
Abstract
Objectives
Superficial venous thrombosis (SVT) is the complications of varicose great saphenous veins (VGSVs), but its pathogenesis remains unclear. This study was designed to measure the changes in expression of matrix metalloproteinases (MMPs) and the tissue inhibitor of metalloproteinases (TIMPs) from SVT, VGSVs, and great saphenous veins (GSVs).
Methods
In the venous walls of the three groups, the expression of MMP-2, MMP-9, TIMP-1, and TIMP-2 proteins, protein-positive expression ratios, mRNA expression, and protein expression were determined by immunohistochemistry, polymerase chain reaction, and western blot.
Results
The MMP-2, MMP-9, TIMP-1, and TIMP-2 protein-positive expression ratios, mRNA and protein expression in the SVT group were significantly higher than those in the VGSV and the GSV groups. The corresponding expression in the VGSV group were significantly higher than those in the GSV group.
Conclusion
Disequilibrium of MMPs and TIMPs in SVT wall occurs due to underlying high hydrostatic pressure and inflammation. These results suggested that MMPs and TIMPs participate in the process of venous wall remodeling.
Keywords
Introduction
Varicose great saphenous veins (VGSVs) are a common venous disorder affecting approximately 25 million adults in the United States. 1 Several risk factors, such as advanced age, female gender, contraceptive pills and estrogen therapy, pregnancy, overweight and obesity, prior leg injury, vein inflammation, and phlebitis, may lead to VGSVs. 2 One of the complications of VGSVs is superficial venous thrombosis (SVT), which is characterized by endothelial, vein wall, and valve inflammation and leukocyte infiltration leading to disruption of vein function and venous thrombosis.3–5 SVT is inflammation of the superficial veins and is associated with venous thrombosis. 6 It has been shown to be higher than that of deep vein thrombosis, which is estimated at about one per 1000. 7
Matrix metalloproteinases (MMPs) are a family of zinc containing proteolytic enzymes that degrade and remodel the extracellular matrix (ECM). 8 Prolonged increases in venous pressure and vein wall tension are thought to increase the expression/activity of MMPs. 9 MMPs are inhibited by tissue inhibitors of metalloproteinases (TIMPs), which form high-affinity, essentially irreversible, non-covalent complexes with the active forms of MMPs and gelatinase zymogens.10–12 MMPs could play a role in various physiological processes and could affect cell proliferation, migration, and differentiation. MMPs could also be involved in cell apoptosis, immune response, tissue repair, and angiogenesis. 13
Previous studies have suggested that mechanical stretch may lead to increases in the expression of MMPs in endothelial cells, smooth muscle cells (SMCs), and fibroblasts, and that prolonged increases in mechanical tension or wall stretch of isolated rings of rat inferior vena cava (IVC) are associated with increased expressions of MMP-2 and MMP-9 in the tunica intima and an increased expression of MMP-9 in the tunica media layer of the vein wall.14,15 Prior experimental studies also has suggested that significantly higher MMP-2, MMP-9 mRNA expression was observed at the femoral vein thrombosis and the thrombosed IVC in rats.16,17 Our previous studies have shown that the MMP-2, MMP-9, TIMP-1, and TIMP-2 protein-positive expression ratios, mRNA and protein expression in the lower segments in the VGSV group were significantly higher than those in the upper and middle segments in the VGSV group. 18
SVT is the complications of VGSVs, but its pathogenesis remains unclear. Regarding the expression of MMPs and TIMPs in SVT, none has reported in the literature. In the present study, the distribution of MMP-2, MMP-9, TIMP-1, and TIMP-2 proteins in the walls of SVT was quantified by immunohistochemistry, polymerase chain reaction (PCR), and western blot. This information may help to understand the association between thrombosis and inflammation in SVT.
Materials and methods
Ethical approval of the study protocol (No. 1538) was obtained from the Human Research Ethics Committee of the 80th Group Army Hospital of People's Liberation Army (Weifang, China). All individuals provided written informed consent to participate in the study.
Materials
Methods
Specimen collection
Paraffin sections were obtained. A 3–4 mm sample was obtained from the trunk of SVT, VGSVs, and GSVs below the knee. Specimens were fixed in 10% formalin, dehydrated, embedded, and serially sliced into 4-µm sections. One slice of each sample was used for hematoxylin and eosin (H&E) staining. Each specimen was cut into 15 slices, with five slices per group. Immunohistochemistry of the sections were performed. In addition, fresh specimens were collected and immediately inserted into EP tubes, and then they were preserved in liquid nitrogen until used for PCR or western blotting.
Immunohistochemistry staining
Immunohistochemistry was performed as follows: Antigen retrieval was performed using ethylenediaminetetraacetic acid (pH = 8.5), 0.3% (v/v) hydrogen peroxide, and goat serum to block the non-specific antigen. The immunohistochemical SP method was performed. Sections were then incubated overnight at 4°C with the primary antibody. Rabbit anti-human MMP-2, MMP-9, TIMP-1, and TIMP-2 monoclonal antibodies (Zhongshan Golden Bridge Co., Ltd., Beijing, China) were used to label the corresponding antigens, followed by incubation at room temperature for 30 min with biotinylated secondary antibody with diaminobenzidine as the substrate, and then they were stained with hematoxylin. The negative control group was studied using the same steps described above, but primary antibody was used instead of phosphate-buffered saline.
Measurement of MMP-2, MMP-9, TIMP-1, and TIMP-2 mRNA expressions by fluorescence quantitative PCR
Venous tissues and total RNA were extracted in each group according to the instructions provided for TRIzol (Invitrogen, Carlsbad, CA, USA), and then they were dissolved in diethylpyrocarbonate (DEPC)-treated demineralized water and kept at 80°C. Fluorescence quantitative PCR was used with the SYBR Green PrimeScript RT-PCR Kit II (TaKaRa Co., Ltd., Dalian, China), using a Bio-RadIQ5 (Bio Rad, Hercules, CA, USA). Related primer sequences were designed and synthesized (TaKaRa Co., Ltd. (Dalian, China) (Table 1), a housekeeping gene (GAPDH) was used as an internal calibration in experiments, and ultrapure water (RNase-free) was used as a negative control. Results were analyzed using 2-ΔΔCt amplification efficiency in each group.
Related primer sequences and amplified fragment length of matrix metalloproteinase (MMP)-2, MMP-9, tissue inhibitor of metalloproteinases (TIMP)-1, and TIMP-2.

GAPDH: Glyceraldehyde phosphate dehydrogenase.
Measurement of MMP-2, MMP-9, TIMP-1, and TIMP-2 protein concentrations by western blot
Total cell protein was extracted according to the kit instructions, stored at 4°C, and centrifuged (2000 × g) for 5 min. The supernatant was added to the protein loading buffer and mixed well, placed in boiling water for 5 min, and then stored at −20°C. The stacking and separating gels were marked. The protein extract (in the form of the supernatant) was added after irrigation and separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis for 3.5 h. A transmembrane was used in a tank transfer system. A 200 mA current source was used for 30–120 min. Skim milk powder was added for 1 h. The primary antibody was incubated at 4°C overnight. Tris-buffered saline-Tween 20 was used for washing, with shaking for 5–10 min/count. The second antibody was incubated at room temperature for 1 h. Tris-buffered saline-Tween 20 was used for washing the gel, with shaking for 5–10 min/count, an enhanced chemiluminescence liquid was dropped, exposed, and colored in a dark room, and the protein bands were scanned and analyzed using Image-J software (National Institutes of Health, Bethesda, MD, USA).
Reagents and equipment
The immunohistochemical expression of MMP-2, MMP-9, TIMP-1, and TIMP-2 markers was purchased from Zhongshan Golden Bridge Co., Ltd. (Beijing, China). The SYBR kit, as well as designed and synthesized primer sequences, was purchased from. MMP-2, MMP-9, TIMP-1, and TIMP-2 ELISA kits were purchased from Jing Mei Co., Ltd. (Shenzhen, China). The following equipment was used: enzyme-labeled instrument ((Versa Max; Molecular Devices, Sunnyvale, CA, USA); light microscope (BX51; Olympus, Tokyo, Japan); fluorescence quantitative PCR (Bio-RadIQ5; Bio Rad, Hercules, CA, USA); and an automatic analyzer (ProteinSimple Ltd., Silicon Valley, CA, USA).
Criterion standard
Cells in which the cytoplasm had brown-yellow or dark brown granules were referred to as positive cells. Cells with no staining or the presence of light-yellow granules were referred to as negative cells. For the histological analysis, five fields per section were examined for staining at 200× magnification. The immunohistochemical expression of MMP-2, MMP-9, TIMP-1, and TIMP-2 immunoreactive proteins was determined according to the intensity of nuclear staining and the density and morphology of chromatin. The results were evaluated by two independent investigators who were blinded to the patients’ clinical findings. MMP-2, MMP-9, TIMP-1, and TIMP-2 immunoreactive proteins considered positive for the protein ratio (number of positive cells/total number of cells per high-power field) were counted. Under light microscopy (×200) (BX51; Olympus, Tokyo, Japan), Image-Pro Plus 6.0 (Media Cybernetics, Silver Spring, MD, USA) was used to calculate the positive protein ratios and determine the positive proteins.
Statistical analyses
Data analyses were performed with SPSS 22.0 software (SPSS, Inc., Chicago, IL, USA). All values are expressed as the mean ± standard deviation. Differences in measurement data were compared using one-way ANOVA and SNK-q method, Student’s t-test with two independent samples, and two paired samples. A P < 0.05 was considered statistically significant.
Results
The patients in the SVT group included 14 men and 8 women (mean age 35 years; range 28–45 years). The VGSV group included 13 men and 9 women (mean age 36 years; range 27–46 years). The control group included 9 men and 4 women (mean age 32 years; range 20–39 years).
Distribution of MMP-2, MMP-9, TIMP-1, and TIMP-2 protein expression in the venous walls of the three groups
In the tunica intima and tunica media layers of the SVT and VGSV groups, positive expression of MMP-2, MMP-9, TIMP-1, and TIMP-2 proteins were densely distributed in the intimal endothelial cells, medial smooth muscle cells, adventitial vasa vasorum, and fibroblast cells in venous walls; and in the SVT group, these expressions were more evident in the venous walls. In the GSV group, MMP-2, MMP-9, TIMP-1, and TIMP-2 proteins were not expressed, while individual or scattered areas of positive expression were observed (Figure 1).

The expression of MMP-2, MMP-9, TIMP-1, and TIMP-2 proteins in venous walls of three groups. The expression of the superficial venous thrombosis (SVT) group (A), the varicose great saphenous veins (VGSV) group (B), and the great saphenous veins (GSV) group (C) in MMP-2 protein; The expression of the SVT group (D), the VGSV group (E), and the GSV group (F) in MMP-9 protein; The expression of the SVT group (G), the VGSV group (H), and the GSV group (I) in TIMP-1 protein; The expression of the SVT group (J), the VGSV group (K), and the GSV group (L); (The arrow indicates positive expression, ×400, magnification, Bars =50 μm.
Determination of MMP-2, MMP-9, TIMP-1, and TIMP-2 protein-positive expression ratios in the venous walls of the three groups
In the SVT group (tunica intima and tunica media), MMP-2, MMP-9, TIMP-1, and TIMP-2 protein-positive expression ratios were significantly higher than those in the VGSV and GSV groups (all P < 0.05). The corresponding protein-positive expression ratios in the VGSV group were significantly higher than those in the GSV group (P < 0.05, Figure 2).
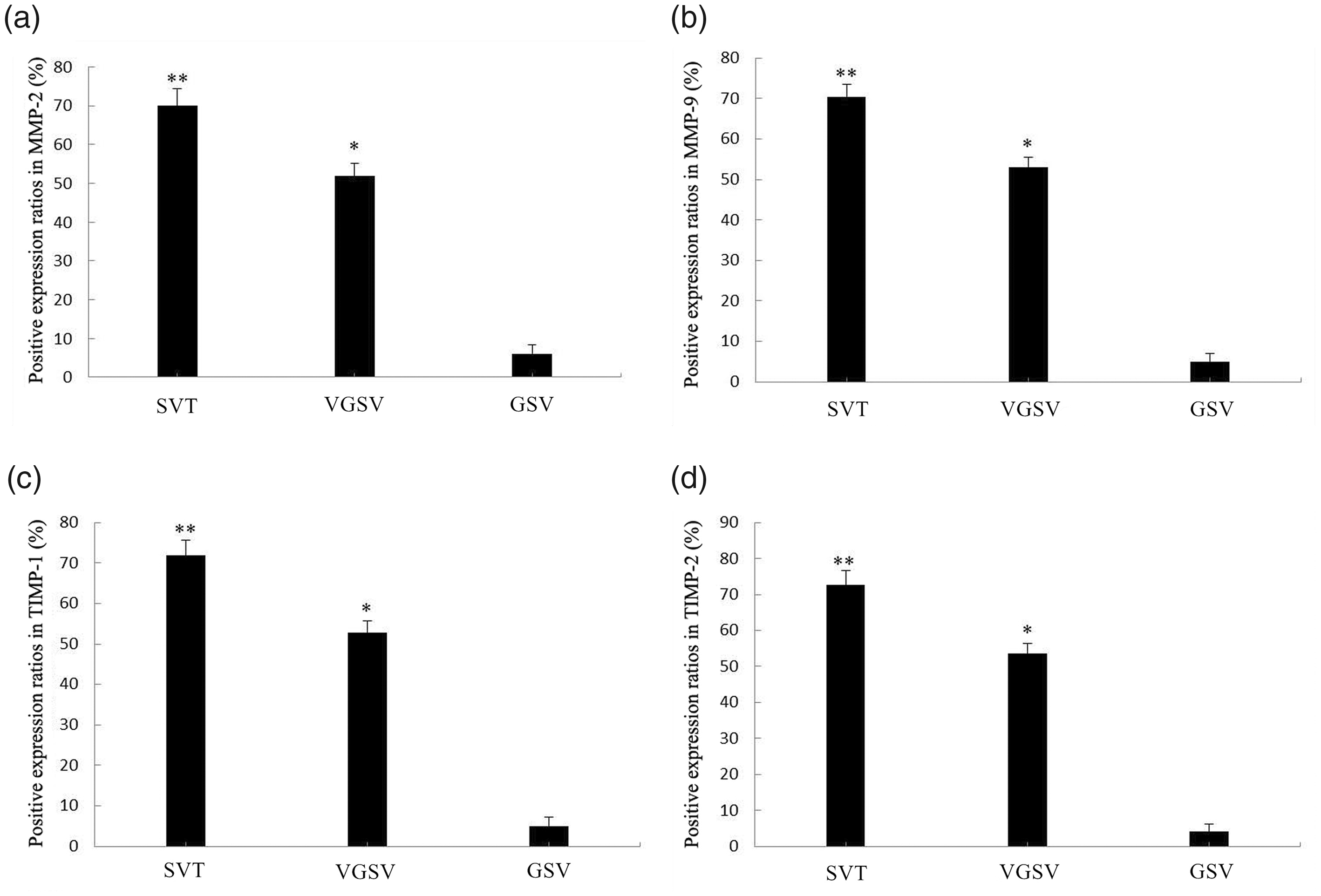
Positive expression ratios of MMP-2, MMP-9, TIMP-1, and TIMP-2 proteins in venous walls of three groups. Positive expression ratios of (A) MMP-2, (B) MMP-9, (C) TIMP-1, and (D) TIMP-2 proteins in the SVT, VGSV, and GSV groups. The error bars show the standard deviation. **P < 0.05, compared with in the VGSV and GSV groups, respectively; *P < 0.05, compared with in the GSV group.
Determination of MMP-2, MMP-9, TIMP-1, and TIMP-2 mRNA expression in the venous walls of the three groups
In the SVT group (tunica intima and tunica media), MMP-2, MMP-9, TIMP-1, and TIMP-2 gene expressions were significantly higher than those in the VGSV and GSV groups (all P < 0.05). The corresponding expression in the VGSV group were significantly higher than those in the GSV group (P < 0.05, Figure 3).
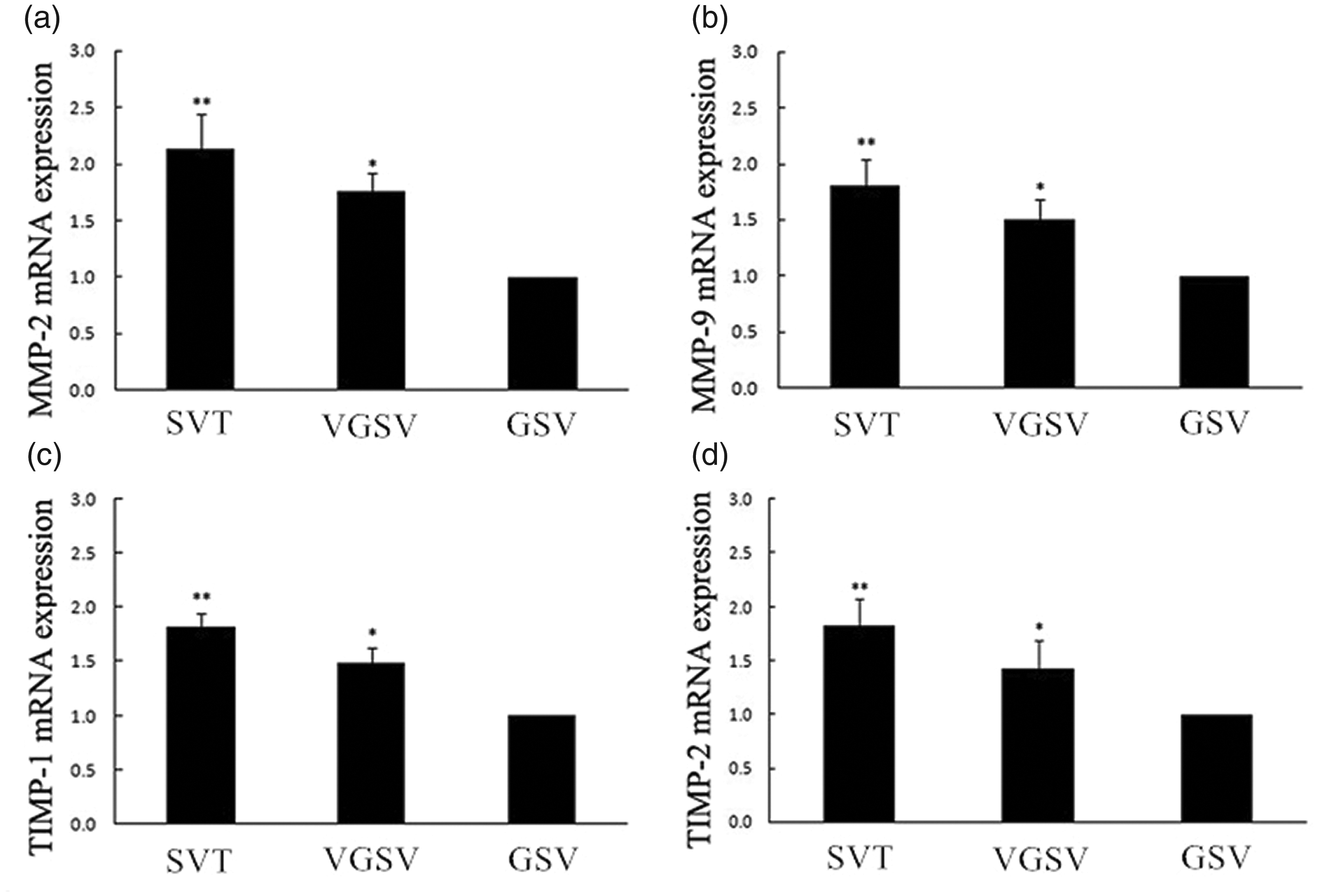
Messenger RNA (mRNA) expression of MMP-2, MMP-9, TIMP-1, and TIMP-2 in venous walls of three groups. mRNA expression of (A) MMP-2, (B) MMP-9, (C) TIMP-1, and (D) TIMP-2 in the SVT, VGSV and GSV groups. The error bars show the standard deviation. **P < 0.05, compared with in the VGSV and GSV groups, respectively; *P < 0.05, compared with in the GSV group.
MMP-2, MMP-9, TIMP-1, and TIMP-2 protein expression in the venous walls of the three groups
MMP-2, MMP-9, TIMP-1, and TIMP-2 protein expression in the SVT group were significantly higher than those in the VGSV and GSV groups (all P < 0.05). The corresponding protein expression in the VGSV group were significantly higher than those in the GSV group (P < 0.05, Figure 4).
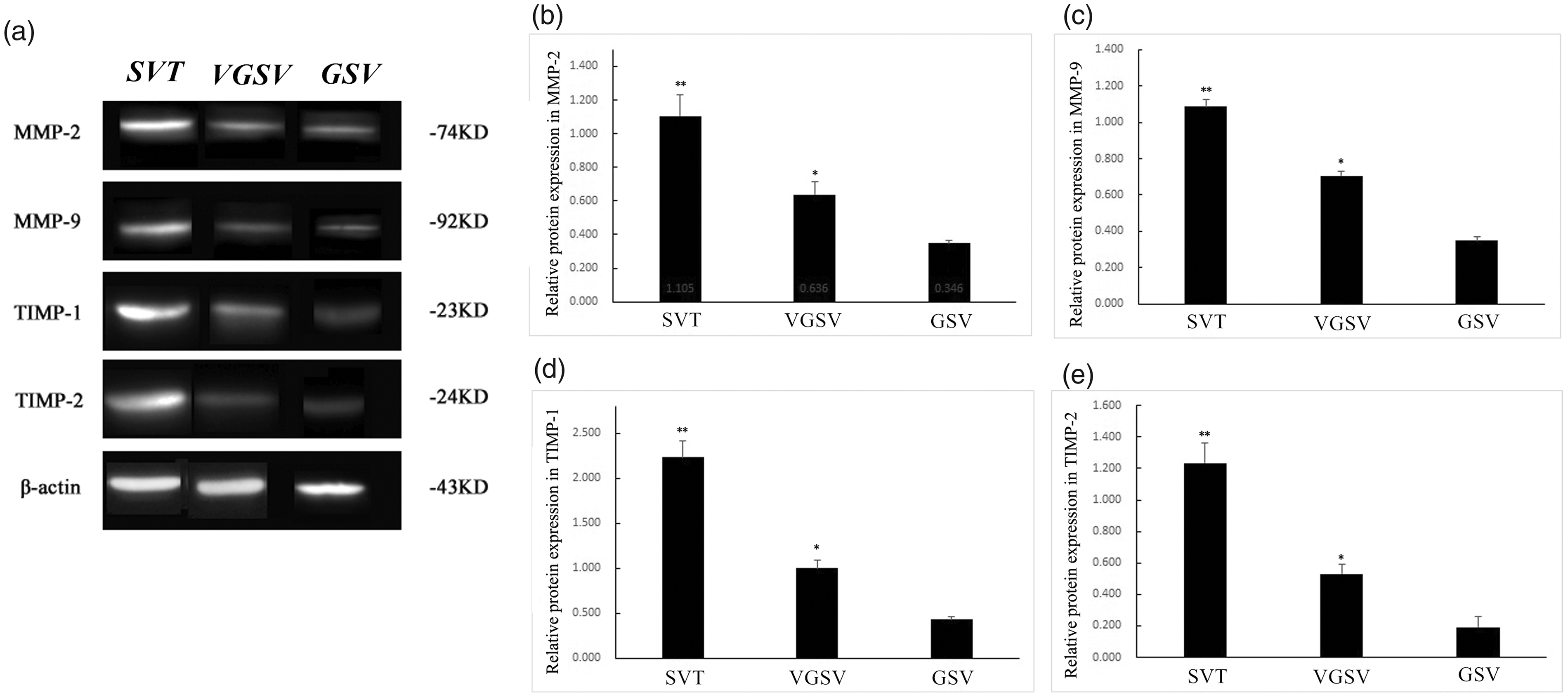
MMP-2, MMP-9, TIMP-1, and TIMP-2 protein expression in venous walls of three groups. (A) Bands of MMP-2, MMP-9, TIMP-1, TIMP-2, and β-actin proteins. The protein expression of (B) MMP-2, (C) MMP-9, (D) TIMP-1, and (E) TIMP-2 in the SVT, VGSV and GSV groups. The error bars show the standard deviation. **P < 0.05, compared with in the VGSV and GSV groups, respectively. *P < 0.05, compared with in the GSV group.
Discussion
Degradation and remodeling of the ECM by MMPs and TIMPs have been implicated in the pathogenesis of SVT. Because of the association of MMPs and TIMPs with VGSVs, we postulated that MMPs and TIMPs may play a role in the pathogenesis of VGSVs.9,19 In this study, we analyzed and compared tissue samples from patients with SVT, VGSVs, and GSVs to determine if there were any differences in the expression and localization of MMP-2, MMP-9, TIMP-1, and TIMP-2. We chose to perform immunohistochemistry of SVT, VGSVs, and GSVs with MMP-2, MMP-9, TIMP-1, and TIMP-2 antibodies. As compared to GSVs, intimal and adventitial expressions of MMP-9 were increased in 31 (49.2%) and 40 (63.5%) cases, respectively. Expression of TIMP-1 was absent in both VGSVs and GSVs, when assessed with MMP-9, and TIMP-1 antibodies. 20
Histologically, several morphological differences were evident between SVT, VGSVs, and GSVs. Focal areas of endothelial denudation, fragmentation, and attenuation of the elastic lamellae, loss of SMCs, and medial fibrosis in areas of thrombus adherence were the characteristic findings in SVT and to a lesser extent in VGSV specimens. The morphological changes in both SVT and VGSVs may only involve part of the circumference of the venous wall. 21 In SVT, distribution of more mast cells was observed around the vasa vasorum, and accumulation occurred in the adventitia, with respect to infiltration of T and B cells; each layer of the vein wall showed an increase. The most obvious increase was observed in the adventitia, followed by the intima. 22 Fragmentation of elastic tissue is a characteristic feature of SVT. It has not been determined whether fragmentation of the elastic tissue lamellae is a precursor to dilatation in the venous wall.
Vascular remodeling may take one of the several forms: (1) neointimal hyperplasia, (2) alteration of the venous structural luminal diameter (inward or outward), and (3) alteration of tunica media mass (eutrophic, hypertrophic, and hypotrophic). 23 In all pathologies involving matrix remodelling, an imbalance between the production of ECM proteins, MMPs and TIMPs, is observed. In VGSVs, the production of MMP-1, MMP-2, MMP-3 and MMP-7 was augmented in the intima and media layers. This increase in MMP expression reflects changes in the regulation of connective tissue. However, the level of TIMP labeling also appeared to be increased in VGSVs. 24 It is suggested that activities of MMPs are also under the control of TIMPs, and changes in the ratio of MMPs/TIMPs are probably involved in vascular wall remodeling and in varicose vein formation. 25 As some studies have suggested that transforming growth factor beta 1 (TGF-b1) may act differently in different phases of the disease resulting in either atrophy (increase in ECM degradation) or fibrosis (decrease in ECM degradation), the net proteolytic activity was not assessed in this simulated acute inflammatory phase. 26 Other studies have documented that MMPs are increased upon stimulation with inflammatory cytokines, such as IL-1β, TNF-α, and IL-6; thus, showing a correlation between MMPs and cytokines. 27 In healthy tissues, TIMPs appear to spare the ECM from degradation, and only following injury/infection, TIMPs appear to indirectly control ECM deposition. 28 These regulatory mechanisms underlying protein expression and post-translational modifications of MMP-2 and MMP-9 can be the targets of therapeutic interventions to ameliorate the abnormal tissue destruction observed in VGSVs. 29
MMPs activities are regulated at the following three different levels: transcriptional, activation of the proenzyme, inhibition by non-specific and specific inhibitors. Transcriptional regulation differs depending on the cell implicated in MMP production. Genes encoding for MMPs are modulated by several stimuli, such as growth factors, cytokines, cell–cell or cell–matrix interactions, stress, and chemical agents. In the cardiovascular system, macrophages and SMCs are induced to express MMPs by some cytokines, such as IL-1β and TNF-α, thrombin, high shear stress, and hypercholesterolemia.30,31 Interestingly, low concentrations of TIMP-2 induce MMP-2 activation, while higher concentrations inactivate MMP-2.Therefore, alterations in the pattern of expressions of MMPs and TIMPs by platelets may contribute to plaque instability, favor thrombus formation, or boost tissue inflammation. 32 It is conceivable that a dysregulated transfer of MMPs and TIMP transcripts from megakaryocytes to platelets may occur under pathologic conditions. 33 MMP-2, TIMP-1, TIMP-2, and plasminogen activator inhibitor type-1 levels were measured by enzyme-linked immunosorbent assay (ELISA). A high TIMP-1 level and a low MMP-2 level/activity were found in VGSVs, resulting in a three-fold increase in the TIMP-1/MMP-2 ratio in VGSVs versus GSVs.These results demonstrate that VGSVs are characterized by a higher TIMP/MMP ratio than GSVs, which may facilitate ECM accumulation in the diseased venous wall. 34
Our study also showed that MMP-2, MMP-9, TIMP-1, and TIMP-2 protein-positive expression ratios, mRNA and protein expression by immunohistochemistry staining, fluorescence quantitative PCR, and western blot in the SVT group were significantly higher than those in the VGSV and GSV groups. The expression of MMP-2, MMP-9,TIMP-1, and TIMP-2 assessed by fluorescence quantitative PCR and western blotting were quite variable and correlated with the immunohistochemical findings. In SVT, Our previous study also showed that the mRNA and protein expression of MMP-2, MMP-9, TIMP-1, and TIMP-2 in SMCs in VGSVs were significantly increased compared with those in GSVs. 35 Studies have shown a prominent increase in the MMP-2 activity in VGSVs. 24 Other studies have shown MMP-9 expression in the intimal and adventitial layers of VGSVs. 36 Other studies have shown an increase in the levels of MMP-1 and -13 in the proximal vs distal segments of VGSVs, with no change in the mRNA expression of MMPs, suggesting that increased levels of MMPs are related to changes in MMP posttranscriptional modification or protein degradation. 37 One study has shown that the levels of active MMP-1 and both pro- and active forms of MMP-2 are decreased in VGSVs. The variability in the levels of MMPs may be due to examination of different vein segments from different regions of VGSVs, i.e., hypertrophic vs atrophic regions, or examination of vein specimens from patients in different stages of VGSVs, or inability to distinguish between pro- and active forms of MMPs. Many factors could modulate the expression/activity of MMPs in VGSVs including increases in lower extremity venous hydrostatic pressure, inflammation of the vein wall, hypoxia, and other factors. 25
It has been reported in the literature that the SVT may have expression of different MMPs compared with VGSVs. 38 A study examining MMP-1, MMP-2, MMP-3, and MMP-9 in the SVT showed an elevated content of MMPs in the vein wall, and increased MMP-1, MMP-2, and MMP-9 activities. These marked changes in the content and activity of MMPs in SVT could lead venous tissue remodeling and alterations in the mechanical properties of the vein wall. 9 Studies have shown increased monocyte/macrophage infiltration in the walls and valves of GSVs from patients with chronic venous disease. 4 Cytokines, including interleukins IL-17 and IL-18,may induce MMP-9 expression via activation of transcription factor activator protein-1 and NF-κB-dependent pathways. 39 Therefore, the differences between our results and literature reports may be related to the location of the material, selected disease period, and age. The MMP-2, MMP-9, TIMP-1, and TIMP-2 protein expression, mRNA and protein expression were increased in the SVT. This finding is presumed to be a physiologic/pathologic compensatory performance, which helps prevent fibrosis of the vessel wall. Thus, it can be confirmed that the histopathological changes lay the molecular biological basis for the imbalance between MMPs and TIMPs. We believe that high pressure caused by a thrombus in the venous lumen and inflammatory cell infiltration in the venous wall are the main factors that lead to severe disequilibrium of MMPs and TIMPs in SVT.
Limitations
This study has several limitations. Evaluation of SVT represents the complications of VGSVs and may not reflect the factors that initiate or contribute to the pathogenesis of these lesions. The amount of tissue available for analysis remains an issue. Because of their rarity, it is unlikely that prospective studies of SVT will be undertaken. A multicenter registry and prospective collection of tissue samples may provide answers to some issues.
Conclusion
We have demonstrated increased expression of MMP-2, MMP-9, TIMP-1, and TIMP-2 in SVT compared with VGSVs and GSVs in this preliminary study. This observation in conjunction with the morphologic changes in the ECM of SVT suggests a possible causal role of MMPs and TIMPs in the pathogenesis of SVT.
Footnotes
Acknowledgements
We would like to thank Institute of Pharmacology and Toxicology in Academy of Military Medical Sciences for their guidance and help to us in the design of this study and data processing.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by Military Logistic Project of China (AWS17J008).
Ethical approval
The ethics committee of the 80th Group Army Hospital of People’s Liberation Army (Weifang, China) approved this study (REC number: 1538).
Guarantors
HX Zhan and YX Zhong.
Contributorship
GY, KL, HZ, and YZ researched literature and conceived the study. YX and HC were involved in protocol development, gaining ethical approval, patient recruitment, and data analysis. GY and KL wrote the first draft of the manuscript. HZ and YZ provided helpful and constructive advices in the revision. All authors reviewed and edited the manuscript. All authors read and approved the manuscript.