
Abstract
We have analysed the long-term effects of adolescent (postnatal day 28–43) exposure of male and female rats to nicotine (NIC, 1.4 mg/kg/day) and/or the cannabinoid agonist CP 55,940 (CP, 0.4 mg/kg/day) on the following parameters measured in the adulthood: (1) the memory ability evaluated in the object location task (OL) and in the novel object test (NOT); (2) the anxiety-like behaviour in the elevated plus maze; and (3) nicotinic and CB1 cannabinoid receptors in cingulated cortex and hippocampus. In the OL, all pharmacological treatments induced significant decreases in the DI of females, whereas no significant effects were found among males. In the NOT, NIC-treated females showed a significantly reduced DI, whereas the effect of the cannabinoid agonist (a decrease in the DI) was only significant in males. The anxiety-related behaviour was not changed by any drug. Both, nicotine and cannabinoid treatments induced a long-lasting increase in CB1 receptor activity (CP-stimulated GTPγS binding) in male rats, and the nicotine treatment also induced a decrease in nicotinic receptor density in the prefrontal cortex of females. The results show gender-dependent harmful effects of both drugs and long-lasting changes in CB1 and nicotinic receptors.
Introduction
Nicotine and cannabis share some pharmacological effects, including modification of brain cognitive function, and are frequently consumed in combination, particularly among adolescents and young adults. Learning and memory impairments are among the most commonly reported behavioural effects of cannabinoids in both human and experimental studies (Ameri, 1999; Nordentoft and Hjorthøj, 2007; Solowij et al., 2002; Solowij and Michie, 2007; Solowij and Battisti, 2008; Trezza et al., 2008). Acute consumption of marijuana is associated with disrupted functioning in a variety of cognitive and performance paradigms, including impaired memory, altered time sense and decrements in tasks such as reaction time, learning perception, motor coordination and attention (Ameri, 1999; Lichtman et al., 2002; Sullivan, 2000). In particular, cannabinoids have been shown to disrupt spatial working memory in a wide range of animal models (Egerton et al., 2006). Cannabinoid CB1 receptors are located in brain regions involved in cognitive processes, notably they are highly expressed in the hippocampus (Herkenham et al., 1990), a brain region that serves a critical role in declarative memory in humans and in processing contextual information (spatial/contextual learning) in animals (Eichenbaum, 2004). In fact, behavioural studies in humans and animals indicate that there is a connection between cannabinoid-mediated memory deficits and impairment of hippocampal function (Sullivan, 2000). More recently, given the key role of the prefrontal cortex (PFC) in diverse aspects of cognitive functions, special attention has been paid to this brain structure where there are also abundant CB1 receptors. In fact, there is evidence suggesting that cannabinoids may alter PFC function (Egerton et al., 2006).
In addition to acute marijuana effects, chronic marijuana/cannabis smoking may cause persistent memory and cognition deficiencies (Solowij et al., 2002). In relation with these findings, chronic exposure to Δ9-tetrahydrocannabinol (THC) or marijuana extracts persistently alters the structure and function of the rat hippocampus (Ameri, 1999) and neuroimaging studies have shown even after an extended washout period, heavy marijuana use results in alteration in the patterns of cortical activation (Lundqvist, 2005). As for the effects of nicotine in cognitive function, animal studies, provide evidence both for positive effects of nicotine and for negative withdrawal effects. In the five-choice serial reaction test of attention in food-restricted rats, it is possible to see improved performance following nicotine administration and impaired performance following nicotine withdrawal (Levin et al., 2006; Shoaib and Bizarro, 2005; Stolerman et al., 2000). The negative effects of withdrawal also appear in humans since a study designed to examine the effect of cigarette smoking and withdrawal showed a deficit in working memory associated with acute abstinence from smoking (Mendrek et al., 2006). These cognitive effects may contribute to the difficulty of smoking cessation.
In spite of the above evidence, and the high incidence of nicotine and cannabis abuse among adolescents, relatively few experimental studies have been devoted to analyse the long-term cognitive effects of these drugs when they are administered alone and in combination in young animals (Marco et al., 2006, 2007). Adolescence is a critical developmental period that involves neurocognitive, hormonal and psychosocial changes that mark the transition from childhood to adulthood. During this developmental phase, maturation and rearrangement of major neurotransmitter pathways (notably the endocannabinoid system (Rodriguez de Fonseca et al., 1993; Wenger et al., 2002)) and functions are still taking place and, therefore, adolescents are particularly vulnerable to the consequences of psychoactive drugs exposure (Adriani and Laviola, 2004; Crews et al., 2007; Spear, 2000). Adolescent daily tobacco smokers experienced acute impairment of verbal memory and working memory after smoking cessation, along with chronic decrements in cognitive performance. These findings have been related to the fact that at early developmental periods neurotoxic effects of nicotine might be enhanced (Jacobsen et al., 2005). Another study by Jacobsen et al. (2007) in adolescents has shown greater disruption in brain function and verbal memory during nicotine withdrawal in those who were also regular cannabis users. A recent study on the effects of simultaneous cannabis and tobacco use in young women indicates that simultaneous use is potentially a marker for more severe psychosocial consequences associated with cannabis use, suggesting that simultaneous users of these drugs are an especially vulnerable population (Agrawal et al., 2009).
The present study was undertaken with two main objectives. First, we aimed to address possible long-term effects of sub-chronic exposure to the cannabinoid agonist CP 55,940, nicotine or both drugs during the periadolescent period on diverse tasks related to the cognitive (memory) function as well as on the anxiety level of the animals. Our second objective was to investigate the long-term effects of the drugs on CB1 cannabinoid receptor and nicotinic acetylcholine receptor (nAChR) densities measured by quantitative autoradiographic binding studies with [3H]CP 55,940 and L-[3H]nicotine, respectively, in several brain areas. In the same brain areas, we also investigated CB1 receptor function by CP 55,940-stimulated [35S]GTPγS binding autoradiography.
Materials and methods
Animals
Wistar albino rats (Rattus norvegicus albinos) of both genders were used in this study. Subjects were the offspring of rats purchased from Harlan Interfauna Ibérica S.A. (Barcelona, Spain), which were mated (one male × two females) in our laboratory approximately 2 weeks after their arrival. All animals were maintained at a constant temperature (22 ± 1°C) and humidity (50 ± 1%) in a reverse 12 h dark/light cycle (lights on at 20:00 h), with free access to food (commercial diet for rodents A04/A03; Panlab, Barcelona, Spain) and water.
On the day of birth (postnatal day (PND) 0) litters were culled and sex balanced to eight pups per dam (four males and four females). At weaning day (PND 22), animals were separated by gender and housed in different cages with four individuals per cage. A total of 10 litters were used, with experimental groups having animals from two or three different litters. All experimental procedures and animal care was in compliance with the European Union Guidelines on Laboratory Animal Care (Directive 86/609/EEC).
Pharmacological treatments
All treatments were administered daily from PND 28 to PND 43. (−)-Nicotine hydrogen tartrate salt (NIC; Sigma) was dissolved in saline and the pH was adjusted to 7–7.4 with NaOH. Nicotine, at a dose of 0.7 mg/kg prepared as free base, or its corresponding vehicle (saline) was administered s.c. twice a day (11:00 h and 17:00 h) at a volume of 2 ml/kg. The synthetic cannabinoid receptor agonist CP 55,940 (0.4 mg/kg, 2 ml/kg; Tocris, Spain) or its corresponding vehicle (ethanol : cremophor : saline 1 : 1 : 18, cremophor; Fluka BioChemiKa) was administered i.p. once per day (17.00 h) before the second nicotine injection. All animals received three injections and were randomly assigned to the following groups: NIC males (n = 8), NIC females (n = 8), CP 55,940 (CP) males (n = 12), CP females (n = 12), NIC-CP males (n = 12), NIC-CP females (n = 12), control (CONT) males (n = 7) and CONT females (n = 8). Animals from the same cage received the same treatment.
Experimental design
By the present animal model and experimental design (Figure 1), we aimed to address the long-term (withdrawal) effects of subchronic adolescent exposure to NIC and/or CP on diverse memory tasks. Thus, pharmacological treatments were administered during the interval PND 28–43 and we evaluated the effects on spatial (object location test, OL) and recognition memory (novel object test, NOT) after approximately 1 month of withdrawal (PND 70–76). In order to evaluate whether the effects on these memory tests could be affected by a potential change in the anxiety level of the animals, we also performed the elevated plus maze (EPM) of anxiety. In addition we evaluated the short term effects of the drugs in the spontaneous alternation task (SAT), which was performed at PND 44. At 81 days, animals were sacrificed and their brains were removed for nicotinic and CB1 receptor assays (see below for details).
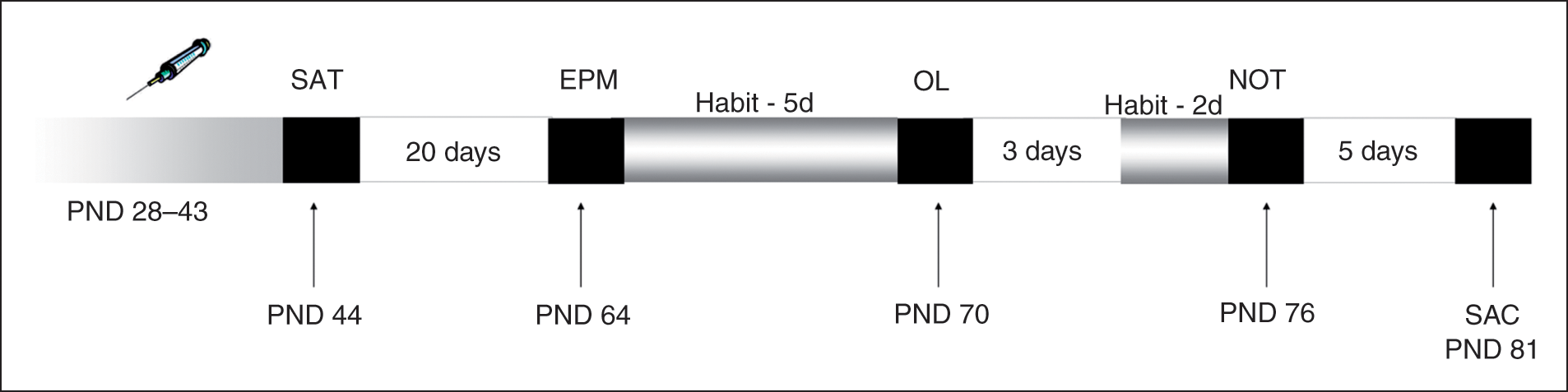
Experimental design. Animals were exposed to nicotine (NIC) and/or CP 55,940 (CP) during the periadolescent period (PND 28–43). At PND 44, short-term memory was assessed in the spontaneous alternation task (SAT). At PND 64 all rats were subjected to the elevated plus maze test of anxiety (EPM). At PND 70, rats performed the object location task (OL) which measures the recognition memory for object location (spatial memory). At PND 76, the animals were tested in the novel object test (NOT) where the recognition memory of novel versus familiar object is evaluated. Previous to each one of these two latter tests, the animals were allowed to habituate (Habit) to the apparatus. At 81 days, rats were sacrificed (SAC) and their brains were removed for nicotinic and CB1 receptor assays.
Behavioural tests
Spontaneous alternation task
At PND 44, short-term memory was assessed by spontaneous alternation behaviour in the Y-maze task. Free-running or continuous spontaneous alternation task (SAT) was performed in a symmetrical black Y-maze (40 cm × 30 cm × 15 cm) made of opaque PVC. Each rat had free access to the three arms of the maze for 8 min. During this time sequences of arm entries were recorded. Analyses were performed on (i) the total number of arm entries, (ii) the sequence of arm entries. An arm entry was recorded when the hind paws of the rat were within the arm. Alternation was defined as entries into all three arms on consecutive occasions. Then, the discrimination index is calculated by dividing the number of alternations by the number of total arm entries minus one.
Elevated plus maze
Twenty days after the SAT (PND 64). All rats were subjected to the elevated plus maze (EPM) test. The animals were habituated in a quiet laboratory for a 30-min period before testing. The apparatus consisted of two open arms (50 cm × 10 cm) and two enclosed arms of the same size with 40-cm-high walls arranged so that the arms of the same type were opposite each other. The junction of the four arms formed a central square area (10 cm × 10 cm). The apparatus was made of hard plastic material and elevated to a height of 62 cm. The test was carried out for 5 min. The measures recorded were frequency and duration of arm visits, separately for open and closed arms. An arm was considered to be entered when the animal entered it with the four limbs. We calculated the open arms entries as a percentage of the total number of entries and the time spent in the open arms as a percentage of the total time in arms. In this test, the percentage of time and entries in the open arms of the maze are the two parameters related to anxiety, whereas the number of closed arms entries provides the best measure of motor activity (File, 1992; Pellow et al., 1985).
Object location task
At PND 70, rats performed the object location task (OL), which measured the recognition memory for object location (spatial memory). The protocol was based on Ennaceur et al. (2005) with some modifications that were introduced according to pilot studies performed under our specific experimental conditions. The apparatus consisted of a square open box (60 cm × 60 cm × 45 cm) with matte-painted metallic walls and a plastic-covered wooden floor. The task included three phases: (i) Habituation (from PND 65 to PND 69, five consecutive days): during this phase, the animals were allowed to explore the apparatus during 5 min each day; (ii) Training session: At PND 70 rats were exposed to two identical objects (transparent glass bottles) and they were allowed to explore both of them during 3 min; and (iii) Test session: At PND 70, 10 min after the training session, the animal was exposed to the same two objects but one of them was moved to a new location in the arena. All rats were placed in an individual cage located in their home room during the inter-trial interval. In all sessions, objects were placed in the back corner of the arena at a distance of 5 cm from the wall. The diverse phases were recorded by a video camera and then measured manually by the experimenter.
Analyses were performed on (i) the total time that the animal spent exploring the two sample objects in the training session, (ii) latency to explore any object, (iii) discrimination index (DI), which is the difference between the time that animals explore the displaced object and the familiar one divided by the total time spent exploring the two objects in the test session, and iv) the total time spent exploring the two sample objects during the test. Exploration of an object was defined as directing the nose to the object at ≤1 cm of distance, and/or touching the object with the nose. However, turning around, climbing or biting the objects were not considerer as exploratory behaviour (Barker et al., 2007; Dix and Aggleton, 1999; Mitchell and Laiacona, 1998; Rossato et al., 2007). Objects were carefully cleaned between rats with 20% ethanol in water solution and their position was randomly changed between the two opposite corners of the arena during sessions to avoid object and/or place preference. Animals exploring less than 10 s during the training phase or that only explored one of the objects during the test phase were filtered for statistical analysis.
Novel object test
Five days after the OL, the animals were tested in the novel object test (NOT), based on the differential exploration of familiar versus new objects. The present protocol was based on Ennaceur and Delacour (1988), with some modifications that were introduced according to pilot studies performed under our specific experimental conditions. The apparatus was the same as the one employed in the OL described above.
The test had three phases:
Habituation (from PND 74 to PND 75): during this phase the animals were allowed to explore the apparatus during 5 min per day.
Training session (PND 76): after the last day of habituation, rats were exposed to two identical objects (that were different from the ones used in the OL; in this case tow plastic boxes, named as F1 and F2) until they explored the objects during 30 s. The maximum duration of this training session was of 4 min.
Test session (PND 76): 4 h after the training session, rats were exposed to two objects for 3 min. One of them was familiar for the animal (F1 or F2) and the other one was a non-familiar (new) object (metallic coloured box, N). During the inter-trial interval animals were returned to their home cage with their litter mates.
In all sessions, objects were placed in the back corner of the arena at a distance of 5 cm from the wall. Objects were carefully cleaned between rats with 20% ethanol in water solution, and their position was randomly changed between the two opposite corners of the arena, during sessions, to avoid object and/or place preference.
Analyses were performed on (i) the total time that the animal spent exploring the two sample objects in the training session (F1 + F2), and (ii) DI, which is the difference between the time exploring the new object and the time exploring the familiar one, divided by the total time spent exploring the two objects in the test session (N − F/N + F). Animals exploring less than 30 s during the training phase or that only explored one of the objects during the test phase were filtered for statistical analysis.
Each test was started by placing the animals in the area of the apparatus considered more behaviourally neutral (at the centre of the apparatus facing one of the enclosed arms (plus maze and SAT), facing the opposite wall with respect to the objects location (NOT and OL)) so that the animal was not artificially induced to perform a significant pattern. Both apparatuses were thoroughly cleaned after each animal finished the test with 20% ethanol diluted in water.
Body weight and food intake control
Body weight (BW) of rats was daily controlled at 09:00 h throughout the experimental period, from PND 25 to PND 63. To minimize confounding effects coming from initial differences on body weight data were expressed as the percentage of body weight gain [percentage BW gain = (BWday – BW25)/BW25 × 100] taking BW at postnatal day 25 as the reference day.
Food intake was daily registered at 09:00 h as previously described (Biscaia et al., 2003; Lamota et al., 2008). An established quantity of pelleted food (250 g) was placed on the top of a cage every morning, and 24 h later the remaining food was weighed again. The difference between weights was considered as the daily food intake of the four animals housed in the same cage. Data are expressed as g/cage.
Binding studies
Drugs
[3H](–)-CP 55,940(1α,2β-(R)-5α]-5-(1,1-dimethyleptyl)-2-[5-hydroxy-2-(3-hydroxy-propyl)cyclohexyl]phenol), L-[3H]nicotine, 5′-O-(3-[35S]thiotriphospate) ([35S]GTPγS), were purchased from Perkin-Elmer Life Sciences (Boston, MA, USA). Guanosine 5′-diphosphate (GDP) GTPγS, (–)-nicotine hydrogen tartrate sodium and (–)-CP 55,940 were obtained from Sigma/RBI (St. Louis, MO, USA) and Tocris (Ballwin, MO, USA), respectively.
Autoradiographic studies
Brain tissue preparation
Five days after completion of behavioural testing, rats (PDN 81) were sacrificed by rapid decapitation, and brains were rapidly removed, immediately immersed in liquid nitrogen and then stored at −80°C until sectioning for autoradiographic studies.
[3H](–)-CP 55,940 and L-[3H]nicotine binding autoradiography
Coronal sections 12–16 µm thick were prepared with a cryostat at −20°C, thaw mounted onto Superfrost Plus slides (BDH, Lutterworth, UK) and stored with desiccant at −20°C until use. Brain regions for analysis, according to the atlas of Paxinos and Watson (1997) corresponding to: cingulate cortex areas 3 and 1 (Cg3 and Cg1) (AP + 3.2), CA1, CA2 and CA3 fields of hippocampus, gyrus dentate (DG) of hippocampus, and amygdala (AP −2.14, −3.14), were chosen (Figure 2) and adjacent sections to those used for autoradiography were collected and stained with Neutral Red to facilitate the identification of the selected brain areas.
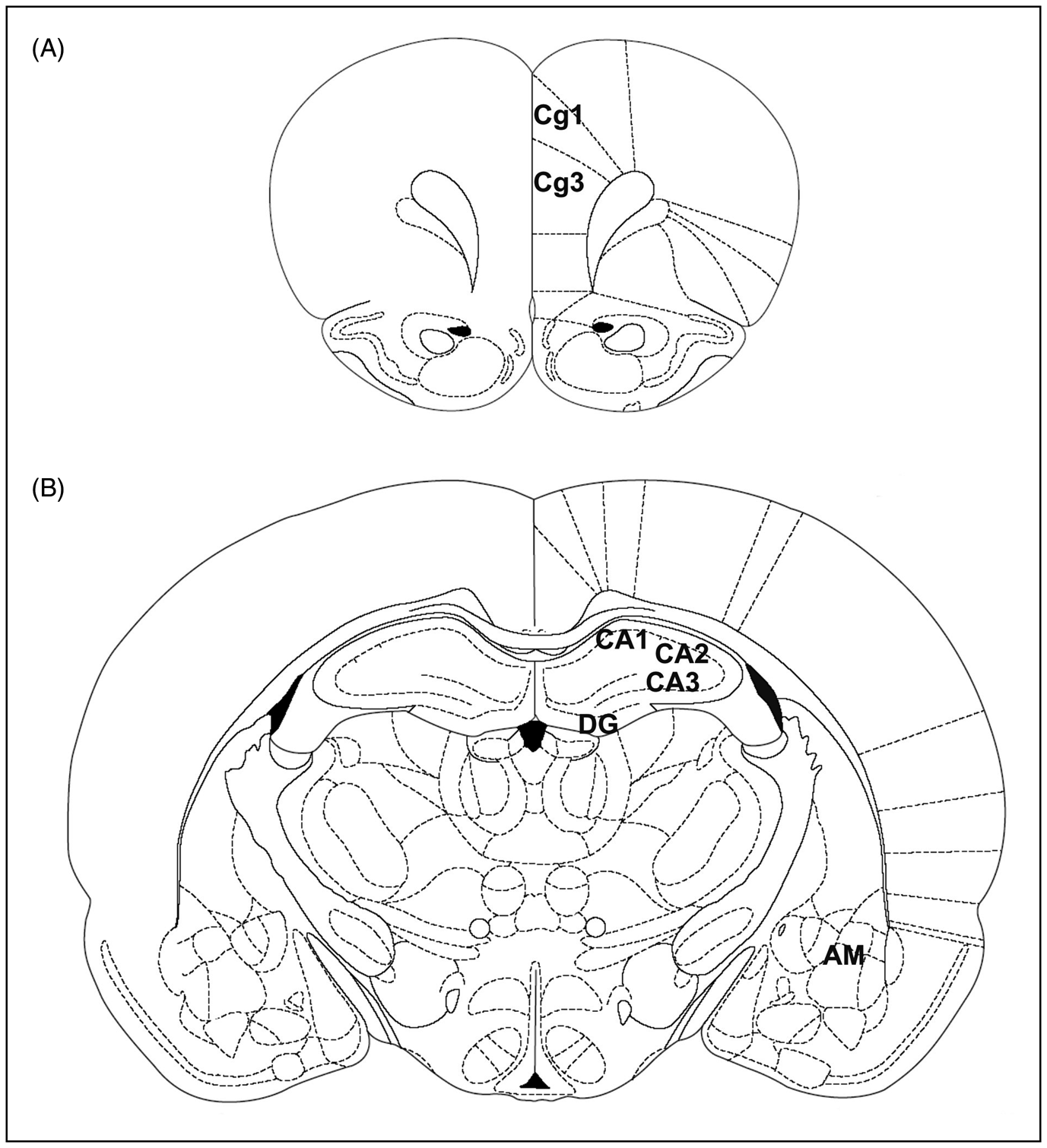
Schematic (adapted from Paxinos G and Watson C (1997) The Rat Brain in Streretaxic Coordinates with permission from Elsevier). of the prefrontal cortex (cingulate cortex areas 3 and 1, Cg3 and Cg1) (a), and hippocampus (b). AM, amygdala; Cg1 and Cg3, cingulate cortex areas 1 and 3; CA1, field CA1 of Ammon’s horn; CA2, field of Ammon’s horn; CA3, field of Ammon’s horn; DG, dentate gyrus of hippocampus.
[3H](–)-CP 55,940 binding autoradiography was performed as previously described by Herkenham et al. (1991). Briefly, tissue slides were incubated at 37°C for 2.5 h in 50 mM Tris-HCl (pH 7.4) containing 5% bovine serum albumin (BSA) and 10 nM of [3H]CP 55,940 (specific activity, 139.6 Ci/mmol, Perkin Elmer, Boston, MA, USA). Non-specific binding was determined in adjacent brain sections in the presence of 10 µM unlabelled CP 55,940. Following incubation, tissue slides were rinsed twice at 4°C for 2 h in ice-cold Tris-HCl buffer (50 mM, pH 7.4) with 1% BSA, once (5 min) with 50 mM Tris-HCl, dipped in ice-cold deionized water and then air dried.
For L-[3H]nicotine-nAChRs autoradiography, as previously described (Mugnaini et al., 2002), sections were preincubated at 4°C in KRH buffer (HEPES 20 mM, NaCl 118 mM, KCl 4.8 mM, CaCl2 2.5 mM, MgSO4 1.2 mM; pH 7.5 with NaOH) for 30 min. Then, sections were incubated in KHR (pH 7.5) containing 20 nM L-[3H]nicotine (specific activity, 75.0 Ci/mmol, Perkin Elmer, Boston, MA, USA) for 90 min at 4°C. Non-specific binding was determined in adjacent brain sections in the presence of 10 µM unlabelled (–)-nicotine hydrogen tartrate salt. Incubation was terminated by two washes (5 s) in KRH buffer and two washes (5 s) in HEPES (pH 7.5), followed by a short dipping in ice-cold deionized water and then tissue slides were air dried.
CP 55,940-stimulated [35S]GTPγS binding in autoradiography
[35S]GTPγS binding in autoradiography was performed as previously described by Rubino et al. (2003). Briefly, tissue slices were preincubated in assay buffer (50 mM Tris-HCl, 3 mM MgCl2, 0.2 mM EGTA, 100 mM NaCl, 0.1% BSA, pH 7.4) at 25°C for 10 min and preincubated further with 3 mM GDP for 10 min in assay buffer at 25°C.
Agonist-stimulated activity was determined by incubating tissue slices for 2 h at 25°C with 5 µM of CP 55,940 in fresh buffer containing 0.04 nM of [35S]GTPγS and 3 mM of GDP. Basal activity was assayed in the absence of CP 55,940 and in the presence of 3 mM GDP. Non-specific binding was determined in adjacent brain sections in the presence of 10 µM of unlabelled GTPγS. Following incubation, tissue slices were rinsed twice in ice-cold Tris-HCl buffer (50 mM, pH 7.4) and once in deionized water, and air-dried.
Image analysis
Dried tissue sections and slide-mounted [3H]micro-scales standards (RPA 501 and 505, Amersham) for [3H](–)-CP 55,940 and L-[3H]nicotine or [14C]micro-scales standards (RPA 504 and 511, Amersham) for [35S]GTPγS binding autoradiography were placed in a Fujifilm BAS cassette with a BAS-5000 imaging plate. The resulting images were analysed with the Fujifilm-BAS 5000 imaging system (AIDA, Raytest, USA), and optical densities were transformed into levels of bound radioactivity (fentomoles per milligram of protein) with gray values generated by co-exposed [3H] or [14C] standards. CP 55,940-stimulated [35S]GTPγS activity in brain sections was calculated by subtracting the optical density in basal sections (incubated with GDP alone) from that of agonist-stimulated sections; results were expressed as percentage basal activity.
Statistical analysis
All results were analysed by three-way ANOVA (factors: gender, nicotine treatment and CP 55,940 treatment), followed by lower-order ANOVAs were appropriate. A repeated measures ANOVA was used for the analyses of body weights. The Tukey HSD test with a level of significance set at p < 0.05 was used for post-hoc comparisons.
Results
Spontaneous alternation task
For the number of arm entries during the spontaneous alternation task, the ANOVA revealed significant effects of nicotine (F(1,67) = 11.76, p = 0.001) and of the cannabinoid agonist (F(1,67) = 22.94, p < 0.01) as well as a significant interaction between the two drugs (F(1,67) = 7.6, p < 0.01). For both, males and females, the NIC group showed a significant higher value for this parameter than their respective control and CP-treated counterparts (Figure 3). As for the DI, the analysis revealed a significant interaction between the cannabinoid agonist (CP) and gender (F(1,67) = 10.04, p < 0.01) and significant interaction between the three main factors (F(1,67) = 6.46, p < 0.05). Post-hoc comparisons showed that spontaneous alternation behaviour was significantly augmented in control females when compared to control males (p < 0.05). When we further analysed males separately, the two-way ANOVA revealed a significant overall effect of CP (F(1,33) = 17.84, p < 0.001). A significant (though modest) increase in the DI of CP and NIC-CP males when compared to control males (Figure 3).
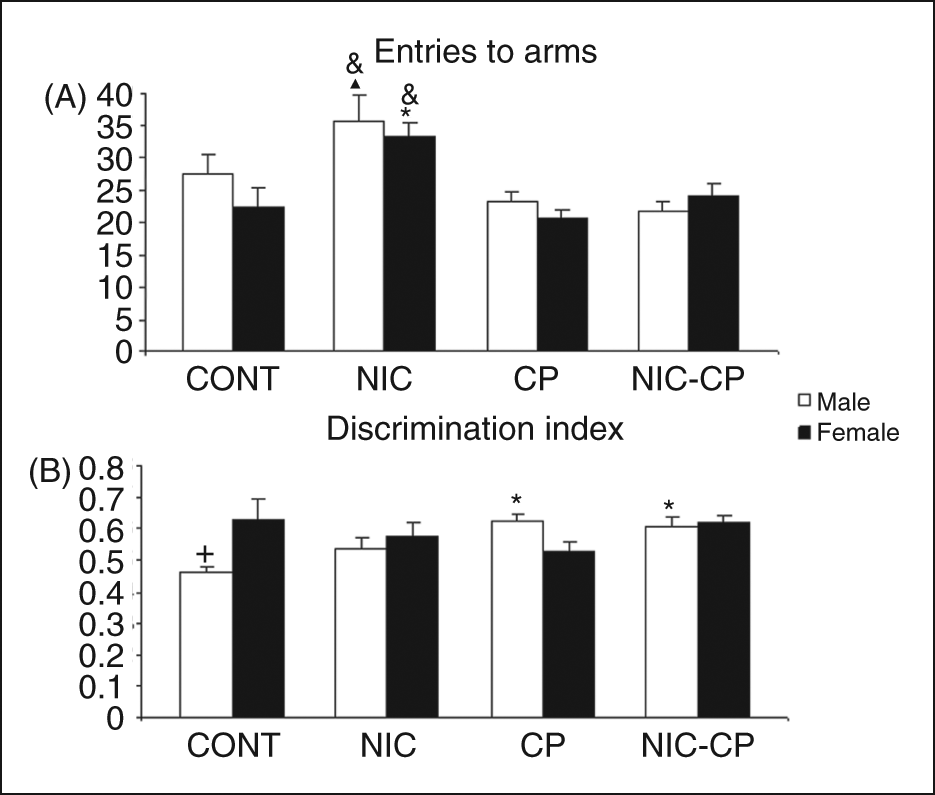
Short-term effects of chronic treatments (PND 28–43) with nicotine (NIC, 1.4 mg/kg) and/or the cannabinoid agonist CP 55,940 (CP, 0.4 mg/kg) on the behaviour of adolescent rats (PND 44) in the spontaneous alternation task. A) Number of arm entries (horizontal activity), B) Discrimination index (index of working memory). Histograms represent the mean ± SEM (6–12 animals per experimental group). Statistically significant differences: *, vs. control (CONT) animals of the same gender; &, vs. CP-treated animals of the same gender; ▴, vs. NIC-CP males; +, vs. females of the same treatment group.
In brief, in the spontaneous alternation task, control females showed a better performance than control males (significantly increased DI), and males administered with the chronic cannabinoid treatment showed a modest though significant increase in the DI when compared with their control group.
Elevated plus maze
The analysis of the parameters measured in the elevated plus maze test (Table 1) rendered significant effects of gender for the percentage of time (F(1,71) = 13.312, p < 0.001) and entries (F(1,71) = 14.815, p < 0.001) in the open arms, with females showing increased percentages than males, and a general effect of nicotine on the closed arm entries of females (F(1,36) = 6.179, p < 0.05). The parameters related to anxiety-related behaviour, i.e., the percentages of time and entries in the open arms, were not changed by any drug.
Long-term effects of adolescent exposure to chronic treatments with nicotine and/or the cannabinoid agonist CP 55,940 on the behaviour of adult rats in the elevated plus maze
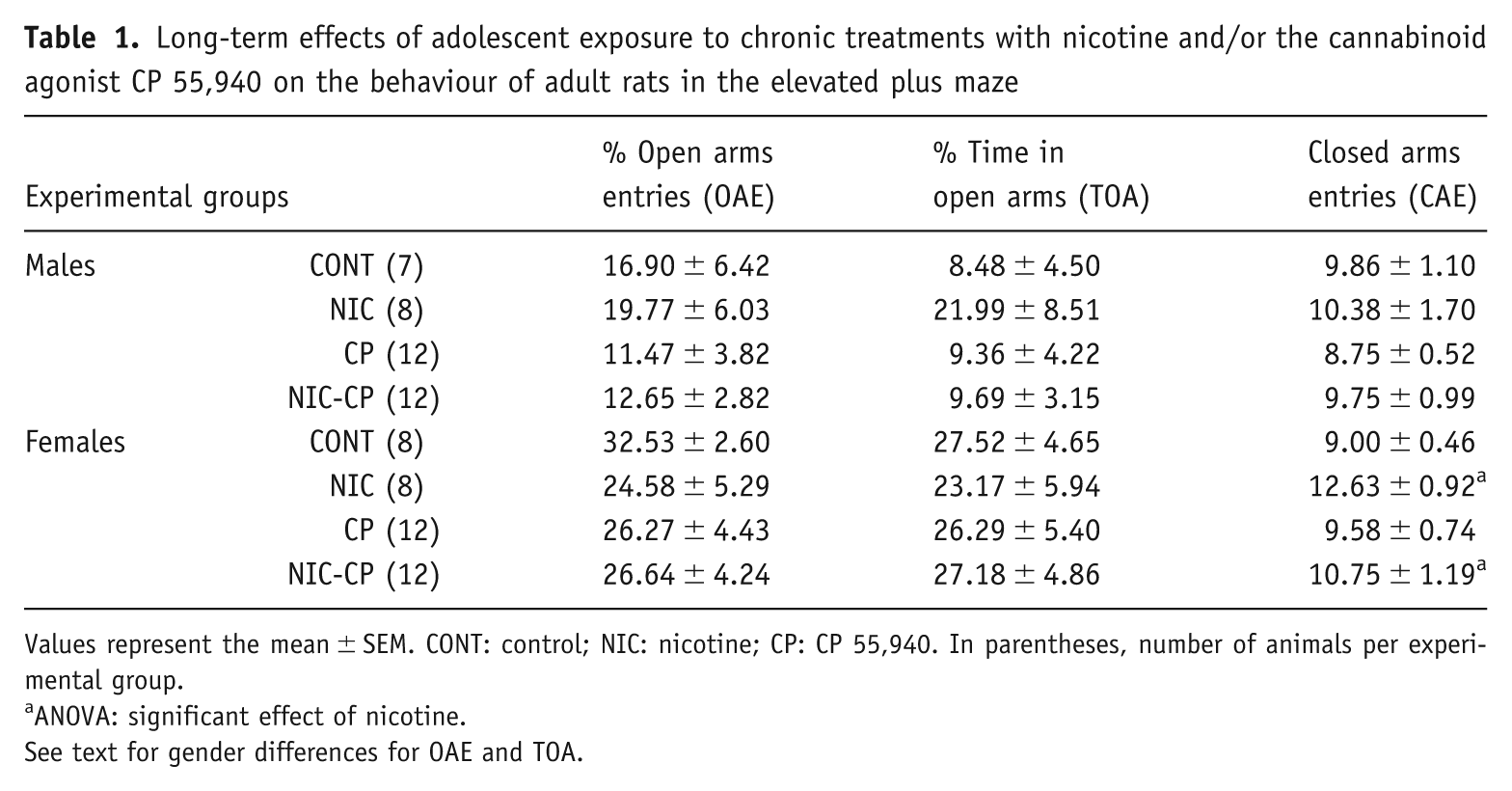
Values represent the mean ± SEM. CONT: control; NIC: nicotine; CP: CP 55,940. In parentheses, number of animals per experimental group.
ANOVA: significant effect of nicotine.
See text for gender differences for OAE and TOA.
Object location task
Among the animals included in the statistical analysis, there were no differences in the total time that they spent exploring the objects during the training phase, which indicates that differences in memory (DI) cannot be attributed to a higher level of interaction with objects during the training phase. Moreover, the analysis of the latency to explore the object did not render any significant difference between groups (Table 2). It is worth noting however, that five males and four females that had been exposed to the cannabinoid agonist in the adolescence were removed from the analysis since they did not explore at all the objects during the training phase, suggesting a markedly enhanced neophobic reaction. As for the DI (Figure 4), significant interactions between NIC and CP (F(1,56) = 10.79, p < 0.01) as well as between NIC, CP and gender (F(1,56) = 9.94, p < 0.01) were found. Post-hoc comparisons rendered significant differences only within females, so that the DI of three treatment groups (NIC, CP and NIC-CP) were significantly lower than the DI corresponding to the control group.
Long-term effects of adolescent exposure to chronic treatments with nicotine and/or the cannabinoid agonist CP 55,940 on the behaviour of adult rats in the object location test
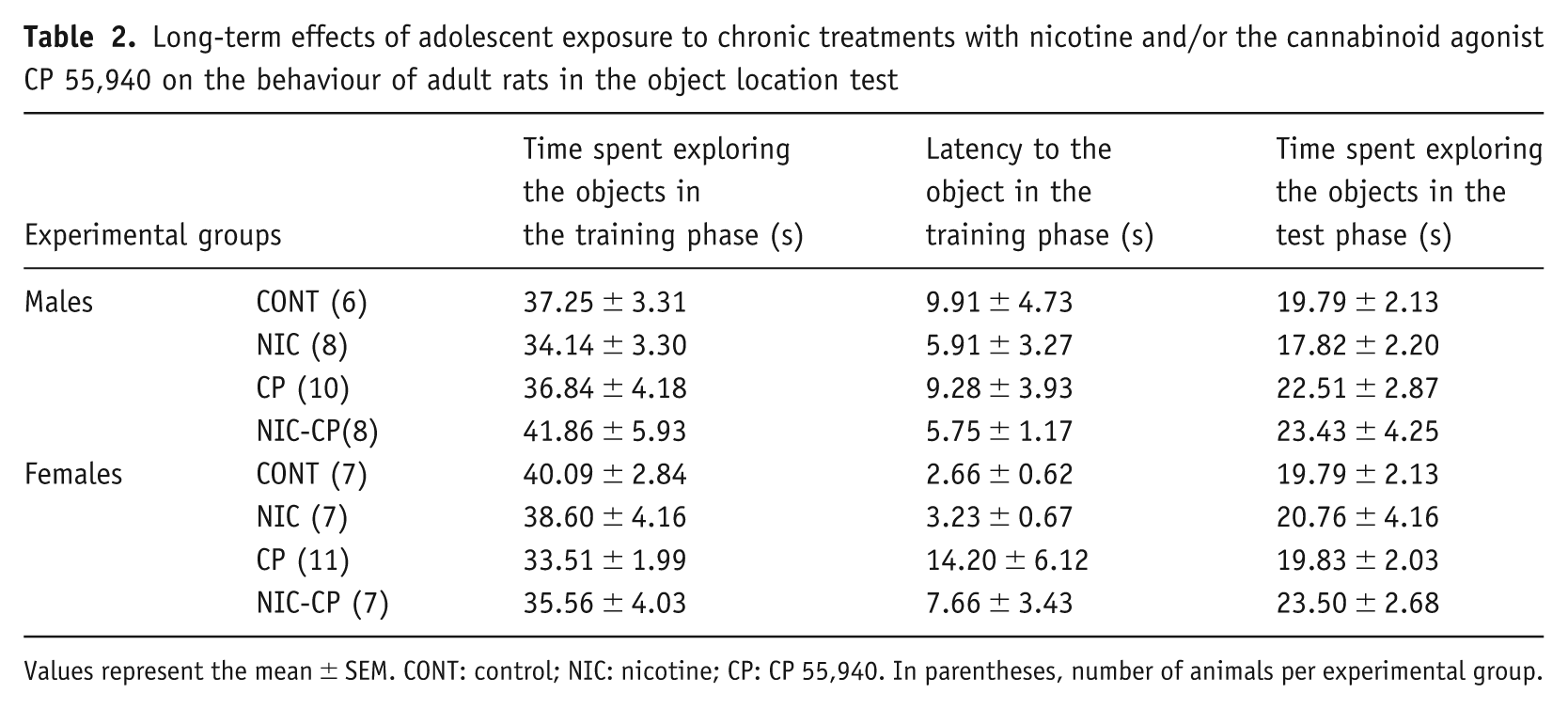
Values represent the mean ± SEM. CONT: control; NIC: nicotine; CP: CP 55,940. In parentheses, number of animals per experimental group.
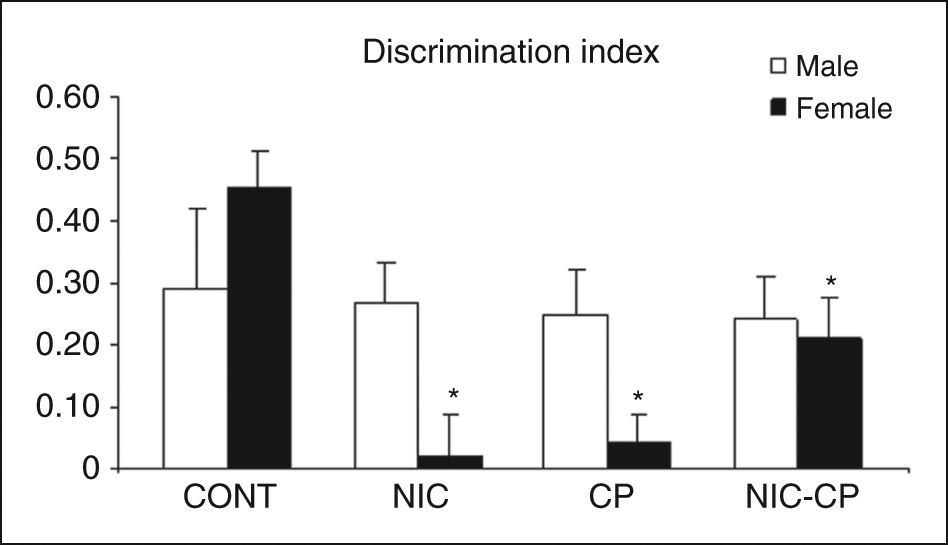
Long-term effects of chronic treatments (PND 28–43) with nicotine (NIC, 1.4 mg/kg) and/or the cannabinoid agonist CP 55,940 (CP, 0.4 mg/kg) on the behaviour of adult rats (PND 70) in the object location task. Histograms represent the mean ± SEM (6–11 animals per experimental group). Statistically significant differences: *, vs. control (CONT) females.
In summary, in the object location task all pharmacological treatments induced significant decreases in the DI of females, whereas no significant effects were found among males.
Novel object test
During the training phase (Table 3), the animals that had been exposed to the cannabinoid agonist (CP) during the periadolescent period, needed to stay more time in the arena to complete the required 30 s of object exploration (F(1,56) = 8.00, p < 0.01). Moreover, as in the OL, four males and three females with an adolescent history of CP were removed from the analysis, since they did not explore the objects during the training phase, supporting their markedly enhanced neophobic reaction. This observation may be related with a neophobic response induced by the drug. As for the DI (Figure 5), the three-way ANOVA showed significant effects of nicotine (F(1,56) = 5.23, p < 0.05) and CP (F(1,56) = 12.82, p = 0.01) as well as significant interactions NIC × CP (F(1,56) = 6.44, p < 0.05) and CP × gender (F(1,56) = 13.27, p < 0.05). Post-hoc comparisons revealed that, in males, the memory index was significantly lower in CP and NIC-CP groups when compared to their control group. Among females, NIC female rats showed a significantly decreased DI when compared to the corresponding female control group. The post-hoc comparison also showed a trend towards signification between male and female NIC (p = 0.065).
Long-term effects of adolescent exposure to chronic treatments with nicotine and/or the cannabinoid agonist CP 55,940 on the behaviour of adult rats in the novel object test
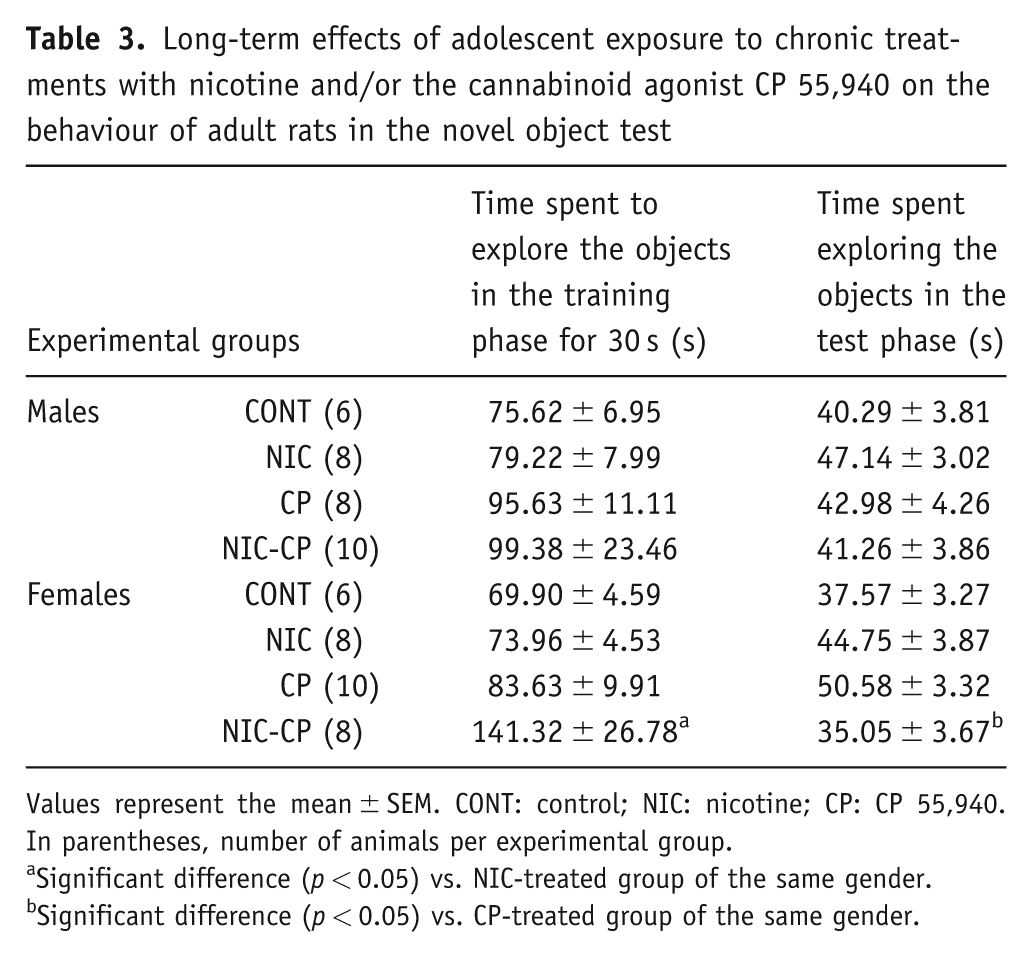
Values represent the mean ± SEM. CONT: control; NIC: nicotine; CP: CP 55,940. In parentheses, number of animals per experimental group.
Significant difference (p < 0.05) vs. NIC-treated group of the same gender.
Significant difference (p < 0.05) vs. CP-treated group of the same gender.
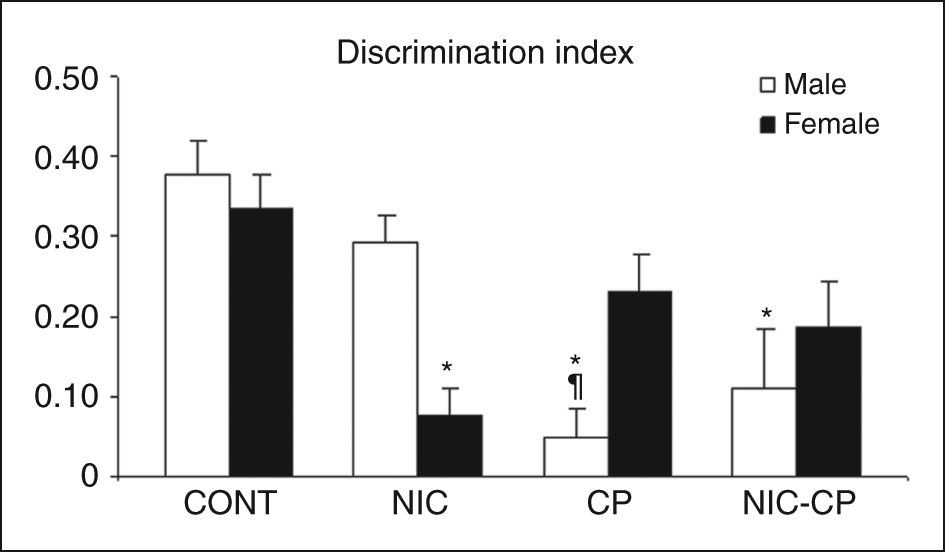
Long-term effects of chronic treatments (PND 28–43) with nicotine (NIC, 1.4 mg/kg) and/or the cannabinoid agonist CP 55,940 (CP, 0.4 mg/kg) on the behaviour of adult rats (PND 76) in the novel object test. Histograms represent the mean ± SEM (6–10 animals per experimental group). Statistically significant differences: *, vs. control (CONT) animals of the same gender; ¶, vs. NIC males.
To sum up, in the novel object test, NIC-treated females showed a significantly reduced DI, whereas the effect of the cannabinoid agonist (a decrease in the DI) was only significant in males.
Body weight
The repeated measures ANOVA rendered significant global effects of gender (F(1,71) = 203.913, p < 0.01) and of the cannabinoid agonist (CP) treatment (F(1,71) = 58.727, p < 0.01) on body weights (Figure 6). Further, we analysed by additional three-way ANOVA the specific effects throughout the treatment and post-treatment periods. At PND 28, the effect of gender was already significant (F(1,71) = 8.228, p < 0.01) and this effect lasted until the last day of the second period, PND 63, i.e., throughout the whole experimental period, (F(1,71) = 568.964, p < 0.01), with males showing significant higher body weights than females, as expected. At PND 29, the effect of CP was already significant (F(1,71) = 4.656, p < 0.05). Post-hoc comparisons showed that these differences affected the male group and it was at PND 32 when the effect was more marked (F(1,71) = 60.499, p < 0.01). In females, the first difference between control and CP animals appeared at PND 33 (F(1,71) = 126.606, p < 0.01). The general effect of CP was significant throughout the treatment period and lasted until PND 47 in males (F(1,71) = 51.404, p < 0.01) and until PND 49 in females (F(1,71) = 26.561, p < 0.01). Moreover, during the post-treatment period, post-hoc comparisons showed that, in females, the group treated with CP (but not nicotine) showed a significantly lower weight gain when compared to the respective control group until PND 63.
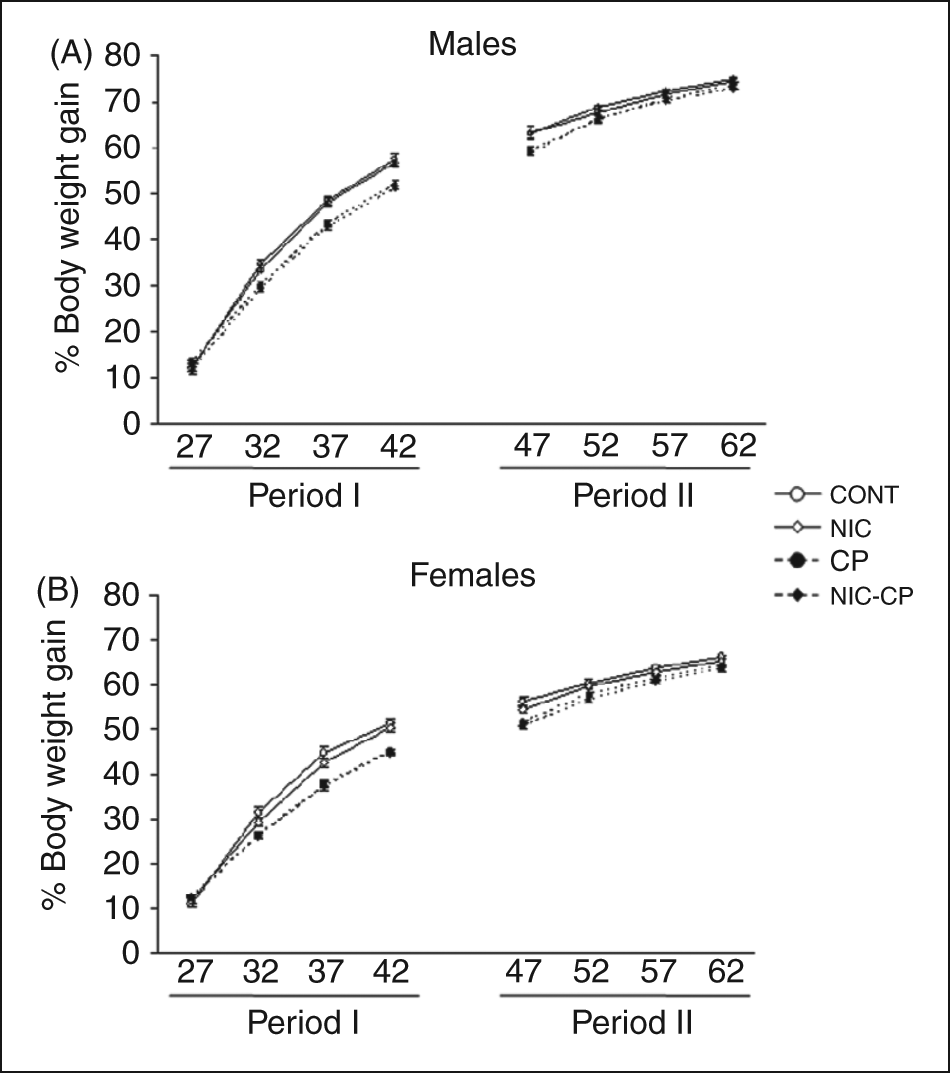
Effects of chronic treatments (PND 28–43) with nicotine (NIC, 1.4 mg/kg) and/or the cannabinoid agonist CP 55,940 (CP, 0.4 mg/kg) on body weight gain. Period I: during treatment; period II: after treatment. ANOVA: significant overall effect of CP (see text for details).
In summary, the results showed that the cannabinoid agonist induced a decrease in body weights that lasted beyond the treatment period and that normal weights were recovered before the adulthood.
CB1 receptor binding and function
Basal levels of [35S]GTPγS binding do not significantly differ between brains from vehicle and drug-treated male or female rats in any region analysed (data not shown). Table 4 shows the densitometric analysis of CP 55,940-stimulated GTPγS binding in selected brain areas of adult male and female rats exposed during the adolescence to different drug treatments. The three-way ANOVA showed significant interactions between cannnabinoid agonist treatment and gender in hippocampal areas CA1 (F(1,43) = 5.96, p = 0.019), CA2 (F(1,41) = 9.95, p = 0.003) and CA3 (F(1,42) = 4.94, p = 0.032). The Tukey post-hoc analysis revealed a significant effect of gender in vehicle-treated rats, so that the CP 55,940-stimulated binding was significantly higher in females than in males in CA2. Indeed, CP 55,940-stimulated GTPγS binding was also higher in females than in males in CA1 and CA3, though the differences did not reach statistical significance. (p = 0.14 and 0.09 for CA1 and CA3, respectively).
Long-term effects of adolescent exposure to chronic treatments with nicotine and/or the cannabinoid agonist CP 55,940 (CP) on CP-stimulated [35S]GTPγS binding in several brain areas of adult rats
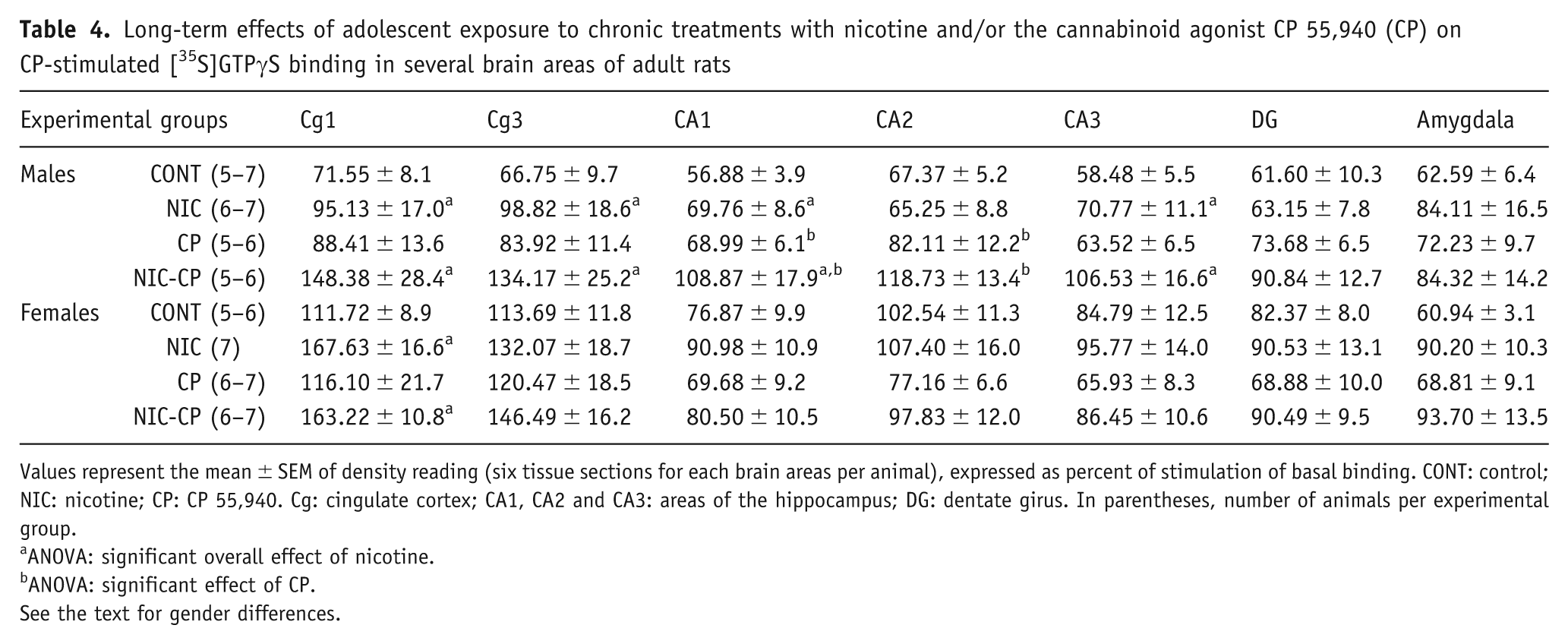
Values represent the mean ± SEM of density reading (six tissue sections for each brain areas per animal), expressed as percent of stimulation of basal binding. CONT: control; NIC: nicotine; CP: CP 55,940. Cg: cingulate cortex; CA1, CA2 and CA3: areas of the hippocampus; DG: dentate girus. In parentheses, number of animals per experimental group.
ANOVA: significant overall effect of nicotine.
ANOVA: significant effect of CP.
See the text for gender differences.
The ANOVA also rendered significant overall effect of nicotine treatment and gender in several brain areas. Specifically, the effect of nicotine treatment was significant in Cg1 (F(1,41) = 14.65, p < 0.001), and in Cg3 (F(1,44) = 6.94, p = 0.012), as well as in hippocampal areas CA1 and CA3 (F(1,43) = 7.58, p = 0.009; F(1,42) = 7.83, p = 0.008, respectively), showing that nicotine induces an increase of stimulated GTPγS binding in these brain regions. The effect of gender was significant in Cg1 (F(1,41) = 10.13, p = 0.003) and in Cg3 (F(1,44) = 7.19, p = 0.010), with the activation of G-proteins by CB1 receptor being more marked in females than in males (Table 4).
To better investigate the influence of gender, data were analysed separately for males and females by two-way ANOVA (factors: nicotine treatment and CP 55,940 treatment). As shown in Table 4, the separate two-way ANOVA for males revealed significant main effects of nicotine treatment in Cg1 (F(1,21) = 6.27, p = 0.021), Cg3 (F(1,21) = 5.86, p = 0.025), CA1 (F(1,20) = 7.67, p = 0.012) and CA3 (F(1,20) = 7.58, p = 0.012), and of CP 55,940 treatment in CA1 (F(1,20) = 7.23, p = 0.014) and CA2 (F(1,19) = 10.18, p = 0.05), while a strong trend in CA3 (F(1,20) = 4.13, p = 0.056) was observed. Thus, sub-chronic adolescent treatment either with NIC or CP 55,940 increased CB1 receptor-mediated G protein activation in specific brain areas of adult male rats. On the other hand, two-way ANOVA for females only revealed a main effect of NIC treatment in Cg1 (F(1,21) = 10.22, p = 0.04) (Table 4).
To determine whether the altered G-protein mediated cannabinoid CB1 receptor function was associated with changes in CB1 receptor density, [3H]CP 55,940 autoradiography was performed in the same brain areas of all treated groups. Table 5 shows the densitometric analysis of [3H]CP 55,940 binding. The three-way ANOVA revealed a significant effect of gender in hippocampal formation (CA1: F(1,44) = 8.60, p = 0.005; CA2: F(1,45) = 4.37, p = 0.042; CA3: F(1,44) = 6.04, p = 0.018) with females showing lower CB1 receptor density than males. Further, a significant overall effect of NIC treatment was found in Cg1 (F(1,44) = 4.31, p = 0.044) and in Cg3 (F(1,45) = 16.86, p < 0.001). The separate analysis for males and females by two-way ANOVA (factors: nicotine treatment and CP 55,940 treatment) revealed a main effect of NIC in Cg3 of males (F(1,22) = 13.02, p = 0.002) and females (F(1,23) = 4.43, p = 0.046), indicating a lower CB1 receptor density in NIC-treated animals when compared to control rats, whereas no significant interactions were found (Table 5).
Long-term effects of adolescent exposure to chronic treatments with nicotine and/or the cannabinoid agonist CP 55,940 on CB1 cannabinoid receptor density in several brain areas of adult rats
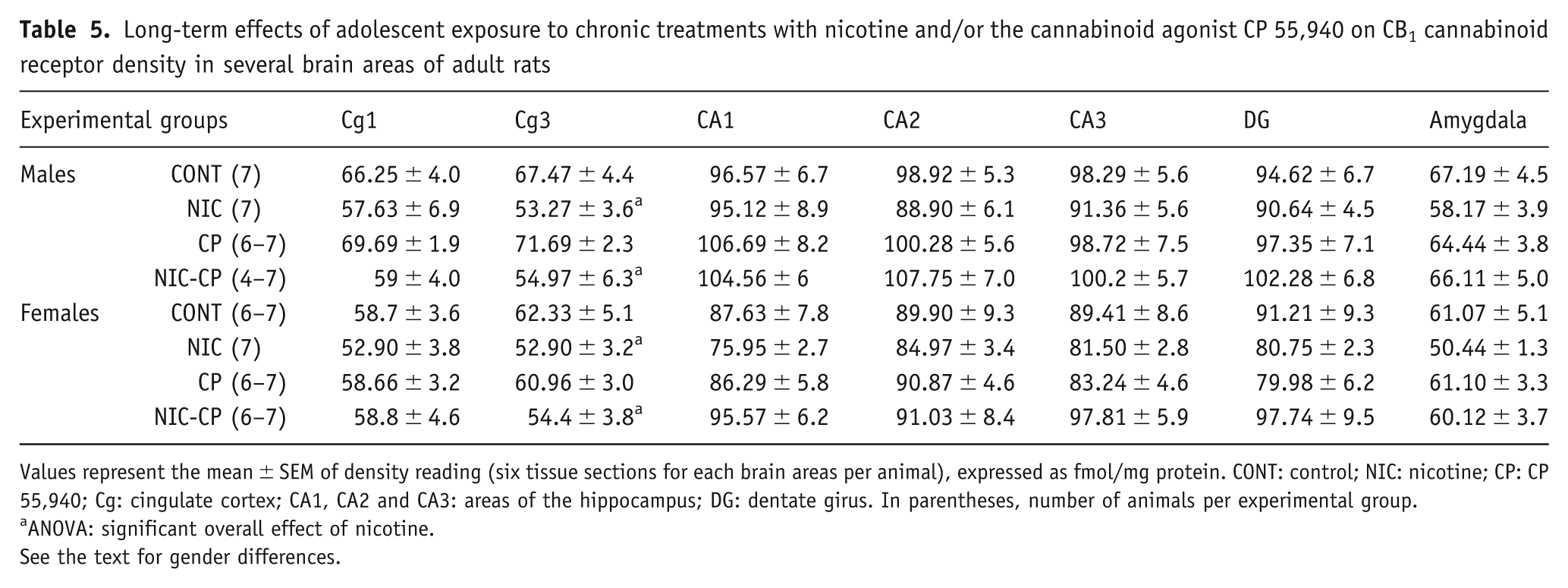
Values represent the mean ± SEM of density reading (six tissue sections for each brain areas per animal), expressed as fmol/mg protein. CONT: control; NIC: nicotine; CP: CP 55,940; Cg: cingulate cortex; CA1, CA2 and CA3: areas of the hippocampus; DG: dentate girus. In parentheses, number of animals per experimental group.
ANOVA: significant overall effect of nicotine.
See the text for gender differences.
[3H]nicotine-nAChRs autoradiography
Densitometric analysis of [3H]nicotine-nAChRs is shown in Table 6. The three-way ANOVA rendered a significant main effect of gender in amygdala (F(1,45) = 7.29, p = 0.010), as well as a significant effect of nicotine in the PFC (F(1,42) = 12.31, p = 0.001), in CA1 (F(1,48) = 5.38, p = 0.025) and in DG (F(1,48) = 4.95, p = 0.031). Separate analyses in each gender by two-way ANOVA (factors: NIC treatment and CP treatment) only revealed a significant effect in PFC for female rats. As shown in Table 6, NIC-treated females showed a decreased [3H]nicotine-nAChRs density in PFC when compared to its corresponding control group.
Long-term effects of adolescent exposure to chronic treatments with nicotine and/or the cannabinoid agonist CP 55,940 on nicotine-nAChRs receptor density in several brain areas of adult rats
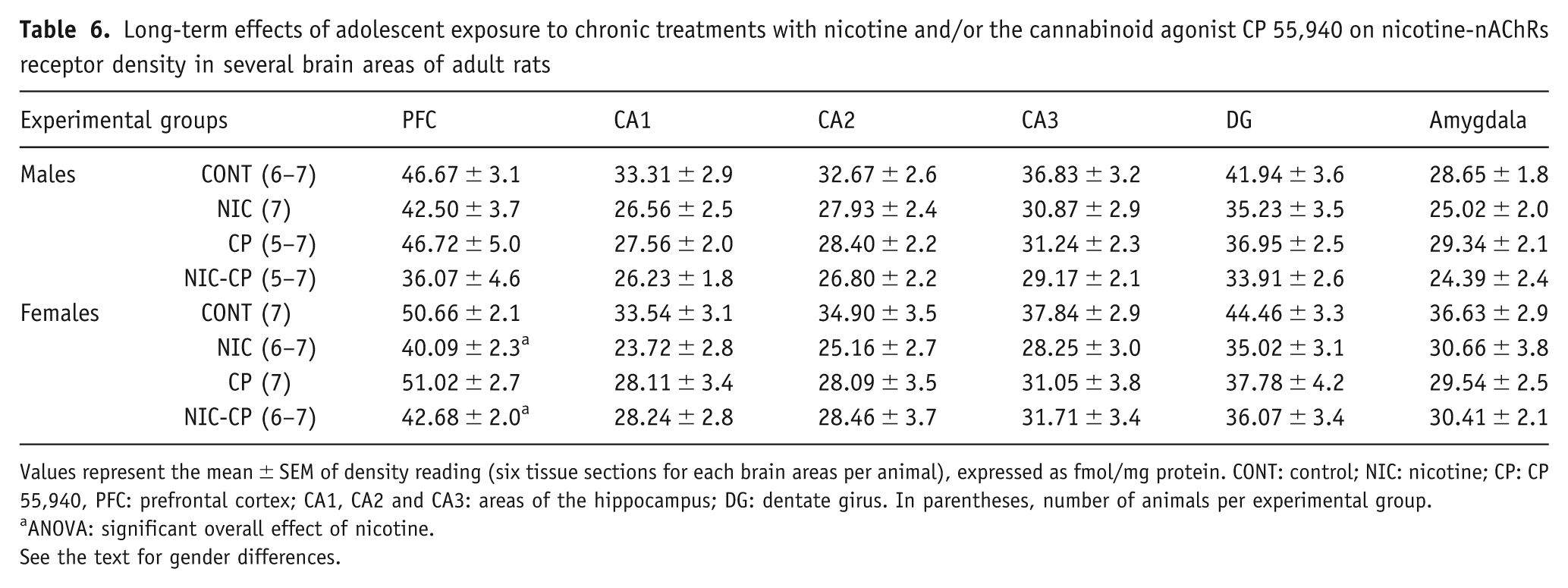
Values represent the mean ± SEM of density reading (six tissue sections for each brain areas per animal), expressed as fmol/mg protein. CONT: control; NIC: nicotine; CP: CP 55,940, PFC: prefrontal cortex; CA1, CA2 and CA3: areas of the hippocampus; DG: dentate girus. In parentheses, number of animals per experimental group.
ANOVA: significant overall effect of nicotine.
See the text for gender differences.
In brief, regarding receptor assays, both, nicotine and cannabinoid treatments induced a long-lasting increase in CB1 receptor activity (CP-stimulated GTPγS binding) in male rats, and the nicotine treatment also induced a decrease in nicotinic receptor density in the prefrontal cortex of females.
Discussion
Behavioural experiments and body weights
The present results show that sub-chronic adolescent exposure to nicotine and/or a cannabinoid agonist during the periadolescent period induced gender-dependent long-term deleterious effects on diverse aspects of memory function. In the object location task, all pharmacological treatments induced significant decreases in the DI of females, whereas no significant effects were found among males. These data suggest that females may be more vulnerable than males to the effects of nicotine and cannabinoids on tasks requiring spatial memory. On the other hand, in the novel object test, which provides information about recognition memory (novel vs. familiar objects), NIC-treated females, but not males, showed a significantly reduced DI, whereas the effect of the cannabinoid agonist (also a decrease in the DI) was only significant in males. Thus, in this specific task, the results indicate a differential drug-dependent vulnerability for males and females. The peripubertal period appears to be critical for the development of cannabinoid CB1 receptors and endocannabinoid levels (Rodriguez de Fonseca et al., 1993; Wenger et al., 2002). Therefore, it is conceivable that chronic interference by cannabinoids with the developing endocannabinoid system during this critical time-interval leads to severe and persistent functional impairments (Schneider and Koch, 2007). There is indeed evidence indicating that chronic administration of cannabinoid agonists during the periadolescent period causes diverse persistent behavioural alterations in adulthood. For instance, a 21-day treatment with CP 55,940 in 30-day-old rats resulted in a lasting impairment of working memory (O’Shea et al., 2004). Interestingly, these later behavioural changes were observed in adolescent but not adult drug-treated rats. In other study chronic pubertal treatment with another cannabinoid agonist, WIN 55,212-2 (WIN), resulted in impaired memory in adulthood (Schneider and Koch, 2003). In line with the study by O’Shea et al. (2004), these authors also showed that the chronic treatment with WIN during adulthood did not lead to behavioural changes (Schneider and Koch, 2003). A more recent study performed in male rats has shown that pubertal, but not adult, chronic WIN administration induced persistent disturbances in object and social recognition memory (indicating impairments in working memory and social memory, respectively) and led to social withdrawal and alterations in social behaviour and self-grooming. Furthermore, acute administration of the synthetic cannabinoid receptor agonist WIN induced more severe effects on behavioural performance in pubertal than in adult rats (Schneider et al., 2008). Thus, the early adolescent period appears to have a unique vulnerability to at least some of the adverse effects of cannabinoids. In support of these experimental data, early-onset cannabis users (who began smoking before age 17) exhibit poorer cognitive performance than late-onset users (who began smoking at age 17 or later) or control subjects, especially in verbal IQ (Pope et al., 2003). On the other hand, O’Shea et al. (2006) found that chronic exposure to the cannabinoid agonist CP 55,940 during perinatal, adolescent or early adulthood induced similar long-term memory impairments in male rats. To explain the different results with respect to their previous study performed in female rats (O’Shea et al., 2004, see above), they claimed that adult males might be more vulnerable than adult females to some detrimental effects of cannabinoids, such as cognitive effects. In line with this proposal, the present results show that, in the novel object test, males were more vulnerable than females to the detrimental effects of the chronic cannabinoid treatment. The present data also indicate that, in the object location task, only the females showed a significantly impaired performance in response to cannabinoid exposure, suggesting that diverse aspects of memory function may be differentially affected in each gender. In addition to cognitive effects, other cannabinoid effects have been also shown to be sexually dimorphic. Thus, Rubino et al. (2008) recently demonstrated that THC chronic administration in adolescent rats induced subtle but lasting alterations in the emotional circuit ending in depressive-like behaviour in adulthood and that this effect was observed in females but not in male rats. In a more recent study performed only in male rats, these same authors (Rubino et al., 2009) showed that a subchronic treatment with THC from 35 to 45 postnatal days, resulted, in the adulthood (PND 75), in a worse performance in the radial maze, though no alteration was found in their aversive memory (passive avoidance). Thus, it seems that, in agreement also with our results, the long-term residual effects of adolescent chronic cannabinoid exposure are gender and task dependent. The duration and onset of the treatments are also important factors that may affect the outcomes but it seems clear that, in all cases, the effects of cannabinoids on cognitive function are deleterious and can be observed after a long wash out period. Human studies also suggest the existence of gender differences as regards cannabis-induced cognitive impairment in young people (Pope et al., 1997), though much more research is necessary evaluating sexual dimorphisms.
It is very likely that long-term cognitive effects of adolescent cannabinoid exposure are related to less synaptic contacts and/or less efficient synaptic connections throughout the hippocampus and this could represent the molecular underpinning of the cognitive deficit induced by adolescent cannabinoid treatment (Rubino et al., 2009). Moreover, it is tempting to speculate that a differential effect on synaptic plasticity could be found depending on the gender of the animals in parallel with the differential behavioural impact. A possible link between the impaired memory observed specifically in males pre-exposed to cannabinoid in the novel object test and the increased functional activity of their CB1 receptors (see below) might be the CB1-mediated inhibition of glutamatergic and GABA-ergic neurons involved in mnemonic circuits (Ferraro et al., 2009; Larkin et al., 2008; Viveros et al., 2007).
In spite of the well-known negative effects of nicotine withdrawal on cognitive processes (Jacobsen et al., 2005, 2006, 2007; Semenova et al., 2007), there are few studies analysing the effects of nicotine withdrawal in rodent models of adolescence. Adult animal studies provide evidence both for positive effects of nicotine and for negative withdrawal effects. For example, in the five-choice serial reaction test of attention in food-restricted rats, it is possible to see improved performance following nicotine administration and impaired performance following nicotine withdrawal (Stolerman et al., 2000). The withdrawal-induced impairments were not long lasting and a complete recovery in performance was achieved (Levin et al., 2006; Shoaib and Bizarro, 2005). It is worth noting that, in the present study, a clear detrimental effect of adolescent nicotine on memory was observed approximately 1 month after the last injection of the drug. Thus, it seems likely that adolescence represents a particularly vulnerable period in relation to persistent cognitive effects of nicotine withdrawal. In support of this claim, adolescent daily tobacco smokers experienced acute impairment of verbal memory and working memory after smoking cessation, along with chronic decrements in cognitive performance. These findings have been related to the fact that at early developmental periods neurotoxic effects of nicotine might be enhanced (Jacobsen et al., 2005). Further, smoking abstinence in adolescent smokers appears to be associated with reductions in the efficiency of working memory neurocircuitry and alterations in the functional coordination between components of the working memory neurocircuit and it has been suggested that these alterations may stem from effects of nicotine exposure on dopaminergic systems during adolescent development (Jacobsen et al., 2007).
The impaired performance of the treated animals in the memory tasks described here is not attributable to an increased anxiety level since no significant differences were found in the elevated plus maze test of anxiety. In accordance with these data, we have previously found that a subchronic nicotine treatment (0.4 mg/kg i.p., once daily) during the periadolescent period (postnatal days 34–43) did not induce any significant change in the anxiety levels of the animals when tested in the plus maze 24 h after the last injection of nicotine (Marco et al., 2006). Moreover, a previous study on adult rats also indicated that a subchronic treatment with nicotine (i.e. 0.8 mg/kg) did not induce anxiogenic-like effects in the elevated plus maze when this test was performed 24 h after the last injection of nicotine (López-Moreno et al., 2007). On the other hand, there are also studies indicating an anxiogenic-like behaviour in nicotine abstinent adult (Irvine et al., 2001; Slawecki et al., 2004) and young (Slawecki et al., 2004) rats treated with different doses and/or employing different routes of administration. It is likely that these factors, as well as the time elapsed between the last nicotine injection and behavioural testing, affect the resulting responses in this test. A similar complex scenario appears in the literature regarding long-lasting effects of cannabinoid agonists on anxiety so that the direction of the effects may depend on the duration of the pharmacological treatment, the time elapsed between the end of the chronic treatment and the beginning of behavioural testing and the specific test employed (see for review Viveros et al., 2005).
Another finding that should be mentioned in relation to the animals exposed to the cannabinoid agonist during the adolescent period is the high percentage of animals from this group that failed to explore the objects during the training phase in both, the object location and the novel object tests. This observation might be somehow related to the social withdrawal shown by adult animals that had been exposed to chronic cannabinoid (WIN) administration during pubertal development (Schneider et al., 2008).
In apparent contradiction with the clearly negative long-term effects of the chronic cannabinoid exposure on the perfomance of the animals in the object location and object recognition tests, we found that, in the spontaneous alternation task that was performed at PND 44 (1 day after finishing the chronic CP treatment), males administered with the chronic cannabinoid treatment showed a modest though significant increase in the DI, suggesting a modestly better performance than their respective control group. We interpret this paradoxical action as a rebound or withdrawal effect. In line with these findings, Schneider et al. (2008) showed that male rats that had received a chronic pubertal cannabinoid treatment, when tested 24 h after the last cannabinoid injection showed certain behavioural alterations that differed completely from acute and long-term cannabinoid effects. This finding was interpreted by the authors as a sign of withdrawal from chronic cannabinoid treatment (Schneider et al., 2008).
It is also worth noting that we found several sexual dimorphisms among control non-treated animals. For instance, in the spontaneous alternation task, control females showed a better performance than control males (significantly increased DI) and in the elevated plus maze, control females showed a lesser level of anxiety than their male counterparts. These results are in accord with previous data showing that, in the elevated plus maze test, female rats showed a reduced aversion to the open arms compared to male rats (Johnston and File, 1991)
The present results also showed that the cannabinoid agonist induced a decrease in body weights that lasted beyond the treatment period, though normal weights were recovered before the adulthood. In a previous study we found that administration of CP during PND 35–45 reduced both body weight and food intake. Moreover, we observed development of tolerance to the effects of CP on motor activity within the period of treatment, whereas the decrease in food intake lasted during the whole treatment period (Biscaia et al., 2003), suggesting that the effect on food intake could not be entirely attributable to a decrease motor activity. By using a similar methodology, THC during adolescence reduced growth in male and female rats, the body weight reduction being related to the reduction of food intake. This effect seemed restricted to the period of THC exposure, and when the rats reached adulthood neither body weight nor food intake differed significantly from controls (Rubino et al., 2008). Other authors have also reported dose-dependent weight loss during (and after) treatment with other cannabinoid agonist, WIN 55,212-2 in adult rats (López-Moreno et al., 2004). Moreover, in a recent study we showed the presence of an active endogenous cannabinoid system regulating energy expenditure and metabolism during adolescence. In fact, we found that the administration of the CB1 receptor antagonist surinabant during adolescence or post-adolescence reduced feeding and body weight gain (Lamota et al., 2008). It might be speculated that there is a compensatory reduced hypothalamic CB1 function during the time interval when the exogenous agonist is being administered which might contribute to reduced food intake and body weight. An additional factor might be the aversive state caused by the acute administration of the drug (McGregor et al., 1996). In any case, since the animals treated with CP recovered normal body weight during the post-treatment period, the long-term behavioural effects of the cannabinoid receptor agonist are not attributable to impaired nutritional state or somatic growth. As for the sexual dimorphisms observed, it is worth mentioning that there is evidence for sex differences in how cannabinoids might acutely regulate various indices of energy homeostasis (i.e., food intake, metabolism, core body temperature, interactions with neural substrates known to modulate appetite) (for a review see Farhang et al., 2009)
Receptor assays
A relevant finding of the present study is the occurrence of gender differences in CB1 receptor function among the control (vehicle-injected) animals, with control females showing higher CB1 receptor-mediated G protein activation in hippocampal formation when compared to control males. On the other hand, CB1 receptor density tended to be lower in females (general effect of gender in the hippocampal formation). Previous studies also indicated a gender difference in hippocampal CB1 levels with males having higher CB1 levels than females (Western blot analysis) (Reich et al., 2009). Thus, it may be that the lower CB1 receptor density in females might be somehow compensated by higher functional activity, at least in certain brain regions. In relation with nAChRs, we only found significant gender differences in nAChRs density of the amygdala with females showing a higher receptor density.
The adolescent subchronic cannabinoid exposure induced an increased CB1 receptor-mediated G protein function in the hippocampal CA1 and CA2 fields of males but not females, indicating both gender- and region-dependent effects of the cannabinoid treatment, whereas no effect on CB1 receptor density was found in any of the regions analysed. These different effects on CB1 receptor density and function have been previously described in adult animals. For example, chronic treatment with the cannabinoid agonists anandamide or WIN produced either CB1 receptor desensitization or sensitization without down- or up-regulation of the receptors (Castelli et al., 2007; Rubino et al., 2003). The few reports available on the long-term effects of adolescent cannabinoid exposure on brain CB1 receptor expression and function have rendered apparently contradictory results. Thus, whereas Ellgren et al. (2007) who only used males, did not find any change, Rubino et al. (2008) showed a permanent down-regulation of CB1 receptor density and function within the nucleus accumbens, ventral tegmental area and amygdala in female rats, whereas male had significant alterations only in amygdala and hippocampus. These two latter studies employed THC for their treatments. Obviously discrepancies among the different studies are attributable not only to the specific agonist employed but also to other methodological differences such as duration of the treatment and time interval between pharmacological treatments and binding studies.
The present results also demonstrate, for the first time, that adolescent subchronic nicotine exposure induced a significant long-lasting increase in CP-stimulated GTPγS binding (i.e., an increased functionality of CB1 receptors) in cingulated cortex of both genders, and in hippocampal regions of male rats. The increased function of CB1 receptor was not accompanied by receptor up-regulation, suggesting that the increased potency of CP 55,940 to stimulate [35S]GTPγS binding might be due to a sensitization of cannabinoid CB1 receptors. This effect may be due to changes in G-protein number and/or changes of their conformational structure, altering the ability of G proteins to bind [35S] GTPγS. Another possible explanation is that the increased function of CB1 receptor reflects a compensatory mechanism to the decreased levels of anandamide induced by nicotine treatment (González et al., 2002).
As for the effects of adolescent nicotine exposure on nAChRs receptor densities, our results indicate that it induced a long-term down-regulation of nAChRs in the prefrontal cortex of females. It is well known that chronic exposure to nicotine in animals, as well as smoking in humans, alters brain nAChRs. In fact, one of the mechanisms of nicotine-induced improvements in cognition may be upregulation of nAChR expression (see for review Viveros et al., 2006). On the other hand, chronic ventral hippocampal infusion of dihydro-beta-erythroidine (DHbetaE), an alpha4beta2 nicotinic receptor antagonist, as a model of nicotinic receptor loss, caused significant spatial memory impairment (Arthur and Levin, 2002). The dynamic changes observed in the cognitive effects (i.e. beneficial effects of nicotine in the short term and negative effects in nicotine withdrawal) may be related to parallel dynamic changes in nicotinic receptors. For example Trauth et al. (1999) reported different effects of adolescent nicotine infusion on brain nAChR at different time points, with down regulation being found only during the treatment. In the present study, long-lasting withdrawal from chronic nicotine treatment during adolescence, resulted in impaired memory which were clearly observed in the two memory tasks explored in this study (in females). It is likely that these gender-dependent cognitive effects are related to the observed gender-dependent down-regulation of nAChRs in the prefrontal cortex of these same animals.
In conclusion, the present results indicate that adolescent nicotine and cannabinoid exposure result in long-term gender-dependent harmful effects on memory abilities as well as in changes in brain CB1 and nicotinic receptors.
Footnotes
Acknowledgements
Red de trastornos adictivos, RD06/0001/1013 (Instituto de Salud Carlos III, Ministerio de Sanidad y Consumo), GRUPOS UCM-BSCH: Grupo UCM 951579, Plan Nacional sobre Drogas Orden SAS/1250/2009, Ministerio de Ciencia e Innovación: BFU2009-10109 (subprograma BFI), and Fondazione Banco di Sardegna. Ricardo Llorente is a predoctoral fellow at the Universidad Complutense.