
Abstract
In the present study the influence of the dopaminergic system(s) of the amygdala on the anxiolytic-like effect of the cannabinoid CB1 receptor agonist, arachydonilcyclopropylamide (ACPA), in male Wistar rats was investigated. An elevated plus-maze test of anxiety was used to assess anxiety-like behaviors. The results showed that bilateral intra-amygdala injections of ACPA (0.125, 1.25 and 5 ng/rat) and the mixed dopamine D1/D2 receptor agonist, apomorphine, at different doses (0.001, 0.01 and 0.1 µg/rat) increased percentage open arm time (%OAT) and percentage open arm entries (%OAE), indicating an anxiolytic-like effect for both of the drugs. In contrast, intra-amygdala administration of the dopamine D1 receptor antagonist SCH23390 (0.5 and 1 µg/rat) and the dopamine D2 receptor antagonist, sulpiride (2 and 3 µg/rat) decreased %OAT and %OAE, suggesting an anxiogenic-like effect for both of the drugs. Interestingly, pretreatment with a sub-effective dose of apomorphine (0.0005 µg/rat) increased, while SCH23390 (0.25 µg/rat) and sulpiride (1.5 µg/rat) decreased the anxiolytic-like effect of ACPA. It can be concluded that the dopaminergic system of the amygdala may be involved, at least partly, in the anxiolytic-like effects induced by ACPA in the rat amygdala.
Introduction
There is an increasing interest in the endocannabinoid system as part of a complex circuitry that regulates anxiety (Viveros et al., 2005). The amygdala has been accepted as an important site in regulation of anxiety (Phelps and LeDoux, 2005; Sah et al., 2003), and the high level of expression of the cannabinoid CB1 receptor within areas of the brain such as amygdala may support the participation of cannabinoids in regulation of anxiety states (Marco and Viveros, 2009). It has also been reported that the amygdala plays a key role in emotional and affective changes experienced during cannabinoid intoxication (Patel et al., 2005); therefore it can be hypothesized that the amygdala contributes to the interaction of cannabinoids with environmental stressors.
There are substantial reports that the effects of cannabinoid agonists on anxiety are biphasic, with low doses being anxiolytic and high doses anxiogenic (Moreira and Lutz, 2008; Viveros et al., 2005). To date, there is no consensus on the mechanism(s) underlying the biphasic profile of action of cannabinoids. A range of alternatives in this regard have been speculated, of which the existence of different cannabinoid receptors with different sensitivity to cannabinoids is just one (Viveros et al., 2005), but alternative hypotheses cannot be excluded. Biphasic effects of cannabinoids on acetylcholine release in the hippocampus has been related to differential, functional association of anatomically distinct subpopulations of CB1 receptors to neurotransmitter systems that have opposing effects on acetylcholine release (Tzavara et al., 2003).
Dopamine is one of the neuromodulators most active in the mechanisms underlying states of fear and anxiety (Brandao et al., 1999; LeDoux, 2000; Rodgers et al., 1994; Zahm and Trimble, 2008). It has been reported that dopamine D1 and D2 receptors mechanisms are important in mediating anxiety (De Oliveira et al., 2006; Inoue et al., 2005; Rodgers et al., 1994; Timothy et al., 1999; Tzavara et al., 2003). It has been demonstrated that systemic injections of mixed D1/D2 dopamine receptors and apomorphine (0.25, 0.5 and 1.0 mg/kg) produce anxiolytic-like effects in the elevated plus-maze (Garcia et al., 2005). Intra-central amygdala administration of apomorphine (0.1–0.3 µg/rat) induced anxiolytic-like effects in the elevated plus-maze, which was prevented by D1 or D2 dopamine receptor antagonists (Rezayof et al., 2009).
Dopaminergic and endocannabinoid interaction at different anatomical levels (i.e. amydgala, nucleus accumbens and striatum) seems to be involved in several neurophysiological responses (Li et al., 2009; Martin et al., 2008; O’Neill et al., 2009; Polissidis et al., 2009; Sperlagh et al., 2009). The amygdala receives an important dopaminergic input from the ventral tegmental area (Amaral and Corbett, 2003; Zahm and Trimble, 2008). It has been suggested that CB1 receptor signaling modulates dopaminergic pathways by influencing, directly or indirectly, the activity of dopaminergic neurons through either post-synaptic or pre-synaptic mechanisms (Laviolette and Grace, 2006). Conversely, there are some reports that endocannabinoid levels have been modulated by drugs affecting D1 and D2 receptors (Giuffrida et al., 1999; Patel et al., 2003). It has also been reported that the dopamine D3 receptor null mice, which exhibit different endocannabinoid levels, show a less anxiety-like behavior in the elevated plus-maze test (Micale et al., 2009), supporting further interaction between dopaminergic and endocannabinoid systems.
Arachydonilcyclopropylamide (ACPA) has been introduced as a selective potent agonist for the cannabinoid CB1 receptors (Hillard et al., 1999). Until now there has been no report about the effect of systemic administration of ACPA on anxiety-like behaviors. We recently reported that intra-central amygdala injection of ACPA induces an anxiolytic-like effect in rat (Zarrindast et al., 2008). Considering the importance of the amygdala in anxiety-related processes and the interaction between cannabinoid and dopaminergic systems, the aim of the present study was to investigate the involvement of the dopaminergic system of the amygdala in anxiolytic-like effect induced by ACPA. For this purpose, at first the effect of intra-amygdala injection of ACPA in the elevated plus-maze was examined. Then, role of amygdala dopamine D1 and D2 receptors in anxiety-like behaviors by themselves and participation of these receptors in the cannabinoid-induced anxiolytic-like effect were evaluated.
Materials and methods
Animals
Male Wistar rats from Pasteur Institute (Tehran, Iran), weighing 250–280 g at the time of surgery, were used. Four animals were housed per each cage (42 cm × 26 cm × 15 cm height) in a room with a 12/12 h light/dark cycle (lights on at 07:00) and controlled temperature (22 ± 2°C). They had free access to food and water, and were allowed to adapt to the laboratory conditions for at least 1 week before the surgery. Experimental groups consisted of eight rats and each animal was tested once. All experiments were performed between 09:00 and 12:00. All procedures were performed in accordance with international guidelines for animal care and use (NIH publication #85−23, revised in 1985).
Drugs
The drugs used in the present study were apomorphine hydrochloride (Sigma, Poole, UK), SCH23390 [(R(+)-7-chloro-8-hydroxy-3-methyl-1-phenyl-2,3,4,5-tetrahydro-1H-3-benzazepine hydrochloride)] (Sigma, St Louis, CA, USA), and sulpiride [(±)-N-1-(Ethylpyrrolidin-2-ylmethyl)-2-methoxy-5-sulfamoylbenzamide, (±)-5-(Aminosulfonyl)-N-[(1-ethyl-2-pyrrolidinyl)methyl]-2-methoxybenzamide] (Sigma, St Louis, CA, USA). ACPA (arachidonylcyclopropylamide; N-(2-cyclopropyl)-5Z, 8Z, 11Z, 14Zeicosatertraenanmide) was purchased from Tocris (Tocris, Cookson, UK). Apomorphine and SCH23390 were dissolved in sterile 0.9% saline. Sulpiride was dissolved in one drop of glacial acetic acid and made up to a volume of 2 mL with sterile 0.9% saline, and then was diluted to the required volume using saline. ACPA was suspended in a vehicle consisted of absolute ethanol, emulphor and 0.9% saline at a ratio of 1:1:18. The drug doses and injection times were based on a pilot study and our previous studies (for ACPA, Zarrindast et al., 2008; for dopaminergic agents, Rezayof et al., 2009; Zarrindast et al., 2003; Zarrindast et al., 2005b).
Surgery
Rats were anesthetized intraperitoneally with a ketamine/xylazine mixture (100 and 10 mg/kg, respectively) and placed in a stereotaxic frame (Stoelting Co, Illinois, USA). A midline scalp incision was made and the skin and underlying periosteum was retracted. Then, two stainless steel guide cannulae (22-gauge, Supa; Iran) were implanted bilaterally 1 mm above the center of the right and left amygdala. The coordinates for the amygdala were: −2.2 mm posterior to bregma; ±4.1 mm lateral to the midline; and −7.8 mm ventral to the dorsal surface of the skull (Paxinos and Watson, 2006). The cannulae were anchored to the skull with a jeweler’s screw and dental cement, and then two stainless steel stylets (27 gauge) were inserted into the guide cannulae to maintain patency prior to microinfusions. The surgery was done with maximal care to minimize infection, and the animals were housed in a clean box after surgery and were given postsurgical antibiotic treatment. After surgery, the animals were allowed 5 days of recovery before the behavioral testing. During the recovery period, the rats were handled for 3 min for each day prior to being submitted to the elevated plus-maze.
Drug microinjections
Intra-amygdala microinjections were performed by means of an injection cannula (27 gauge, Supa, Iran) connected by a polyethylene tube to a 1 µL Hamilton syringe. The length of the injection canula was 1 mm more than the guide cannulae, terminating 1 mm below the tip of the guide cannulae at the center of the amygdala. The animals received bilateral microinjections of 1 µL/rat (0.5 µL/side, bilaterally) of each solution over a 60 s period, and then the injection cannula was left in the place for an additional 60 s to allow diffusion of the solution from the tip of the injection cannula. The rate of infusion was controlled manually by the researcher, and after each injection and removal the Hamilton microsyringe was filled again and vacated onto paper to confirm drug delivery. In case of two injections, the second drug was injected immediately after the first using another Hamilton microsyringe. Animals were randomly allocated to the treatment conditions and were tested in a counterbalanced order.
Elevated plus-maze
The apparatus was the same as that described previously (Zarrindast et al., 2005a, 2008). Briefly, the elevated plus-maze was a wooden cross-shaped maze consisting of four arms (two open arms and two closed arms), arranged in the shape of a plus sign. The open arms had no walls (50 cm × 10 cm), but a rim of Plexiglas (0.5 cm high) surrounded the perimeter of the open arms to prevent the rats from falling. The closed arms were enclosed by walls 40 cm high (50 cm × 10 cm × 40 cm). Where the four arms intersected, there was a square platform of 10 cm × 10 cm without any walls. The whole apparatus was elevated 50 cm from the floor.
Behavioral testing
At least 1 h before testing, the rats were placed in the experimental room with sound and light attenuated. After treatments, each rat was placed in the center of the maze facing a closed arm and was allowed free exploration during the 5 min of testing. The experimental sessions were recorded by a video camera interfaced with a monitor and a VCR in an adjacent room. Videotapes were scored by a highly trained observer (intra-rater reliability ≥0.9) who was blind to the treatment conditions. The number of entries (with all four paws) into open and closed arms, and the total time in the open and closed arms were recorded. The percentage of open arm time (%OAT) and open arm entries (%OAE), used as the standard anxiety indices (Rodgers and Johnson, 1995), were calculated using the following formulae: %OAT (the ratio of the total time spent in the open arms to the total time spent in four arms × 100) %OAE (the ratio of the total entries into the open arms to the total entries in four arms × 100)
Total closed arm entries were also recorded as an index of general locomotor activity (Rodgers and Johnson, 1995).
Experimental design
Eight rats were used in each experimental group. Total doses of the drugs were dissolved in a volume of 1 µL, so bilateral intra-amygdala microinfusion of drugs was in a volume of 1 µL/rat (0.5 µL per each side). Control groups received vehicle (1 µL/rat or 0.5 µL per each side, bilaterally), and whenever the drug was dissolved in saline, the vehicle used was saline. The test session was performed 5 min after the final intra-amygdala injections, and during the testing %OAT, %OAE and locomotor activity parameters were recorded for each animal.
Experiment 1
Five groups of animals received intra-amygdala injections of vehicle or ACPA at the different doses (0.06, 0.125, 1.25 and 5 ng/rat).
Experiment 2
Five groups of rats received intra-amygdala injections of saline or apomorphine at the different doses (0.0005, 0.001, 0.01 and 0.1 µg/rat). The other four groups of rats received saline or SCH23390 at the different doses (0.25, 0.5 and 1 µg/rat), and the last five groups of rats received vehicle or sulpiride at the different doses (1, 1.5, 2 and 3 µg/rat).
Experiment 3
In this experiment, four groups of rats received intra-amygdala injections of saline–vehicle or saline–ACPA at the different doses (0.125, 1.25 and 5 ng/rat). The other four groups of rats received apomorphine (0.0005 µg/rat) plus either vehicle (apomorphine–vehicle) or ACPA at the different doses (apomorphine–ACPA).
Experiment 4
In this experiment, four groups of rats received intra-amygdala injections of saline–vehicle or saline–ACPA at the different doses (0.125, 1.25 and 5 ng/rat). The other four groups of rats received SCH23390 (0.25 µg/rat) plus either vehicle (SCH23390–vehicle) or ACPA at the different doses (SCH23390–ACPA).
Experiment 5
In this experiment, four groups of rats received intra-amygdala injections of vehicle–vehicle or vehicle–ACPA at the different doses (0.125, 1.25 and 5 ng/rat). The other four groups received sulpiride (1.5 µg/rat) plus either vehicle (sulpiride–vehicle) or ACPA at the different doses (sulpiride–ACPA).
Statistical analysis
Normality of the data was evaluated by Kolmogorov–Smirnov test. The result of this test showed the normality of data for all parameters in all groups (p > 0.05). One-way analysis of variance (ANOVA) was used for between-group comparisons of different doses of apomorphine, ACPA, SCH23390 and sulpiride by themselves. Two-way ANOVA was used for evaluation of interactions between drugs (apomorphine, SCH23390 or sulpiride with ACPA). Following a significant p-value, post-hoc Tukey test was performed to assess paired-groups comparisons. After a significant interaction between drugs, in some cases a Student’s t-test was also used for between-group comparisons (whenever using a Student’s t-test, it has been mentioned in the text and legends). Statistical significance levels of p < 0.05 were used throughout.
Results
Effect of intra-amygdala injection of ACPA on anxiety-like behaviors in the elevated plus-maze
Figure 1 shows the effects of intra-amygdala administration of the cannabinoid CB1 receptor agonist, ACPA on anxiety-like behaviors in the elevated plus-maze. One-way ANOVA revealed that ACPA altered %OAT (F (4,35)=5.31, p < 0.01) and %OAE (F (4,35)=6.32, p < 0.01), but caused no change in locomotor activity (F (4,35)=1.38, NS). Post-hoc comparisons indicated that intra-amygdala injections of ACPA produced anxiolytic-like effects at doses of 0.125, 1.25 and 5 ng.
The effects of intra-amygdala injections of ACPA on anxiety-like behaviors. Five groups of animals received intra-amygdala injections of vehicle (1 µL/rat) or ACPA (0.06, 0.125, 1.25 and 5 ng/rat). Each bar represents mean ± S.E.M. (n=8) for %OAT (A panel), %OAE (B panel) and locomotor activity (C panel). *p < 0.05 and **p < 0.01 when compared to the vehicle control group in the respective A or B panels.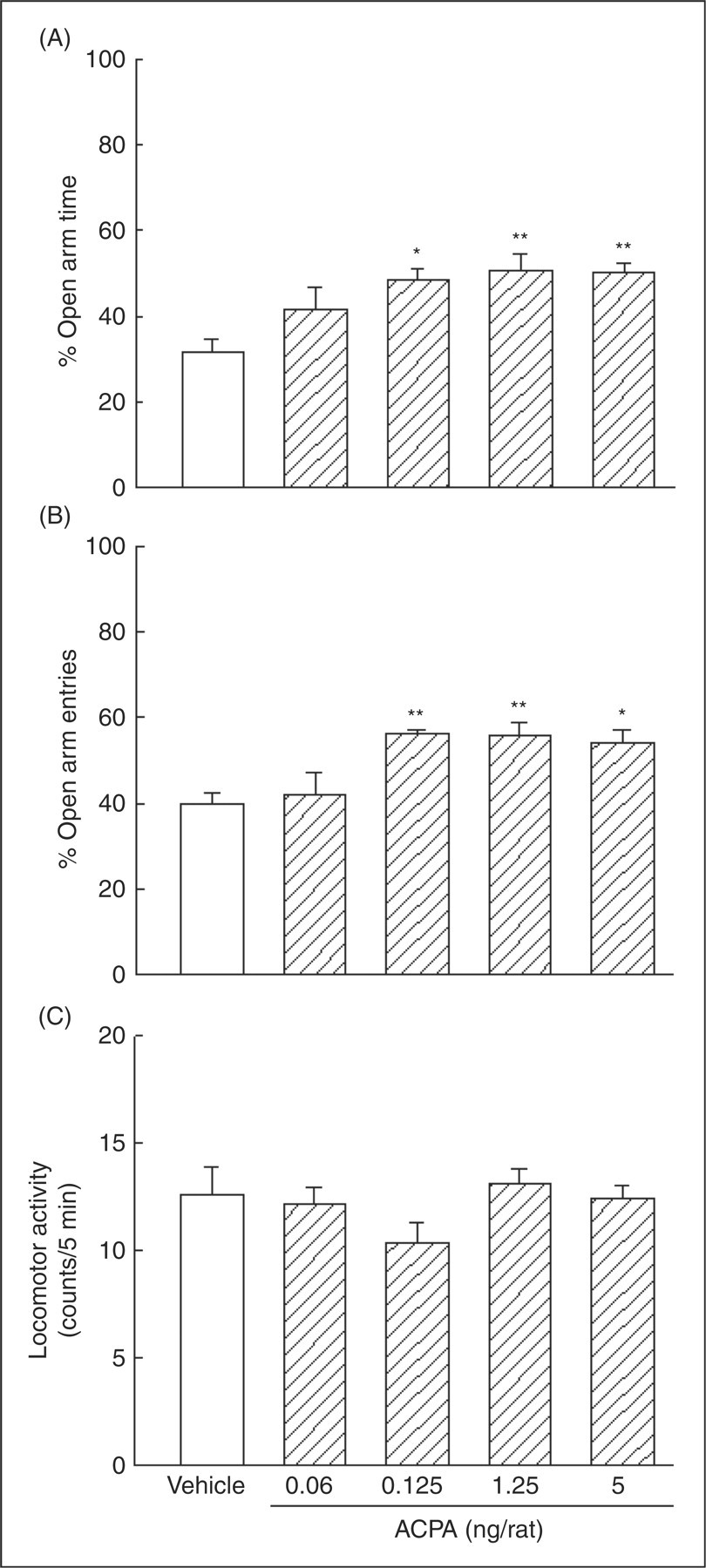
Role of dopamine D1/D2 receptors within the amygdala in anxiety-like behaviors as measured in the elevated plus-maze
Figure 2 shows the effects of intra-amygdala administration of the mixed dopamine D1/D2 receptor agonist, apomorphine (left A, B and C panels), the dopamine D1 receptor antagonist, SCH23390 (middle A, B and C panels), and the dopamine D2 receptor antagonist, sulpiride (right A, B and C panels) on anxiety-like behaviors in the elevated plus-maze. One-way ANOVA revealed that intra-amygdala administration of apomorphine altered %OAT (F (4,35)=9.56, p < 0.001) and %OAE (F (4,35)=6.7, p < 0.001), but no significant change was observed in locomotor activity (F (4,35)=0.78, NS). Post-hoc test showed that apomorphine by itself at the doses of 0.001, 0.01 and 0.1 µg/rat produced anxiolytic-like effects.
The effects of intra-amygdala injection of apomorphine (left A, B and C panels), SCH23390 (middle A, B and C panels), and sulpiride (right A, B and C panels) on anxiety-like behaviors. Five groups of animals received saline (1 µL/rat) or apomorphine at different doses (0.0005, 0.001, 0.01 and 0.1 µg/rat). The other four groups of rats received saline (1 µL/rat) or SCH23390 at different doses (0.25, 0.5 and 1 µg/rat) and the last five groups received vehicle (1 µL/rat) or sulpiride at different doses (1, 1.5, 2 and 3 µg/rat). Each bar represents mean ± S.E.M. (n = 8) for %OAT (A panel), %OAE (B panel) and locomotor activity (C panel). *p < 0.05, **p < 0.01 and ***p < 0.001 when compared to the saline control group in the respective A or B panels. +p < 0.05, when compared to the relative saline control group in the respective A or B panels. #p < 0.05 and ##p < 0.01, when compared to the relative vehicle control group in the respective A or B panels.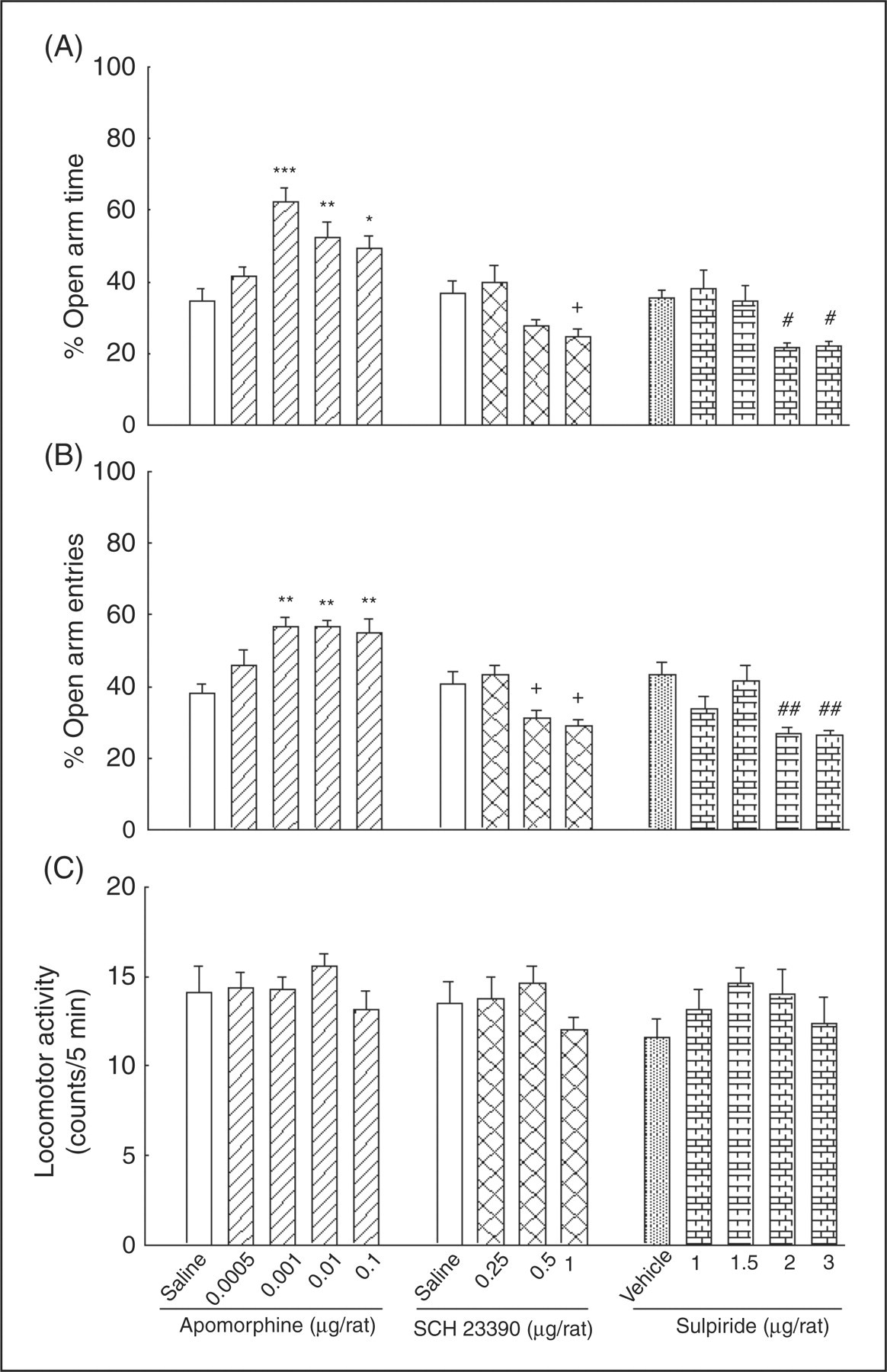
One-way ANOVA showed that intra-amygdala injections of SCH23390 altered %OAT (F (3,28)=5.75, p < 0.01) and %OAE (F (3,28)=7.92, p < 0.01), but not locomotor activity (F (3,28)=1.1, NS). Post-hoc comparisons showed that intra-amygdala administration of SCH23390 at the dose of 1 µg/rat decreased %OAT, and at doses of 0.5 and 1 µg/rat decreased %OAE.
One-way ANOVA also revealed that intra-amygdala injections of sulpiride altered %OAT (F (4,35)=5.99, p < 0.01) and %OAE (F (4,35)=7.1, p < 0.001), but not locomotor activity (F (4,35)=0.75, NS). Post-hoc comparisons revealed that intra-amygdala administration of sulpiride at dose of 2 and 3 µg/rat decreased %OAT and %OAE. The data indicated that intra-amygdala administration of the dopamine D1 and D2 receptor antagonists by themselves induced an anxiogenic-like effect.
Effect of activation of the amygdala dopamine D1/D2 receptors on the cannabinoid-induced anxiolytic-like effect in rats
Figure 3 shows the effects of intra-amygdala injections of a sub-effective dose of apomorphine along with ACPA on anxiety-like behaviors. Two-way ANOVA revealed that there was no significant interaction between apomorphine (factor A) and ACPA (factor B) (for %OAT, F (3,56)=0.46, NS; for %OAE, F (3,56)=2.42, NS; for locomotor activity, F (3,56)=2.69, NS). Comparing saline–saline and saline–ACPA groups, one-way ANOVA indicated that ACPA altered anxiety-like parameters (for %OAT, F (3,28)=6.86, p < 0.01; for %OAE, F (3,28)=5.86, p < 0.01; but not for locomotor activity F (3,28)=1.62, NS). Post-hoc tests showed that intra-amygdala injections of the highest doses of ACPA (1.25 and 5 ng/rat) increased %OAT, and for %OAE the maximum effect was achieved by the intermediate dose (1.25 ng/rat). Comparing apomorphine–vehicle and apomorphine–ACPA groups, one-way ANOVA also revealed a difference between groups in the anxiety-like parameters (for %OAT, F (3,28)=16.66, p < 0.001; for %OAE, F (3,28)=15.36, p < 0.001; but not for locomotor activity, F (3,28)=0.38, NS). Post-hoc comparisons showed no significant effect for the apomorphine–vehicle group on anxiety-like parameters compared with the saline–vehicle control group (p > 0.05). According to the post-hoc Tukey test there was no significant interaction between apomorphine and ACPA on anxiety-like behaviors, but apomorphine showed a strong tendency to increase the anxiolytic-like effect of ACPA. Comparing apomorphine–ACPA at a dose of 1.25 ng/rat with the respective saline–ACPA group with a Student’s t-test, we also found that apomorphine increased %OAE (p < 0.01).
The effects of intra-amygdala co-administration of apomorphine with ACPA on anxiety-like parameters. Four groups of animals received intra-amygdala injections of saline plus either vehicle (1 µL/rat) or ACPA at different doses (0.125, 1.25 and 5 ng/rat). The other four groups of animals received apomorphine (0.0005 µg/rat) plus either vehicle (1 µL/rat) or ACPA at the different doses. Each bar represents mean ± S.E.M. (n = 8) for %OAT (A panel), %OAE (B panel), and locomotor activity (C panel). *p < 0.05, **p < 0.01 and ***p < 0.001 when compared to the saline–vehicle group in the respective A or B panels. +p < 0.05, ++p < 0.01 and +++p < 0.001 when compared to the apomorphine–vehicle group in the respective A or B panels. ##p < 0.01 when compared to the respective saline–ACPA group in the respective A or B panels (using student t-test).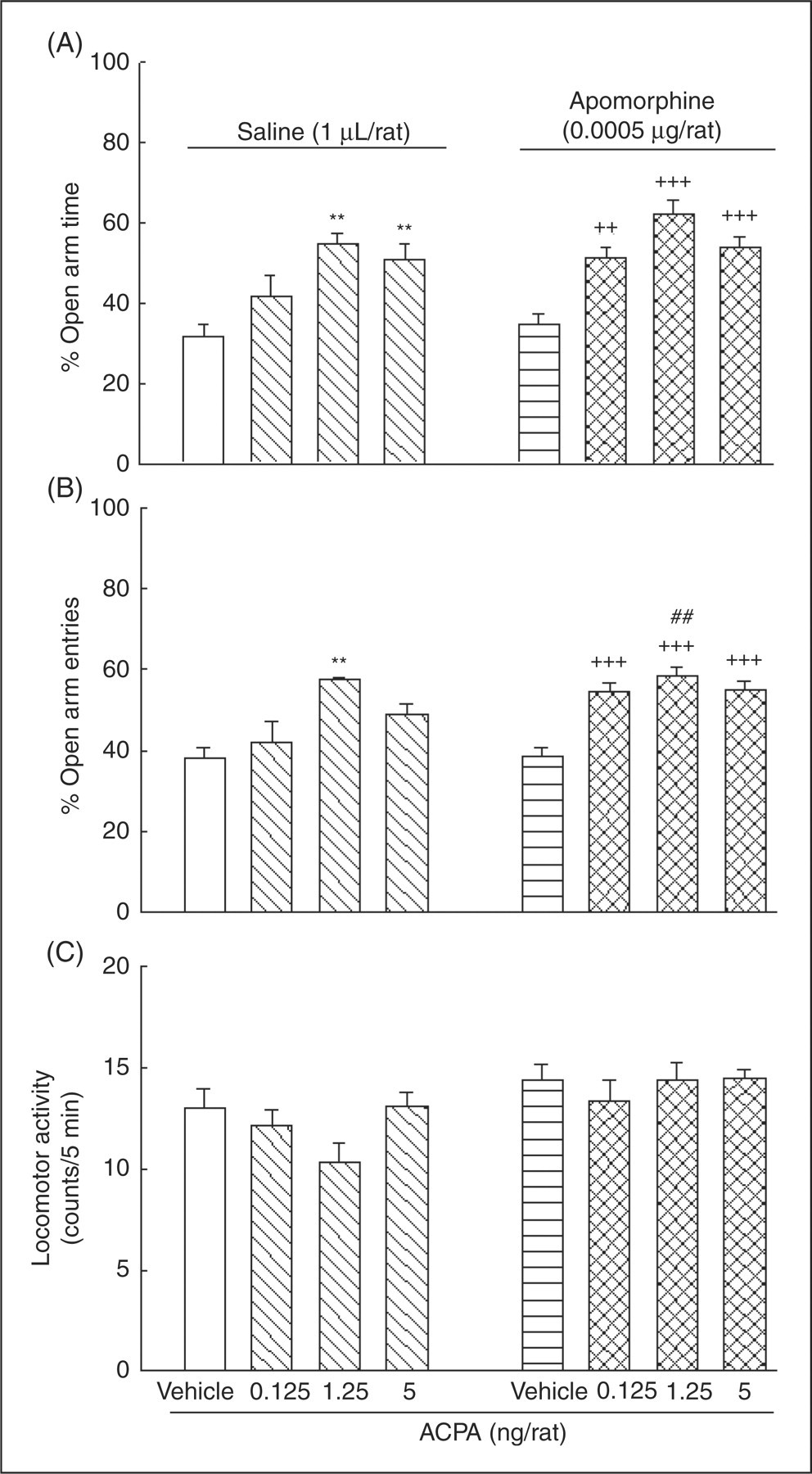
Effect of blockade of the amygdala dopamine D1/D2 receptors on the cannabinoid-induced anxiolytic-like effect in rats
Figure 4 indicates the influence of intra-amygdala injection of an ineffective dose of SCH23390 on the effect of different doses of ACPA on anxiety-like behaviors. Two-way ANOVA showed a significant interaction between SCH23390 (factor A) and ACPA (factor B) on anxiety-like parameters (for %OAT, F (3,56)=6.92, p < 0.001; for %OAE, F (3,56)=7.35, p < 0.001), but not for locomotor activity (F (3,56)=1.12, NS). Post-hoc analyses with Tukey's test showed that saline–ACPA at a dose of 1.25 ng/rat increased %OAE, and at doses of 1.25 and 5 ng/rat increased %OAT compared with the saline–saline control group. When administered along with ACPA into the amygdala, SCH23390 showed a tendency to prevent the anxiolytic-like effect of ACPA. Comparing SCH23390-ACPA with the respective saline–ACPA group, the post-hoc Tukey test revealed that the ineffective dose of SCH23390 (0.25 µg/rat) significantly prevented the anxiolytic-like effect of ACPA (1.25 ng/rat). Student’s t-test also revealed that the anxiolytic-like effect of ACPA at doses of 1.25 and 5 ng/rat was decreased in SCH23390–ACPA groups compared with the respective saline–ACPA groups.
The effects of intra-amygdala co-administration of SCH23390 with ACPA on anxiety-like parameters. Four groups of rats received intra-amygdala injections of saline–vehicle or saline–ACPA at different doses (0.125, 1.25 and 5 ng/rat). The other four groups received intra-amygdala injections of SCH23390 (0.25 µg/rat) plus either vehicle (SCH23390–vehicle) or the different doses of ACPA (SCH23390–ACPA). Each bar represents mean ± S.E.M. (n = 8) for %OAT (A panel), %OAE (B panel), and locomotor activity (C panel). *p < 0.05 and **p < 0.01 when compared to the saline–vehicle control group in the respective A or B panels. $$p < 0.01 and $$$p < 0.001 when compared to the relative saline–ACPA group in the respective A or B panels. ##p < 0.01 and ###p < 0.001 when compared to the relative saline–ACPA group in the respective A or B panels (using student t-test).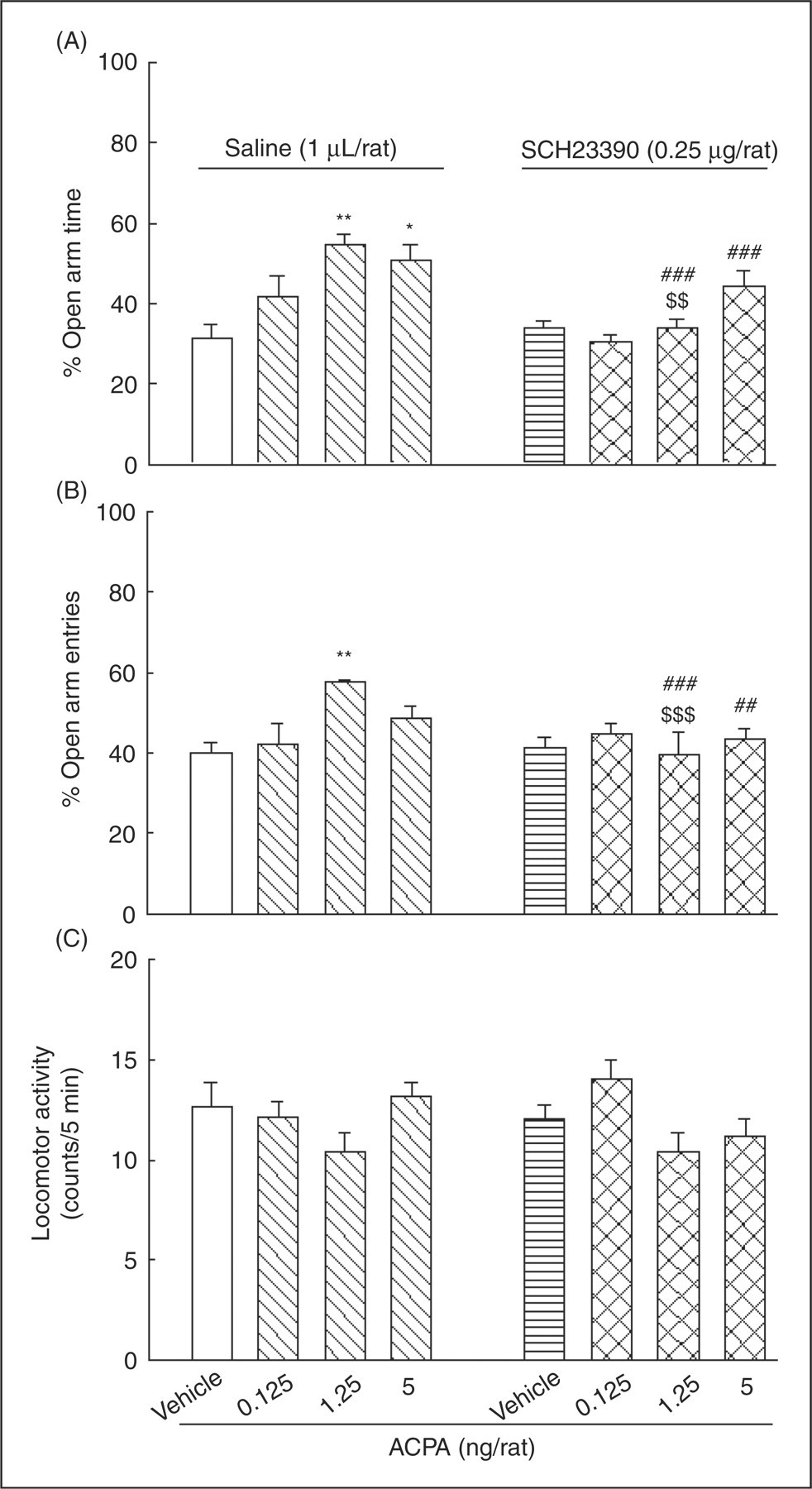
Figure 5 shows the influence of intra-amygdala injection of an ineffective dose of sulpiride on the effect of different doses of ACPA on anxiety-like behaviors. Two-way ANOVA revealed that, similar to SCH23390, there was a significant interaction between sulpiride (factor A) and ACPA (factor B) on anxiety-like parameters (for %OAT, F (3,56)=6.95, p < 0.001; for %OAE, F (3,56)=7.29, p < 0.001; but not for locomotor activity, F (3,56)=1.1, NS). Post-hoc analyses with Tukey test indicated that vehicle–ACPA at doses of 1.25 and 5 ng/rat increased %OAT, and at a dose of 1.25 ng/rat increased %OAE when compared with the vehicle–vehicle control group. In addition, comparing sulpiride–ACPA with the respective vehicle–ACPA groups, post-hoc Tukey tests also showed that sulpiride at a dose of 1.5 µg/rat, which had no effect by itself, prevented the anxiolytic-like effect of ACPA (1.25 ng/rat). Student’s t-test also revealed that anxiolytic-like effects of ACPA at doses of 1.25 and 5 ng/rat in sulpiride–ACPA groups compared with the respective saline-ACPA groups were decreased. These results may support the idea that the amygdala dopamine D1 and D2 receptors are involved, at least in part, in the anxiolytic-like effect of ACPA.
The effects of intra-amygdala co-administration of sulpiride and ACPA on anxiety-like parameters. Four groups of animals received injections of vehicle (1 µL/rat) plus either vehicle (1 µL/rat) or ACPA (0.125, 1.25 and 5 ng/rat). The other four groups received injections of sulpiride (1.5 µg/rat) plus either vehicle (sulpiride–vehicle) or ACPA at the different doses (sulpiride–ACPA). Each bar represents mean ± S.E.M. (n = 8) for %OAT (A), %OAE (B), and locomotor activity (C). **p < 0.01 when compared to the vehicle–vehicle control group in the respective A or B panels. $$p < 0.01 when compared to the relative vehicle–ACPA group in the respective A or B panels. ##p < 0.01 and ###p < 0.001 when compared to the relative vehicle–ACPA group in the respective A or B panels (using student t-test).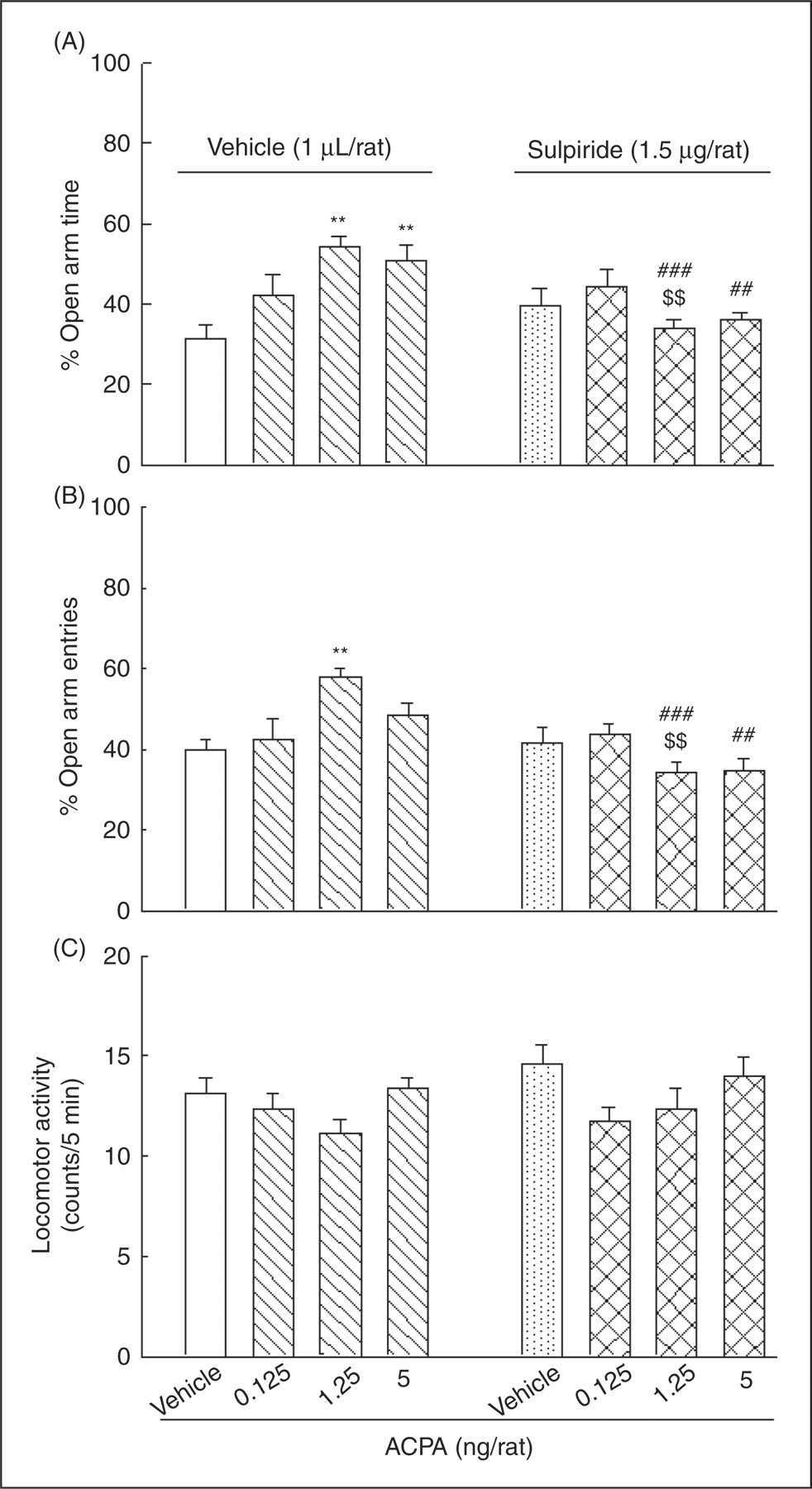
Histology
After the testing sessions the rats received 0.5 µl/side of a methylene blue solution (1%) and approximately 5–10 min later the animals were decapitated and their brains were removed. The brains were fixed in a 10% formalin solution for 5 days before sectioning. Sections were examined to determine the location of the cannulae aimed for the amygdala. The cannulae placements were verified using the atlas of Paxinos and Watson (2006). Only data from animals with correct cannulae implants were included in the statistical analyses. For animals with incorrect cannulae implants some other animals were substituted, so the final number of animals was eight per each experimental group.
Discussion
In the present study, involvement of the dopaminergic system of the amygdala in the anxiolytic-like effect of the potent agonist of cannabinoid CB1 receptor ACPA, using an elevated plus-maze test of anxiety, has been investigated. The result of the present experiments indicated that intra-amygdala administration of ACPA increased percentage preference for open arm time (%OAT) and open arm entries (%OAE) of the elevated plus-maze, indicating an anxiolytic-like effect for the drug. This result may support the idea that the cannabinoid CB1 receptors in the amygdala are important in modulating anxiety-related behaviors. In the present study, low doses of ACPA were administered into the amygdala and an anxiolytic-like effect was observed. A possible biphasic action of ACPA on anxiety-like behaviors in the amygdala requires more investigations using higher doses of the drug.
It has been reported that mice treated with systemic injections of the cannabinoid receptor agonist, nabilone, spend a greater time in the open arm of the elevated plus-maze (Onaivi et al., 1990). An anxiolytic-like effect in the plus-maze test was also induced by systemic administration of a low dose (1 µg/kg) of the cannabinoid receptor agonist, CP 55940, whereas a higher dose (50 µg/kg) of the drug induced an anxiogenic-like effect in rat (Marco et al., 2004). It has also been demonstrated that systemic administration of a low dose of Δ9-tetrahydrocannabinol (THC) produces clear anxiolytic-like responses in the light–dark box in mice (Berrendero and Maldonado, 2002). A similar biphasic profile has also been reported for central administrations of cannabinoids. Rubino et al. (2008) showed that low doses of THC injected into the prefrontal cortex and ventral hippocampus of rat induced an anxiolytic-like effect in the elevated plus-maze, while a higher level of the drug lost the anxiolytic effect and even seemed to switch into an anxiogenic-like effect. Conversely, low doses of THC injected into the basolateral amygdala produced an anxiogenic-like response, whereas higher doses were ineffective (Rubino et al., 2008). In contrast, it has been demonstrated that intra-central amygdala administration of cannabinoid CB1 receptor agonist, ACPA, induces an anxiolytic-like effect in the elevated plus-maze test in rat (Zarrindast et al., 2008).
Although there is a great consistency between the anxiolytic-like results which have been obtained for cannabinoid CB1 receptor agonists in the studies cited above, there are also discrepancies. Some possible explanations for discrepant results may include using different agonists of cannabinoid CB1 receptors, potency of the cannabinoid compounds, dosage and different injection sites even at the same structure, particularly the basolateral versus the central amygdala. In addition, the anxiolytic-like effects of ACPA might not be directly mediated via CB1 receptors, as previously demonstrated by intra-central amygdala administration of the cannabinoid CB1 receptor antagonist AM251 (Zarrindast et al., 2008).
The results of the present study also indicated that intra-amygdala administration of a mixed dopamine D1/D2 agonist, apomorphine, increased %OAT and %OAE without significant change in locomotor activity in the elevated plus-maze. Therefore, it seems that apomorphine has exerted an anxiolytic-like effect. Conversely, blockade of the amygdala D1 or D2 dopamine receptors by SCH23390 and sulpiride, respectively, reduced %OAT and %OAE parameters, suggesting an anxiogenic-like effect for these drugs.
The amygdala receives dopaminergic projections from the ventral tegmental area (VTA) and sends glutamatergic projections to the VTA and the medial prefrontal cortex (mPFC). It has been shown that GABAergic inhibitory interneurons can modulate glutamatergic projections from the amygdala (for detailed information see Gardner, 2005; Laviolette and Grace, 2006; Swanson and Petrovich, 1998; Zahm and Trimble, 2008). A simplified diagram of the anatomical circuits mentioned above has been summarized in Figure 6, but for simplicity some connections with other structures have been excluded.
Simplified diagram showing some of the functional and anatomical connections of the amygdala with the ventral tegmental area (VTA) and the medial prefrontal cortex (mPFC). The sign of (+) on axon terminals symbolizes an excitatory post-synaptic effect whereas the sign of (−) symbolizes an inhibitory post-synaptic effect. Inhibitory GABAergic interneurons in the amygdala receive dopaminergic (DA) afferents from the VTA and glutamatergic (Glu) afferents from the mPFC. Dopaminergic neurons of the VTA receive glutamatergic afferents from the mPFC and the amygdala. Glutamatergic excitatory axon terminals originated from the mPFC to GABAergic interneurons in the amygdala are pre-synaptically inhibited by dopaminergic afferents from the VTA.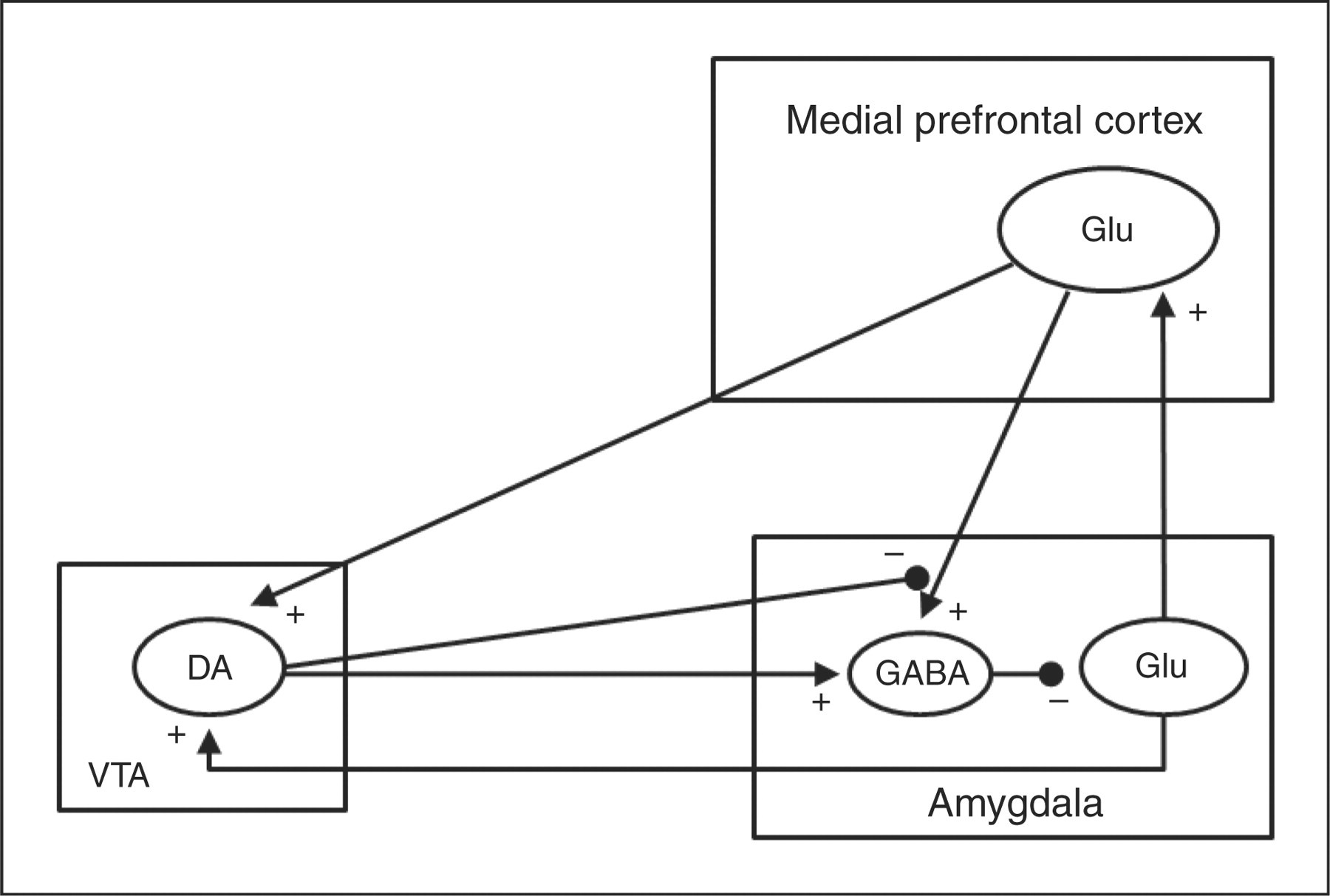
There was a lack of clear dose dependency for ACPA and apomorphine in the present study, i.e. the lowest dose tested had no effect but there was partly no difference in magnitude of responses to the higher doses. The lack of dose dependency for ACPA might suggest a permissive rather than directive role for presynaptic CB1 receptors. It has been demonstrated that dopaminergic, glutamatergic and GABAergic axon terminals in the amygdala express cannabinoid CB1 receptors (Katona et al., 2001). It has been reported that endocannabinoids, by acting as retrograde inhibitors, modulate the release of several neurotransmitters such as gamma-aminobutyric acid (GABA) or glutamate (Piomelli, 2003). Katona et al. (2001) showed that activation of CB1 receptors on GABAergic inhibitory interneurons in the amygdala reduced GABA release, thus increasing the excitability of principal cells. Conversely, there are also some reports that activation of cannabinoid CB1 receptors by the cannabinoid agonist WIN 55212-2 caused a reduction of glutamate excitatory transmission within the amygdala (Azad et al., 2003; Domenici et al., 2006). It seems that this effect of cannabinoids is mediated through presynaptic CB1 receptors on glutamaterigic axon terminals in the amygdala.
It has also been reported that the pre-synaptic distribution of dopamine receptors on glutamatergic terminals in the amygdala can excite amygdala pyramidal neurons by inhibiting pre-synaptic excitatory glutamatergic input from the mPFC to these interneurons (Rosenkranz and Grace, 2002). Therefore, the similar effects of apomorphine and ACPA in the present study may be due to pre-synaptic inhibition of the glutamatergic inputs to the amygdala. It has also been demonstrated that GABAergic interneurons in the amygdala contain cholecystokinin (CCK), therefore it is possible that CCK also be involved in the effect of cannabinoids on anxiety-like behaviors. In support of this suggestion, it has been shown that CCK plays a role in the modulation of anxiety in the basolateral amygdala (Rotzinger and Vaccarino, 2003). Taken together, the anxiolytic-like effects of intra-amygdala administration of ACPA in the present study may result from affecting the balance of neurotransmission through pre-synaptic CB1 receptors on dopaminergic, GABAergic and glutamatergic axon terminals in the amygdala.
The role of apomorphine in stressful conditions has been studied using several laboratory tests (Furlan and Brandao, 2001; Jaworski et al., 2001; Troncoso et al., 2003), but little is known about effects of apomorphine in the elevated plus-maze. Garcia et al. (2005) reported that systemic injection of apomorphine in rat induced an anxiolytic-like effect in the elevated plus-maze. Using tests of ‘illuminated area’ and ‘threatening situation’ avoidance by rats, apomorphine attenuated the state of alarm which was prevented by sulpiride, a dopamine D2 antagonist (Talalaenko et al., 1994). It has been reported that blockade of dopamine D1 receptors by systemic administration of SCH23390 causes no effects on anxiety behaviors using either elevated plus-maze test in mice (Rodgers et al., 1994) or ultrasonic vocalization paradigm in rat (Bartoszyk, 1998). On the other hand, it has been shown that systemic administration of SCH23390 inhibits the acquisition of conditioned freezing in rat as an index of fear and anxiety (De Oliveira et al., 2006; Inoue et al., 2005). It has been shown that systemic blockade of D2-like dopamine receptors by sulpiride in mice induces an anxiolytic-like effect in a conditioned fear task (Ponnusamy et al., 2005), and the elevated plus-maze (Rodgers et al., 1994). Using the white and black box test of anxiety in rat, it has been demonstrated that intra-amygdala injections of SCH233390 exerts an anxiolytic-like effect (De la Mora et al., 2005). Conversely, microinjection of SCH23390 or sulpiride into the central amygdala caused no significant change in anxiety-like behaviors in the elevated plus-maze (Rezayof et al., 2009). The discrepancy of the results obtained for dopamine receptor agents might result from the different route of administration, site of central injections, anxiety assessing tests and/or different subjects.
The results of the present study also showed that when an ineffective dose of apomorphine was co-administered with ACPA into the amygdala, a potent tendency to increase the anxiolytic-like effect was observed. This may support the hypothesis that cannabinoid CB1 receptors and the dopaminergic system in the amygdala modulate anxiety-like behaviors of the elevated plus-maze in rat. In support of this suggestion, the present results also indicated that intra-amygdala co-administration of SCH23390 and sulpiride with ACPA decreased the anxiolytic-like effect of the latter drug.
In conclusion, the findings of the present experiments show that there is a link between dopaminergic and cannabinergic systems of the amygdala in modulating anxiety-like behaviors. Considering anatomical connections within the amygdala, investigation of possible interactions of cannabinoids with the other neurotransmitter systems on anxiety-like behaviors is recommended for future research.
Footnotes
Funding
This work received no specific grant from any funding agency in the public, commercial or not-for-profit sectors.