
Abstract
Mice overexpressing CB2r (CB2xP) were exposed to open field (OF), light–dark box (LDB) and elevated plus maze (EPM) tests. Corticotropin-releasing factor (CRF) and pro-opiomelanocortin (POMC) mRNA were measured in paraventricular (PVN) and arcuate (ARC) nuclei of the hypothalamus after 30 minutes of restraint stress (RS). Anxiolytic effects of alprazolam (45 or 70 µg/kg, ip) were evaluated. GABAAα2 and GABAAγ2 mRNA were measured in the hippocampus (HIPP) and amygdala (AMY) of CB2xP and wild type (WT) mice. No differences were observed in the total distance travelled by CB2xP and WT mice in OF. Central and peripheral distances travelled significantly increased and decreased in CB2xP mice. Overexpression of CB2r reduced anxiety-like behaviours in LDB and EPM. In WT mice, RS increased CRF (82%) and POMC (42%) mRNA in the PVN and ARC nuclei, respectively. In CB2xP mice, RS also increased POMC (22%) mRNA in the ARC nucleus, but had no effect on CRF mRNA in the PVN nucleus. Administration of alprazolam was without effect in CB2xP mice. An increase of GABAAα2 and GABAAγ2 mRNA in the hippocampus and amygdala of CB2xP mice was observed. Our findings revealed that increased expression of CB2r significantly reduced anxiogenic-related behaviours, modified the response to stress and impaired the action of anxiolytic drugs.
Introduction
The characterization of the endocannabinoid system has identified two Gi/o-protein coupled receptors, CB1r and CB2r (Devane et al., 1988; Matsuda et al., 1990; Munro et al., 1993). CB1r is widely distributed throughout the rodent and human brain (Herkenham et al., 1991; Mackie, 2005; Tsou et al., 1998). The presence of a high density of CB1r in brain areas regulating emotional behaviours suggested that this receptor may have a potential role in anxiety and mood-related disorders. Genetic studies developed in mice lacking CB1r reveal increased anxiety associated with neuroendocrine and molecular adaptations in different key elements, such as the hypothalamic–pituitary–adrenal (HPA) axis (Cota et al., 2007; Haller et al., 2002; Steiner and Wotjak, 2008; Urigüen et al., 2004). This assumption was supported by several observations: (1) reduced basal corticosterone secretion, (2) increased corticotropin-releasing factor (CRF) and pro-opiomelanocortin (POMC) gene expression in the paraventricular (PVN) nucleus and anterior pituitary gland, respectively, (3) decreased glucocorticoid receptor gene expression in the CA1 of the hippocampus, (4) enhanced circadian HPA-axis-activity peak and (5) impairment of glucocorticoid feedback and hypersensitivity to stress (Aso et al., 2008; Haller et al., 2004; Steiner and Wotjak, 2008; Urigüen et al., 2004). It is interesting that direct intracerebroventricular or systemic administration of the cannabinoid CB1r antagonist SR141716A increases the concentrations of adrenocorticotropic hormone (ACTH) and corticosterone in rats (Finn et al., 2004; Manzanares et al., 1999; Patel et al., 2004; Steiner et al., 2008; Steiner and Wotjak, 2008).
In contrast, few studies have evaluated the role of CB2r in anxiety and stress-related disorders. This cannabinoid receptor has been considered to be a key element in the regulation of the immune response due to its initial identification in rat spleen and a leukocyte subpopulation in humans (Galiègue et al., 1995; Munro et al., 1993). In addition, CB2rs were identified in the brain only under pathological conditions, such as senile plaques in Alzheimer disease (Benito et al., 2003), in activated microglial cells/macrophages of multiple sclerosis (Benito et al., 2007), amyotrophic lateral sclerosis spinal cord (Yiangou et al., 2006) and near tumours (Guzmán et al., 2001; Joosten et al., 2002). Further descriptive studies in rats identified a wide distribution of CB2r in different brain areas, including the spinal nucleus, hippocampus, olfactory nucleus, cerebral cortex, amygdala, striatum, thalamus and cerebellum (Gong et al., 2006; Onaivi, 2006). The presence of CB2rs in areas involved in the response to stress and anxiety suggests the involvement of this receptor in the regulation of emotional behaviours.
Recently, Onaivi et al. indicated that intracerebroventricular administration of an antisense oligonucleotide sequence directed against CB2 mRNA reduced anxiety-like behaviours in mice (Onaivi et al., 2008a). In addition, an association with cannabinoid CB2r polymorphism Q63 R was also detected in Japanese depressed and alcoholic subjects (Onaivi et al., 2008b).
To investigate the role of CB2r in anxiety- and stress-related behaviours, transgenic mice overexpressing CB2r (CB2xP) were challenged with acute anxiogenic-like stimuli (open field [OF], light–dark box [LDB] and elevated plus maze [EPM] tests). The effects of restraint stress on CRF and POMC gene expression were studied in the PVN and arcuate nucleus (ARC) of the hypothalamus, respectively. The response to the anxiolytic drug alprazolam was evaluated in both CB2xP and wild-type (WT) mice. GABAAα2 and GABAAγ2 receptor subunit gene expression was also analysed in the hippocampus and amygdala of basal CB2xP and WT mice.
Methods
Animals
Male mice overexpressing CB2 receptors (CB2xP) on a Swiss ICR congenic background, prepared in our laboratory as described elsewhere (Racz et al., 2008b), and their corresponding Swiss ICR littermates (WT mice) were used in all experiments. At the beginning of the experiments, mice were 2–3 months old and weighed 25–35 g. All animals were maintained under controlled temperature (23 ± 2°C) and lighting conditions (light–dark cycle from 08:00 to 20:00), with free access to food (commercial diet for rodents A04 Panlab, Barcelona, Spain) and water. All the experiments were carried out between 09:00 and 13:00. The same conditions were maintained for all the behavioural tests. All studies were performed in compliance with Spanish Royal Decree 223/1998 of 14 March (BOE. 8 18) and Ministerial Order of 13 October 1989 (BOE 18), as well as the European Council Directive of 24 November 1986 (86/609/EEC).
Drugs
Alprazolam (8-chloro-1-methyl-6-phenyl-4 H-s-triazolo[4,3-a][1,4]benzodiazepine) (Sigma, Madrid, Spain) was dissolved in NaCl. The drug was administered at 45 or 70 µg/kg (0.3 mL, ip) 30 minutes before the corresponding experimental test.
Motor activity-open field test
The OF test consists of a transparent square cage 25 cm × 25 cm × 25 cm with a white Plexiglas floor. The base of the cage was divided into peripheral (12) and central (4) squares. Testing was conducted in a silent room with constant light. Mice were individually placed in the centre of the apparatus to initiate a 30-minute test session. Each session was recorded with a video camera and analysed directly with the SMART (Spontaneous Motor Activity Recording & Tracking) v.2.0 software system (Panlab, Barcelona, Spain) on a computer. Peripheral, central and total distances travelled in 30 minutes were analysed.
Light–dark box test
The LDB test uses the natural aversion of rodents to bright areas compared with darker ones (Crawley and Goodwin, 1980). In a two-compartment box, rodents will prefer dark areas, whereas rodents treated with anxiolytic drugs should increase the time spent in the light compartment. The apparatus consisted of two methacrylate boxes (20 cm × 20 cm × 15 cm), one transparent and one black and opaque, separated by an opaque tunnel (4 cm). Light from a 60 W desk lamp placed 25 cm above the light box provided room illumination. Mice were individually tested in 5-minute sessions. The floor of each box was cleaned between sessions. At the beginning of the session, each mouse was placed in the tunnel facing the dark box. The time spent in the light area by each mouse was recorded for 5 minutes. A mouse whose four paws were in the new box was considered as having changed boxes.
Elevated plus maze
The EPM consisted of two open arms and two enclosed horizontal perpendicular arms 50 cm above the floor (Lister, 1987). The junction of four arms formed a central squared platform (5 cm × 5 cm). The test began with the animal being placed in the centre of the apparatus facing one of the enclosed arms and allowed to explore freely for 5 minutes. Arm entry was considered as entry of four paws into the arm. The time spent in open arms (as a percentage of the total test time) and the number of open-arm entries were recorded.
Restraint stress procedure
Unanaesthetized mice were confined in an acrylic cylindrical tube (inner size 10 cm × 3 cm × 3 cm) for 30 minutes (Kvetnansky et al., 1966). After this period of restraint stress, the animals were removed from the tubes and housed in their cages. Control animals remained in their home cages until decapitation. Potential changes in the level of gene expression were studied by decapitating mice and removing their brains 2 hours and 30 minutes after restraint stress.
In situ hybridization histochemistry (ISHH)
Brain sections of 20 µm were cut at different levels containing the regions of interest according to Paxinos and Franklin (2001) and stored at −80°C until the day of the assay. ISHH was performed as described previously (Young et al., 1986) using synthetic oligonucleotide probes complementary to proopiomelanocortin (POMC) (CTT CTT GCC CAC CGG CTT GCC CCA GCG CCA GCG GAA GTG CTC CAT GGA GTA GGA) (Thermo Fisher Scientific, Ulm, Germany) and CRF (GGC CCG CGG CGC TCC AGA GAC GGA TCC CCT GCT CAG CAG GGC CCT GCA) (Thermo Fisher Scientific, Ulm, Germany) of the rat POMC and CRF gene mRNA, respectively. Oligonucleotide probes were labelled using terminal deoxytransferase (Amersham, Madrid, Spain) to add 35S labelled deoxyATP (1000 Ci mmol_1; Amersham, Madrid, Spain) tail to the 3’ end. The probe (in 50 µL of hybridization buffer) was applied to each section and left overnight at 37°C for hybridization. Following hybridization, sections were washed four times for 15 minutes each in 0.15 M NaCl, 0.015 M sodium citrate, pH 7.2 (1X saline sodium citrate, SSC) at 55°C, followed by two 30-minute washes in 1X SSC at room temperature, one brief water dip and blow-drying with air. The dried slides were apposed on Kodak BioMax MR-1 film (Amersham, Madrid, Spain) for 14 days. Autoradiograms were analysed with a PC computer using the public domain NIH Image program (developed at the US National Institutes of Health and available on the Internet at http://rsb.info.nih.gov/nih-image). Optical densities were calculated from the uncalibrated mode by subtracting the background from each measurement and were expressed in grey scale values. The background measurement was taken from the area of the slice with the lowest non-specific hybridization signal and subtracted from the hybridization signal measurement in the same slice. Results were presented considering mean control values as 100%. Additional brain sections were co-hybridized with 100-fold excess of cold probe or with RNAse to assert the specificity of the signal. As expected, no hybridization signal was detected in these sections (data not shown).
Analysis of GABAAα2 and GABAAγ2 receptor subunits gene expression
The gene expression of the main anxiolytic subunits of the GABAA receptor, GABAAα2 and GABAAγ2, was examined by real time PCR (RtPCR) in the amygdala and hippocampus of basal CB2xP and WT mice. Briefly, mice were decapitated and their brains were removed from the skull and frozen over dry ice. Brain sections were cut at 500 µm at different levels containing the regions of interest according to Paxinos and Franklin (2001), mounted onto slides and stored at −80°C. Sections were dissected following the method described by Palkovits (1983). Total RNA was obtained from brain punches using Biozol® Total RNA extraction reagent (Bioflux, Inilab, Madrid, Spain). After DNAse digestion, reverse transcription was carried out following the instructions of the manufacturer (Epicentre, Tech. Corp., Madison, Wisconsin). GABAAα2 and GABAAγ2 gene expression was measured using Taqman Gene Expression assay (Mm00433435_m1 and Mm00433489_m1) (Applied Biosystems, Madrid, Spain) as a double-stranded DNA-specific fluorescent dye and performed on the AbbiPrism 7700 Real Time Cycler (Applied Biosystems, Madrid, Spain). The reference gene was 18 S rRNA and was detected using Taqman ribosomal RNA control reagents. All primer-probe combinations were optimized and validated for relative quantification of gene expression. Briefly, the data for each target gene were normalized to the endogenous reference gene, and the fold change in target gene abundance was determined using the 2−ΔΔCt method (Livak and Schmittgen, 2001).
Statistical analyses
Statistical analyses were performed using the Student’s t-test when comparing two groups, one-way analysis of variance followed by the Student’s Newman–Keul test when comparing three groups and two-way analysis of variance followed by the Student’s Newman–Keul test when comparing four groups. Differences were considered significant if the probability of error was less than 5%.
Results
Motor activity-open field
No differences were found between CB2xP and WT mice in the total distance travelled (Student’s t-test, t = −1.042, p = 0.303, 42 df) (n = 26–27) (Figure 1A). However, the central distance travelled increased significantly (Student’s t-test, t = −3.504, p = 0.001, 45 df) (n = 26–27/group; Figure 1B) and the peripheral distance travelled decreased significantly (Student’s t-test, t = 2.427, p = 0.020, 41 df) (n = 26–27/group) (Figure 1C) in CB2xP mice.
Assessment of spontaneous motor activity of CB2xP and WT mice in the open field test. Motor activity (total, central and peripheral activities) was measured in a 30-min session. Columns represent the means and vertical lines ± SEM of total, central and peripheral distance (cm). *Values from CB2xP mice that are significantly different from WT mice (Student’s t-test, p < 0.05) (n = 26–27/group).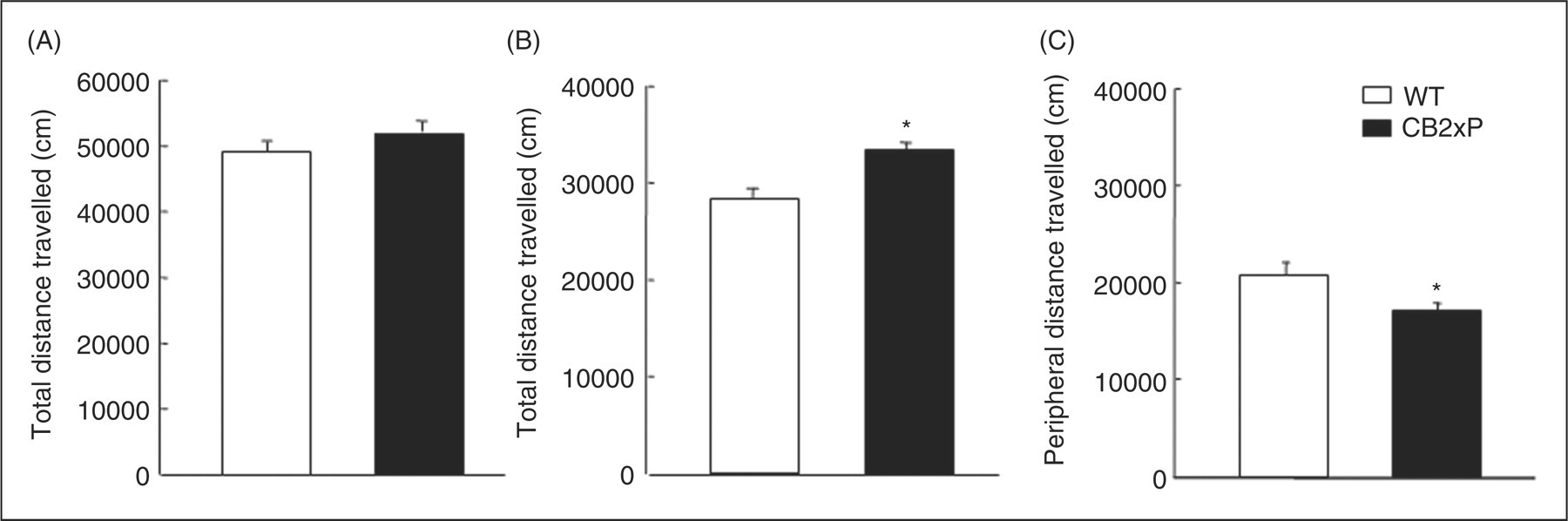
Assessment of anxiety-like behaviours
Light–dark box test
CB2xP mice spent significantly more time in the light box (Student’s t-test, t = −2.134, p = 0.045, 21 df) (n = 12–14) (Figure 2A) and displayed a significant increase in the number of transitions (Student’s t-test, t = −2.221, p = 0.039, 19 df) (n = 12–14) (Figure 2B).
Assessment of anxiety-like behaviours of CB2xP and WT mice in the light-dark box (A and B) and elevated plus maze tests (C and D). Behaviour in each test was evaluated for a period of 5 min. Columns represent the means and vertical lines ± SEM of (A) time in the light box (s), (B) number of transitions, (C) % time in open arms and (D) number of transitions in 9–14 mice. *Values from CB2xP mice (black columns) that are significantly different from WT mice (white columns) (Student’s t-test, p < 0.05).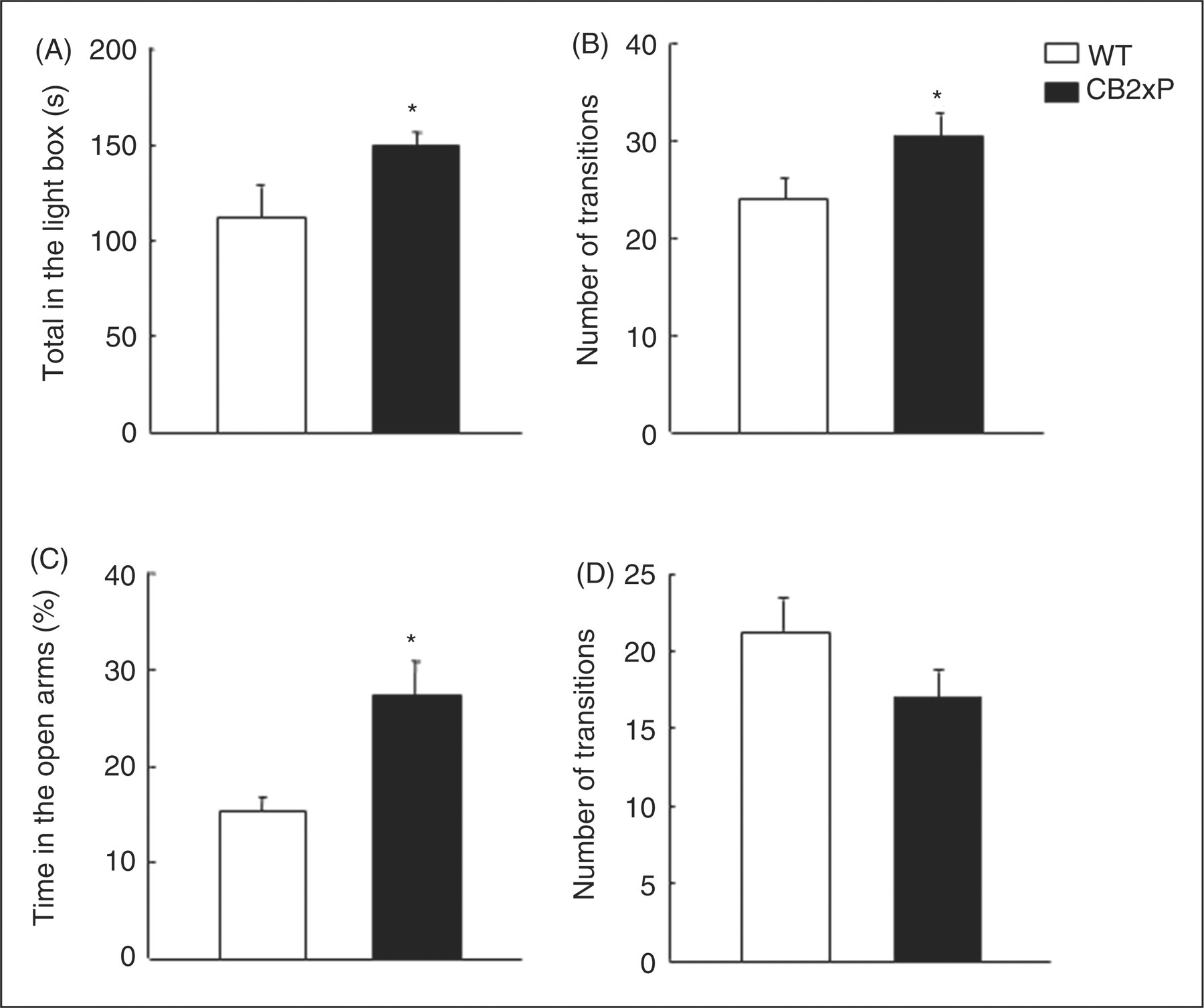
Elevated plus maze test
In the elevated plus maze test, the percentage of time spent in the open arms significantly increased in CB2xP mice (Student’s t-test, t = −3.300, p = 0.004, 17 df) (n = 9–11) (Figure 2C). No differences were observed in the number of transitions between compartments (Student’s t-test, t = 1.629, p = 0.126, 14 df) (n = 9–11) (Figure 2D).
Evaluation of CRF and POMC gene expression
CRF gene expression
In the WT mice, the results revealed that restraint stress markedly increased (82%) CRF gene expression in the PVN nucleus of the hypothalamus (Figure 3A and B). Interestingly, restraint stress failed to produce any alteration in CB2xP mice In addition, no differences were observed between non-stressed CB2xP and non-stressed WT mice (two-way ANOVA followed by the Student’s Newman–Keul test, genotype F
(1,28) = 2.216, p = 0.149, stress F
(1,28) = 7.206, p = 0.013, genotype × stress F
(1,28) = 8.568, p = 0.007) (n = 8–12/group) (Figure 3A and B).
Evaluation of CRF and POMC gene expression under basal and restraint stress conditions. Columns represent the means and vertical lines ± SEM of CRF mRNA levels (% of control) (A, B) and POMC mRNA levels (% of control) (C, D) in PVN and ARC nuclei, respectively. *Values from stressed-WT and stressed-CB2xP that are significantly different from non-stressed-WT mice. #Values from stressed-CB2xP mice that are significantly different from non-stressed-CB2xP mice (two-way ANOVA followed by the Student’s Newman–Keul test, p = 0.05). Representative autoradiograms for CRF in PVN (B) and POMC in ARC (D) of stressed and non-stressed-CB2xP and WT mice. Bar represents 1 mm.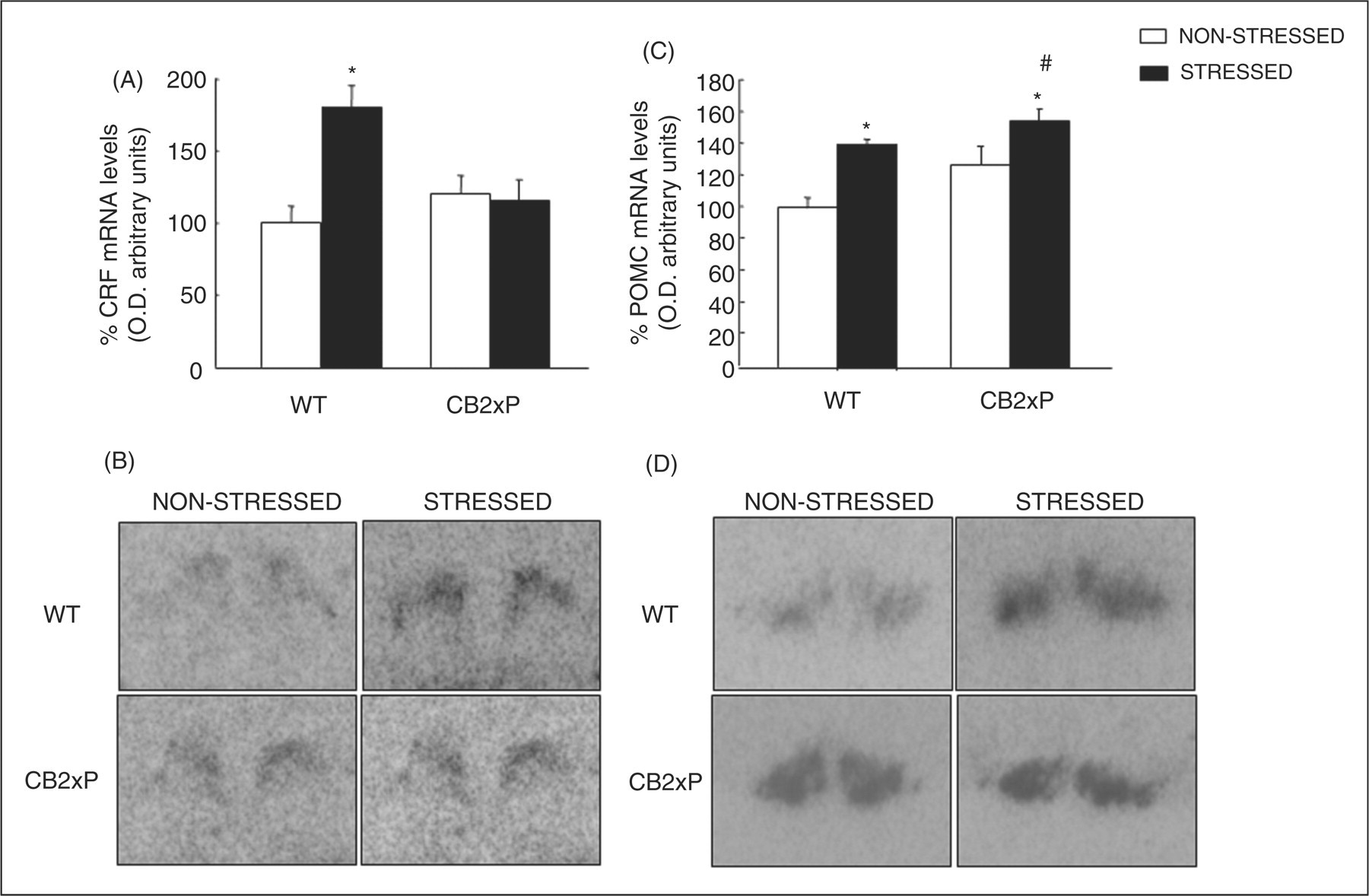
POMC gene expression
Restraint stress significantly increased (42%) POMC gene expression in the ARC nucleus of the hypothalamus in stressed WT compared with non-stressed WT mice (Figure 3C and D). It was noteworthy that restraint stress reduced the increase (22%) in POMC gene expression in CB2xP mice. In addition, no difference was observed between non-stressed CB2xP and non-stressed WT mice (two-way ANOVA followed by the Student’s Newman–Keul test, genotype F (1,17) = 4.334, p = 0.056, stress F (1,17) = 13.677, p = 0.002, genotype × stress F (1,17) = 0.568, p = 0.464) (n = 5–10/group) (Figure 3C and D).
Dose-response effects of alprazolam in the light–dark box test
The administration of alprazolam (45 and 70 µg/kg) significantly increased the time spent in the light box (one-way ANOVA followed by the Student’s Newman–Keul test, F
(2,17) = 15.709, p < 0.001) (n = 8–10/group) (Figure 4A) and the number of transitions of WT mice (one-way ANOVA followed by the Student’s Newman–Keul test, F
(2,21) = 10.909, p < 0.001) (n = 8–10/group) (Figure 4B). However, no differences in the time in the light box (one-way ANOVA followed by the Student’s Newman–Keul test, F
(1,23) = 0.0824, p = 0.921) (n = 8–10/group) (Figure 4C) and in the number of transitions (one-way ANOVA followed by the Student’s Newman–Keul test, F
(2,28) = 1.104, p = 0.346) (n = 8–10/group) (Figure 4D) were observed in CB2xP mice at either of the doses used.
Dose response effects of alprazolam in light–dark box in WT (A-B) and CB2xP (C-D). Mice were injected with alprazolam (45 or 70 µg/kg, ip) or its saline vehicle, and 30 min later were exposed to light-dark box for 5 min. Columns represent the means and vertical lines ± SEM of (A, C) time (s) in the light box and (B, D) number of transitions in (n = 8–10/group). *Values from drug-treated WT mice that are significantly different from corresponding vehicle-treated WT mice.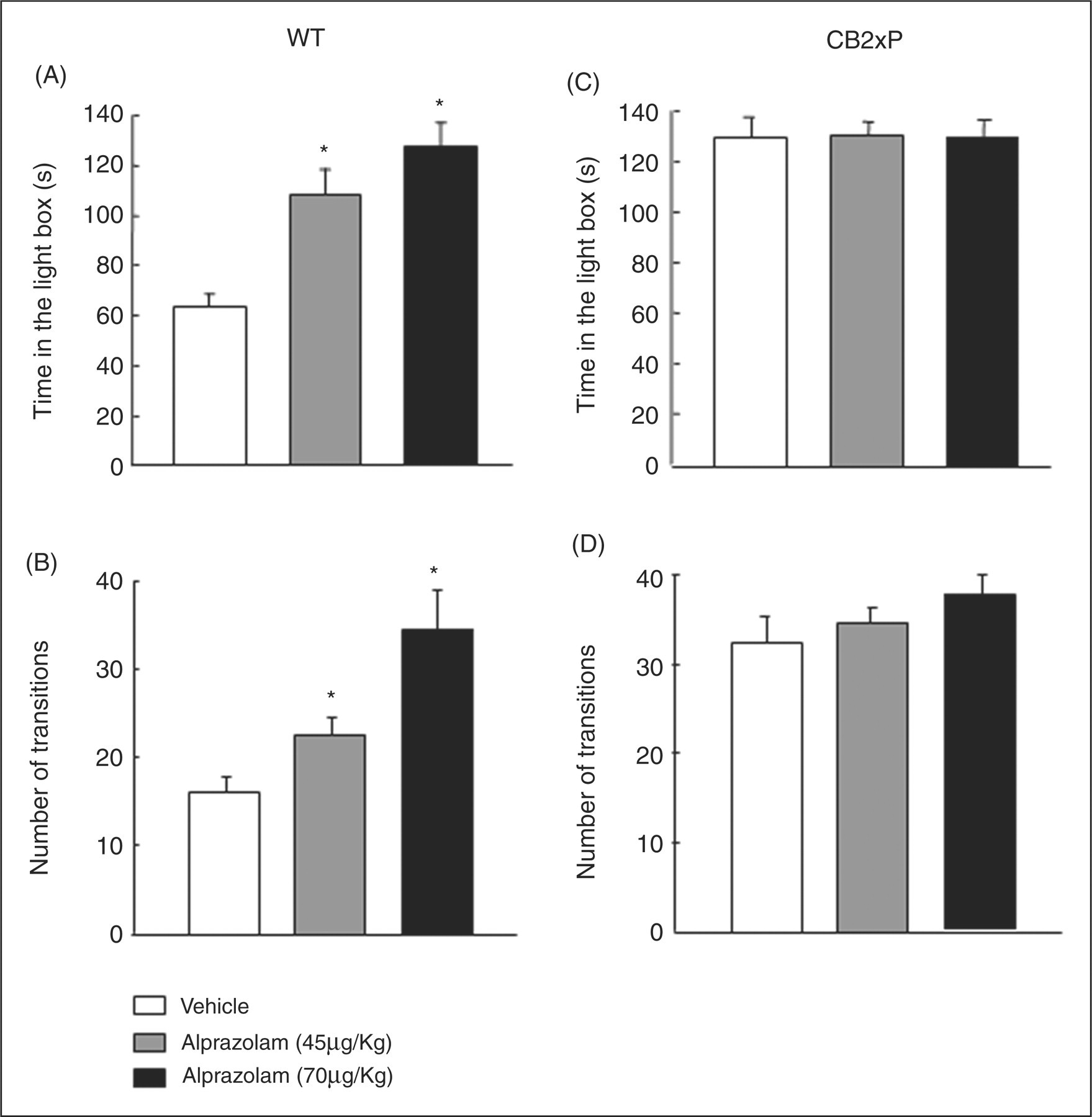
Evaluation of GABAAα2 and GABAAγ2 receptor subunit gene expression
GABAA
α
2 and GABAAγ2 receptor subunit gene expression was studied in the amygdala and hippocampus of CB2xP and WT mice. The results revealed that CB2xP mice presented increased expression of both subunits in the amygdala (Amy) and hippocampus (Hipp) compared with WT mice (GABAAα2, Student’s t-test, Hipp t = 2.523, p = 0.025, 13 df; Amy t = 2.605, p = 0.022, 13 df) (GABAAγ2, Student’s t-test, Hipp t = 2.164, p = 0.050, 13 df; Amy t = 2.209, p = 0.046, 13 df) (Figure 5) n = 7–8/group).
Relative GABAAα2 and GABAAγ2 gene expression in hippocampus (A, B) and amygdala (C, D) of CB2xP and WT mice. Columns represent the means and vertical lines the ± SEM of relative GABAAα2 and GABAAγ2
mRNA levels. *Values from CB2xP mice that are significantly different from WT mice (Student’s t-test, p < 0.05) (n = 7–8/group).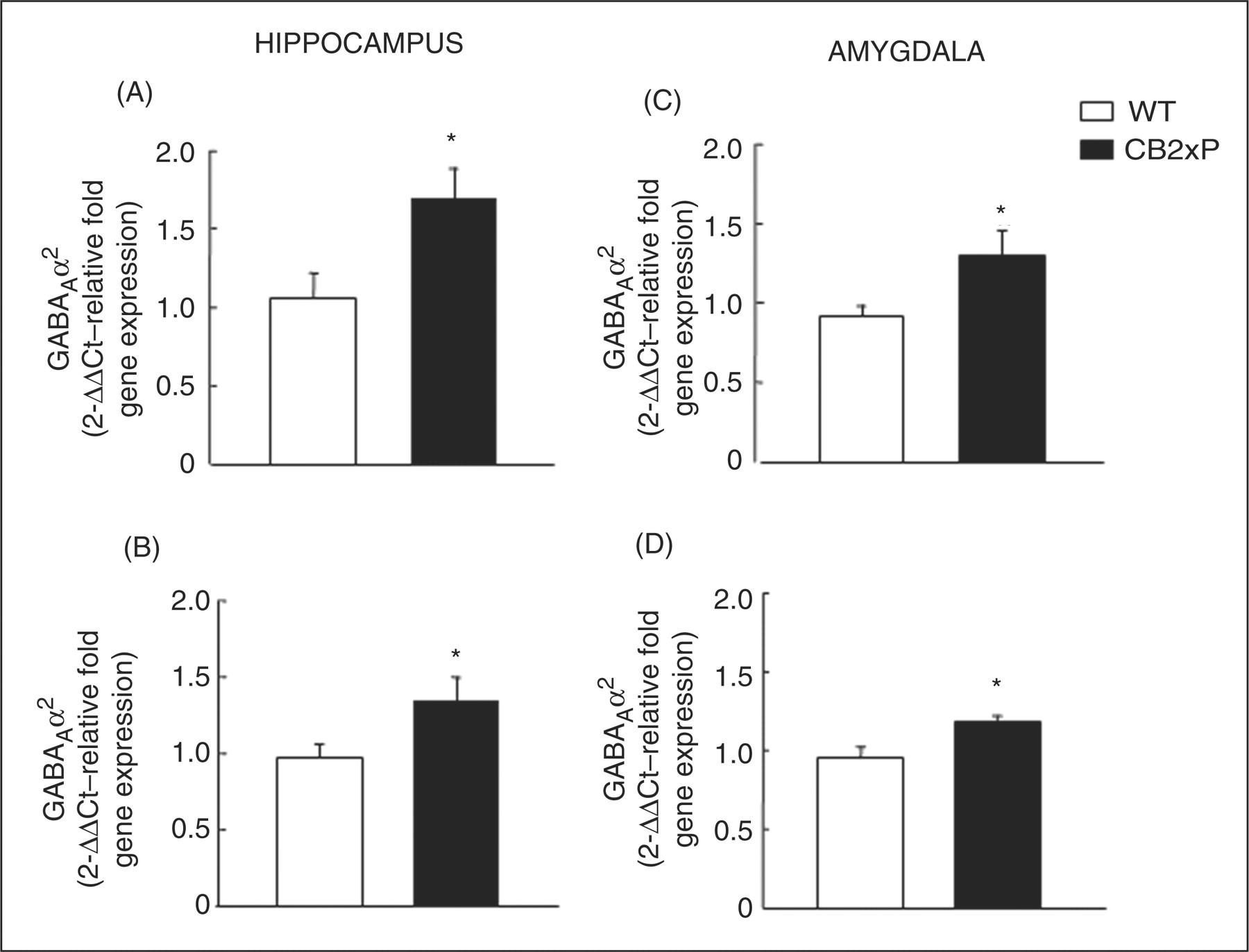
Discussion
The results of the present study provide unequivocal information about the pivotal role of the CB2r in the regulation of emotional behaviours. This assumption is supported by several observations: (1) the overexpression of the CB2r resulted in a behavioural endophenotype resistant to anxiogenic-like stimuli; (2) the effects of restraint stress on CRF and POMC gene expression were differentially altered in WT and CB2xP mice; (3) the administration of alprazolam was devoid of anxiolytic action in CB2xP mice; (4) the gene expression of GABAAα2 and GABAAγ2 receptor subunits was higher in the hippocampus and amygdala of CB2xP mice.
Previous studies evaluating the role of CB2r in the regulation of neuropathic pain and depressive-like behaviours have partially described these CB2xP mice (García-Gutiérrez and Manzanares, 2009; García-Gutiérrez et al., 2010; Racz et al., 2008a; Racz et al., 2008b). In these studies, CB2r gene expression was identified in the thalamus, periaqueductal grey matter (PAG), cervical and thoracic spinal cord, and in the different brain nuclei analysed (caudate-putamen nucleus [CPu], Nucleus Accumbens [Acc], Cingulated cortex [Cg], amygdala, hippocampus, ventromedial nucleus of hypothalamus [VMN], arcuate nucleus of hypothalamus [ARC], Substantia nigra [SN], Dorsal raphe [DR] and Medial raphe [MnR]) of WT mice. CB2r gene expression was significantly increased in these brain and spinal cord regions compared to WT mice (CPu [157%], Acc [182%], Cg [198%], amygdala [64%], hippocampus [144%], VMN [134%], ARC [158%], SN [277%], DR [50%], Mn] [57%], thalamus [95%], PAG [56%], cervical spinal cord [205%], thoracic spinal cord [176%]). The distribution of CB2r in the central nervous system suggests a potential role of CB2r in a wide variety of physiological functions. In fact, the presence of CB2r in the amygdala, hippocampus, DR and MnR (areas related to stress, anxiety and depression) suggests that re-evaluation of the possible roles of CB2r in the regulation of anxiety-like disorders would be worthwhile.
CB2xP and WT mice were exposed to the OF test to evaluate potential alterations in motor activity. The analysis of the results revealed that overexpression of CB2r did not change the total distance travelled compared to WT mice. However, CB2xP mice presented a significant increase in the central distance travelled and a decrease in the peripheral distance travelled. Considering the absence of changes observed in the total distance travelled, the differences found in the central and peripheral areas of CB2xP mice could be due to reduced vulnerability to anxiogenic stimuli such as the central area of the open field.
In order to investigate the potential involvement of CB2r in the regulation of emotional behaviours, the response of CB2xP mice to anxiogenic-like stimuli was examined in the LDB and EPM tests. The analysis of the results revealed that overexpression of CB2r resulted in a behavioural endophenotype resistant to acute anxiogenic-like stimuli (increased time spent in the light area and in the percentage of time spent in the LDB and EPM tests, respectively). These results are in contrast with those reported by Onaivi et al. (2008b). The discrepancies between the two studies may be due to the fact that the Onaivi group studied the effects of intracerebroventricular administration of CB2 antisense oligonucleotide (20 µg in 5 µL, twice a day during three days) in mice. The effects of intracerebroventricular administration of CB2 antisense oligonucleotide could act on different brain regions that may differ from those where CB2rs are overexpressed in CB2xP mice. These different actions in possibly different brain regions could underlie the discrepancies observed between the two studies. Furthermore, Onaivi et al. (2008b) used mice generated from a different genetic background (DBA/2, C57BL/6 and BALB/c strain) compared to the mice analysed in our study (Swiss ICR strain).
The marked behavioural alterations occurring in CB2xP mice are probably associated with neuroendocrine and molecular adaptations in key elements regulating anxiety. In this respect, the HPA axis is a crucial system involved in the endocrine, autonomic and behavioural response to stress. Dysfunctions of the HPA axis have been associated with stress- and anxiety-related behaviours in rodents (Arborelius et al., 1999; Dunn and Berridge, 1990; Smith et al., 1998; Stenzel-Poore et al., 1994; Timpl et al., 1998) and anxiety disorders in humans (Altemus et al., 1992; Banki et al., 1992; Bremner et al., 1997; Chappell et al., 1996; Fossey et al., 1996; Heim et al., 1997, 2000; Jolkkonen et al., 1993; Roy-Byrne et al., 1986; Smith et al., 1998; Yehuda, 1997).
The effects of 30 minutes of restraint stress on CRF and POMC gene expression in the PVN and ARC nuclei, respectively, were examined to explore potential alterations in the regulation of CRF and POMC gene expression (both of which are involved in the regulation of the HPA axis) between CB2xP and WT mice. Restraint stress significantly increased CRF (82 %) and POMC (42 %) gene expression in the PVN and ARC, respectively, in WT mice. In contrast, restraint stress failed to alter CRF gene expression and produced a lower increase in POMC gene expression (22%) in CB2xP mice. Although the precise mechanisms underlying these changes remain to be elucidated, it is tempting to speculate that CB2xP mice could present alterations in the feedback regulatory processes (synthesis and release of CRF, corticotropin, corticosterone, and glucocorticoid receptor function) responsible for maintaining the activity of the HPA axis. It is interesting to note that recent studies have pointed out the participation of the endocannabinoid system in the regulation of the HPA axis. In fact, pharmacological and genetic studies support a ‘homeostatic’ function for CB1r in the control of HPA axis (Corchero et al., 1999; Cota et al., 2007; Steiner and Wotjak, 2008; Urigüen et al., 2004). The results of this study suggest that CB2r could also contribute to maintaining steady-state control of the HPA axis. Considering that the increased expression of CB2r blocked the effects of stress on CRF gene expression in the PVN, it can be hypothesized that the mechanisms controlling the HPA axis in CB2xP mice may be acting at the level of the synthesis or release of CRF. Pharmacological studies with agonists or antagonists acting on CB2r and the examination of additional elements at different levels of the HPA axis in WT mice may clarify the mechanisms involved in the interaction between CB2r and this stress axis.
The GABAergic system has been considered an important element regulating emotional states and, therefore, one of the most important therapeutic targets in the control of anxiety. In fact, the effects of stress on GABAA receptors may be relevant to better understanding the alterations and therapeutic outcome in a wide variety of psychiatric disorders, such as anxiety disorders, schizophrenia and depression. The GABAA receptor is a complex receptor formed by the assembly of 5 subunits out of 16 subunits that have been described (six α (α1–α6), four β (β1–β4), three γ (γ1–γ3, two splice variants; γ2short, γ2long), one δ, one ɛ, and one θ subunit (Whiting, 2003). Only certain subunit combinations can form functional receptors that reach the plasma membrane, including αβ and αβγ/∂/ɛ/θ combinations (Connolly et al., 1996; Gorrie et al., 1997; Kittler et al., 2000; Pritchett et al., 1989). Benzodiazepines, acting through their binding on the interface of the α and γ subunits of the GABAA receptor complex, are known to act as anxiolytics promoting the inhibitory actions of the GABA neurotransmitter in the central nervous system (Da Settimo et al., 2007). The activation of the GABAA receptors results in neuronal hyperpolarization via the opening of chloride-permeable ion channels. Recent studies suggest that GABAA receptors containing the α2 and γ2 subunits, enriched in corticolimbic structures, mediate the anxiolytic effect of benzodiazepines (Löw et al., 2000). The results of the present study clearly demonstrate that the administration of alprazolam, a well-known anxiolytic benzodiazepine, markedly increased the time in the lighted area in WT mice whereas it failed to produce any effect in CB2xP mice. The lack of this effect of benzodiazepine in mice overexpressing CB2r may be related to functional alterations occurring in GABAA receptor subunits. Indeed, GABAA α 2 and GABAAγ2 receptor subunit gene expression was increased in the hippocampus and amygdala of CB2xP mice. These findings suggest that overexpression of CB2r may produce functional alterations in the GABAA receptors of brain areas controlling emotional behaviours. In addition, the present results led us to speculate that CB2r may potentially be implicated in the regulation of the GABAergic system. It is possible that CB2r may modify GABAergic release from neuronal terminals. In fact, recent studies suggest the suppression of GABAergic inhibitory signalling in the hippocampus following the administration of the cannabinoid CB2r agonist JWH133. These effects could be blocked by prior administration of the cannabinoid CB2r antagonist AM630 (Morgan et al., 2009). Additional studies are needed to explain the exact nature of the interaction between CB2r and the GABAergic system and the molecular mechanisms responsible for these functional alterations in CB2xP mice.
In summary, the results presented here reveal that CB2r plays a pivotal role in the regulation of the neurobiology and treatment of anxiety-related behaviours. Overexpression of CB2r modulated anxiety behaviour and produced pronounced alterations in the HPA axis and GABAergic system. Taken together, these findings strongly support the role of CB2r in the regulation of anxiety disorders and indicate that this receptor could be a potential new key target in the treatment of different mood-related disorders. Further pharmacological studies with cannabinoid CB2 agonists or antagonists are necessary to explore the potential therapeutic use of cannabinoid CB2r.
Footnotes
Acknowledgements
We thank Patricia Rodríguez and Analía Rico for excellent technical assistance.
Funding statement
This research was supported by grants from the Ministry of Science and Innovation (SAF 2008–01106) and Ministry of Health (RETICS RD06/0001/1004 and PNSD 2007/061). MSGG is a predoctoral fellow of the Ministry of Science and Innovation (MICINN).