
Abstract
Objective
Disordered programmed cell death may play a role in the development of superficial venous incompetence. We have determined the number of cells undergoing apoptosis and the alterations in the apoptotic level in the wall of different segments of the great saphenous varicose vein.
Methods
Twenty-one varicose great saphenous veins (VGSVs) (varicose group) and 12 normal great saphenous veins (GSVs) (control group) were collected, and the apoptosis level in the upper, middle, and lower segments were immunohistochemically stained with antibodies (anti-Bax and anti-Bcl-xl). Apoptosis was evaluated by the TUNEL assay and immunofluorescence staining. The morphology of apoptotic cells was observed with an electron microscope.
Results
Quantitative analysis showed that the apoptotic ratios in venous walls (intima and media) of the varicose group were significantly lower than the corresponding regions in the control group (all P < 0.05). A significantly higher apoptotic rates of the venous walls was observed in control group within the upper compared with the lower segment (P < 0.05). Significantly higher positive proteins expression rates of Bcl-xl/Bax were also detected in the VGSVs compared with the GSVs within the three segments, respectively (P < 0.01). Electron microscopic observations confirmed that endothelial and smooth muscle cells in varicose and normal vein walls exhibited apoptotic morphologic features, such as fuzzy mitochondrial cristae, medullary changes, and margination of the nuclear chromatin.
Conclusion
VGSV walls were found to have a significant decrease in apoptotic rate compared with that of GSVs. The rate of apoptosis in the intima and media within the upper segment was increased more than the middle and lower segments in the GSVs. Our findings confirm that programmed cell death is down-regulated in primary varicose veins.
Introduction
Lower extremity varicose veins are one of the most common medical conditions affecting people worldwide. Despite extensive studies, the etiology of varicose veins is unclear, as are the molecular mechanisms that could potentially lead to dilation of the vein wall. Studies in human renal, coronary, and carotid arteries have shown increased apoptosis and an increased number of intrinsic pathway promotors, including the apoptotic promoter, Bax. 1 Controversial data exist with respect to the role of apoptosis in varicose veins. Several authors have reported a reduction in the apoptosis rate in varicose veins compared with non-varicose veins.2–4 In contrast, Bujan et al. reported an increase in apoptosis of smooth muscle cells (SMCs) in varicose veins. 5 Urbanek et al. described an increase in the SMC apoptotic index in the media of distal varicose veins of patients <50 years of age, but not in an older age group. 6 Simovart et al. observed an increasing trend of the number of apoptotic SMCs in the walls of varicose veins with the advancing age. 7 In contrast, much less attention has been paid to the apoptotic regulation of endothelial cells (ECs) in varicose veins. In the present study, we used the terminal deoxynucleotidyl transferase-mediated nick-end labeling (TUNEL), immunofluorescence and the immunohistochemical expression of pro-apoptotic Bax and anti-apoptotic Bcl-xl markers, as well as electron microscope assays, to quantitatively analyze alterations in the apoptotic level in the wall of different segments of varicose great saphenous veins (VGSVs) to investigate the relationship between apoptosis of ECs and SMCs and varicose venous walls.
Materials and methods
Ethical approval of the study protocol was obtained from the Human Research Ethics Committee of Weifang Medical University (Weifang, China). All individuals provided written informed consent to be involved in the study.
Materials
Ninety-nine tissue samples were collected from patients in the 89th Hospital of the People's Liberation Army (Weifang, China) from January 2010 to May 2013. Twenty-one cases (12 males; mean age: 58 years; age range: 55–68 years) with VGSVs who had received high ligation and stripping plus rotary cutting were included in the varicose group. All 21 cases were confirmed to have primary VGSVs using color Doppler ultrasound before surgery. Patients were classified as C2–C3 according to the clinical, etiologic, anatomic, and pathophysiologic (CEAP) classification, with a disease duration of 4–7 years. A total of 12 normal great saphenous veins (GSVs) were obtained from 12 patients (seven males; mean age: 59 years; age range: 56–69 years) who were undergoing distal arterial bypass grafting surgery. The exclusion criteria were as follows: severe lower limb edema, varicose veins with skin lesions, thrombophlebitis, phlebitis without thrombus, arterial disease, and diabetes.
Specimen collection and processing
Paraffin sections were made. A 3–4 mm sample was obtained from the upper (the thigh root), middle (below the knee joint), and lower (above the malleolus medialis) segments of great saphenous veins. Specimens were fixed in 10% formalin, dehydrated, embedded, and serially sliced into 4 µm sections. One slice of each sample was used for H&E staining (H&Ekit; Labest Biotech Co. Ltd, Beijing, China). Each specimen was cut into 15 slices, with five slices per group. Immunohistochemistry and immunofluorescence for these sections were performed.
A total of 1 mm3 of fresh specimens from all the groups were obtained. The specimens were fixed in 3% glutaraldehyde (Lebang Biotech Co. Ltd, Suzhou, China) for 24–48 h, dehydrated, embedded, and dried. The specimens were then made into ultrathin (70 nm) slices (UC7 ultrathin slicing machine; Leica). Slices were cleansed with water and soaked in a saturated aqueous solution of uranyl acetate (Zhenpu Biotech Co. Ltd, Shanghai, China). The slices were then cleaned with doubledistilled water and soaked in lead citrate solution (Tianyuan Co. Ltd, Yingkou, China). The ultrastructural changes of venous structure cells were observed using a Hitachi H-7500 transmission electron microscope (Hitachi, Tokyo, Japan).
Immunohistochemistry
The immunohistochemical techniques which were used have been described elsewhere.4,8 For the histologic analysis, five fields per section were examined for staining at 400 × magnification. The immunohistochemical expression of apoptotic cells was determined according to the intensity of nuclear staining and the density and morphology of chromatin. The results were evaluated by two independent investigators who were blinded to the patients’ clinical findings. Based on TUNEL labeling and immunofluorescence in situ, Bax and Bcl-xl immunoreactive cells were considered positive for apoptosis in the intima and media layers were counted manually at 400 × magnification and five random fields per section were counted. Two to five sections per specimen were evaluated. Protease K, apop A and apop B were provided by Roche Biotechnology Company. The immunohistochemical stain for pro-apoptotic Bax and anti-apoptotic Bcl-xl markers was purchased from Zhongshan Golden Bridge Biotechnology.
Statistical analysis
SPSS (version 17.0; SPSS, Inc., Chicago, IL, USA) was used. Data are expressed as the mean ± standard deviation. Differences in measurement data were compared using Student’s t-test with two independent samples and two paired samples. A P < 0.05 was considered significant.
Results
Distribution of TUNEL and immunofluorescence positive cells
The results obtained with the TUNEL assay were the same as the results obtained with the immunofluorescence assay. In the VGSV group, only scattered apoptotic cells were present in the venous walls (intima and media), which were almost uniformly distributed within the three segments (Figure 1 (a)–(f)). While there were considerably more apoptotic cells in the venous walls (intima and media) in the GSV group, apoptosis was more evident in the upper segments than the lower segments (Figure 1(g) to (l)).
The positive expression of apoptotic cells in venous walls within three segments.
Determination of TUNEL and immunofluorescence positive cells rates
Comparison of apoptotic ratio in venous walls (intima and media) within the three segments (%,
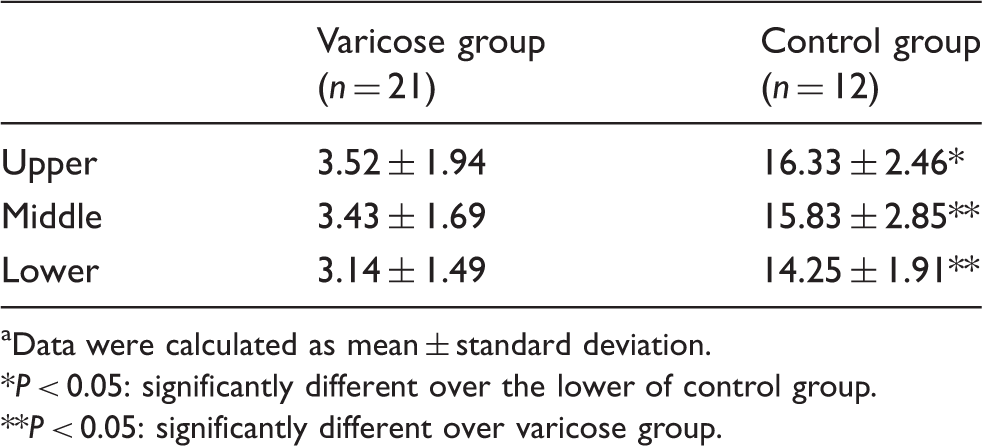
Data were calculated as mean ± standard deviation.
P < 0.05: significantly different over the lower of control group.
P < 0.05: significantly different over varicose group.
Distribution of the apoptotic cell proteins expression
In the VGSVs group, only scattered cells with positive expression of Bax were present, while there were considerably more cells with positive expression of Bcl-xl in the venous walls (intima and media) within the three segments (Figure 2(a) to (f)). In the GSV group, the results obtained with positive expression of Bax and Bcl-xl were opposite that of the VGSV group within the three segments. Cells with positive expression of Bax and Bcl-xl were more evident in the upper segments than the lower segments (Figure 2(g) to (l)).
The positive expression of apoptotic proteins in venous walls within three segments.
Determination of the apoptotic cell proteins expression rates
Comparison of apoptotic cell proteins expression ratios in venous walls (intima and media) within the three segments (%,
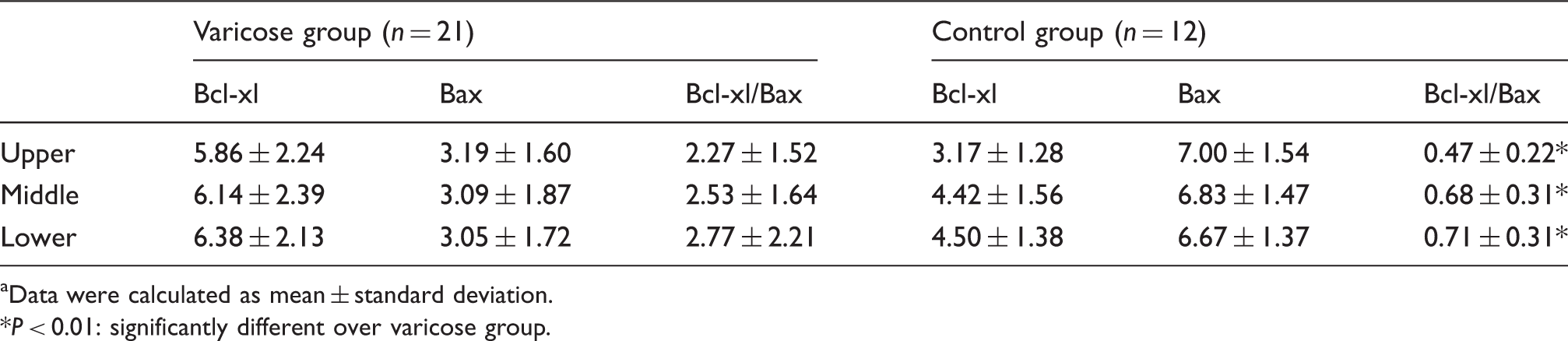
Data were calculated as mean ± standard deviation.
P < 0.01: significantly different over varicose group.
Ultrastructural changes of apoptotic cells
In the VGSV and GSV groups, the apoptotic morphologic features (including fuzzy mitochondrial cristae, crista breaking, medullary changes, rough endoplasmic reticulum thickening, degranulation, and margination of the nuclear chromatin) were present in SMCs and ECs of venous walls within the three segments (Figure 3(a) to (c), (e) to (g)); normal morphologic features, including normal mitochondria, rough endoplasmic reticulum, and nuclear chromatin were also noted (Figure 3(d) and (h)).
Ultrastructural changes of smooth muscle cells (SMCs) and endothelial cells (ECs) in venous walls within three segments.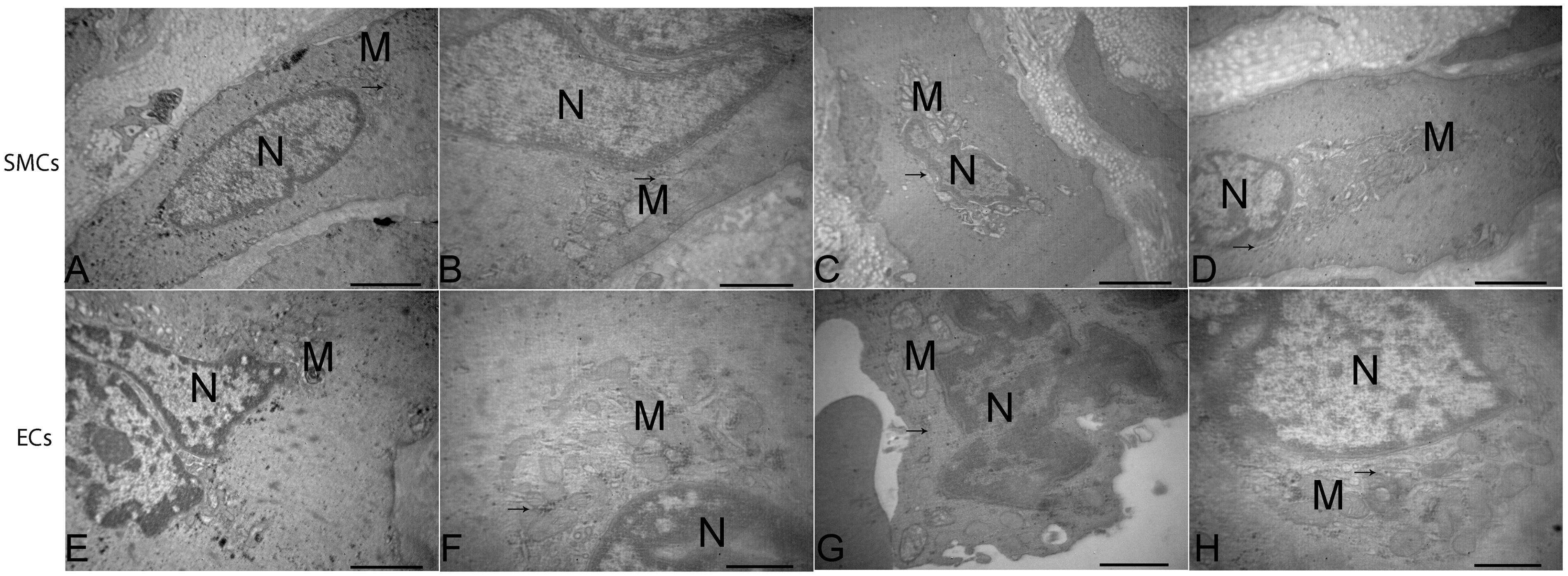
Discussion
Kerr et al. first proposed the concept of apoptosis, which is physiologic programmed cell death. 9 Apoptosis is a very complex, tight cascade reaction process, which plays a role in the development and senility of tissue and organs, in maintaining the stability of tissue homeostasis and reducing cell renewal. Apoptosis includes endogenous and exogenous channels. Endogenous channels are also referred to as mitochondrial channels, and can regulate apoptosis priming proteins (Bax or Bcl-2) and specific caspases (caspase-9) to stimulate mitochondrial release of cytochrome C, which then combines with apoptotic protease-activating factor-1 (APAF-1) within the cytoplasm. Exogenous channels are also referred to as transmembrane channels. Under the stimulation of apoptosis signals, FasL and tumor necrosis factor-alpha (TNF-α) can lead to apoptosis through binding with their corresponding receptors.10–12 In vascular surgery, apoptosis was first observed in peripheral atherosclerotic occlusive disease, and apoptosis participates in the stages of atherosclerotic plaques.13,14 The apoptosis index is increased in atherosclerotic walls of renal, coronary, and common carotid arteries. 1 Green et al. suggested that disordered apoptosis is mainly mediated by endogenous channels in varicose venous walls. 15 Apoptosis is down-regulated in the media layer of varicose veins. Dysregulation of the cellular mechanism that maintains normal tissue integrity is mediated through the intrinsic apoptotic pathway. 4 Other scholars believed that this phenomenon may be related to the turbulent structure of walls and abnormal distribution of collagen and elastic fibers.9,16 It is evident that apoptosis participates in remodeling of varicose venous walls.
It has been verified that abnormal apoptosis exists in venous walls, but the results differ, and little is known about apoptosis and the regulatory mechanism governing apoptosis. Ascher et al. conducted a morphologic analysis of venous walls and found that apoptotic cells in varicose venous walls were >3% in only 15% of patients with varicosities, but 48% in the control group. 2 By detecting apoptosis-related factors, the expression of Bax in the intima and media of varicose venous walls has been shown to be decreased and the expression of poly-ADP ribose polymerase (PARP) inhibited; Bax and PARP down-regulate apoptosis through endogenous channels. 3 It has been shown that apoptosis is the same in varicose spermatic venous walls and VGSV walls; specifically, the walls are thickened, the lumen is expanded, the apoptosis index is decreased, and the expression of HIF-1α and Bcl-2 increased. 17 Knaapen et al. and Urbanek et al. showed that the level of apoptosis is decreased in the walls of proximal GSVs, and ER-β, p21, p53, and Fas mRNA is increased.18,6 Ducasse et al. studied the trunk and branches of GSVs and showed that the medium of the mitochondrial channel decreased, the apoptosis channel was blocked, and apoptosis was down-regulated in the media; however, some scholars have contested the relationship between apoptosis and varicose veins. 19 Bujan et al. found that apoptosis of SMCs in varicose venous walls was higher compared with normal veins, also noting collagen accumulation and reduced wall elasticity. 5 Electron microscopic observations confirmed that SMCs in varicose venous walls showed typical apoptotic morphologic features, such as chromatin condensation, cytoplasmic hypervacuolization, and apoptotic bodies. 20 Filis et al. found that the expression of Bax, caspase-3, Bcl-xl, Bcl-xs, and Ki-67 in VGSVs walls increased, apoptosis increased, and apoptosis was more obvious in the distal trunk than in the proximal trunk. 21 It has been reported that the counts of apoptotic ECs, SMCs, and ages had a positive correlation.7,22
Our research showed that in the varicose group, only scattered apoptotic cells were found in the venous walls (intima and media), which were almost uniformly distributed at the three segments, while there were considerably more apoptotic cells in the venous walls (intima and media) in the control group by TUNEL assay and immunofluorescence staining. In the varicose group, only scattered cells with positive expression of Bax were present, while there were considerably more cells with positive expression of Bcl-xl in the venous walls (intima and media) within the three segments; control group results were opposite that of the varicose group within the three segments. Cells with positive expression of Bax and Bcl-xl were also more evident in the upper segments than the lower segments. Quantitative analysis showed that the apoptotic ratios in venous walls (intima and media) of the varicose group were significantly lower than the corresponding regions in the control group. A significantly higher apoptotic rate of the venous walls was observed in control group within the upper compared with the lower segment. Significantly higher proteins expression rates of Bcl-xl/Bax were also detected in VGSVs compared with GSVs within the three segments, respectively. Electron microscopic observations confirmed that endothelial and smooth muscle cells in varicose and normal vein walls exhibited apoptotic morphologic features, such as fuzzy mitochondrial cristae, medullary changes, and margination of the nuclear chromatin. The results indicate that apoptosis is down-regulated in varicose venous walls, while the apoptotic rate of all normal great saphenous vein segments showed an opposite tendency compared with venous pressure. Our results also showed that expression of Bax proteins was opposite the positive expression of Bcl-xl proteins in both two groups within the three segments. We also found that varicose veins from the saphenous trunk within the three segments contained fewer Bax and Bcl-xl positive cells than healthy veins. No difference existed in the Bax and Bcl-xl detection indices in the saphenous trunk within the three segments. The results confirmed that immunoreactivity, as an indicator of apoptosis, was less prominent in the media layer of varicose veins than healthy veins, suggesting that fewer specific mediators regulate the intrinsic apoptosis pathway in varicose veins. Our findings in varicose veins contrast with the increased apoptosis and increased number of intrinsic pathway promotors, including the apoptotic promoter, Bax, reported by others in varicose veins of the oldest age group (>50 years); the percentage of apoptotic ECs and SMCs in the medial layer was increased. 22 These discrepancies presumably reflect the different tissues examined. The fibrotic degradation in the varicose venous wall is associated with dysregulation of apoptosis due to a decreased turnover of ECs and SMCs in the media layer.
We now provide evidence implicating the mitochondrial (intrinsic) pathway in the pathogenesis of human primary varicose veins. These findings suggest that deregulated programmed cell death through the intrinsic mitochondrial apoptosis pathway promoter is among the possible causes of primary varicose veins in humans. The intrinsic pathway can be induced by mitochondrial release of cytochrome c and various stimuli, including elevated levels of the pro-apoptotic Bcl-2 family and Bax. 4 By associating Bax and Bcl-xl in our study, we clearly showed that the decreased cell turnover was mediated by the intrinsic pathway. We suggest that hypoxia caused by venous stasis and high pressure could inhibit the apoptotic channels. 23 Indeed, hypoxia could cause cellular stress, including an increase in anti-apoptosis protein (Bcl-2), caspase-9, and mitochondrial outer membrane permeability, which block the mitochondrial channel.24,25 In contrast, Fas and caspase-8 are activated, which block the transmembrane channel.26,27 It has been confirmed that venous high pressure and hypoxia can reduce cell metabolism, and restrain apoptosis of varicose venous walls. Down-regulation of apoptosis in varicose venous walls could lead to cell proliferation, wall thickening, and an increase in the lumen.6,28
In conclusion, our results provide evidence of decreased programmed cell death in the medial layer within the three segments of human primary varicose veins. Generally, the apoptotic rate was higher in the upper segment than the lower segment and higher in the intima than the media. Assay of specific proteins indicated that the physiologic decrease in normal apoptosis was modulated through the intrinsic mitochondrial-dependent pathway. This observational study implicates apoptotic down-regulation in the vein walls as a cause of varicose veins, but does not exclude the possibility that other mechanisms are also responsible.
Footnotes
Authors’ contribution
XY and HW equally contributed to this paper.
Acknowledgments
We thank Bo Zhang and Lin Hou for technical assistance, Department of Pathology, and Medical Department of the Beijing University.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Supported by the Weifang City Sci-tech Development Project of China (Grant No. 2014zj1058).