
Abstract
Objective:
This study was aimed to evaluate the effects of diosgenin on myocardial ischaemia-reperfusion injury and the potential involvement of mitochondrial KATP (mitoKATP) channel and nitric oxide (NO) system blockades in this field.
Materials and methods:
After isolation of hearts of male Wister rats, the study was conducted on control and diosgenin- receiving hearts in the presence or absence of 5-HD and L-NAME (as antagonists of mitoKATP channel and NO system, respectively) in an isolated buffer-perfused heart model. Global ischaemia was induced by 30-min occlusion of aortic flow followed by 90-min reperfusion. Cardiac haemodynamics were recorded throughout the experiment using a PowerLab data acquisition system.
Results:
The levels of creatine kinase (CK-MB) and lactate dehydrogenase (LDH) in the coronary effluents were estimated colourimetrically. Diosgenin pre-administration significantly decreased the release of LDH and CK-MD into the coronary effluent as compared the with the control group (P<0.05). The left ventricular developed pressure (LVDP) and contractility (±dP/dt) were significantly improved and restored to pre-ischaemic values in the diosgenin-receiving group (P<0.05). There were no significant differences in left ventricular end-diastolic pressure, coronary flow and heart rate between the control and diosgenin-treated groups during the pre-ischaemic and reperfusion periods. Blocking the mitoKATP channels by 5-HD completely eliminated the positive effect of the diosgenin on the LVDP and ±dP/dt (P<0.05). However, blocking the NO system by L-NAME slightly reduced the diosgenin effects and the inhibitory effect of L-NAME was less than 5-HD.
Conclusion:
The results showed that diosgenin may have cardioprotective effects against myocardial reperfusion injury through activating the mitoKATP channels.
Introduction
Acute myocardial infarction is a leading cause of morbidity and mortality worldwide and threatens human health. 1 This condition results from an interruption of blood flow to the myocardial tissue (ischaemia), which may be followed by either spontaneous or therapeutically induced resumption of the blood supply (reperfusion). This reperfusion, although key to cardiomyocyte survival, is also associated with myocardial damage and may lead to ischaemia-reperfusion (I/R) injury that, consequently, decreases cardiac contractility and its function; thus, finding effective ways to minimize the deleterious effects of reperfusion is very important. 2
Studies of the molecular consequences of I/R injury suggest that opening of the mitochondrial ATP-sensitive potassium (mitoKATP) channels in the mitochondrial inner membrane is considered to be a pivotal step in the mechanisms of pre- and post-conditioning-induced cardioprotection.3,4 Recent research has demonstrated that activation of mitoKATP channels suppresses mitochondrial calcium overload and overproduction of oxygen-derived free radicals during simulated I/R; therefore, a mitoKATP opener may modulate mitochondrial function during I/R. 5 On the other hand, it has been shown nitric oxide (NO) is a mediator/protector of I/R injury. The protective action of NO during I/R is due to its potential as an antioxidant and anti-inflammatory agent. 6 NO acts as an oxygen radical scavenger and also inhibits mitochondrial respiration and, thereby, reduces the generation of oxygen-derived free radicals during I/R injury. 7
Diosgenin (3β-hydroxy-5-spirostene) is a plant-derived sapogenin, isolated from a variety of plants, such as wild yam, fenugreek and soybean. It is structurally similar to estrogen and seems to have some estrogen-like effects in the body (as a phytosterogen). Diosgenin has been used as a dietary supplement in traditional medicine to treat hyperglycemia, hyperlipidemia, inflammation and gastrointestinal problems.8–10 Studies have also suggested a lower incidence for cardiovascular diseases in humans who have a high consumption of diet rich in diosgenin.11,12 Therefore, due to its potential in cardiovascular medicine, the aim of this study was to investigate the cardioprotective mechanisms of diosgenin against I/R injury in rat isolated hearts, with focus on the mitoKATP channels and nitric oxide system.
Materials and Methods
Animals
Healthy adult male Wistar rats weighing 300–350 g (12 weeks old) were used in this study. They were obtained from the animal center of Tabriz University of Medical Sciences and housed under standard animal room conditions (12-h light/dark cycle at 25°C). All animal experiments and procedures were conducted in accordance with the guidelines of the local Animal Ethics Committee and all experimentations were approved by this committee.
Materials
Diosgenin was obtained from Sigma (St. Louis, MO, USA). The selective mitoKATP channel blocker, 5-hydroxydecanoate (5-HD) and a NO synthase blocker, L-Nitro-Arginine Methyl Ester (L-NAME) were purchased from Tocris Bioscience (Anonmouth, UK). All other chemicals and reagents used were obtained from commercial sources in the highest quality available.
Langendorff perfusion setting
All animals were anaesthetized intraperitoneally with sodium pentobarbital (6 mg/100 g) and heparinised with sodium heparin (300 IU). The hearts were excised rapidly via a thoracotomy and immersed in ice-cold Krebs-Henseleit solution (K-H). Then, the hearts were cannulated via the aortas and perfused with K-H solution that contained (in mmol/L): 118 NaCl, 4.8 KCl, 1.2 MgSO4, 1.0 KH2PO4, 27.2 NaHCO3, 10 Glucose and 1.25 CaCl2. A mixture of 95% O2 and 5% CO2 was bubbled through the perfusate at a pH range of 7.35–7.45. A thermostatically controlled water circulator (SatchwellSunvic, Hamilton, UK) maintained the perfusate and bath temperatures at 37°C. The hearts were perfused at a constant mean pressure of 75 mmHg throughout the experiment.
Ischaemia-reperfusion injury protocol
We performed the I/R injury protocol as described previously. 13 The hearts on the Langendorff apparatus were stabilized for about 20 minutes in order to obtain baseline measurements. Then, global ischaemia was conducted by interrupting the aortic flow for 30 min, followed by reperfusion with K-H solution for up to 90 min.
Experimental design
The animals were randomly divided into the following groups (n=10/each group).There was no statistically significant difference in bodyweights among all the experimental groups.
1) Con (control)—in which, after the surgical preparation and stabilization periods, the isolated hearts of animals were perfused with a normal K-H solution for 20 min (to obtain the time-matched controls) and then subjected to a 30-min global ischaemia and 90-min reperfusion with a normal K-H solution.
2) EL-C (Cremophor-EL)—in which the condition was similar to the control group except that the hearts were perfused with a K-H solution containing 0.1% Cremophor-EL (EL-C, as a diosgenin solvent) 20 min before ischaemia.
3) Dio—in which the condition was similar to the control group except that the hearts were perfused with a K-H solution containing 0.001 μM diosgenin for 20 min before ischaemia.
4) 5-HD—in which the condition was similar to the control group except that the hearts were perfused with a K-H solution containing 100 μM 5-hydroxydecanoate (5-HD, as a mitochondrial KATP channel blocker) 20 min before ischaemia.
5) Dio plus 5-HD—in which the condition was similar to the control group except that the hearts were perfused with a K-H solution containing both 100 μM 5-HD and 0.001 μM diosgenin for 20 min before ischaemia.
6) L-NAME—in which the condition was similar to the control group except that the hearts were perfused with a K-H solution containing 100 μM L-Nitro-Arginine Methyl Ester (L-NAME, a nitric oxide synthase blocker) 20 min before ischaemia.
7) Dio plus L-NAME— in which the condition was similar to the control group except that the hearts were perfused with a K-H solution containing both 100 μM L-NAME and 0.001 μM diosgenin for 20 min before ischaemia.
It should be noted that the data acquired in groups receiving EL-C (group 2), 5HD (group 4) and L-NAME (group 6) did not significantly differ from those of the control group in all parameters and the trends of alterations of parameters in these three groups were similar to the controls; therefore, the data of the EL-C, 5HD and L-NAME groups were omitted from the Results section for simplicity in the interpretation of differences between the main four groups.
Measurement of myocardial haemodynamics
During the stabilization period, a latex balloon (Harvard Apparatus Ltd, Edenbridge, UK) attached to the end of a piece of stiff polyethylene tubing was inserted into the left ventricle through the mitral valve. The balloon and tubing were connected to a pressure transducer and filled with normal saline to produce a left ventricular end-diastolic pressure (LVEDP) of 5–10 mmHg at baseline and the balloon volume was maintained constant throughout the experiment. The LVEDP, LV peak systolic pressure (LVSP) and the peak rates of positive and negative changes in LV pressure (±dP/dt) were measured with a Power Lab System (AD Instruments, Newcastle, NSW, Australia). The LV developed pressure (LVDP) was calculated as follows: LVDP=LVSP–LVEDP (in mmHg). The heart rate (HR) was calculated using a bioelectric amplifier (AD Instruments, Australia) from the electrocardiogram that recorded via two electrodes attached to the apex and the right ventricle of the heart and one reference electrode attached to the aortic cannula. All haemodynamic variables were continuously recorded in all groups before ischaemia and during the reperfusion period (10, 60, 90 minutes). The coronary flow was measured by timed collection of coronary effluent during the experiment.
Exclusion criteria
In the Langendorff setting, the hearts were excluded from the experiment if their baseline left ventricular pressures were lower than 70 mmHg or they showed severe arrhythmias and irregular heart rates. The excluded hearts were replaced with the other ones. The exclusion (and replacement) rate for groups was as follows: control = 1 heart; EL-C group = 0; diosgenin group s = 1 heart; 5-HD group = 2 hearts; L-NAME group = 1 heart. Weak contraction or arrhythmias may be related to failure in the surgical and isolation procedures.
Biochemical assays
The coronary effluent was collected during the reperfusion period to measure the heart muscle damage indicators, including myocardial creatine kinase (CK-MB) and lactate dehyrogenase (LDH). Then, the levels of these enzymes were determined using commercial kits (Parsazmoon Co., Karaj, Iran) by auto analyzer (Alcyon 300, Abbott Labs, Santa Clara, CA, USA) in accordance with the manufacturer’s protocol. The absorbance of the solution for LDH was detected at 492 nm and for CK-MB at 340 nm by a spectrophotometer. The results were reported in U/L.
Statistical analysis
All quantitative data are presented as mean±SEM. Data on cardiac function were subjected to a two-way analysis of variance (ANOVA) with the Bonferroni correction test. The CK-MB and LDH levels were analyzed using one-way ANOVA and a subsequent Turkey’s post hoc test to determine the statistical significance between groups. A level of P<0.05 was accepted as statistically significant.
Results
LDH and CK-MB release in control and diosgenin-treated hearts
Pre-administration of diosgenin in I/R hearts decreased both LDH and CK-MB release into the coronary effluent compared with those of the control hearts (P<0.05) (Figure 1). After blocking the mitoKATP channels by using 5-HD, diosgenin failed to significantly affect these enzymes (P<0.05). Inhibiting the NO system through the administration of L-NAME also significantly abolished the effect of diosgenin on LDH, but not the CK-MB level (Figure 1). The data for 3 groups (EL-C, 5-HD, and L-NAME) were similar to those of the control group and, therefore, they have not been shown in the Figures.
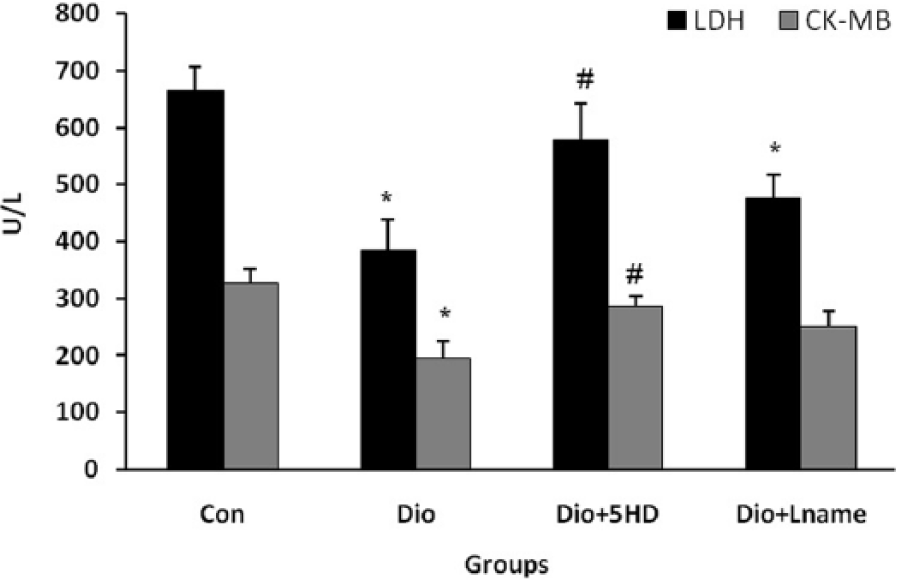
The levels of lactate dehydrogenase (LDH) and creatine kinase (CK-MB) release into the coronary effluent of treated and untreated I/R hearts (n=10). *P<0.05 as compared with the respective control (Con) groups; and # P<0.05 as compared with the respective diosgenin (Dio) groups. 5HD: 5-hydroxydecanoate; Lname: L-nitroarginine methyl ester.
Alteration of HR in control and diosgenin-treated hearts
The baseline values for HR in all groups were similar and this variable reached zero in the ischaemic phase in all groups (Figure 2). However, after reopening of aortic flow to the myocardium, the HR in all the experimental groups recoverd to its baseline values. In addition, there were no statistically significant differences in HR during the reperfusion phase between the treated and untreated groups.
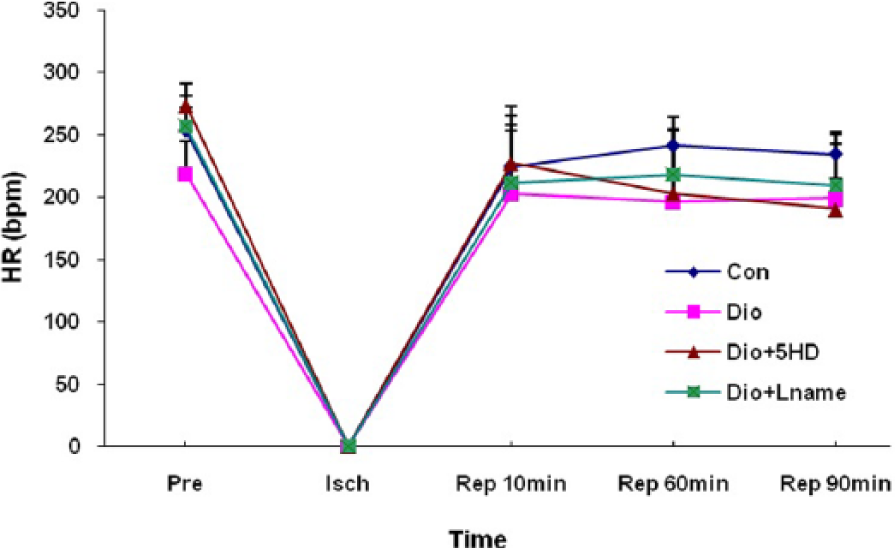
Heart rate (HR) changes in treated and untreated I/R hearts. The data are expressed as mean±SD (n=10). Pre: pre-ischaemic; Isch: ischaemia; Rep: reperfusion; Con: control; Dio: diosgenin; 5HD: 5-hydroxydecanoate; Lname: L-nitroarginine methyl ester.
LVEDP changes in control and diosgenin-treated hearts
In all groups, reperfusion of the ischaemic hearts led to increased LVEDP at all reperfusion times as compared with its pre-ischaemic values (Figure 3). Although diosgenin tended to prevent the elevation of LVEDP in the reperfusion phase, this effect was not statisticaly significant in comparison with the control group. Administration of 5-HD or L-NAME with diosgenin significantly increased the LVEDP in the first phases of reperfusion compared with those of the diosgenin group (P<0.05). The effects of 5-HD or L-NAME were similar with the control group.
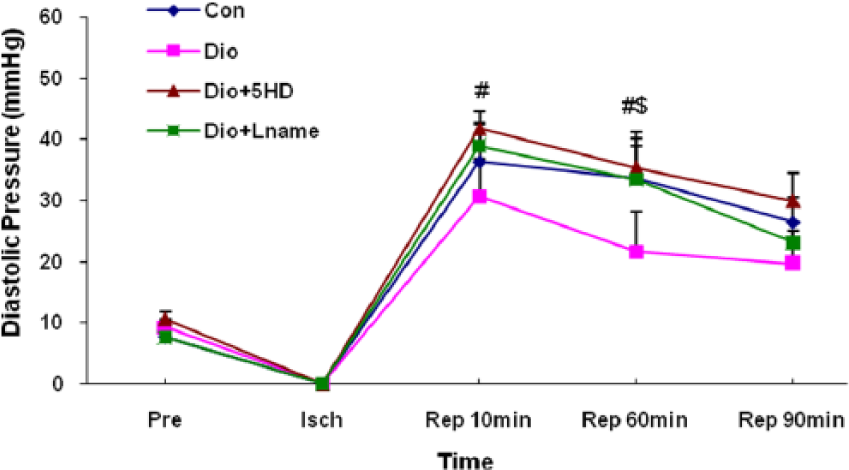
Left ventricular end-diastolic pressure (LVEDP) changes in treated and untreated I/R hearts. The data are expressed as mean±SD (n=10). # P<0.05 Dio+5HD vs. Dio groups; and $ P<0.05 Dio+Lname vs. Dio groups. Pre: pre-ischaemic; Isch: ischaemia; Rep: reperfusion; Con: control; Dio: diosgenin; 5HD: 5-hydroxydecanoate; Lname: L-nitroarginine methyl ester.
LVDP changes in control and diosgenin-treated hearts
Induction of global ischaemia brought the LVDP from its pre-ischaemic values to zero in the experimental groups (Figure 4). After reopening of perfusion supply to the hearts, the LVDP was significantly greater in the diosgenin-receiving group as compared with controls (P<0.05). Administration of 5-HD completely blocked the protective effect of diosgenin at all reperfusion times. Similarly, the administration of L-NAME reduced the effects of diosgenin on LVDP; however, in comparison with 5-HD, the blocking influence of L-NAME on diosgenin action was not strong and considerable (Figure 4).
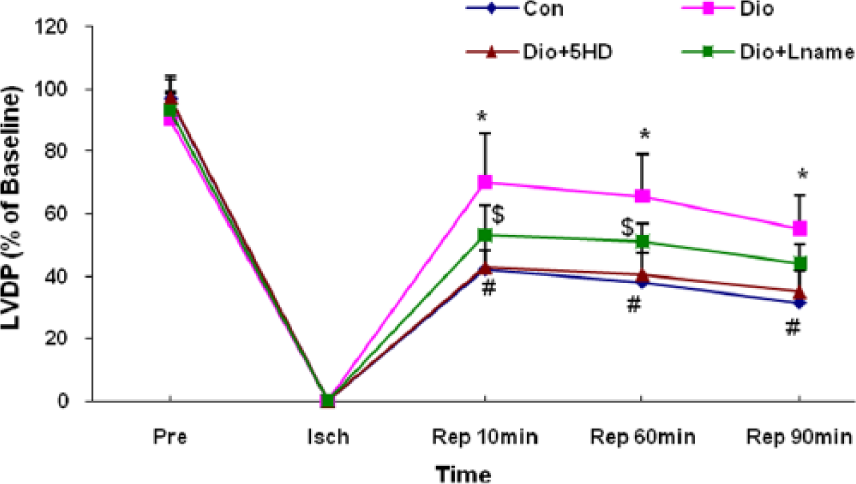
Left ventricular developed pressure (LVDP) changes in treated and untreated I/R hearts. The data are expressed as mean±SD (n=10). *P<0.05 Dio vs. Con groups; # P<0.05 Dio+5HD vs. Dio groups; and $ P<0.05 Dio+Lname vs. Dio groups. Pre: pre-ischaemic; Isch: ischaemia; Rep: reperfusion; Con: control; Dio: diosgenin; 5HD: 5-hydroxydecanoate; Lname: L-nitroarginine methyl ester.
±dP/dt changes in control and diosgenin-treated hearts
Since dP/dt is a function of LVDP and HR alterations, the trend of ±dP/dt alterations approximately mimicked the LVDP trend. As shown in Figure 5, diosgenin significantly recovered the ±dP/dt, especially in the early minutes of the reperfusion phase compared to those of the control untreated hearts (P<0.05). Blocking the mitoKATP channels significantly abolished the effect of diosgenin on +dP/dt in 10 and 60 minutes of reperfusion and on -dP/dt only in 10 minutes of reperfusion. However, the administration of L-NAME had a weaker effect and reduced the diosgenin effect only on +dP/dt in 10 minutes of reperfusion (figure 5).
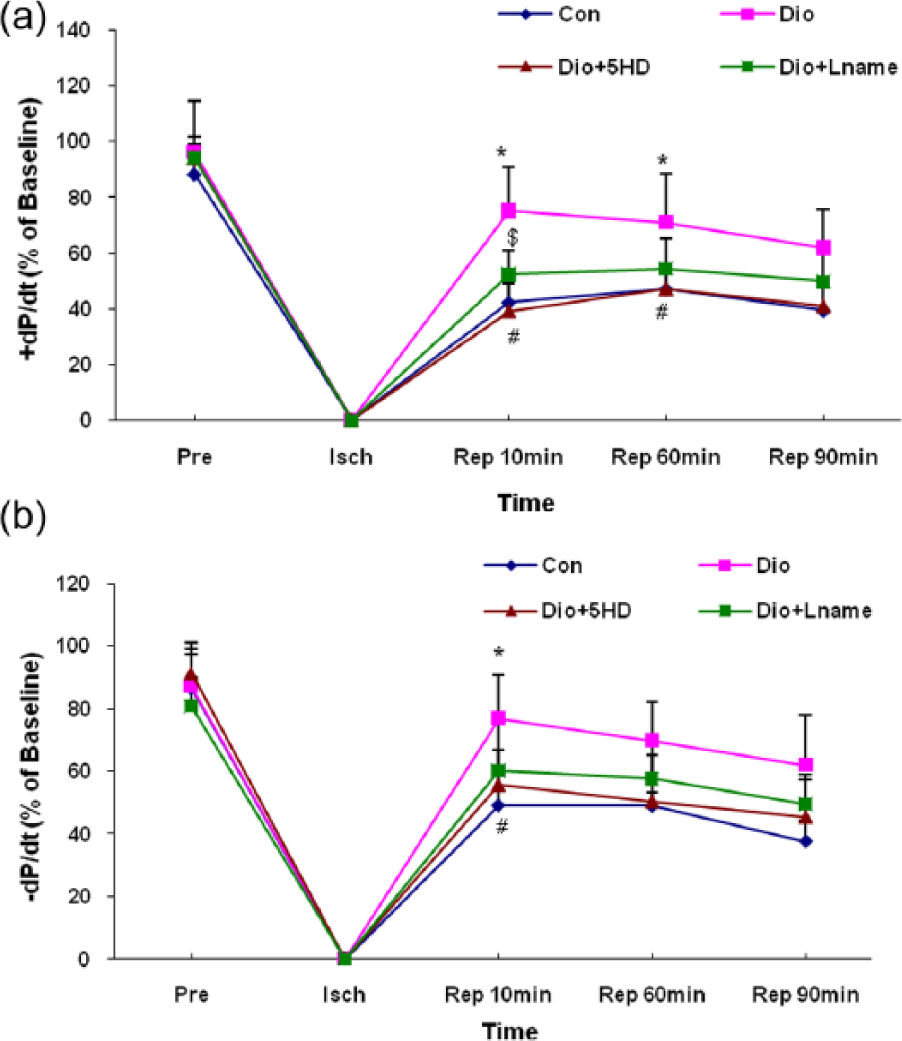
Left ventricular contractility (±dP/dt) changes in treated and untreated I/R hearts. The data are expressed as mean±SD (n=10). *P<0.05 Dio vs. Con groups; # P<0.05 Dio+5HD vs. Dio groups; and $ P<0.05 Dio+Lname vs. Dio groups. Pre: pre-ischaemic; Isch: ischaemia; Rep: reperfusion; Con: control; Dio: diosgenin; 5HD: 5-hydroxydecanoate; Lname: L-nitroarginine methyl ester.
Coronary flow changes in control and diosgenin-treated hearts
The alterations of coronary flow are shown in Figure 6. Although the recovery of coronary flow in the diosgenin-receiving group was greater than other groups, there were no statistically significant differences in coronary flow at baseline, ischaemic phase or all points of the reperfusion phase between treated and untreated hearts.
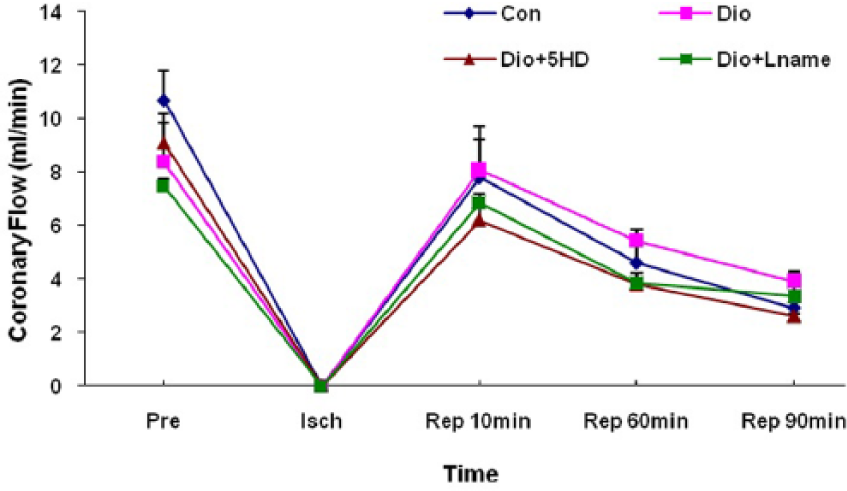
Coronary flow (CF) changes in treated and untreated I/R hearts. The data are expressed as mean±SD (n=10). Pre: pre-ischaemic; Isch: ischemia; Rep: reperfusion; Con: control; Dio: diosgenin; 5HD: 5-hydroxydecanoate; Lname: L-nitroarginine methyl ester.
Discussion
In the present study, we investigated the effects of diosgenin on cardiac function and tissue injury markers in ischaemia-reperfusion injury, when 5-HD and L-NAME were used as the blockers of mitoKATP channels and the NO system, respectively. Briefly, our results showed that LDH and CK-MB were significantly reduced and LVDP and ±dP/dt were significantly improved in the diosgenin group. In addition, blocking the mitoKATP channels completely and blocking the NO system moderately eliminated the positive effects of diosgenin on myocardial contractility and myocardial enzymes in the reperfusion phase. In addition, diosgenin had partial effects on LVEDP, CF and HR.
Clinical and experimental research reported that diosgenin improves peripheral circulation, suppresses platelet aggregation and increases coronary flow and blood supply to the cardiac muscle.11,14–16 In a previous study, we showed that administration of diosgenin before myocardial ischaemia exerted a cardioprotective effect during reperfusion by lowering the levels of inflammatory mediators IL-1β, IL-6, and TNF-α in injured myocardium. 17 In addition, Jayachandran et al. 15 investigated the effect of diosgenin on lysosomal hydrolases, membrane-bound enzymes and electrolytes during isoproterenol-induced myocardial necrosis in rats. They concluded that the protective action of diosgenin might be due to the antioxidant and membrane stabilizing potential of diosgenin. 15 In another study, Son et al. evaluated the possible hypolipidemic and antioxidative effects of diosgenin on rats fed with high-cholesterol diet. They reported that diosgenin could be a very useful compound to control hypercholesterolaemia, which is the main leading cause of cardiovascular diseases, by both improving lipid profile and modulating oxidative stress. 18 Anti-thrombosis activity of diosgenin has been also suggested, which is mediated through improving the anticoagulation function, inhibiting platelet aggregation and thrombosis. 12 In the present study, we showed the cardioprotective effects of diosgenin from other aspects. Our results showed that pre-administration of diosgenin in rat isolated hearts with I/R injury improved cardiac mechanical activities by increasing LVDP and ±dP/dt levels. Additionally, LDL and CK-MD levels were decreased in coronary effluent releasing from isolated hearts in the diosgenin-receiving group; this reflects the reduction in tissue injury, which is a consequence of myocardial infarction. In the mechanistic insight, we found that blocking mitoKATP channels completely reversed the cardioprotective effects of diosgenin.
Mitochondria play an important role in cell death and cardioprotection. During ischaemia, when ATP is progressively deleted, ion pumps cannot function, resulting in a rise in calcium (Ca2+), which further accelerates ATP depletion. 19 The resultant low ATP and altered ion homeostasis and, thus, mitochondrial dysfunction result in rupture of the plasma membrane and cell death.20,21 Therefore, many cardioprotective mechanisms converge on the mitochondria to reduce cell death. One of these mechanisms that reduce I/R injury is the opening of the mitoKATP channels.
Since discovery, many studies have suggested that the mitoKATP channels play a key role in the process of cardioprotection and prevention of apoptosis.22–24 It is not clearly understood how the opening of the mitoKATP channels would lead to cardioprotection. However, there are three hypotheses to explain the link between mitoKATP channel opening and cardioprotection:25–27 (1) a decrease in the mitochondrial Ca2+ uptake, (2) swelling of the mitochondrial matrix and changes in ATP synthesis, and (3) changes in the levels of reactive oxygen species (ROS).
Regarding the first hypothesis, 5-HD could reverse the decrease in the mitochondrial Ca2+ uptake. This change in mitochondrial Ca2+ uptake is thought to be mediated by a partial depolarization of ΔΨm in response to mitoKATP opening. In our previous study, on the other hand, we reported that blocking the mitoKATP channels reversed the anti-inflammatory actions of diosgenin and, thus, the activation of mitoKATP channels may be involved in the cardioprotective effect of diosgenin.
In addition to mitoKATP channels, it appears that the NO system, another mediator involved in I/R injury, also plays a moderate role in the cardioprotective actions of diosgenin. Previous studies indicated that NO plays protective roles in the ischaemic heart by several mechanisms and early studies proposed that NO increased the activity of the mitoKATP channels.28–30 These studies suggest that the effects of NO on mitoKATP channels are mediated via protein kinase (PK) G and PKCϵ. Endothelial NO synthase generates NO, resulting in the activation of guanylyl cyclase and, thereby, PKG which is reported to activate a mitochondrial PKCϵ, ultimately leading to the opening of mitoKATP channels. 31 Exogenous NO reduced mitochondrial calcium overload during simulated ischaemia. 32 However, in the present study, L-NAME, as a blocker of the NO system, did not have a strong effect. Therefore, we hypothesize that there may be other potent regulatory pathways rather than NO controlling the mitoKATP channel opening by diosgenin.
ROS and inflammation are known as important mediators of I/R injury. There is a reverse relationship between ROS generation and mitoKATP channel activation. Therefore, the anti-inflammatory and anti-oxidative properties of diosgenin reported in previous studies10,13,17,18 may explain the cardioprotective influence of diosgenin which is achieved by the activation of mitoKATP channels. In addition, the survival protein kinases involved in the opening of mitoKATP and cardioprotective signalling pathways may be the targets of diosgenin; this hypothesis needs future investigation to be clarified. Furthermore, it has been reported that diosgenin may decrease the vascular tone and increase blood flow. The vascular relaxation induced by diosgenin was significantly inhibited after removal of the endothelium or after pretreatment with L-NAME.11,16 In our study, although the increased coronary flow by diosgenin was not statistically significant, this may indicate the potential of diosgenin to act, at least partly, on the coronary arteries to increase the blood supply to the myocardium. Increased effective blood flow at reperfusion can lead to the effective supply of oxygen and nutrients to the mitochondria and attenuate the severity of I/R injuries.
In conclusion, diosgenin administration before myocardial ischaemia lowered LDH- and CK-MB-releasing as markers of tissue injury and improved cardiac function during the reperfusion period. In this field, mitoKATP channel blockade reversed the positive effects of diosgenin in I/R hearts. Therefore, diosgenin may act as a cardioprotective agent by opening mitoKATP channels and may have the potential to be a useful pharmaceutical component to decrease the outcomes of myocardial reperfusion injury.
Footnotes
Declaration of conflicting interest
The authors have no conflicts of interest in regard to this research or its funding.
Funding
This research has been supported with a financial grant from Tabriz University of Medical Sciences, Tabriz-Iran.