
Abstract
Deep hypothermic cardiopulmonary bypass (CPB) is used in selected procedures such as pulmonary endarterectomy (PEA) and aortic arch repair. Hypothermia induces metabolic and physicochemical changes that affect hydrogen ion concentration ([H+]), haemoglobin–dioxygen (O2) binding, O2 consumption, and carbon dioxide (CO2) production. Modern blood gas analysers provide instantaneous measurements of blood pH, PO2, and PCO2, enabling rapid bedside assessment of acid–base disturbances. Accurate interpretation, however, requires an understanding of the underlying physical and chemical principles. Acid–base management during hypothermic CPB is typically guided by either a pH-stat (interpreting blood gases at the patient’s actual temperature) or an alpha-stat strategy (interpreting blood gases at 37°C, independently of the patient’s actual temperature). The optimal strategy remains debated. This review clarifies key physiological principles of acid–base regulation, the effects of temperature on blood pH and gas solubility, and their clinical implications, with the aim of supporting accurate interpretation of blood gas values during hypothermic CPB.
Keywords
Introduction
As intensive care clinicians and anaesthesiologists, we routinely work with scientific concepts drawn from physics, biochemistry, biology, and mathematics. Ventilator management and the assessment of blood oxygenation rely on fundamental physical principles, particularly gas laws. Likewise, interpreting acid–base disorders requires an understanding of key physicochemical concepts such as electroneutrality, conservation of mass and thermodynamics. 1 Since their introduction in the 1960s in intensive care units (ICUs) and subsequently in operating rooms (ORs), arterial blood gas (ABG) analysers have transformed the assessment of acid–base disorders and gas exchange by providing rapid bedside measurements of pH, PO2, PCO2, and derived variables. 2 This has enabled a more dynamic, physiology-based interpretation of acid–base disorders. However, it has also led to misunderstandings when complex biochemical processes are approached with overly reductionist interpretations. In adults, deep hypothermic cardiopulmonary bypass (CPB) is used for procedures requiring circulatory arrest, such as pulmonary endarterectomy (PEA) and aortic arch repair. 3 Cerebral protection relies largely on metabolism slowing through temperature reduction, sometimes as low as 18°C. 3 Yet deep hypothermia markedly alters acid–base status and gas solubility and affects several metabolic functions.3,4 Two main strategies are used for acid–base/CO2 management during hypothermia: (1) a pH-stat strategy, which interprets ABGs at the patient’s actual temperature; and (2) an alpha-stat strategy, which interprets ABGs at 37°C, independently of the patient’s actual temperature. Which approach yields better clinical outcomes remains contested.5–8 Beyond this debate, our objective is to clarify the physiological concepts required for accurate interpretation of ABG values during hypothermic CPB.
General reminders about fundamental concepts
Basic physicochemical principles
An aqueous solution is neutral when the concentrations of hydrogen ([H+] or [H3O+]) and hydroxyl ions ([OH-]) are equal. 1 Neutral pH depends on system conditions such as temperature and pressure. For example, water has a neutral pH of 7.0 at 25°C, but not at 37°C. 1 Under physiological conditions, the extracellular fluid is alkaline ([OH-] > [H+]), with a pH of about 7.4 at 37°C. 1 Intracellular fluid pH is closer to neutrality, typically ranging between 6.8 and 7.2 at 37°C, although values vary widely across cell types and even between different organelles.9,10
Water dissociates into two ions, H+ and OH-, but its dissociation constant is very low at 37°C; as a result, water behaves almost as a nonelectrolyte solution.
1
Its concentration in the human body is roughly 55 mol/L at 37°C, vastly exceeding the concentration of free H+, and thus serving as an inexhaustible reservoir of H+ (Figure 1).
1
Dissociation and reassociation of water occur rapidly; consequently, H+ does not diffuse freely, nor does it cross cellular membranes via specialized transporters. Instead, it is generated and consumed quasi-instantaneously within biochemical reactions.1,10 Determinants of [H+] and pH in human body fluids. pH is the negative decimal logarithm expression of hydrogen ion concentration. It is a dependent variable determined by three main factors: the strong ion difference (SID), the partial pressure of CO2 (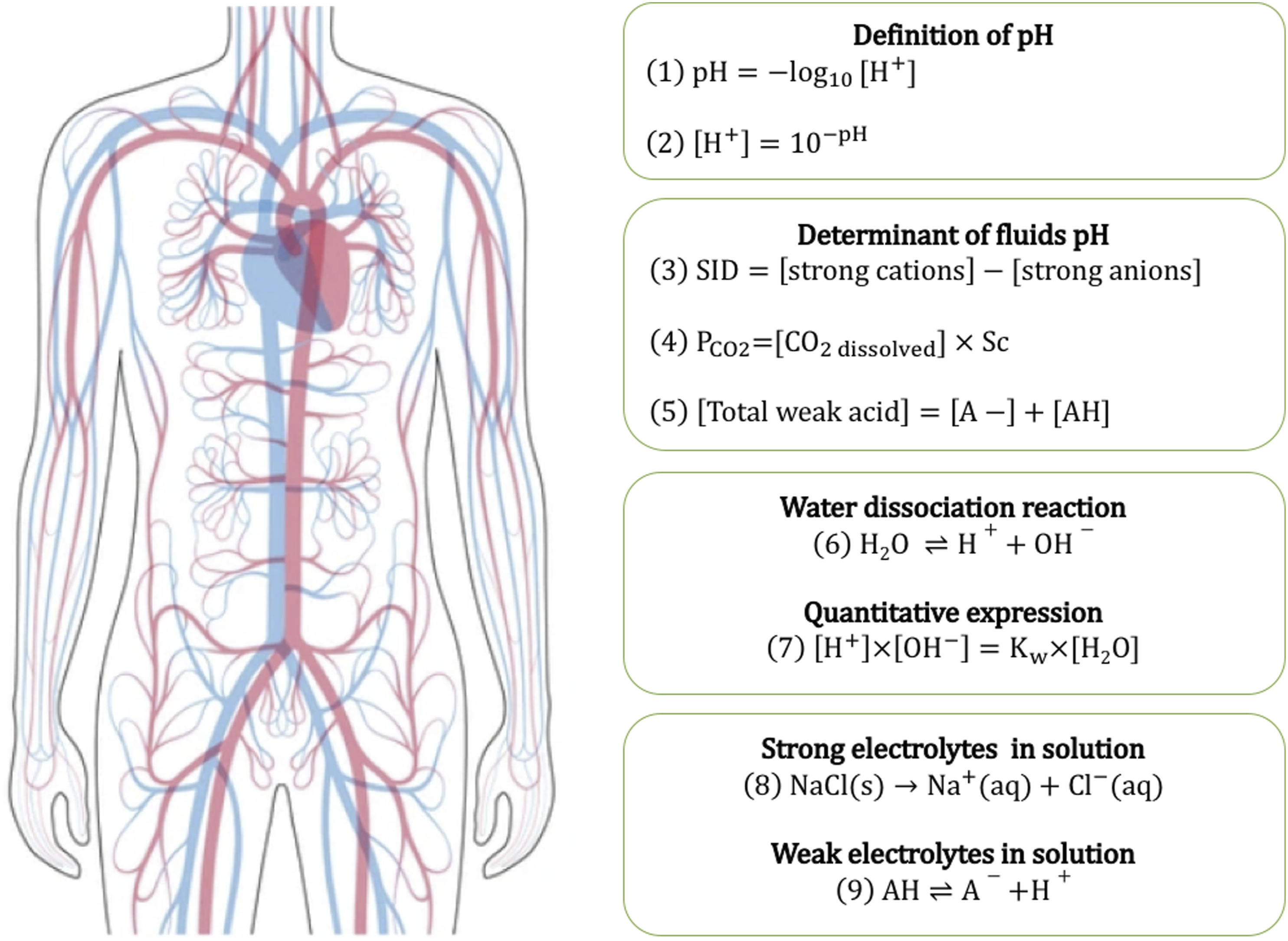
In biological fluids, pH (i.e., [H+]) is determined by three main factors: the strong ion difference (SID; the difference between strong cations and strong anions), the concentration of dissolved CO2 (PCO2 in an open CO2 systems), and the total concentration of weak acids (Atot) (Figure 1). 1 Quantitatively, SID is the most important determinant of pH.1,10
The composition of intracellular and extracellular fluids is quite different. The intracellular space contains higher levels of potassium (K+) and magnesium ions, and relatively lower levels of sodium (Na+) and chloride (Cl−) ions.1,10 The SID is higher in intracellular than in extracellular fluids because of the abundance of proteins. Molecule exchange between compartments is tightly controlled; only water and CO2 diffuse freely across cell membranes. 11
Most metabolic reactions occur intracellularly and are highly dependent on pH, making intracellular pH the variable of interest.9,10 Unfortunately, only plasma pH is available at bedside. Plasma and intracellular pH are related, but not interchangeable.9,10,12
CO2 metabolism
CO2 and water are formed by the reduction of O2; this process is aerobic cellular metabolism. CO2 production is proportional to metabolic rate, and CO2 clearance is regulated through ventilation.1,9 CO2 diffuses across the cellular membrane into the bloodstream, through which it is transported to the lungs for elimination. Once CO2 is dissolved in blood, it undergoes a series of reversible reactions: hydration to carbonic acid (H2CO3), dissociation into H+ and bicarbonate (HCO3-), and further conversion into carbonate (CO32−). Because PCO2 is in equilibrium with H2CO3, it is proportional to H2CO3. [HCO3−] and [CO32−] depend on both PCO2 and SID.1,10 A small fraction of CO2 can also bind to proteins, forming carbamino compounds. 1
Atmospheric PCO2 remains relatively constant. Because the body functions as an open system for CO2, it maintains the concentration of dissolved CO2 in body fluids within a narrow range. Dissolved CO2 in blood equilibrates with alveolar CO2. Alveolar CO2 can rapidly change through adjustments in alveolar ventilation in response to variations in CO2 production—for example, during physical exertion. This dynamic regulation ensures that increases in CO2 only transiently modify pH.1,9
Dissolved CO2 in body fluids, conveniently referred to as PCO2, is an independent variable that determines pH. In a closed system, however, the total CO2 content− including all forms of CO2− becomes the independent variable. When CO2 is added, it will cause a sustained modification in pH.1,9
Blood is a tissue made up of red and white blood cells, platelets, and plasma. Haemoglobin in RBCs makes up most intracellular weak acids. The affinity of CO2 for haemoglobin depends on several factors, including O2, pH, temperature, 2,3 diphosphoglycerate (2,3 DPG) concentration, and diphosphoglycerate mutase activity. 13
What occurs inside RBCs does not directly influence plasma pH because RBCs are impermeable to proteins like haemoglobin. Consequently, haemoglobin does not function as a buffer for plasma pH. Instead, RBCs can interact indirectly with plasma pH by modulating the SID through active exchange of strong ions such as Na+, K+, and Cl− (Hamburger effect).1,14
Temperature considerations
Temperature shifts the equilibrium of chemical reactions according to Le Chatelier’s principle and, in doing so, alters the pH of water-based solutions. From a thermodynamic standpoint, an increase in temperature favours endothermic reactions (reactions that consume heat), whereas a decrease in temperature favours exothermic reactions (reactions that release heat). Consequently, hypothermia slows water dissociation and decreases CO2 production, which tends to raise pH. 1
Temperature also affects the function of macromolecules by altering their structure, three-dimensional conformation, and binding sites. 15 Humans are homeothermic (endothermic) vertebrates, capable of maintaining a stable internal temperature through continuous heat production and tight thermoregulatory control.15,16 However, body temperature varies slightly with circadian rhythm and is not uniformly distributed across body regions. 17 There are also inter-individual variations related to factors such as body surface area, adipose tissue content, and environmental conditions. 18
During hypothermic CPB, the targeted degree of hypothermia may be mild, moderate, deep, or even profound. In the present review, we used the following classification: Mild hypothermia corresponds to temperatures ranging between 28.1–34°C; moderate hypothermia to temperatures ranging between 20.1°–28°C; deep hypothermia for temperatures ranging between 14.1–20°C; and profound hypothermia for temperatures below 14°C. 19
Partial pressure of a gas
According to Dalton’s law, the partial pressure of a gas is defined as the pressure that the gas would exert if it were the only gas occupying the volume considered. According to Henry’s law, the concentration of an ideal gas dissolved in a liquid is proportional to its partial pressure in the gas phase above the liquid, if temperature and pressure remain constant. This is a thermodynamic process: the partial pressure of a gas reflects the kinetic energy (or molecular agitation) of its particles, corresponding to the frequency of their collision with the walls of the gas container. As temperature increases, molecular kinetic energy increases, and the partial pressure of the gas rises accordingly. The amount of gas dissolved in a liquid depends not only on its partial pressure but also on the physicochemical properties of the liquid and on ambient temperature and pressure.
Blood gas analysers measure PO2, PCO2 and pH using a dedicated electrochemical or optical technology. PO2 is assessed either with a Clark polarographic electrode or an optical sensor.20,21 In the Clark electrode, O2 molecules, diffuse from the blood towards a platinum cathode, where they undergo reduction. This reaction generates an electric current which is directly proportional to the concentrations of dissolved O2 and therefore reflects PO2. Optical sensors, in contrast, rely on luminescence-based properties: specific dyes emit photons when excited, and the presence of O2 quenches this fluorescence. 21 The degree of fluorescence quenching enables a direct measure of dissolved O2 concentrations. 21
PCO2 is measured using a pH-sensitive electrode. CO2 diffusing into the measurement chamber undergoes hydration to H2CO3, which dissociates into HCO3- and H+. The resulting change in pH is proportional to the PCO2 of the blood sample.
All measurements are performed at 37°C inside the analyser. A temperature-correction algorithm is then applied to calculate values adjusted to the patient’s actual temperature, yielding the reported ¨temperature-corrected¨ blood gas results.
Finally…
The alpha-stat hypothesis refers to a conceptual framework describing how pH, PCO2, and total CO2 content vary with temperature. First proposed by Reeves in 1972, it states that pH changes with temperature in a manner that preserves a constant ratio between the deprotonated and protonated forms of imidazole groups on histidine residues within proteins.22,23 In other terms, the temperature-induced variation in pH parallels the temperature-dependent change in the negative logarithm of the dissociation constant (pKa) of the histidyl–imidazole group.22,23 Maintaining this constant imidazole dissociation ratio preserves the net charge of proteins, thereby supporting optimal enzymatic activity and stabilizing the three-dimensional (functional) conformation of proteins.22,23
Effect of temperature on blood pH
Temperature dynamically modifies extracellular pH in both ectotherms (cold-blooded) and endotherms (warm-blooded) vertebrates. In their pioneering work, Austin et al. observed in alligators an arterial pH variation (dpH/ΔT) of −0.0018 pH unit per °C. 24 Ectotherms often use the alpha-stat regulation to maintain their physiological state. However, not all ectotherms follow this pattern; for instance, the black racer snake (Coluber constrictor) maintains a constant arterial pH despite temperature changes. 25
In contrast, hibernating mammals adopt a pH-stat regulation during torpor, maintaining a relatively constant pH despite decreased metabolic demands and body temperature. As a result, they develop hypercapnia and acidosis. 26 Interestingly, some hibernating mammals shift to alpha-stat regulation during arousal. 26
In non-hibernating mammals, the situation is more complex. Thermoregulatory mechanisms usually maintain a stable internal temperature, limiting the natural expression of alpha-stat behaviour. However, when these mechanisms are overridden or blunted—such as during anaesthesia—alpha-stat regulation may be predominant.26,27 In alpha-stat organisms, pH increases by approximately 0.0015–0.0018 units for each 1°C decrease in temperature.22,23,28,29 Physiological studies have shown that isolated human blood and dog blood follow alpha-stat behaviour during hypothermia: at a constant total CO2 content, PCO2 decreases while pH increases.30,31
Through its effect on water dissociation, CO2 production and elimination and the SID, temperature dynamically alter pH. The total amount of non-volatile weak acid (Atot), another determinant of pH, can reasonably be considered stable across temperature changes.
A rise in temperature drives the water dissociation forward, increasing [H+] and lowering pH. Conversely, hypothermia reduces molecular kinetic energy, shifts the reaction backwards, lowers [H+], and increases pH (Figure 2). Effect of hypothermia on [H+] and dissolved gases. Hypothermia affects the pH of water-based solutions through several mechanisms. First, it shifts the water dissociation equilibrium towards reduced dissociation, leading to a decrease in [H+] and an increase in pH. Second, hypothermia slows cellular metabolism, thereby reducing CO2 production. In addition, hypothermia decreases molecular kinetic energy, which increases the solubility of gases. In blood, this leads to increased concentrations of dissolved O2 and CO2 despite lower corresponding partial pressures. Hypothermia also shifts the oxyhaemoglobin dissociation curve to the left, reflected by a decrease in P50 (the arterial PO2 at which 50% of haemoglobin is saturated), indicating increased haemoglobin affinity for O2.
Temperature also affects metabolism, and thus CO2 production. 32 It is important to note that the metabolic response to cold varies across species, among mammals, and according to the level of exposure. In unanaesthetised mammals, the initial response to cold exposure is an increase in heat production to counteract heat loss. Consequently, O2 consumption (VO2) rises, and alveolar ventilation and/or O2 extraction increases accordingly.26,27
From this point onward, the term hypothermia refers to a fall in core temperature beyond the body’s thermoregulatory capacity, as occurs in anaesthetised patients undergoing hypothermic CPB.
Hypothermia slows metabolism and CO2 production.32,33 The human body normally behaves as an open system for CO2.1,10 PCO2 is determined by ventilation, whereas total body CO2 content remains relatively constant.1,10 The concentrations of CO2 in its other forms (HCO3−, CO32−, H2CO3) depend primarily on SID, while PCO2 is determined solely by cellular production and ventilation.1,10
Hypothermia alters pulmonary mechanics by reducing lung compliance and depressing O2 and CO2 chemoreceptor function.26,34 In unanaesthetised, non-hibernating mammals experiencing hypothermia, alveolar ventilation −and consequently CO2 elimination − decline in parallel with VO2, and through the direct depressant effect of hypothermia on ventilation.34–36
However, in mechanically ventilated subjects or during CPB, ventilation is controlled by the clinician or the perfusionist; the body is therefore no longer a “fully” open CO2 system. Hypothermia markedly increases the solubility of CO2 and decreases plasma buffering capacity. An identical change in alveolar ventilation produces a larger pH shift in hypothermia than in normothermia.14,37 As a result, minute ventilation or sweep gas flow must be carefully adjusted at each temperature step according to the targeted PCO2 (alpha or pH-stat strategy).
Hypothermia also induces mineral and metabolic changes that can modify SID, and therefore, pH. For example, lactate may accumulate due to shivering, or tissue hypoxia. 14 Additionally, hypothermia enhances renal Na+ excretion and impairs tubular transport, limiting the kidney’s ability to regulate acid-base balance.14,38
Effect of temperature on dissolved gas in blood
To better understand the effect of temperature on O2 and CO2 concentrations in blood, it is helpful to first consider how an ideal gas behaves when dissolved in a liquid. Let us take the example of O2 dissolved in water at atmospheric pressure and constant temperature (T0). Over time, the system reaches equilibrium, where the rate at which gas molecules enter the liquid equals the rate at which they leave. At this point, the relationship between the different variables is described by Henry’s law:
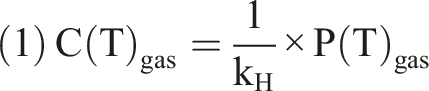
CO2 undergoes hydration and dissociation (forming H2CO3, HCO3−, CO32−) and binds to proteins and haemoglobin. O2 binds extensively to haemoglobin and is continually consumed in mitochondria.
Furthermore, Henry’s law applies only when true thermodynamic equilibrium is reached. In contrast, in living organisms, dissolved gases are continuously added and removed through alveolar ventilation, cardiac output, tissue perfusion, and metabolic demands (VO2 and CO2 production). Consequently, equilibrium is never truly achieved in vivo, and Henry’s law provides only a partial approximation of gas behaviour in blood.
We can now consider the effect of temperature on blood gases such as O2 and CO2. Hypothermia decreases CO2 production and increases CO2 solubility, which lowers arterial PCO2. It also decreases VO2, increases O2 solubility, and shifts the oxyhaemoglobin dissociation curve (ODC) to the left. Clinically, these effects on O2 are highly relevant: during deep hypothermia, because O2 becomes more soluble and therefore more available in its dissolved form, a substantial proportion of metabolic demand can be met by dissolved O2 alone.5,39–42
During hypothermia, acid–base management can be achieved using either the pH-stat or alpha-stat strategy, each leading to very different pH and PCO2 values.8,37 However, a fundamental subtlety is often overlooked when discussing dissolved gases in blood: outside the lungs, blood has no interface with a gas phase. 1 The lungs are the only organ where blood gases reach equilibrium with an actual gas phase—in the alveoli—after diffusing across a thin cellular and extracellular barrier. 1 In healthy subjects, end-tidal PCO2 is approximately equal to arterial PCO2; however, the corresponding concentration of dissolved CO2 depends not only on the solubility coefficient of CO2, but also on the extent of its chemical interactions with plasma, RBCs, water, and proteins. For convenience, we routinely express the amount of dissolved gas in terms of partial pressure. 1 However, this is a conceptual abstraction − an indirect and simplified representation of molecular concentrations in blood. It does not fully reflect the underlying physiology and has no literal gas-phase analogue in the bloodstream. 1 This raises an important question: does this conceptual gap matter in clinical practice?
Partial pressure or concentration: why does it matter?
PO2 and PCO2 are widely used to assess pulmonary gas exchange and blood oxygenation. Blood gas analysers report O2 and CO2 as partial pressures, which do not represent direct measurements of total gas content but rather reflect the gaseous tension of O2 and CO2 dissolved in plasma, as they equilibrate with the alveolar gas. As such, these values are primarily meaningful for evaluating the efficiency of gas exchange between blood and lungs. Nevertheless, PO2 and PCO2 are frequently interpreted as surrogates for gas concentrations in blood, which can lead to oversimplifications and misunderstandings of physiological and metabolic processes. There are two main reasons for this:
Arterial blood gas during deep hypothermia.

Table 1 shows the results of an arterial blood gas analysis performed at 18°C in a patient undergoing pulmonary endarterectomy and highlights the marked differences between temperature-corrected and uncorrected partial pressure values. Blood gas analysers warm the sample to 37°C, resulting in an apparent increase in partial pressure in the gas phase. However, in terms of concentration, the number of O2 molecules per unit volume remains unchanged regardless of temperature. Changes in temperature do not generate additional O2 molecules; they only modify O2 solubility and the position of the oxyhaemoglobin dissociation curve. At the very high values observed in this example, haemoglobin binding sites are fully saturated, even after rewarming. Because O2 solubility increases with hypothermia, the concentration of dissolved O2 is higher at 18°C than at 37°C.
Hypothermia also reduces CO2 production and increases its solubility, while simultaneously altering the equilibria of CO2-related reactions - in water-based solutions. As a result, the concentration of dissolved CO2 cannot be reliably inferred from the partial pressure reported by the temperature-corrected blood gas values.
The behaviour of O2 is somewhat simpler because it interacts predominantly with haemoglobin. When haemoglobin is fully saturated, O2 approximates ideal gas behaviour, and dissolved O2 is approximately proportional to PO2. However, even for O2, this relationship breaks down under altered physiological conditions, such as hypothermia, where dissolved O2 concentration increases while PO2 decreases.
In conclusion, metabolic processes are driven by the molecular concentration of dissolved gases rather than their partial pressures. Partial pressures are useful surrogates for evaluating pulmonary gas exchange, but only under conditions in which Henry’s law applies.
Temperature, solubility, and gas formation
If a gas is dissolved in a liquid under constant pressure and temperature, the liquid can hold only a finite maximal amount of that gas; this state is referred to as saturation. When the amount of dissolved gas exceeds this limit, the system is oversaturated. In oversaturated liquids, dissolved gas may undergo a phase transition and form bubbles as it separates from the liquid phase. This phenomenon is central to the pathophysiology of decompression sickness, which affects divers and individuals exposed to hyperbaric environments. 46 To illustrate the mechanism, consider a diver breathing compressed air. During descent, ambient pressure increases by one bar for every 10 meters of seawater, increasing the partial pressures of inhaled gases—particularly nitrogen (N2). This promotes diffusion of N2 into blood and tissues down its concentration gradient. 46 Tissue N2 content rises with depth and duration of exposure. If ascent is too rapid, ambient pressure falls faster than N2 can be eliminated via the lungs, causing PN2 in blood and tissues to transiently exceed ambient pressure. This oversaturation favours bubble formation and may lead to decompression sickness. 46
Fortunately, at the surface—under normobaric conditions (atmospheric pressure ≈ 101.3 kPa, 1 bar, or 760 mmHg, with body temperature around 37°C)—blood cannot become oversaturated with N2 or O2, even when breathing 100% O2. Two factors explain this. First, arterial PO2 is always lower than alveolar PO2, which itself is necessarily below atmospheric pressure because of water vapour pressure. Second, O2 is continuously consumed by cells, preventing accumulation in blood and tissues, in contrast to N2, which is biologically inert and not metabolised.
During deep hypothermic CPB, the situation differs. Changes in blood temperature can promote endogenous bubble formation, particularly during two critical phases: cooling—when cold blood exiting the oxygenator mixes with warmer aortic blood—and rewarming. 47 As discussed above, hypothermia increases gas solubility in blood. Depending on the inspired O2 fraction (FiO2) and the duration of hypothermia, blood may accumulate high dissolved quantities of N2 and/or O2. During subsequent rewarming, rising temperature reduces gas solubility, allowing dissolved gases to undergo a phase transition and form microbubbles. These gas emboli may contribute to postoperative neurological injury.
Several studies have examined gas emboli during hypothermic CPB to define safer cooling and rewarming strategies. In a pioneering study, Donald et al. reported that gaseous microemboli are released during both cooling and rewarming, and recommended minimising temperature gradients between CPB arterial outflow and venous inflow. 47 Animal studies subsequently confirmed endogenous microemboli formation when temperature gradients between blood and the heat exchanger reached values as high as 10°C.48,49 More recent investigations have suggested that up to 63 mL of gas may be released during the rewarming phase alone. 50 Accordingly, guidelines emphasise maintaining a temperature gradient of less than 10–12°C between the blood and the heat exchanger. In addition, it is recommended to keep the arterial–venous blood temperature gradient below 10°C when blood temperature is below 30°C, and below 4°C when blood temperature exceeds 30°C.47,51
Key points and clinical implications
Interpreting blood gas variables during hypothermic CPB is challenging. The following physiological concepts help structure clinical reasoning and avoid common misconceptions. (1) Patients under deep sedation, mechanical ventilation, and CPB are unable to mount physiological adaptive ventilatory responses to acute changes in temperature and [H+], because they are no longer a fully “open” system for CO2 exchange. Extracellular—and intracellular—pH is therefore significantly influenced by temperature, through both changes in water dissociation and the associated decrease in CO2 production. Correction of pH can be achieved by adjusting ventilation or CPB sweep gas flow, modifying SID, or directly adding CO2 to the CPB circuit. (2) Temperature-corrected PO2 and PCO2 values have limited relevance for understanding gas diffusion between blood and cells. Instead, CO2 and O2 should be considered in terms of molar or volumetric concentrations (i.e., the number of gas molecules or volume per volume of blood). According to physical laws, hypothermia increases the solubility of both O2 and CO2 in blood. Consequently, for a given O2 and CO2 content, the concentrations of dissolved O2 and CO2 increase, resulting in lower PO2 and PCO2 values in the hypothetical gas phase surrounding the blood.52,53
It must be emphasized that temperature-corrected blood gas values are not directly measured but calculated, and that no standardized reference ranges exist for PO2 or PCO2 at temperatures other than 37°C. Furthermore, O2 and CO2 do not strictly obey Henry’s law because of their chemical interactions with water and blood components. As a result, estimating gas concentrations from partial pressure is unreliable, except for O2 when haemoglobin is fully saturated. (3) Markers such as oxygen delivery (DO2), veno-arterial O2 difference, and VO2 are useful for assessing tissue oxygenation during CPB. Deep hypothermia markedly increases haemoglobin affinity for O2, limiting O2 release to tissues. Several studies suggest that, during deep hypothermic CPB, tissue oxygenation may rely predominantly on dissolved O2 rather than haemoglobin-bound O2.5,39–42
Accordingly, increasing FiO2 during cooling may increase dissolved O2 availability and help reduce the risk of tissue hypoxia, particularly during periods of circulatory arrest. This approach may also limit endogenous microemboli formation by reducing tissue N2 accumulation. 42
Caution is warranted when interpreting venous O2 saturation (SvO2) during hypothermic CPB. High SvO2 values are expected because of increased O2 solubility and the leftward shift of the ODC. Thus, SvO2 does not reliably reflect the balance between O2 supply and metabolic demand and should not be used as a surrogate marker of adequate tissue oxygenation in this context.41,42
Classic biological markers such as serum lactate remain useful indicators of impaired O2 utilisation; however, they must be interpreted cautiously. Hypoxia—defined as reduced mitochondrial O2 availability—may occur in the absence of hyperlactatemia.54,55 Conversely, hyperlactatemia may reflect mechanisms unrelated to tissue hypoxia, such as reduced hepatic lactate clearance or increased production due to shivering.14,56
Urine output, although often used to assess the adequacy of blood flow relative to metabolic demand, is a poor marker of metabolic adequacy during deep hypothermic CPB. CPB can impair renal function through multiple mechanisms, including hypoperfusion, non-pulsatile flow, systemic inflammation, haemolysis, and inadequate venous drainage. The effects of moderate and deep hypothermia on tubular function remain poorly understood.14,57 Glomerular filtration rate is tightly coupled to systemic blood flow and typically decreases during cooling. 14
However, polyuria with dilute urine is frequently observed during cooling. The mechanism is unclear but may involve sympathetic nervous system activation and hypothermia-induced tubular dysfunction.14,58 Therefore, current evidence is insufficient to support urine output as a reliable indicator of renal function or metabolic demand during deep hypothermic CPB. (4) Most proteins are intracellular, where intracellular pH is tightly regulated.9,59 CO2 diffuses readily across cellular membranes.
1
Consequently, during pH-stat management, changes in extracellular CO2 will also influence intracellular pH. The addition of CO2 to the blood should therefore be considered carefully, as the resulting increase in dissolved CO2 concentration may be difficult to predict. CO2 also crosses the blood–brain barrier and can decrease cerebral intracellular pH, leading to cerebral vasodilatation. Some authors argue that this may be beneficial during hypothermic CPB by improving cerebral blood flow and cooling, whereas others suggest it could increase the risk of cerebral oedema or exacerbate reperfusion injury.37,60 (5) The controversy regarding the optimal pH management strategy remains unresolved. Both strategies have potential benefits and drawbacks, and clinical studies have yielded inconsistent results (Figure 3).5,60–63 Advantages and limitations of alpha-stat and pH-stat strategies. Figure 3 depicts the potential advantages and risks associated with the pH-stat and alpha-stat strategies with respect to cerebral blood flow and metabolism (Image(s) provided by Servier Medical Art (https://smart.servier.com), licensed under CC BY 4.0 (https://creativecommons.org/licenses/by/4.0/).
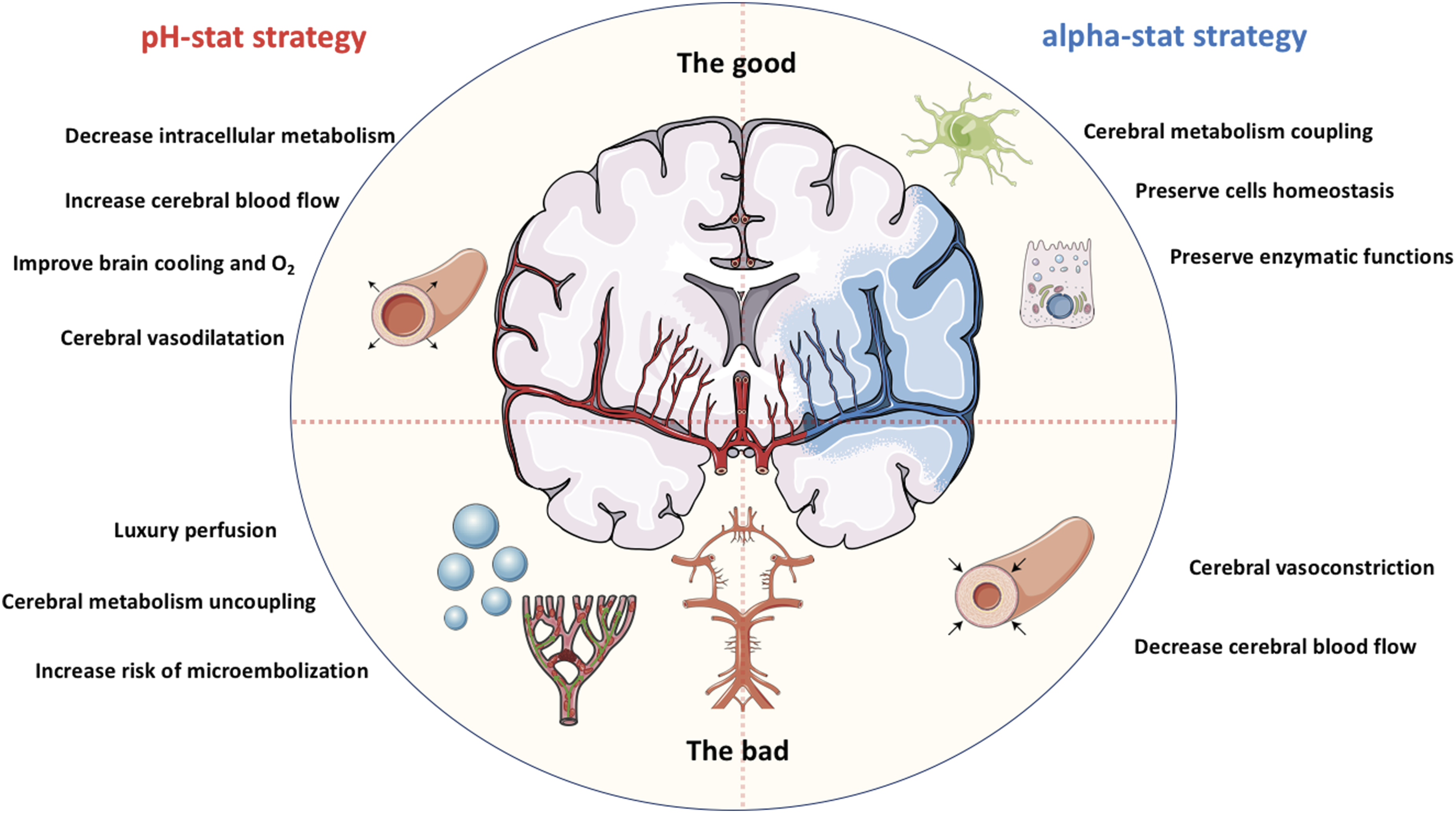
Potential benefits of pH-stat management include several mechanisms. First, it leverages the vasodilatory effect of CO2 on the cerebral vasculature, which increases cerebral blood flow and may enhance brain cooling.9,31 Second, intracellular acidosis can inhibit cellular metabolism, thereby reducing VO2. Third, the extracellular decrease in pH promotes O2 release from haemoglobin (Bohr effect).60,64,65 Fourth, at the cellular level, pH-stat management decreases intracellular pH and may activate protective pathways, including improved mitochondrial stability and enhanced antioxidant defences.31,66,67 However, the metabolic inhibition associated with pH-stat management may become clinically relevant only after a certain duration, which can exceed the length of many surgical procedures.9,32,60 Moreover, pH-stat management has been associated with cerebral hyperperfusion (“luxury perfusion”), which may increase the risk of cerebral microembolisation. 60
Experimental and clinical studies have yielded divergent results depending on patient population and depth of hypothermia. Animal studies have shown that pH-stat management increases cerebral blood flow, improves brain cooling, and improves oxygenation—through the Bohr effect—and has been associated with better preservation of cerebral metabolism.7,68–70 These findings have also been reported in infants undergoing deep hypothermic circulatory arrest. 5 In contrast, in adults undergoing moderate hypothermic CPB (approximately 26°C), alpha-stat management has been associated with superior neurological outcomes compared with pH-stat management.60–62
From an evolutionary and physiological perspective, alpha-stat regulation may better preserve cerebral metabolic coupling, cellular homeostasis, and enzymatic structure and function in endothermic vertebrates.9,22,32 Importantly, there is no obligation to apply a single acid–base management strategy throughout the entire procedure. Several authors have proposed a hybrid approach tailored to surgical phases and guided by cerebral monitoring tools such as near-infrared spectroscopy (NIRS) or transcranial Doppler.9,53,71 For example, some suggest using a pH-stat strategy during cooling and up to circulatory arrest, then transitioning to alpha-stat management before circulatory arrest or during rewarming.53,71 (6) During deep hypothermic CPB, numerous factors other than temperature can influence acid–base status. These include the nature and volume of administered fluids, the degree of haemodilution, the adequacy of pump-delivered blood flow relative to metabolic demand, changes in blood viscosity and microcirculatory perfusion, and renal dysfunction secondary to systemic inflammation, haemolysis, or inadequate perfusion. Each of these variables can independently or synergistically alter metabolic and acid–base balance and should be considered when interpreting blood gas and metabolic variables during hypothermic CPB.
Conclusion
Acid–base and oxygenation management during hypothermic CPB are complex and clinically high-stakes. Hypothermia profoundly alters water dissociation, CO2 production, and gas solubility, which disrupts the usual relationship between partial pressure and dissolved gas concentration. Clinically, this means that temperature-corrected blood gas values should not be interpreted in isolation; management should be guided by physiological targets (e.g., pH strategy, CO2 handling, oxygen content and delivery) and by the surgical phase. Practically, clinicians must anticipate stepwise changes in pH and PCO2 with cooling and rewarming and actively adjust ventilation or CPB sweep gas flow, SID, and—when appropriate—FiO2 to maintain adequate O2 delivery, which during deep hypothermia may depend largely on dissolved O2. In parallel, limiting temperature gradients during cooling and rewarming is essential to mitigate endogenous microbubble formation and its neurological consequences. The optimal acid–base strategy remains controversial, with no definitive consensus on whether alpha-stat or pH-stat management is superior; a phase-adapted, individualised hybrid approach guided by cerebral and metabolic monitoring (e.g., NIRS, transcranial Doppler, DO2/VO2, lactate trends) is likely to be the most clinically pragmatic and physiologically coherent option.
Take home message
In biological solutions, hypothermia slows water dissociation and reduces CO2 production, resulting in an increase in pH. - Hypothermia increases gas solubility. For a given O2 and CO2 content, the concentrations of dissolved O2 and CO2 increase, while PO2 and PCO2 decrease. - Temperature-corrected PO2 and PCO2 values have limited relevance for understanding gas diffusion between blood and cells. CO2 and O2 must be considered in terms of their respective concentrations. - The pH-stat strategy lowers intracellular pH because CO2 freely crosses cell membranes. Its effects on cerebral perfusion are difficult to predict and therefore require close monitoring. - The choice between alpha-stat or pH-stat strategies remains controversial, and there is no obligation to apply a single strategy throughout the procedure. A hybrid approach, tailored to surgical phases and guided by cerebral monitoring, may be the most appropriate.
Footnotes
Acknowledgement
We want to thank all the members of our perfusion team, for their work and day to day commitment.
Author contributions
SD and RM wrote the first draft of the manuscript. JT and MCK reviewed it critically and made substantial corrections. All authors approved the final version of the manuscript.
Funding
Financial/Non-financial disclosure: Support was provided solely from institutional and/or departmental sources.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.