
Abstract
We employed two event-related functional magnetic resonance imaging tasks using the pictures of mild and intense facial emotions of fear or happiness. The sample comprised 16 chronic schizophrenia patients treated with risperidone long-acting injections (RLAI), 16 patients treated with conventional antipsychotic depots (CONV) and 16 healthy controls (HC). The HC and RLAI groups demonstrated greater activation in the left amygdala in response to intensively fearful faces, and in right cerebellum to intensively happy faces compared with CONV patients. The CONV group demonstrated under-activation in the right temporal pole in response to intensively happy faces (compared with HC) and over-activation in ventro-medial prefrontal cortex (VMPFC) in response to both intensively happy and fearful expressions, compared with HC and RLAI groups. Our results suggest that networks implicated in the allocation of attentional resources (VMPFC) and emotion processing (amygdala, cerebellum) are differentially affected in patients on CONV versus RLAI.
Introduction
The ability to recognize facial emotional expressions constitutes an important aspect of social cognition (Green et al., 2008) and is involved in guiding interpersonal relationships. Facial emotion processing deficits have been well documented in people with schizophrenia (Edwards et al., 2002; Kee et al., 2006; Schneider et al., 2006; Tsoi et al., 2008). Recent neurocognitive (Sergi et al., 2006, 2007) and neuroimaging (Pinkham et al., 2008) studies have added to the understanding of how abnormal social cognition may contribute to impaired social functioning.
Importantly, facial emotion recognition deficits have been found to be stable over the course of illness, being present in the acute state and in remission (Wolwer et al., 1996; Gur et al., 2003). These deficits do not seem to be merely an effect of antipsychotic medication, since they are also found in medication-free first-episode patients (Herbener et al., 2005).
Interest in this aspect of the neuropsychology of schizophrenia is supported by the assertion that the above deficits may be biomarkers of the illness (Gur et al., 2007a) given the reports on the heritability of facial emotion discrimination (Bediou et al., 2007), emotion intensity discrimination (Gur et al., 2007b) and composite measures of emotion recognition dysfunction (vocal, facial) which have been found in the first-degree relatives of people with schizophrenia (Kee et al., 2004).
Functional neuroimaging studies have provided information on brain abnormalities associated with facial affect processing in schizophrenia. For example, investigators have found reduced or absent amygdalar and hippocampal responses to fearful facial expressions (Gur et al., 2002; Michalopoulou et al., 2008; Li et al., 2009) associated, in particular, with paranoid symptoms (Phillips et al., 1999; Williams et al., 2004; Habel et al., 2005a) and have demonstrated reduced amygdala and orbitofrontal cortex activity to both sad and happy faces. There are also reports of a relative increase in amygdalar response to happy but not fearful expressions (Kosaka et al., 2002) and increased right amygdala activation during passive viewing of fearful and neutral faces (Holt et al., 2006). Thus, the neuroimaging findings show various kinds of abnormalities in brain response to facial emotions which may reflect clinical heterogeneity of samples as well as differential effects of the paradigms employed.
There is no consensus whether or not the abnormalities in emotional expression processing are amenable to treatment. If they are, what is the treatment of choice? Facial stimuli have served as a useful neuroimaging tool in the investigation of the effect of treatment in other disorders, e.g., major depression (Fu et al., 2004, 2007; Keedwell et al., 2009); however, there has been less research on the effects of psychopharmacotherapy on emotion processing in schizophrenia.
An earlier study (Lewis and Garver, 1995) showed that short-term treatment with haloperidol did not improve the facial affect recognition deficit in individuals with schizophrenia. There are indications of a beneficial effect of risperidone versus haloperidol on the ability of schizophrenia patients to recognize expressed emotions (Kee et al., 1998). This differential effect was corroborated by a study reporting improved visual scan paths to facial emotion expressions following treatment with risperidone versus haloperidol (Williams et al., 2003). A recent randomized controlled prospective study (Fakra et al., 2007) found that patients treated with risperidone, but not haloperidol, improved in facial affect recognition, which was not accounted for by the improvement in visuospatial abilities. The injectable form of risperidone (risperidone long-acting injections [RLAI]) provides for reliable drug delivery potentially combined with the beneficial effects of a second-generation antipsychotic.
To the best of the authors’ knowledge, there have been no published studies looking at the effect of RLAI on emotion processing aspects of social cognition. In this study we compared brain responses detected by functional magnetic resonance imaging (fMRI) during implicit emotional processing experiments in patients treated with conventional depot antipsychotics and those on RLAI.
Methods and materials
Participants
Thirty-two patients with a DSM-IV diagnosis of schizophrenia and 16 healthy control volunteers (HC) entered the study. Diagnosis was verified by the researcher using the Structured Clinical Interview for DSM-IV (SCID) (First et al., 2001). Patients were included in the study if they were 18–65 years old, and were undergoing maintenance treatment with either conventional depot or RLAI for schizophrenia for at least three months. All patients were recruited from a geographically defined catchment area. None had experienced a significant clinical exacerbation in the preceding three months. All study subjects were right-handed, as assessed by the Edinburgh inventory (Oldfield, 1971). Exclusion criteria for all participants were previous history of head injury resulting in a loss of consciousness for more than 5 min, contraindication to undergoing a MRI scan, drug or alcohol dependence for at least three months prior to the assessment as determined by a review of the participants medical records. The patients with schizophrenia did not present with diagnosable comorbid depression, anxiety disorder or personality disorders. The HC were recruited from the same community and were screened for past psychiatric disorder.
Among the patients with schizophrenia, 16 were treated with RLAI, and 16 with conventional depot medication (CONV). Conventional depots included: haloperidol decanoate (1 patient, 25 mg per week), flupentixol decanoate (12 patients, average dose 26.25 mg per week), fluphenazine decanoate (two patients, average dose 25 mg per week), and pipothiazine palmitate (one patient, dose 12.5 per week). The patients on conventional depot should have been on the current medication for at least three months. Approximate mean chlorpromazine equivalents of conventional depot were 232 mg/day (Goldberg and Murray, 2006). The doses of RLAI ranged from 25 to 37.5 mg bi-weekly. These are suggested to be as effective as oral doses of 2–6 mg of risperidone (Goldberg and Murray, 2006).
Prior to commencing on RLAI, all patients of that group had been on conventional depot antipsychotics for at least three months. These included flupentixol decanoate (eight patients, mean weekly dose 20 mg), fluphenazine decanoate (two patients, mean weekly dose 25 mg), haloperidol decanoate (two patients, mean weekly dose 22.5mg), and zuclopenthixol decanoate (four patients, mean weekly dose 75 mg).
The allocation of patients to the treatment group was naturalistic, as determined by the responsible clinicians. The study was approved by the Ethics Committee of Institute of Psychiatry and South London & Maudsley NHS trust. All subjects gave their informed written consent prior to participation in the experiments; the study was carried out in accordance with the Declaration of Helsinki.
Clinical rating scales
Demographic and clinical data
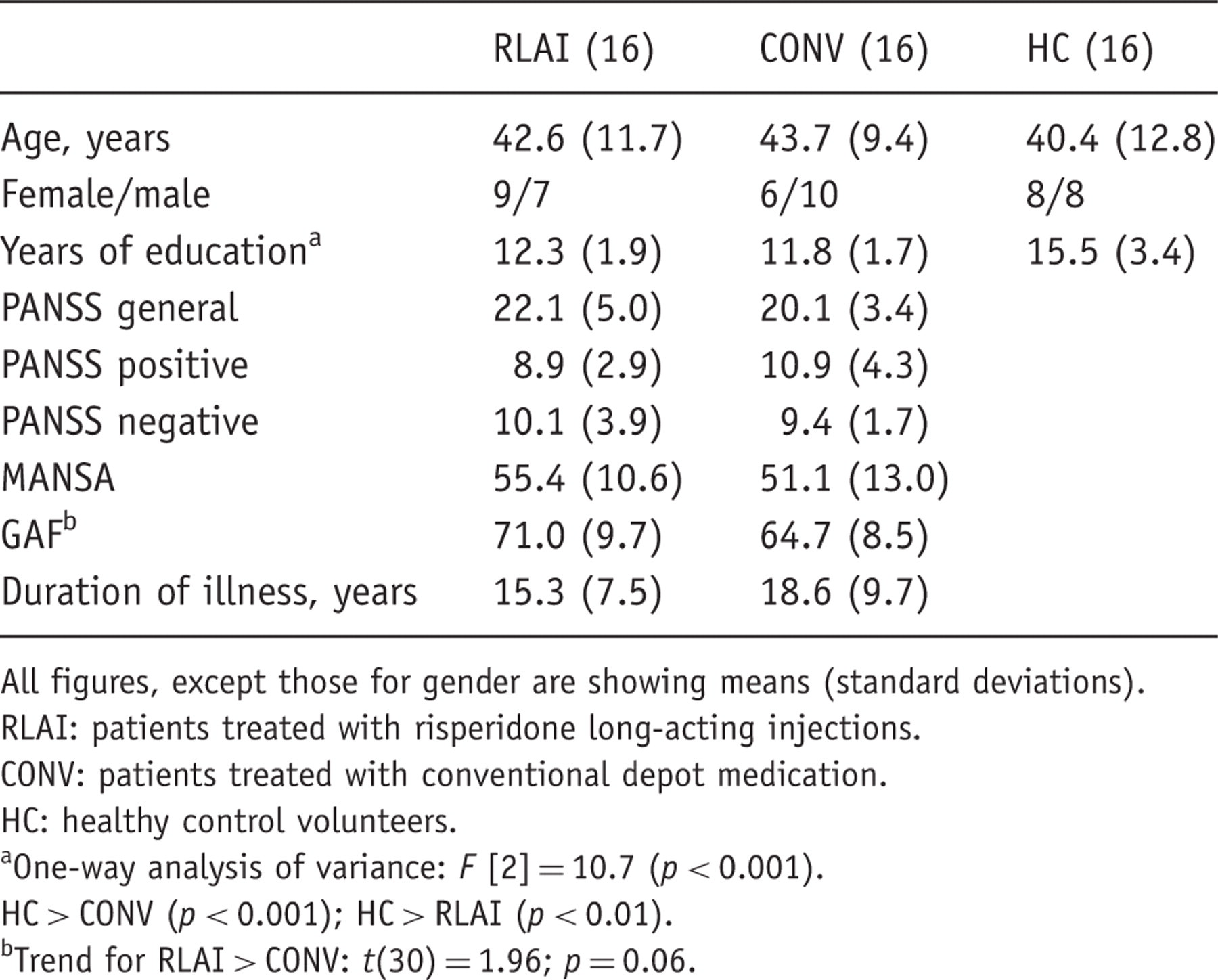
All figures, except those for gender are showing means (standard deviations).
RLAI: patients treated with risperidone long-acting injections.
CONV: patients treated with conventional depot medication.
HC: healthy control volunteers.
One-way analysis of variance: F [2] = 10.7 (p < 0.001).
HC > CONV (p < 0.001); HC > RLAI (p < 0.01).
Trend for RLAI > CONV: t(30) = 1.96; p = 0.06.
Experimental design and acquisition parameters
Study subjects participated in two 6-minute experiments employing event-related fMRI. In each experiment, subjects were presented with facial identities depicting morphed expressions of one emotion (either happiness or fear), and with faces expressing no emotion (neutral faces). For example, in an experiment with fearful facial expressions, there were 20 neutral faces, 20 with mildly (50%) fearful and 20 with intensively (100%) fearful expression, interspersed with the fixation cross. Another experiment included 20 neutral, 20 mildly (50%) happy and 20 intensively (100%) happy faces. The facial expression stimuli were chosen from the standard set of pictures from ‘Facial expressions of emotion: stimuli and tests’ (FEEST) (Young et al., 2002). We adopted an event-related design with a variable inter-stimulus interval to reduce habituation of the BOLD response in regions such as the amygdala, since habituation is a problem in block designs with highly repetitive and predictable stimulus presentation. The task employed in this study has proved to be sensitive in detecting an effect of emotion in different groups, including individuals with schizophrenia (Surguladze et al., 2006).
Each facial stimulus was presented for 2 seconds. During the inter-stimulus interval, the duration of which was varied from 3 to 8 seconds according to a Poisson distribution with an average interval of 5.9 seconds, subjects viewed a fixation cross. Since the identities were the same in both fear and happy emotion tasks, the tasks were counterbalanced across the subjects, to avoid the effects of identity recognition. The gender of actors was counterbalanced across the trials. There were 36 trials with female and 24 with male faces.
Subjects were requested to decide upon the sex of each face and press one of two buttons accordingly with the right thumb with no reference made to emotion. Previous studies have demonstrated that this implicit emotion processing task is reliably associated with responses in limbic and extrastriate cortical regions (Morris et al., 1996; Phillips et al., 1997).
Gradient echo echoplanar imaging (EPI) data were acquired on a GE Signa 1.5 T system (General Electric, Milwaukee, WI, USA) at the Maudsley Hospital, London. A quadrature birdcage headcoil was used for RF transmission and reception. One hundred and eighty T2 *-weighted images depicting BOLD contrast (Ogawa et al., 1992) were acquired over each of 16 near-axial non-contiguous 7-mm-thick planes parallel to the inter-commissural (AC-PC) line: TE 40 ms, TR 2 s, in-plane resolution 3.44 mm, inter-slice gap 0.7 mm. In the same scanning session a gradient echo EPI dataset was acquired at 43 near-axial 3-mm-thick planes parallel to the AC-PC line: TE 73 ms, TI 80 ms, TR 3 s, in-plane resolution 1.72 mm, inter-slice gap 0.3 mm. This higher-resolution EPI dataset provided whole brain coverage and was later used to register the fMRI datasets acquired from each individual in standard stereotactic space.
Individual and group brain activation analysis
The fMRI data were analysed with XBAM software developed at the Institute of Psychiatry (Bullmore et al., 1996; Brammer et al., 1997), using a non-parametric approach to minimize assumptions (see http://brainmap.it).
Data were first processed (Bullmore et al., 1999a) to minimize motion-related artefacts and correct for the slice time acquisition delay. A 3D volume consisting of the average intensity at each voxel over the whole experiment was calculated and used as a template. The 3D image volume at each timepoint was then realigned to this template by computing the combination of rotations (around the x-, y- and z-axes) and translations (in x, y and z) that maximized the correlation between the image intensities of the volume in question and the template. Following realignment, data were then smoothed using a Gaussian filter (full-width half-maximum [FWHM] 7.2 mm) to improve the signal-to-noise characteristics of the images.
Three different levels (neutral face, mild emotion and intense emotion) of stimulation were presented, in a randomized order and contrasted with a fixed background (fixation cross). Responses to the experimental paradigms were then detected by first convolving each component of the experimental design with each of two gamma variate functions (peak responses at 4 and 8 seconds, respectively). The best fit between the weighted sum of these convolutions and the time series at each voxel was computed using the constrained BOLD effect model suggested by Friman et al. (2003). This reduces the possibility of the model fitting procedure giving rise to mathematically plausible, but physiologically implausible results. Following computation of the model fit, a goodness-of-fit statistic was computed. This consisted of the ratio of the sum of squares of deviations from the mean image intensity (over the whole time series) due to the model to the sum of squares of deviations due to the residuals (SSQratio). This statistic is used to overcome the problem inherent in the use of the F (variance ratio) statistic that the residual degrees of freedom are often unknown in fMRI time series due to the presence of coloured noise in the signal. It has also been shown (Edgington, 1995) to behave equivalently to F under permutation testing. Following computation of the observed SSQratio at each voxel, the data are permuted by the wavelet-based method described and extensively characterized in Bullmore et al. (2001). Repeated application of this method at each voxel followed by recomputation of the SSQratio from the permuted data allows (by combination of results over all intracerebral voxels) the data-driven calculation of the null distribution of SSQratios under the assumption of no experimentally determined response. Using this distribution it is possible to calculate the critical value of SSQratio needed to threshold the maps at any desired type I error rate. The detection of activated voxels is extended from the voxel to the cluster level using the method described in detail by Bullmore et al. (1999b). In addition to the SSQratio, the size of the BOLD response to each experimental condition is computed for each individual at each voxel as a percentage of the mean resting image intensity level. In order to calculate the BOLD effect size, the difference between the maximum and minimum values of the fitted model for each condition is expressed as a percentage of the mean image intensity level over the whole time series. The observed and permuted SSQratio maps for each individual, as well as the BOLD effect size maps are transformed into the standard space of Talairach and Tournoux (1988) using the two-stage warping procedure described in detail by Brammer et al. (1997). This involves first computing the average image intensity map for each individual over the coarse of the experiment. The transformations required to map this image to the structural scan for each individual and then from ‘structural space’ to the Talairach template are then computed by maximizing the correlation between the images at each stage. The SSQratio and BOLD effect size maps are then transformed into Talairach space using these transformations. Group activation maps are then computed by determining the median SSQratio at each voxel (over all individuals) in the observed and permuted data maps (medians are used to minimize outlier effects). The distribution of median SSQratios over all intracerebral voxels from the permuted data is then used to derive the null distribution of SSQratios and can be thresholded to produce group activation maps at any desired voxel- or cluster-level type I error rate. Cluster-level maps are thresholded at less than half the expected type I error cluster per brain. The computation of a standardized measure of effect SSQratio at the individual level, followed by analysis of the median SSQratio maps over all individuals treats intra- and inter-subject variations in effect separately, constituting a mixed effect approach to analysis which is deemed desirable in fMRI.
Group by condition interaction analysis
The data pertaining to fearful and happy faces experiments were analysed separately. Voxel- and cluster-wise between-group differences in BOLD signal change to the three different types of facial expression versus baseline were examined using a 3 × 3 repeated measures analysis of variance (ANOVA), with intensity (neutral, 50% emotion, 100% emotion) as the within-subject variable, and group (RLAI, CONV, HC) as the between-group variable. This analysis allowed for the examination of any main effect or interaction between group and expression intensity, using whole brain statistical maps. In further between-groups analyses we used non-parametric Kruskal–Wallis and Kolmogorov–Smirnov (K–S) tests. The latter is considered to be as powerful as the parametric t-test, but has an advantage in analysing relatively small samples allowing for a comparison of the entire distribution, not just measures of central tendency (Pett, 1997).
Since we were specifically interested in amygdala functioning (among other structures) under different medications, we explored amygdala activation, using a region of interest (ROI) approach. To obtain the BOLD effect values pertaining to amygdala activation across the three groups, the data were extracted from the spherical regions with the centre coordinates: x = ±22, y = −5, z = −16 and diameter of 6 mm, based on atlas coordinates (Talairach and Tournoux, 1988). This mask was then applied to the data pertaining to each participant.
Results
Socio-demographic and clinical data
Healthy individuals did not differ from the schizophrenia patient groups in terms of age, although they had spent significantly more years in education. The two patient groups did not differ in either of these variables. There was no significant difference between the three groups in race and sex distribution. There was no significant difference between the patients groups in terms of duration of illness, or quality of life (MANSA), but there was a trend towards better GAF scores in the RLAI group compared with the CONV group: t(30) = 1.96; p = 0.06 (Table 1).
There was no significant difference between the patient groups in total scores of the PANSS or for positive, negative or general subscales.
On-line performance
One-way ANOVAs were performed for the per cent correct measure of gender decision for each condition in each group. There was no significant difference between groups in any condition: F[2,44] = 0.63, p = 0.54 in the fearful faces condition and F[2,45] = 2.2, p = 0.13 in the happy faces condition.
Brain activation during processing of fearful faces
Generic brain activation to fearful faces: per group and condition
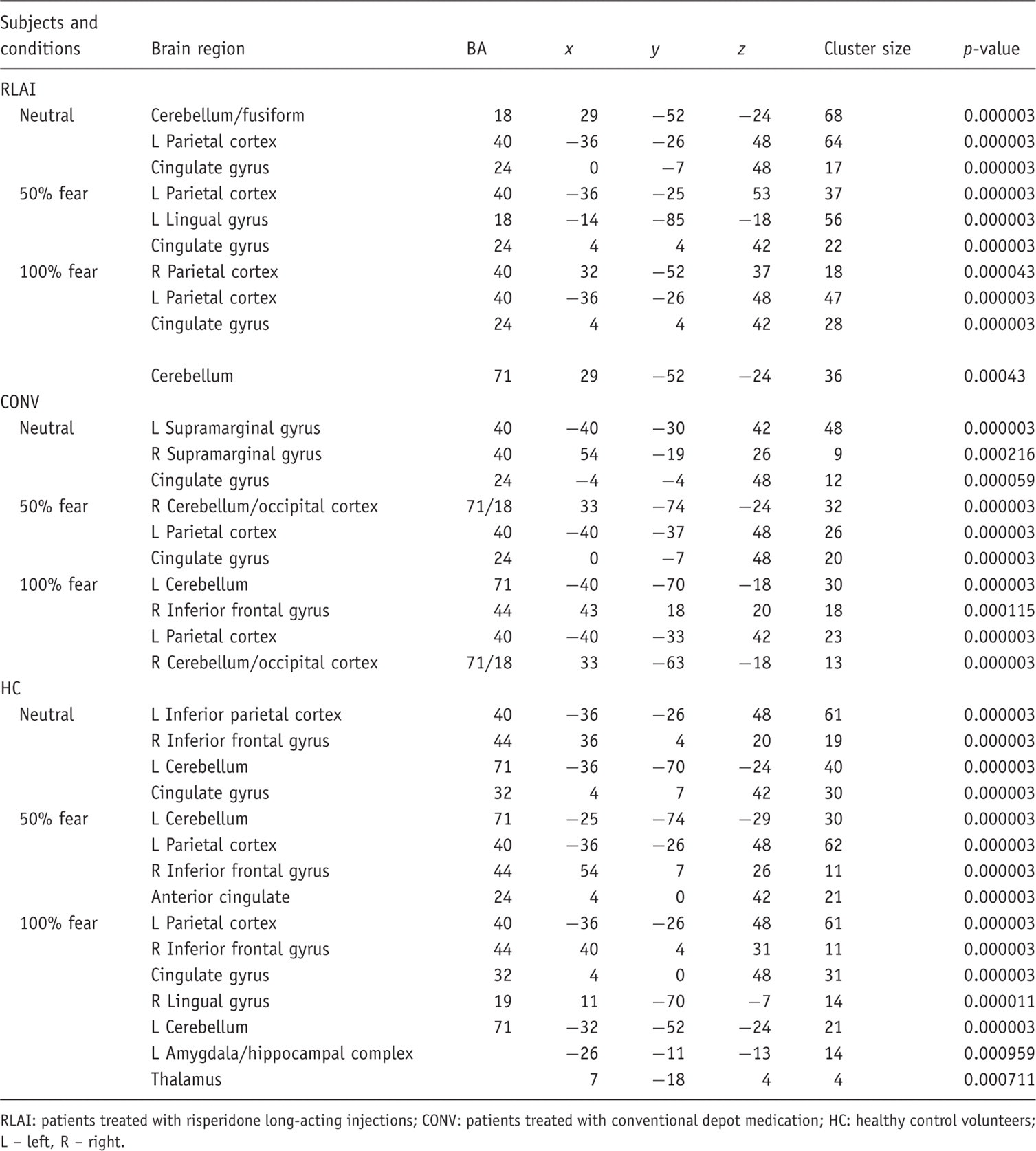
RLAI: patients treated with risperidone long-acting injections; CONV: patients treated with conventional depot medication; HC: healthy control volunteers; L – left, R – right.
To identify specific brain areas differentially activated in the three groups with regards to three emotion conditions, 3 × 3 repeated ANOVA of the whole brain activation data was performed with intensity (neutral, mild, intensive fear) as a within-subject variable and group (RLAI, CONV, HC) as a between-group variable. At the significance level p = 0.001, there was a probability of less than one false a positive cluster to be observed per whole brain map. The analysis produced a cluster reflecting a group by condition interaction in ventro-medial prefrontal cortex (VMPFC), BA 10/11, voxel size 120, with the centre of mass in x = −4; y = 44; z = −13, the borders of this cluster extended from x = −4/7; y = 44/52; z = −7/–18.
To clarify what group and condition accounted for the above-mentioned differential activation, further analyses were carried out. A Kruskal–Wallis test for three independent groups showed a significant effect for intensively fearful faces: χ2 [2] = 7.3; p = 0.026. In this condition, two-sample K-S tests showed significantly greater activation in CONV compared with HC (Z = 1.6; p = 0.013) and CONV compared with RLAI (Z = 1.4; p = 0.037); see Figure 1.
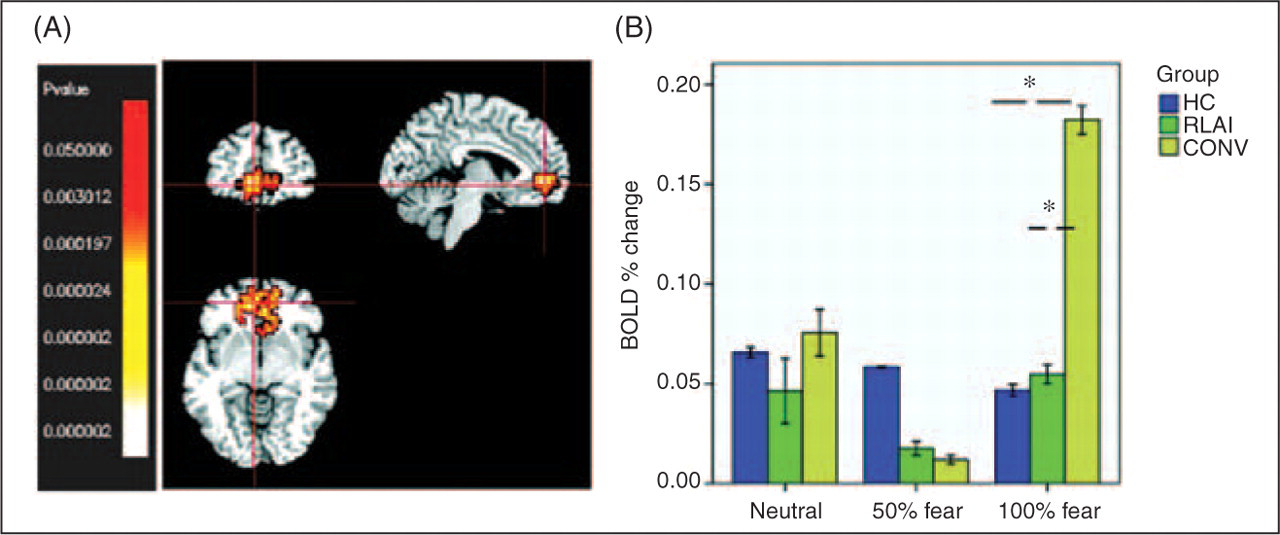
There was no significant difference between groups in activation of the right amygdala to any facial stimuli (Kruskal–Wallis test). The left amygdala demonstrated a differential activation in the intensively fearful face condition: Kruskal–Wallis test χ2 [2] = 9.2; p = 0.01. Two sample K-S tests showed the greater activation of left amygdala in RLAI compared with CONV patients (Z = 1.4; p = 0.037), and the HC compared with the CONV group (Z = 1.4; p = 0.037); see Figure 2.
Brain activation during processing of happy faces
Generic brain activation to happy faces – per group and condition
RLAI: patients treated with risperidone long-acting injections; CONV: patients treated with conventional depot medication; HC: healthy control volunteers; L – left, R – right.
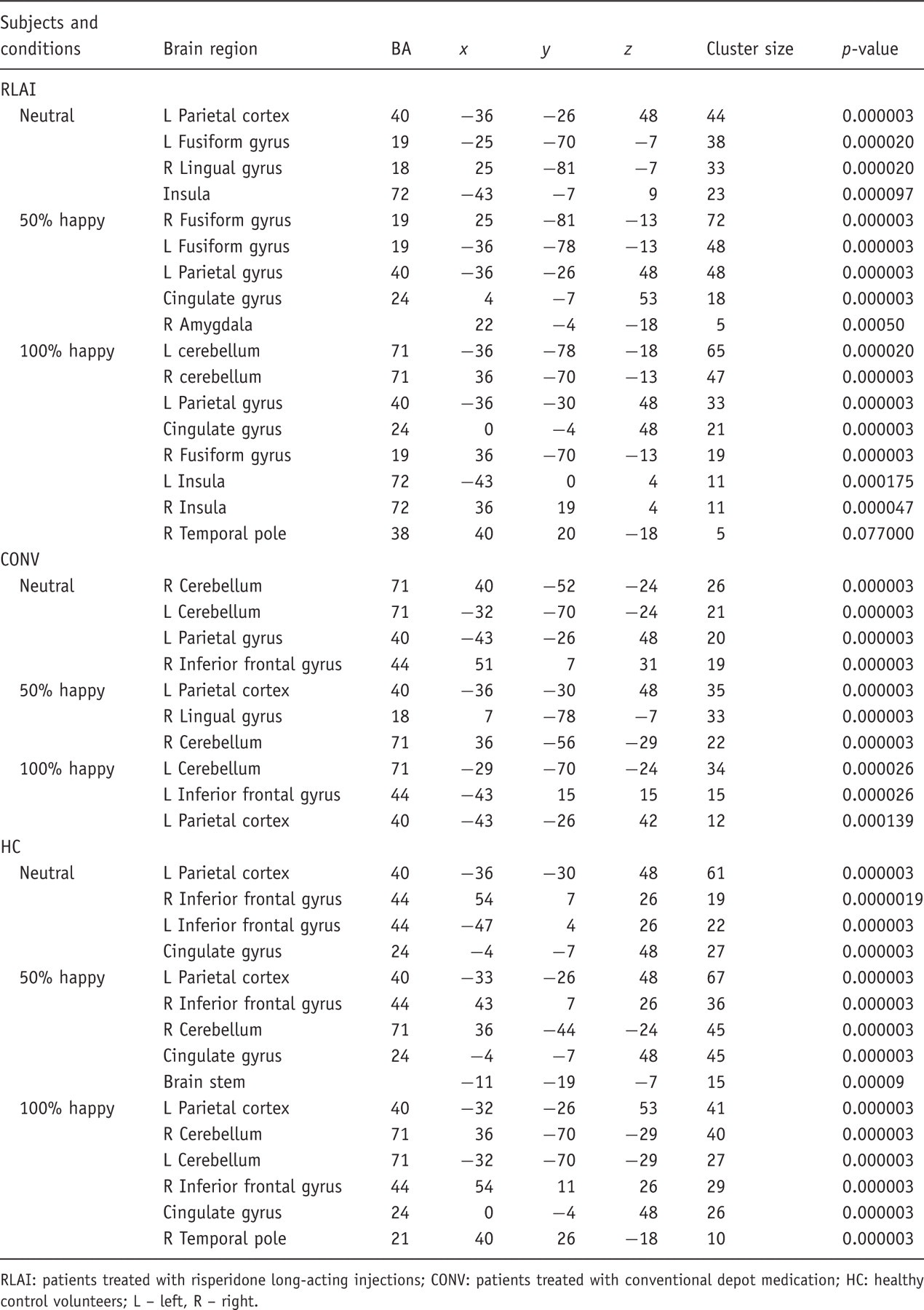
To find out whether there was a significant activation difference in three intensity conditions across the subject groups, a 3 × 3 repeated measures ANOVA with happy emotion intensity (neutral, mild, intensive) as a within-subject variable and group (RLAI, CONV, HC) as a between-group variable was performed. At the significance level p = 0.001 there was a probability of less than one-half false positive cluster to be observed per the whole brain map. This analysis produced three clusters reflecting interactions of group by condition: a cluster in VMPFC (BA 10) with coordinates x = 7, y = 48, z = −7 (cluster size 50 voxels) overlapping with the cluster found in the fearful face analysis; a cluster in right temporal pole (TP) BA 38 (x = 39, y = 15, z = −24; cluster size 92 voxels) and a cluster in right cerebellum (x = 39, y = −48, z = −35, cluster size 48 voxels). Each cluster was analysed separately for the effect of group and condition.
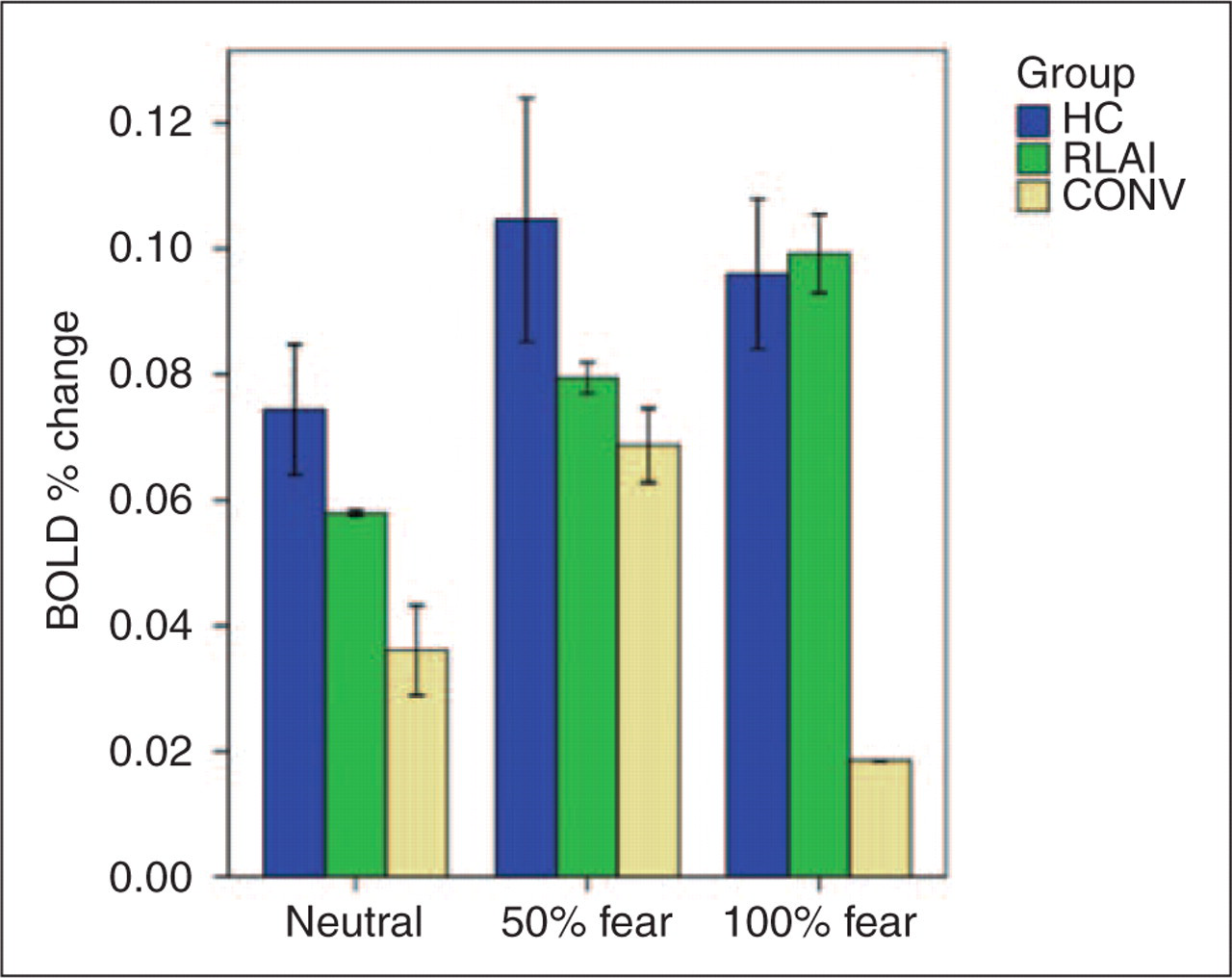
A Kruskal–Wallis test of activation in VMPFC showed a significant group effect in intensively happy condition (χ2 = 6.1; p < 0.05). This effect was accounted for by the greater activation in CONV compared with the HC group (between-groups K-S tests: Z = 1.4; p = 0.037) and greater activation in CONV compared with RLAI patients (K-S test: Z = 1.4; p = 0.037); see Figure 3.

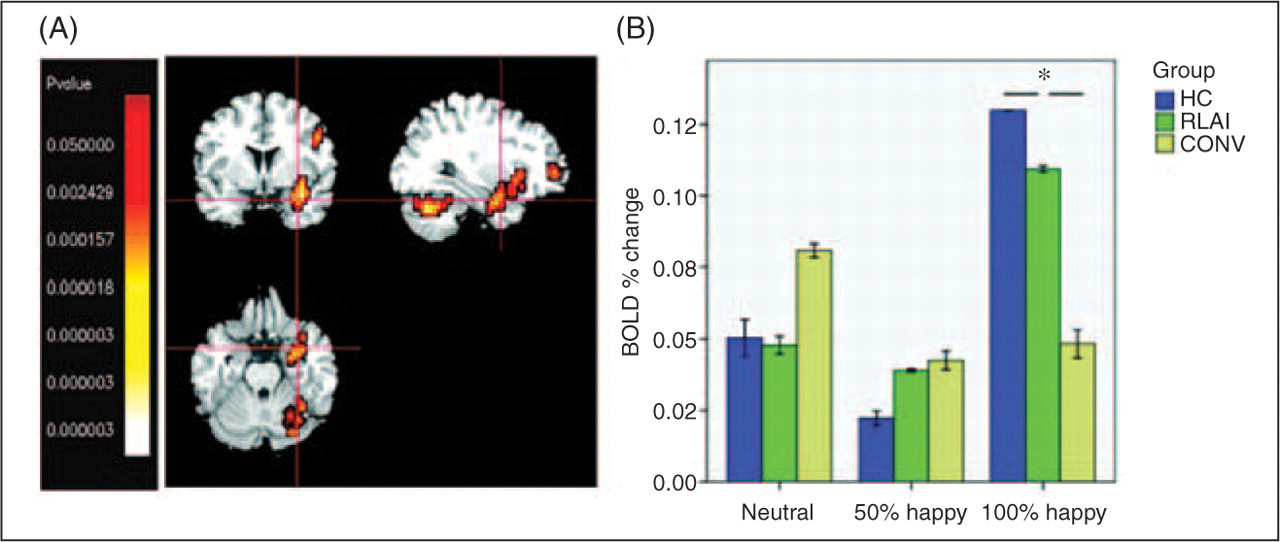
A Kruskal–Wallis test of activation in TP (BA 38) showed a significant group effect in intensively happy condition: χ2 [2] = 8.8; p = 0.012; K-S showed a significantly greater activation in HC compared with CONV: Z = 1.4; p = 0.03; see Figure 4.
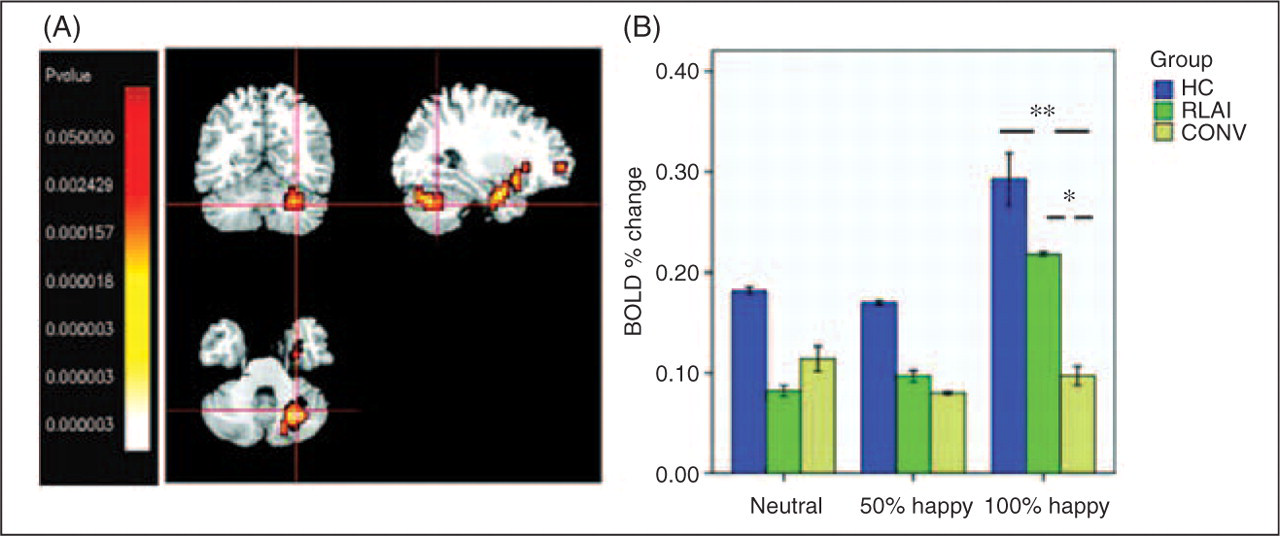
A cluster in the right cerebellum represented a significant interaction of group by condition. A Kruskal–Wallis test showed an effect of group in the intensively happy condition: χ2 [2] = 8.8; p = 0.012. K-S test showed a significantly greater activation in HC compared with CONV (Z = 1.8; p = 0.004), and greater activation in RLAI compared with CONV (Z = 1.6; p = 0.013); see Figure 5.
Using the ROI-extracted data on the amygdala activation, the one-way ANOVA for happy faces processing task did not reveal a significant group effect in the right or left amygdala.
Discussion
Overall, the groups showed differential activation in the conditions with intensive facial expressions of emotion (rather than mild or neutral). This suggests the relevance of emotion-specific systems rather than general face processing systems.
To summarize our findings, in response to intensively fearful faces the schizophrenia patients treated with conventional depot medication (CONV group) demonstrated increased activation in VMPFC and decreased activation in left amygdala compared with both the HC and RLAI participants. In response to intensively happy faces, CONV patients demonstrated increased activation in VMPFC and reduced activation in right cerebellum compared with both RLAI patients and HC, as well as reduced activation in middle temporal cortex compared with HC. The pattern of activation in middle temporal cortex in the RLAI group showed levels of activation midway between CONV and HC groups.
Importantly, the differential activation between the three groups occurred when the on-line performance on all emotional tasks was not different. The patients in each group (RLAI and CONV) were able to correctly identify the gender of posers depicted in the facial stimuli. We have also tested the possible effect of global functioning on the measures of BOLD response in left amygdala to fearful faces and VMPFC in both happy and fearful conditions. There were no significant correlations between GAF scale score and any of the above measures. This rules out the possible contribution of GAF to the differential BOLD response in amygdala or VMPFC.
The increased VMPFC activation to both fearful and happy emotional faces in the CONV but not in RLAI and HC groups, matched results obtained with the same sample but performing a completely different, i.e. working memory task. In that study (Surguladze et al., 2007), CONV patients demonstrated suboptimal performance in association with an increase of activation in a large cluster in VMPFC.
VMPFC has been characterized as being central to the ‘default mode’ area of the cortex, that tends to de-activate during the performance of cognitive tasks, as cognitive resources are redirected (Raichle et al., 2001; McKiernan et al., 2003). This decrease in ‘default mode’ area activation has been demonstrated not only in cognitive tasks, but also during the implicit processing of emotional pictures (Gusnard et al., 2001).
There have been indications of possible abnormal functioning of ‘default mode’ structures in schizophrenia in response to emotional pictures (Taylor et al., 2002, 2007). A recent review of the brain’s default network (Buckner et al., 2008) has highlighted its relevance to schizophrenia and suggested that overactivity of the default network in schizophrenia may be due to the disruption of the system controlling external attention.
The question arises as to what were the mechanisms that might have ‘protected’ RLAI patients from the abnormal prefrontal cortical activation evident in the CONV group? We suggest that this differential effect of RLAI is related to its facilitation of mesocortical dopaminergic transmission, abnormality of which is the primary source of prefrontal dysfunction in schizophrenia (Davis et al., 1991). The studies exploring the effect of (non-depot) antipsychotics on medial prefrontal cortex have indicated that risperidone but not haloperidol increased an extracellular dopamine levels in rat medial prefrontal cortex (Kuroki et al., 1999). A study of Ichikawa et al. (2001) showed that the synergistic 5-HT(2A) and D2 blockade induced by SGAs (including risperidone) facilitated medial prefrontal dopamine release via stimulation of 5-HT(1A) receptors. We acknowledge that 12 out of 16 patients in CONV group were treated with flupentixol decanoate which also has some 5-HT2A-blocking properties. However, a study directly comparing the receptor occupancy of flupentixol, risperidone and haloperidol (Reimold et al., 2007) demonstrated that the mean 5-HT2A receptor occupancy under flupentixol treatment was 19.5% versus 87.9% in risperidone group, with receptor occupancy under haloperidol being not significantly different from zero. Thus, the 5-HT2A blocking effect of CONV medication on prefrontal cortex functioning could be considered as small, compared with that of RLAI. We suggest that the difference between RLAI and CONV observed on VMPFC activation may be accounted for by RLAI-driven facilitation of dopamine release whereas conventional antipsychotics have a weak effect on this action. Subsequently, the abnormal increase in VMPFC activation observed in the CONV group may represent an over-recruitment of the VMPFC in order to perform the tasks. This over-activation of VMPFC was not specific to either of the emotional tasks: it was observed in both experiments with fearful and happy faces. Moreover, this abnormality seems to emerge even in non-emotional tasks, since we have demonstrated the same pattern in the CONV group performing the working memory experiments (Surguladze et al., 2007). We suggest therefore that this over-recruitment of VMPFC represents a general response to any attention-demanding task, which may be remediated by RLAI.
We found that the HC and RLAI groups had greater activation in the left amygdala compared with the CONV group in response to intensely fearful expressions. There are some possible explanations for this differential response, based on differences in dopaminergic transmission in the striatum and amygdala. Importantly for its antipsychotic effect, the D2 receptor-blocking properties of long-acting risperidone in striatum have been found to be comparable with those of conventional antipsychotics (Remington et al., 2006; Uchida et al., 2008). On the other hand, Carboni et al. (2000) reported that risperidone (similarly to some other SGA) dose-dependently stimulated extracellular dopamine release in the extended amygdala, whereas haloperidol and raclopride were ineffective. It is known that dopamine per se does not cause excitation or inhibition, it is supposed to facilitate the current state in the neural networks, cf. ‘activity-dependent actions of dopamine’ (Abi-Dargham and Moore, 2003). Thus, one could speculate that normalization of amygdala activation observed in our RLAI patients might be associated with this facilitatory dopaminergic effect of RLAI.
The results based on ROI analysis of amygdala activation should be considered with caution, since the amygdala is a small brain area with a high variability in its size and location and our acquisition parameters did not allow for its optimal mapping. Thus, our results on the beneficial effects of RLAI on amygdala activity warrant replication.
What could be the neurobiological basis for differential activation of cerebellum in RLAI and CONV groups? The cerebellum has been implicated not only in motor coordination, but in emotional and attentional tasks (Schmahmann and Sherman, 1998; Allen et al., 2005). In the neuroimaging literature, cerebellar activation has been reported in healthy subjects during perception of emotional signals (Phan et al., 2002; Bermpohl et al., 2006), induction of feelings of sadness, anxiety (Liotti et al., 2000; Reiman et al., 1997) or happiness (Habel et al., 2005b). Paradiso et al. (2003) reported under-activation of cerebellum in unmedicated schizophrenia patients evaluating emotional pictures. Malfunctioning of a cortico–cerebellar–thalamic–cortical circuit, ‘cognitive dysmetria’, has been proposed to underlie the neurobiology of schizophrenia (Andreasen, 1999). There is no agreement among authors, however, with regard to the cerebellar involvement in emotional pathology in schizophrenia (Picard et al., 2008).
Considering the possible mechanisms of action of RLAI on cerebellar functioning, we suggest that it could be accounted for by 5-HT(2A) receptor antagonism by risperidone. Recent research shows significant 5-HT(2A) receptor density in the cerebellum (Eastwood et al., 2001). Importantly for our interpretation, the beneficial effect of SGA (as opposed to FGA) in terms of reversal of the abnormal subcellular distribution of 5-HT(2A) has been demonstrated previously in pyramidal neurons (Willins et al., 1999).
Thus, while findings of the similar cerebellar activation in response to happy faces in the RLAI and HC groups (versus those on conventional FGAs) were unexpected, they appear consistent with the reports of the 5-HT(2A) receptor antagonism effects of risperidone.
Regarding other activations observed in our sample, there were differences in activation of right temporal pole where CONV patients demonstrated lower activation to intensively happy faces compared with HC. RLAI group activation did not differ significantly from either the HC or CONV groups.
The temporal pole is considered as a paralimbic region involved in social and emotional (including facial emotion) processing; for a review see Olson et al. (2007). There is strong evidence for the reduction of grey matter volume (Gur et al., 2000; Kasai et al., 2003) of the temporal pole in schizophrenia. A recent study (Reske et al., 2007) found under-activation bilaterally in temporal poles of patients during the happy mood induction paradigm employing facial expressions as cues. This under-activation was not amenable to psychopharmacotherapy with low doses of either haloperidol or risperidone (although the sample size in the treatment groups was small, five in each). Our finding of deficient activation in the middle temporal cortex in the CONV group is in line with these results. However the activation in the RLAI group did not differ from either CONV or HC groups which precludes us from inferring a beneficial effect of RLAI on temporal cortex functioning.
Limitations
The assignment to CONV or RLAI was not random and was based on a clinician’s judgement. This prevents us reaching strong conclusions regarding the neurobiological mechanisms of BOLD response differences associated with one group of medication versus another. We considered it inappropriate to risk either denying atypical depot treatment to patients who might benefit from it, or disturbing patients satisfactorily managed on conventional depot medication, by a randomized design. Thus, although the results of differential activation in groups are significant, we consider them as preliminary. These results, however, may pave the way for further studies where randomized controlled designs could be implemented. Another problem, indirectly related to the naturalistic (rather than randomized) design is that we could not control for the potential effect of the depot medication preceding the RLAI. Only two of the patients were treated with haloperidol decanoate before commencing on RLAI although it has been shown (Nyberg et al., 1997) that D2 receptor occupancy may persist for four months after discontinuation of that agent. Hence, we argue that the effect of a slow elimination of haloperidol in our CONV group patients would have been minimal. On the other hand, the haloperidol-related delayed elimination of the D2 occupancy in striatal areas would not affect our results, since they were demonstrated in other brain structures than striatum, i.e. MPFC, amygdala, cerebellum and temporal pole.
The task we employed has a good track record as it has been successfully used in a number of studies of emotion processing in various psychiatric disorders. However, we acknowledge that the black–white photographs lack the ecological validity and new paradigms, possibly employing dynamic emotional expressions would be useful. We are currently testing a task with short emotional clips on healthy volunteers.
We did not specifically address the issue of gender-related differences in emotion processing across our participants. We emphasize that the groups did not differ in terms of gender distribution. Theoretically, one cannot rule out the effect of gender × emotion × diagnostic group. Unfortunately, our sample size would not allow for this to be tested adequately. Finally, although we did our best to minimize the potentially confounding effects of a range of clinical variables we could not completely exclude the possibility of, for example, previous substance dependence in some participants.
A practical outcome of this study is that it adds to the neurobiology of evidence on improvement of facial emotion processing associated with risperidone treatment of schizophrenia (Kee et al., 1998; Fakra et al., 2007; Williams et al., 2003) which helps clinicians reach a more informed choice when considering the treatment regimens.
Footnotes
Acknowledgments
The work was supported by an investigator-initiated grant from Janssen Cilag to ASD; the company had no input into this report. ASD acknowledges financial support from the Department of Health via the National Institute for Health Research (NIHR) Specialist Biomedical Research Centre for Mental Health award to South London and Maudsley NHS Foundation Trust (SLaM) and the Institute of Psychiatry at King’s College London. CMcD received support from a Medical Research Council (UK) Pathfinder Award. The authors have no conflict of interest to disclose. We thank Dr V Giampietro for his helpful comments on the manuscript.