard haemolysis assays for the analysis of complement function employ sensitised non-human erythrocytes (e.g. from the sheep, guinea-pig or rabbit), the use of which raises animal welfare concerns. To provide an alternative to the use of such animal-derived products for complement function assays, we developed a method that employs modified human erythrocytes to evaluate the activity of complement pathways. Human erythrocytes were subjected to various chemical and/or proteolytic treatments involving 2,4,6-trinitrobenzene sulphonate (TNBS) and pancreatin. Haemolysis assays demonstrated that sequential treatment with TNBS and pancreatin resulted in significantly greater complement-mediated haemolysis, as compared to TNBS or pancreatin treatment alone. Evidence that lysis of the modified erythrocytes was complement-mediated was provided by the chelation and subsequent restoration of calcium in the plasma. Thus, such modified human erythrocytes could be used as an alternative to animal-derived erythrocytes in haemolysis assays, in order to evaluate complement activity in human plasma during, for example, the screening of patients for complement deficiencies and other abnormalities in a clinical setting.
Keywords
Introduction
The complement system plays an integral role in both innate and adaptive immune responses. It serves as a first line of defence against foreign, or damaged, cells and invading microorganisms. Consisting of at least 50 proteins, the complement system is activated via three known pathways — the classical pathway (CP), the alternative pathway (AP) and the lectin pathway (LP) — all of which generate C3 convertases that proteolytically cleave complement component C3 to yield fragments C3a and C3b. The C3b fragment subsequently binds to existing C3 convertases to form C5 convertases, which in turn proteolytically cleave complement component C5 to yield fragments C5a and C5b. Thus, the C5 convertases initiate the final common pathway of complement activation, which culminates in the formation of the terminal membrane attack complex (MAC) that causes target cell lysis. 1,2 The complement cascade also mediates other homeostatic and effector functions, including opsonisation, tissue repair, angiogenesis and coagulation. 3
Mechanistically, the complement system is reminiscent of the blood coagulation system in that, in both systems, activation entails a cascade of proteolytic reactions catalysed by serine proteases (most notably, in the complement system, the C3 and C5 convertases themselves). 4 –6 Moreover, both systems can interact to form a positive feedback loop, whereby complement activation favours coagulation (via C5a-induced upregulation of tissue factor expression in leucocytes) that in turn drives further complement activation (via cleavage of C3 and C5 by thrombin, factors IXa, Xa, XIa, and plasmin). 6,7 This can result in a vicious circle of hypercoagulability and complement-mediated damage.
Given the potential of the complement system to cause cell damage, it is unsurprising that maladaptive activation underlies a range of pathological conditions and processes that are relevant globally in terms of their human cost. 8,9 As a case in point, increasing evidence suggests that a maladaptive immune response including complement C3 activation promotes the exacerbation of acute respiratory distress syndrome (ARDS) in the 2019 coronavirus disease (Covid-19). Such findings have contributed to the emergence of the complement system as a therapeutic target, 10 –12 and have also highlighted the need for methods to evaluate complement activity in human plasma. These methods could be applied to the screening of complement deficiencies, the monitoring of complement activity in disease states and the evaluation of therapeutic strategies to control complement activation. While these goals can be achieved with functional complement assays, exemplified by haemolysis assays, 13 such assays typically utilise erythrocytes from non-human animal sources (e.g. sheep, guinea-pig or rabbit). 6,14 –16 This presents an ethical problem, in that the use of animals for these assays is arguably unnecessary. In addition, their use may yield results that are potentially misleading, as the object of study is human immune function.
In principle, these animal erythrocytes could be replaced with human erythrocytes in haemolysis assays, but normal human erythrocytes are not susceptible to haemolysis mediated by autologous complement activity. This is due to the presence of the regulatory proteins CD55 (decay-accelerating factor (DAF)), CD59 (membrane attack complex-inhibiting protein (MAC-IP)) and C8-binding protein (C8 bp), which are attached to the erythrocyte membrane via glycosylphosphatidylinositol (GPI) anchors and protect against autologous complement-mediated haemolysis. The inactivation of complement regulatory proteins through proteolytic treatment has been shown to transform normal human erythrocytes into complement-sensitive ones, mimicking those in paroxysmal nocturnal haemoglobinuria (PNH). This disease is caused by a genetic mutation in the phosphatidylinositol glycan-class A gene, whose protein product is involved in the synthesis of GPI anchors. 17,18 In vitro complement-mediated lysis of erythrocytes from PNH patients occurs more efficiently at an acidic pH of 6.3 to 6.5. 18,19 While the inactivation and/or elimination of these GPI-anchored proteins from the surface of erythrocytes could permit the initiation of the AP through the slow spontaneous hydrolysis of C3, the other pathways (i.e. the CP and LP) require activators, such as immune complexes, for the complement cascade to be initiated. Despite having different triggering molecules, the CP and the LP both activate the same complement factors of the complement cascade. The functional activity of the CP and the LP have been measured by pre-coating sheep erythrocytes with antibodies and mannan, respectively. 14 Typically, the CP is activated when C1q, a component of the C1 complex, binds to antigen–antibody complexes. However, other agents such as C-reactive protein (CRP) can also induce CP activation. 20 On another note, a study on human and sheep erythrocytes has demonstrated that they can be labelled with haptenic groups that trigger CP activation in an antibody-independent manner. 21
Pancreatin is an enzyme preparation that contains a mixture of trypsin and chymotrypsin, which are homologous serine proteases. Despite their common evolutionary origin, these two enzymes exhibit different substrate preferences: whereas trypsin tends to cleave peptide bonds on the carboxyl terminal side of basic amino acid residues, chymotrypsin tends to cleave peptide bonds on the carboxyl terminal side of large hydrophobic amino acid residues. 22 As a pharmaceutical grade commercial product that is used for digestive and metabolic disorders, 23 pancreatin is widely available and can easily be obtained at a lower cost compared to laboratory grade protease preparations. To the best of our knowledge, the use of pancreatin as an agent for the modification of human erythrocytes to render them sensitive to human complement-mediated lysis has not been reported in the scientific literature.
The reagent 2,4,6-trinitrobenzene sulphonate (TNBS) is commonly used in amino acid analysis, due to its reaction with the amino groups of proteins and phospholipids. It is widely employed for the detection and quantification of protein amino groups. 24 TNBS dissolved in bicarbonate buffer (pH 8.6) has been observed to react optimally with the primary amino groups of erythrocyte cell surface proteins through nucleophilic aromatic substitution — with a reaction time of 30 to 120 minutes at 4°C. Under these conditions, TNBS is able to react with primary amino groups of the cell surface proteins, but it does not penetrate intact cell membranes. However, longer incubations result in the penetration of TNBS through the membrane, reaching the inner leaflet of the lipid bilayer and haemoglobin. 25 In view of these properties, as well as ease of use and availability, TNBS was selected as the chemical treatment agent for the human erythrocytes used in the current study.
Through combination of these proteolytic and chemical treatments, the aim of the current study was to develop an effective method for modifying human erythrocytes to render them susceptible to complement-mediated haemolysis when exposed to human plasma. This human-based method could then be used for the evaluation of total complement and specific AP activity.
Materials and methods
Blood product processing
Whole blood was collected via venepuncture from a healthy blood type O+ female donor (written informed consent was obtained) and placed in 15-ml polypropylene tubes (10 ml whole blood per tube); this blood was used as the source of erythrocytes. To prevent coagulation, 1.4 ml citrate phosphate dextrose adenine (CPDA-1) solution 26 was immediately added to each tube and the contents gently mixed. The separation of plasma was performed by light-spin centrifugation at 1500 g for 15 minutes. After removing the plasma and buffy coat by pipetting, the remaining packed erythrocytes were stored at 4°C, for up to 7 days after blood extraction, until further use in experiments.
Twenty fresh frozen plasma (FFP) units (blood type O+) were obtained from the Philippine General Hospital Blood Bank for use as complement source in this study. The FFP units were stored as 1 ml aliquots in polypropylene microcentrifuge tubes at –20°C until further use. Prior to the experiments, a 1 ml aliquot from each of the 20 FFP units was thawed and then pooled, giving a total pooled plasma volume of 20 ml.
Optimisation of pancreatin treatment
Initially, pancreatin treatment was optimised to determine the most appropriate concentration and incubation period to be used in subsequent experiments (this stage did not involve the use of TBNS). Pancreatin solutions were prepared in phosphate buffered saline, pH 7.4 (PBS) at 2.5%, 5% and 10% w/v; and 200 µl of the previously prepared packed normal human erythrocytes were incubated with 1 ml of each pancreatin solution at 37°C, for either 4 or 16 hours. The erythrocytes were washed three times with 10 ml PBS, and were resuspended to a final concentration of 1% v/v in HEPES buffered saline, pH 7.4 (HBS) prior to incubation with the pooled citrated human plasma (from 20 donors) as the complement source. The pooled citrated human plasma was diluted in acidic HEPES buffered saline (at pH 6.3) to maximise activation of the AP and determine the pancreatin treatment conditions that would result in the highest haemolysis. All subsequent experiments were performed at a physiological pH of 7.4.
Modification of human erythrocytes
In light of the results obtained with the pancreatin treatment optimisation experiment, throughout the study, human erythrocytes were incubated in a 5% w/v solution of pancreatin (Natural Organics Laboratories, Inc., Amityville, NY, USA; Universal Product Code 097467043800) in PBS, at 37°C for 4 hours. All erythrocyte samples were washed three times in 10 ml PBS before use in further treatments or assays.
Given the narrow range of time and temperature conditions previously outlined for the optimal reaction of TNBS with only the surface protein amino groups located externally on the erythrocytes, treatment of the human erythrocytes with TNBS was performed according to a published method. 25 Briefly, 200 µl of packed normal human erythrocytes were incubated with 8 ml of 2 mM TNBS (Sigma-Aldrich, St Louis, MO, USA; P2297) prepared in buffer A (120 mM sodium bicarbonate and 40 mM sodium chloride, pH 8.6), at 4°C for 1.5 hours. Prior to use in experiments, the erythrocytes were washed three times with buffer A.
TNBS (1.5-hour incubation at 4°C) and pancreatin (4-hour incubation at 37°C) treatments were performed individually or in series (TNBS treatment before pancreatin). The erythrocytes were resuspended to a final concentration of 1% v/v in HBS prior to incubation with the human plasma (which served as the complement source).
Haemolysis assay
Erythrocytes subjected to different treatments were compared in terms of their susceptibility to haemolysis mediated by human complement. Haemolysis was assessed directly (i.e. without a centrifugation step) in a 96-well microtitre polystyrene plate (Kartell 2620) by measuring absorbance at 650 nm, which was used to indicate haemolysis as a result of light scattering due to the remaining intact erythrocytes in each well, according to the protocol of Muzykantov et al., 27 with modifications. Briefly, 50 µl of complement source (i.e. pooled plasma, serially diluted in the appropriate buffer) were added to each microtitre plate well. For total complement activity, gelatin–HEPES buffer with calcium and magnesium, pH 7.4 (GHB++) was used, while gelatin–HEPES buffer with magnesium and EGTA (a stronger chelator of Ca2+), pH 7.4 (GHB Mg EGTA) was used for the specific evaluation of the calcium-independent AP. The same volumes of 1% v/v Triton™ X-100 and HBS were added to separate wells, and served as the positive and negative controls, respectively. To account for the turbidity of the plasma, the optical density at 650 nm (OD650) in each well was first measured by using a microplate reader (Thermomax, Sunnydale, CA, USA) prior to the addition of 50 µl of 1% erythrocyte suspension (v/v) in HBS. After a 1-hour incubation, the OD650 was measured again. The OD650 values obtained before incubation were subtracted from the corresponding OD650 values after incubation. The percentage haemolysis was calculated by using the following formula:
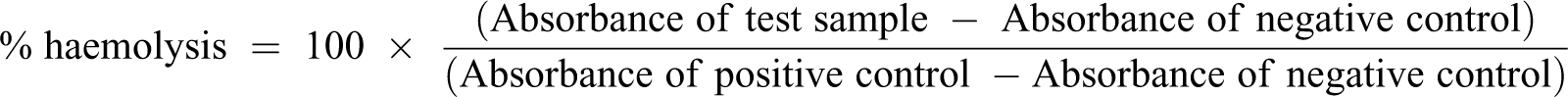
The CH50 (total complement activity) and AH50 (specific AP activity) values represent the respective volumes of plasma required for 50% haemolysis.
Inhibition of complement activity in human plasma
To demonstrate that the haemolysis of the modified human erythrocytes was being mediated by the complement system, complement activity was inactivated by using known methods. Varying amounts of EDTA were added to the plasma in order to chelate calcium, which is necessary for activation of the CP. Haemolysis of the modified human erythrocytes in the presence of complement-inactivated plasma was compared to that of normal plasma, to further demonstrate that lysis of the modified human erythrocytes was complement-mediated.
Statistical analysis
All statistical analyses were performed with GraphPad Prism 8. Four-parameter logistic (4PL) curve-fitting was performed to determine the CH50 and AH50 values for conditions that achieved ≥ 50% haemolysis. Since not all conditions yielded 50% haemolysis, one-way analysis of variance (ANOVA) with Tukey’s multiple comparisons was employed to compare the different treatments. A p value < 0.05 was considered statistically significant.
Results
Pancreatin treatment optimisation
Treatment with 10% pancreatin for 16 hours resulted in significant haemolysis and loss of intact erythrocytes. As a certain quantity of intact erythrocytes within the sample was required for the experiments, this set of conditions (i.e. concentration and length of incubation) was subsequently excluded.
The human erythrocytes were treated with various concentrations of pancreatin for 4 or 16 hours, prior to incubation with increasing amounts of the pooled plasma (i.e. the source of complement). A haemolysis profile, plotted in relation to the amount of plasma added to the pre-treated sample, was obtained for each of the pancreatin treatment conditions and used to compare their efficacy (Figure 1). The various pancreatin treatment conditions tested did not result in significantly different results. However, erythrocytes treated with 2.5% pancreatin for 4 hours led to a notably lower maximum haemolysis (83.7%) compared to the other treatments. Hence, based on the pancreatin optimisation experiment (which did not involve TNBS), treatment with 5% pancreatin for 4 hours was selected for all subsequent experiments.
A comparison of complement-mediated lysis profiles of human erythrocytes pre-treated with pancreatin solution under various conditions. Erythrocytes were treated with a pancreatin solution (2.5%, 5% or 10% (w/v)) for 4 or 16 hours, prior to incubation with acidified citrated human plasma pooled from 20 donors (at pH 6.3). No TNBS treatment was included in this optimisation experiment. No significant differences were observed among the pancreatin treatment conditions. However, the lowest percentage of haemolysis was observed in erythrocytes treated with 2.5% pancreatin for 4 hours.
Sensitivity of modified human erythrocytes to human complement-mediated haemolysis
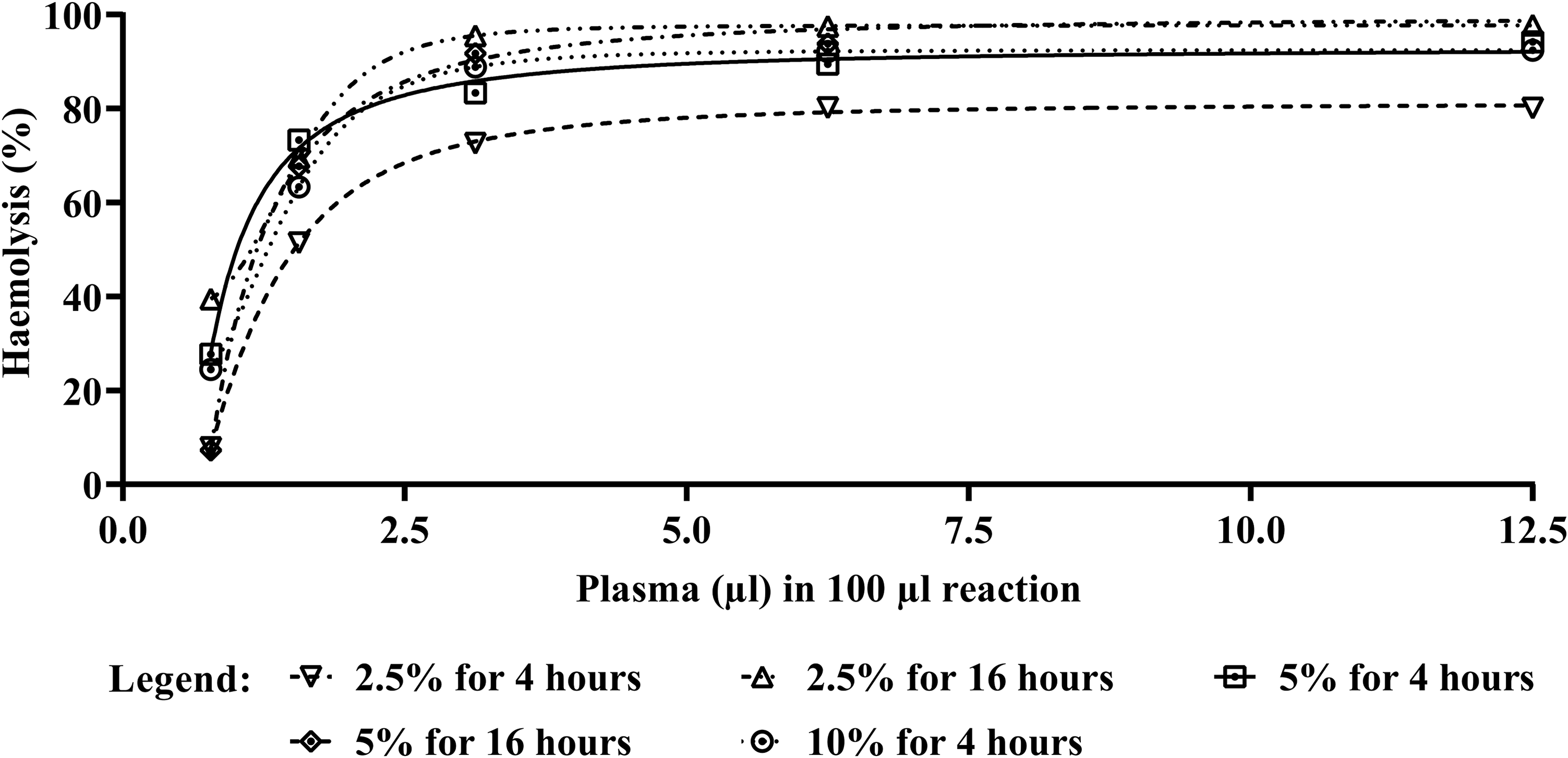
Haemolysis was observed in human erythrocytes pre-treated with TNBS and pancreatin according to the optimised conditions (TP-Es), with a CH50 value of 1.01 µl and a level of 96.10 ± 1.98% haemolysis achieved at the highest plasma concentration. Exposing pancreatin only-treated human erythrocytes to plasma led to some haemolysis, but the level did not reach 50% (maximum observed haemolysis was 35.28 ± 11.41%), even at the highest plasma concentration tested (12.5%). The effect of reaction pH (vis-à-vis maximising AP activity) on the extent of haemolysis is also noteworthy here: During the pancreatin optimisation step (see Figure 1), erythrocyte exposure to plasma was carried out at the acidic pH of 6.3 in order to maximise AP activity. The extent of haemolysis of the 5%/4-hour treated erythrocytes in this case was very much higher than that of identically treated erythrocytes exposed to plasma at the physiological pH of 7.4, as can be seen in Figure 2.
A comparison of the extent of complement-mediated lysis of human erythrocytes after various modification protocols. Human erythrocytes were modified by treating with 5% pancreatin only (for 4 hours at 37°C), 2 mM TNBS only (for 1.5 hours at 4°C), or a combination of both, prior to incubation with citrated human plasma pooled from 20 donors (at pH 7.4). Each data point represents the mean of three technical replicates with error bars indicating the standard error of the mean (SEM). The TNBS–pancreatin sequential treatment resulted in the most extensive haemolysis, with a CH50 value of 1.01. All of the other treatments did not lead to 50% haemolysis upon incubation with pooled human plasma, hence the absence of CH50 values. The asterisks indicate significant differences between the treated and untreated human erythrocytes (*p < 0.05; ****p < 0.0001).
Untreated and TNBS only-treated erythrocytes did not exhibit sensitivity to complement-mediated lysis when exposed to human plasma (Figure 2). Statistical analysis showed that the percentage haemolysis exhibited by TNBS–pancreatin-treated human erythrocytes was significantly higher than that of untreated erythrocytes (p < 0.0001), and of those treated with TNBS only (p < 0.0001) or with pancreatin only (p = 0.0037). These differences applied to most of the plasma proportions tested.
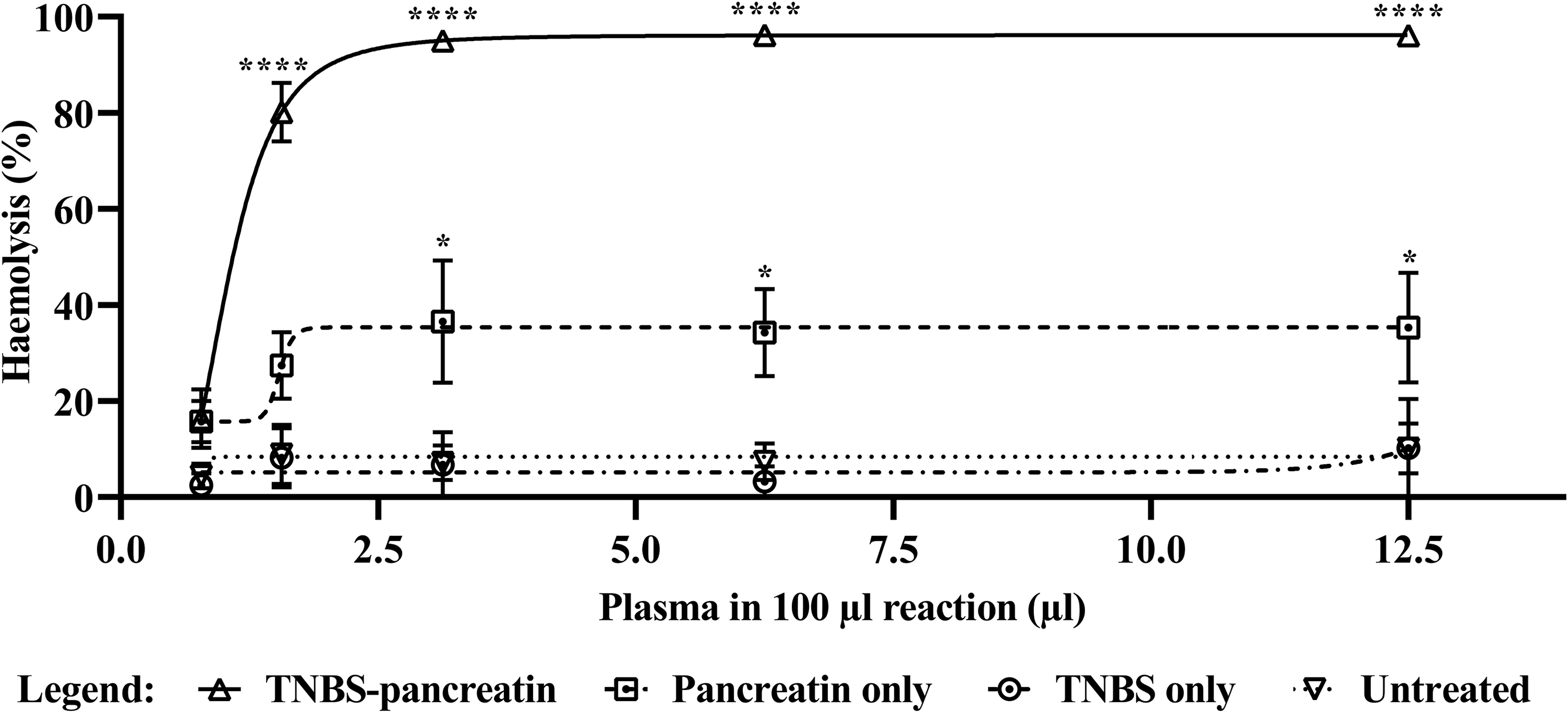
TP-Es were also used to evaluate the specific activity of the AP, by using GHB Mg EGTA (instead of GHB++) for serial dilution of plasma to support only activation of the AP. In this experiment, it was shown that TP-Es are also sensitive to AP-mediated haemolysis, with an AH50 value of 6.67 µl and a level of 56.79 ± 0.48% haemolysis achieved at the highest plasma concentration (Figure 3).
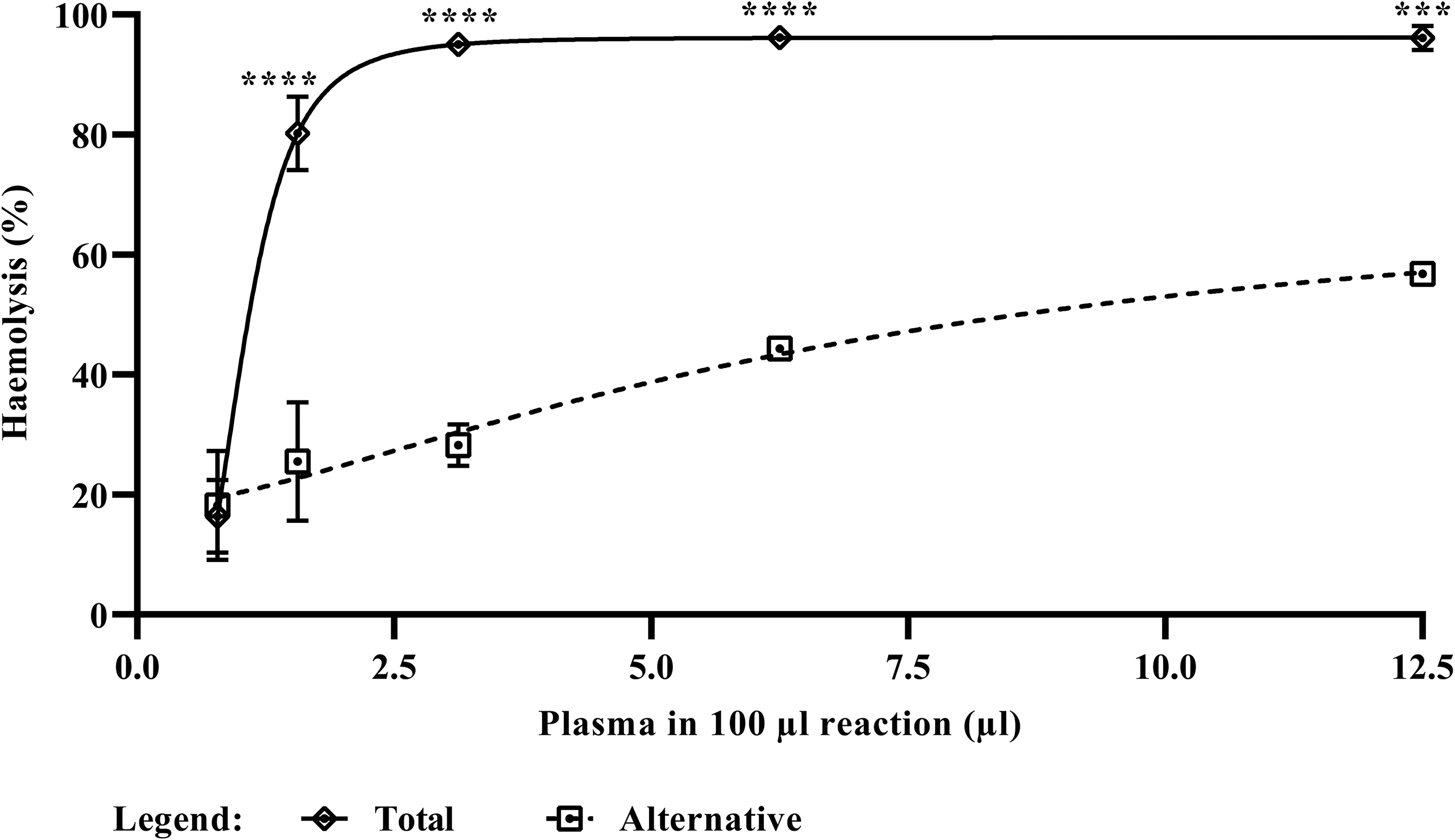
Differential activation of the total complement system or the alternative pathway. A comparison of the complement-mediated lysis of TP-Es after incubation with pooled citrated human plasma (from 20 donors), in either GHB++ (for the assessment of total complement activity) or GHB Mg EGTA (for the assessment of alternative pathway (AP) activity). Each data point represents the mean of three technical replicates with error bars indicating the standard error of the mean (SEM). The TP-Es were also observed to be sensitive to AP-mediated haemolysis, but to a lesser extent (AH50 = 6.67 μl). The asterisks indicate significant differences between total complement activity and AP activity (***p < 0.001; ****p < 0.0001).
Calcium-dependency of the observed effects
To confirm that haemolysis of the TP-Es was complement-mediated, EDTA was used to chelate the calcium cation, which is required for activation of the CP. The chelation of divalent cations through the addition of EDTA resulted in a concentration-dependent decrease in haemolytic activity, achieving a 50% inhibition of haemolysis with 3.82 mM EDTA. This inhibitory effect was reversed by supplementing the plasma with calcium ions, which resulted in the restoration of the plasma’s haemolytic activity (Figure 4).
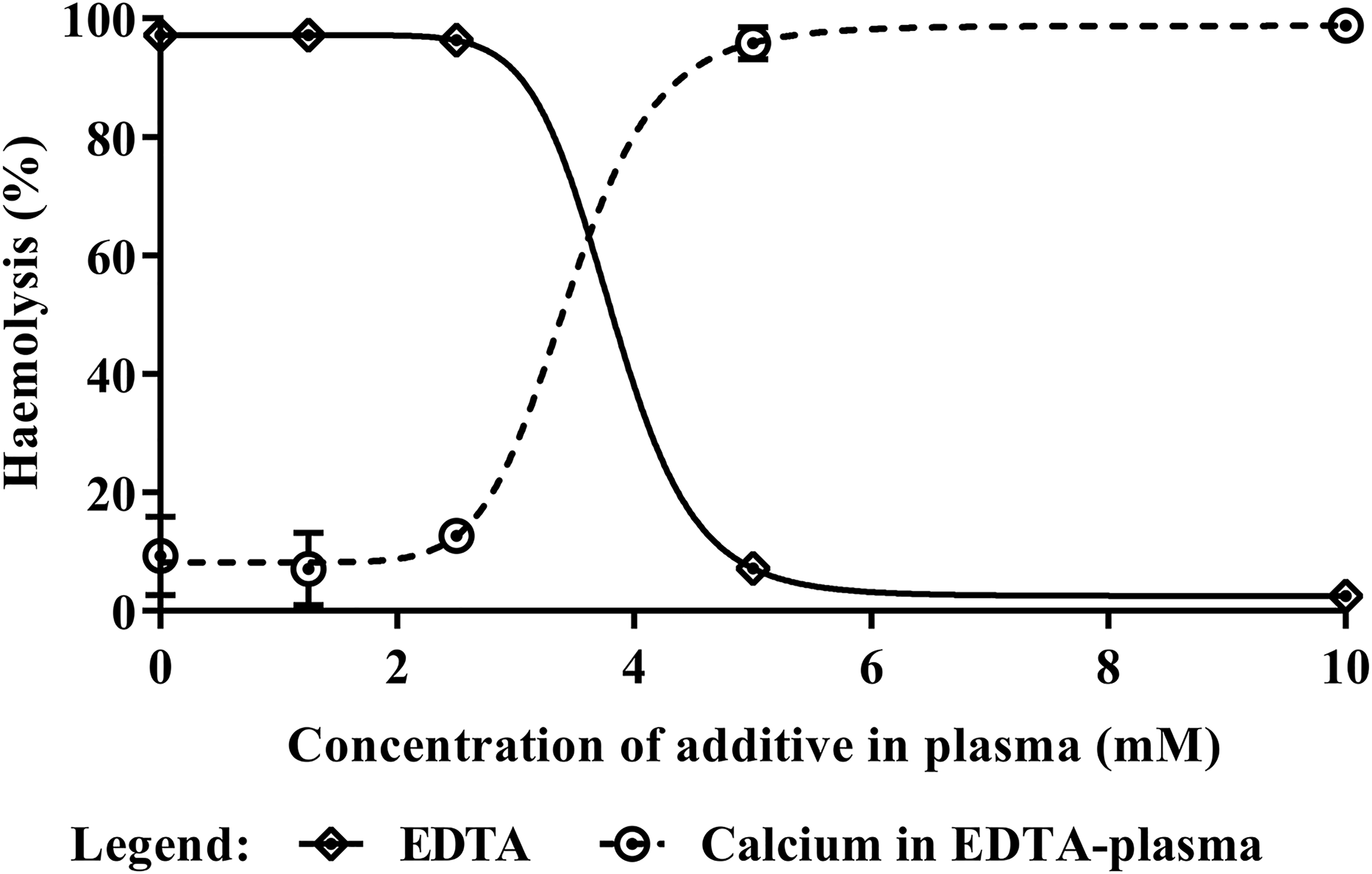
Confirmation that the haemolysis was complement-mediated. The complement-mediated nature of the haemolysis was confirmed by cation chelation and subsequent re-addition of the calcium cation. Each data point represents the mean of three technical replicates with error bars indicating the standard error of the mean (SEM). The chelation of divalent cations through the addition of EDTA resulted in a concentration-dependent decrease in haemolytic activity, with a 50% decrease in the presence of 3.82 mM EDTA. This effect was reversed by supplementing the plasma with calcium, which restored the haemolytic activity of the complement.
Discussion
In this proof-of-concept study, we showed that modified human erythrocytes can be used as an alternative to non-human (i.e. animal-derived) erythrocytes for the evaluation of complement activity. The optimised pre-treatment conditions resulted in modification of the human erythrocytes, which was shown to render them susceptible to human complement-mediated lysis.
It has previously been shown that human erythrocytes subjected to proteolytic treatment become susceptible to complement-mediated haemolysis, via different pathways. Treatment of human erythrocytes with pancreatin alone led to some degree of susceptibility to complement-mediated lysis upon exposure to human plasma. This effect may have been due to the removal of complement regulatory proteins CD55 and CD59, which are located on the surface of erythrocytes, allowing the constitutively active AP to cause haemolysis. This is known to occur readily in undiluted human serum that has been slightly acidified (pH 6.3–6.5), and is also observed in paroxysmal nocturnal haemoglobinuria (PNH) patients. 18,19 In this study, however, experiments were carried out at physiological pH, which could explain the lower levels of haemolysis observed in pancreatin-only treated erythrocytes.
The remarkable sensitivity of TP-Es to complement-mediated lysis is consistent with antibody-independent complement activation by trinitrophenylated erythrocyte surface biomolecules in the setting of proteolytically inactivated complement regulatory proteins (e.g. CD55 and CD59). This synergistic effect is further demonstrated by the use of a TNBS concentration that is lower than the one shown to facilitate antibody-independent activation of the CP in previous studies (2 mM versus 50 mM). 21 Despite the use of a lower concentration of TNBS for treating human erythrocytes, the TP-Es exhibited high levels of haemolysis when exposed to complement (in the form of human plasma). In addition, in terms of protease treatment, a shorter incubation period (4 hours) achieved higher levels of haemolysis than those reported in previous studies following longer incubation. According to the literature, erythrocytes require a 16-hour incubation with trypsin (which is the main component of pancreatin) to achieve 65% haemolysis. 18
The use of EDTA for the chelation of divalent cations, which are necessary for complement activation, reduced haemolysis in a concentration-dependent manner. Subsequent addition of calcium ions restored the haemolytic activity, also in a concentration-dependent manner. The finding that divalent cations are essential for the occurrence of haemolysis indicates that the process is complement-mediated.
While previously published methods permit the distinction of the activities of individual complement pathways, such methods entail the use of various types of animal-derived erythrocytes and different treatment protocols. Employing a single method for the modification of human erythrocytes (i.e. treatment with TNBS and pancreatin) and the evaluation of both total complement and AP activity could potentially reduce cost and time, as both studies could be performed in parallel. Furthermore, the use of the TP-Es in additional assays could also provide information on possible complement factor deficiencies (e.g. C2 or C4 deficiency) or on pathways that might have been inactivated or inhibited by a patient’s treatment.
One major advantage of the method presented in this paper is that it is a potential replacement method for a haemolysis-based complement activity assay (i.e. one that replaces the use of animal-derived products, specifically erythrocytes, with human cells). However, pancreatin, with its wide availability and low cost, was one of the main components used here for the production of TP-Es. The use of this enzyme preparation, which is derived from porcine pancreas, 28 makes the current method — as it stands — a partial replacement method.
The protocol could be refined further, to make it a full replacement method, through the use of alternative proteases from non-animal sources. For example, microorganisms and plants could be explored as sources of proteases to replace pancreatin. Other forms of trypsin itself may be obtained from microbial sources, such as bacteria (including Bacillus licheniformis, Streptomyces griseus and Streptomyces erythraeus), or produced through recombinant protein production. 29 –32 Trypsin-like activity has also been reported in an enzyme found in the envelope extract of Bacteroides forsythus, a bacterial species commonly associated with periodontal disease. 33 Pronase, which is a cocktail of proteases, renders human erythrocytes sensitive to complement-mediated lysis when exposed to acidified serum. 18 Pronase is sourced from the extracellular fluid of Streptomyces griseus. 34 Moreover, treatment with papain, derived from Carica papaya, has been reported to result in the removal of CD55 but only minimal haemolysis, due to the retention of CD59. 18 Several other plant-derived proteases are available, including bromelain from Ananas comosus (pineapple), actinidin from Actinidia spp. (kiwifruit) and ficin from Ficus carica (fig). 35 Treatment of human erythrocytes with these proteases in combination with TNBS could be explored in future studies, in order to completely replace animal-derived materials in the protocol.
Conclusion
This study has demonstrated that human-derived TP-Es can be used as an alternative to animal-derived erythrocytes for the evaluation of total complement and AP activity in human plasma. The replacement of sheep, guinea-pig and rabbit erythrocytes in complement assays will reduce the number of animals potentially used for research purposes. This method could be applied to the screening of complement deficiencies and evaluation of the effects of therapies to regulate the complement system, and thereby alleviate various pathologies where complement dysregulation plays a role (including Covid-19).
In addition to investigating alternative proteolytic treatments, further work will focus on the characterisation of treated erythrocytes to demonstrate the loss of various surface complement regulatory proteins (e.g. CD55 and CD59) through flow cytometry. Moreover, the potential of this method to make use of surplus donated packed red blood cells, thus reducing the need for blood extractions from volunteers, will also be explored in future studies. Cryopreservation of TP-Es is another interesting prospect to be explored, so that the number of production batches can be minimised to ensure a more consistent product quality. The post-cryopreserved TP-Es would also be evaluated for their efficacy in the complement assays.
Footnotes
Acknowledgements
The authors wish to express their appreciation to the Department of Science and Technology-Philippine Council for Health Research and Development (DOST-PCHRD) for providing support through scholarship and dissertation grants to the first three authors, under the MD-PhD (Molecular Medicine) Program of the College of Medicine, University of the Philippines Manila. The authors thank Dr Francisco M. Heralde III, Dr Nina G. Gloriani, Dr Jose B. Nevado Jr and Dr Paul Mark B. Medina, for devoting time and effort to impart valuable feedback and insights. The authors are likewise grateful to the Section of Blood Banking and Transfusion Medicine, University of the Philippines-Philippine General Hospital, for generously providing human plasma samples.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
Ethical approval for the conduct of this study was given by the University of the Philippines Manila Research Ethics Board under study protocol code UPMREB 2016-498-01.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by dissertation research grants from the Department of Science and Technology Philippine Council for Health Research and Development (DOST-PCHRD).