
Abstract
Nicotine enhances attentional functions. Since chronic nicotine exposure through smoking induces neuroadaptive changes in the brain at a structural and molecular level, the present functional MRI (fMRI) study aimed at investigating the neural mechanisms underlying visuospatial and sustained attention in smokers and non-smokers. Visuospatial attention was assessed with a location-cueing paradigm, while sustained attention was measured by changes in response speed over time. During invalid trials, neural activity within the basal forebrain was selectively enhanced in smokers and higher basal forebrain activity was associated with increased parietal cortex activation. Moreover, higher levels of expired carbon monoxide in smokers before scanning were associated with higher parietal cortex activation and faster responses to invalidly cued targets. Smokers showed a slowing of responses and additionally recruited an area within the right supramarginal gyrus with increasing time on task. Activity decreases over time were observed in visual areas in smokers. The data provide evidence for altered attentional functions in smokers as compared with non-smokers, which were partly modulated by residual nicotine levels and were observed at a behavioural level for sustained and at a neural level for spatial and sustained attention.
Introduction
The brain’s cholinergic neurotransmitter system emanating predominantly from neurons in the basal forebrain, the pontomesencephalon and the striatum, is critically involved in attentional functions. It has been shown that nicotine, which binds to the cholinergic nicotinic receptors in the brain, can enhance both visuospatial and sustained attention (see e.g. Hahn et al., 2009; Heishman et al., 2010; Mancuso et al., 1999; Newhouse et al., 2004; Rezvani and Levin, 2001). In non-smokers (Thiel et al., 2005; Thiel and Fink, 2008; Vossel et al., 2008) and smokers (Murphy and Klein, 1998; Witte et al., 1997) nicotine facilitates the detection of stimuli when attention has been misdirected by invalid spatial cues in the location-cueing paradigm (i.e. it speeds attentional reorienting in space) (Posner, 1980). Moreover, nicotine has been shown to reduce omission errors and response time (RT) variability in the Continuous Performance Test (CPT) (Levin et al., 1998) and the Rapid Visual Information Processing (RVIP) task (Lawrence et al., 2002) which both assess the ability to sustain attention over longer periods of time (see Heishman et al., 2010 for a review).
The combination of functional MRI (fMRI) with psychopharmacological challenges allows for the in vivo characterization of pharmacological modulations of task-related neural activity (Honey and Bullmore, 2004). Pharmacological fMRI studies in non-smokers have repeatedly shown that nicotine-induced faster reorienting to invalidly cued targets in the location-cueing paradigm is accompanied by reduced activity in parietal brain regions (Giessing et al., 2006; Thiel et al., 2005; Thiel and Fink, 2008; Vossel et al., 2008), presumably reflecting more efficient information processing under nicotine due to enhanced cholinergic signalling. In contrast, one study in smokers has reported opposite effects of nicotine (i.e. increased activity) in attention-related parietal brain areas during the RVIP task (Lawrence et al., 2002). This may point to different actions of nicotine in smoking and non-smoking subjects at the neural level.
Chronic nicotine exposure through cigarette smoking has been shown to induce neuroadaptive changes in the brain (see e.g. Ferrea and Winterer, 2009 for a recent review). For instance, it has been observed that grey matter volumes in prefrontal and cingulate brain regions are reduced in smokers as compared with non-smokers, which might reflect effects of chronic smoking (or alternatively predisposing traits that lead to smoking, or a combination of these factors (Brody et al., 2004)). At the receptor level, in vitro studies have shown that nicotinic acetylcholine receptors in the brain (in particular α2β4 and α7 subtypes) are increased and upregulated in response to chronic nicotine exposure or chronic smoking (Buisson and Bertrand, 2002; Perry et al., 1999; Staley et al., 2006).
Despite this evidence for smoking-related brain changes at an anatomical and receptor level, there is so far no clear evidence for performance differences between smokers and non-smokers in attentional tasks under drug-free conditions (Hahn et al., 2007, 2009; Lawrence et al., 2002), although performance decrements in smokers as compared with never-smokers have occasionally been reported (Foulds et al., 1996). These between-group differences may crucially depend on the smokers’ deprivation status (and hence on residual bodily levels of nicotine present in smokers, but not in non-smokers) (Parrott et al., 1996). For example, it has been shown that the time needed to reorient attention in the location-cueing paradigm increases with the duration of smoking abstinence and with salivary levels of the nicotine metabolite cotinine (Shirtcliff and Marrocco, 2003).
Furthermore, little is known about baseline differences in attention task-related neural activation patterns of smokers and non-smokers under drug-free conditions. Here, Lawrence et al. (2002) observed reduced activity in parietal brain regions in smokers as compared with non-smokers when smokers were nicotine-free for 2.5 hours.
Thus, the aim of the present study was twofold: first, it was intended to investigate visuospatial attention in the location-cueing paradigm as well as sustained attentional functions (as reflected in the ability to maintain a constant level of performance over time) in non-smokers and mildly deprived smokers. Here, both behavioural performance and brain activation patterns were recorded. Second, the relationship between these measures and residual nicotine levels (as indexed by levels of expired carbon monoxide (CO) before scanning) was examined to test for nicotine-dependent behavioural and neural effects in the group of smokers.
Materials and methods
Subjects
Twenty-five subjects gave written informed consent to participate in the study after having received detailed instructions and explanations regarding the study. The subjects were investigated in the context of a multi-session pharmacological fMRI study (ClinicalTrials.gov Identifier: NCT00618280). The analyses presented here focus on the first experimental session and on the placebo condition only as part of an interim analysis. The study was conducted in compliance with the ethical principles of the World Medical Association (Declaration of Helsinki) and was approved by the ethics committee of the Heinrich-Heine University Düsseldorf and the federal drug agency of Germany (Bundesinstitut für Arzneimittel und Medizinprodukte). All subjects were recruited from a large population-based database in Germany and had no history of medical, neurological or psychiatric illness or alcohol or drug abuse as assessed by a full medical interview and examination, routine laboratory tests, a drug screening test, an electrocardiogram and a standardized psychiatric interview (SCID) (First et al., 1995). Subjects were moreover excluded from the study if they met the following criteria: age below 18 or above 55 years, intake of medication, participation in other pharmacological studies, positive urine drug screening (cannabis, amphetamines, opiates and cocaine), drug abuse during the last 6 months; for women additionally: pregnancy, lactation. Alcohol abuse was additionally assessed with the Alcohol Use Disorder Identification Test (AUDIT) (Saunders et al., 1993). Smokers were required to have a score ≥4 in the Fagerström Test for Nicotine Dependence (FTND) (Heatherton et al., 1991), non-smokers were defined by ≤20 cigarettes per lifetime. Simultaneous EEG recordings were conducted during fMRI scanning. EEG data will not be reported in the current study. Two subjects had to be excluded from further analyses due to excessive movement or technical problems during scanning, respectively. Therefore, data from 23 subjects were analyzed. Of these, 13 subjects were non-smokers (7 females, 6 males, mean age ± standard deviation (SD): 32.1 ± 9.24 years). Ten subjects were smokers (6 females, 4 males, mean age ± SD: 36.3 ± 12.5 years; mean FTND score ± SD: 5.2 ± 1.6, range 4–8) who had smoked on average for 19.9 ± 12.8 years. The smokers reported to smoke on average 17 ± 6 (range from 7–25) cigarettes per day. Data from the AUDIT were available for 10 of the 13 non-smokers and for all smokers and revealed that the two groups did not significantly differ in their AUDIT score (non-smoker: mean ± SD: 4.6 ± 5.1; smokers: 2.9 ± 1.7; t(18) = 0.99, p = .333). With the exception of one ambidextrous subject in the group of non-smokers, all subjects were right-handed.
The subjects were allowed to smoke their last cigarette 2 hours before the start of the experiment. Subjects were not required to smoke, but were told that they were now given the last opportunity to have a cigarette. Craving was assessed with the Questionnaire on Smoking Urges (QSU) (Tiffany and Drobes, 1991). Levels of CO in the breath of the subjects were measured using a Micro 4 Smokerlyzer® (Bedfont Scientific Ltd.) 2 hours before the start of the experimental task in the MRI scanner (i.e. after they were given the last opportunity to smoke). Blood samples were collected for cotinine immunoassay measurements (DRI® Cotinine Assay, Microgenics, Passau, Germany).
Stimuli and experimental paradigm
We employed a location-cueing task with central cues (Posner, 1980). The stimuli were shown on a thin-film transistor (TFT) monitor behind the MR scanner and were presented to the subjects by means of a mirror-system. Viewing distance was approximately 245 cm. Subjects were presented with two horizontally arranged boxes (1° wide and 4° eccentric in each visual field). A central diamond (0.5° eccentric from the centre to the left and right edge of the diamond) was placed in between, serving as a fixation point. These elements constituted the baseline display and were shown on the screen throughout the experiment. Cues consisted of a 200 ms brightening of one side of the diamond depicting an arrowhead pointing to one of the peripheral boxes. After a variable cue-target interval of 700 or 1000 ms the cue was followed by the presentation of the target (a black diamond) appearing for 100 ms in one of the two lateral boxes. The cues were valid in 80% of the trials. Subjects were instructed to respond as quickly as possible to the target by a button press with the index finger of their right hand. Trials were presented in a randomized sequence, every 3200 ms. Approximately one third of the trials were ‘null events’ (Josephs and Henson, 1999) where the baseline stimulus was displayed, leading effectively to variable stimulus onset asynchronies (SOAs). In addition to validly and invalidly cued trials we included catch trials in which the cue was not followed by any target to prevent anticipatory responses to the cues. The experiment consisted of 284 trials (144 valid, 36 invalid and 14 catch trials), including 90 null events, and lasted for 15 minutes. Prior to performing the task in the MR scanner the subjects were informed about the different conditions and completed a short practice session.
Data acquisition
T2*-weighted echoplanar (EPI) images with blood oxygen level-dependent (BOLD) contrast (matrix size 64 × 64, voxel size 3.1 × 3.1 × 3.0 mm3, field of view (FOV) 200 mm × 200 mm, 90° flip angle) were obtained using a 3 T MRI System (Trio, Siemens, Erlangen, Germany). Additional high-resolution anatomical images (voxel size 1 × 1 × 1 mm3) were acquired using a standard T1-weighted 3D magnetisation prepared rapid gradient echo (MP-RAGE) sequence (176 sagittal slices, repetition time/echo time 2250/3.03 ms, matrix size 64 × 64, FOV 200 mm × 200 mm, 9° flip angle).
Four hundred and sixty-five EPI volumes of thirty-three 3 mm thick axial slices were sequentially acquired (repetition time 2.0 s, echo time 30 ms). The first five volumes were discarded to allow for T1 equilibration effects. The data were pre-processed and analyzed with Statistical Parametric Mapping software SPM5 (Wellcome Department of Imaging Neuroscience, London) (Friston et al., 1995). To correct for interscan movement, the images were spatially realigned to the first of the remaining 460 volumes and subsequently re-realigned to the mean of all images. Then the mean EPI image for each subject was computed and spatially normalized to the standard EPI template volume in SPM5. The resulting normalization parameters (which define the transformations that are necessary to move the subject’s data into the Montreal Neurological Institute (MNI) standard coordinate space) were subsequently applied to the individual EPI volumes as well as to the T1 scan, which had been coregistered to the mean of the realigned EPIs beforehand. The functional images were hereby transformed into standard stereotaxic space and resampled at 2 × 2 × 2 mm3 voxel size. The normalized images were spatially smoothed using an 8 mm full-width half-maximum (FWHM) Gaussian kernel to meet the statistical requirements of the General Linear Model and to compensate for residual macroanatomical variations across subjects.
Statistical analysis of imaging data
Data were analyzed with SPM5 employing a random effects model. Four regressors were defined at the single-subject level (validly and invalidly cued targets, catch trials and missed/incorrect responses). The event types were time-locked to the onset of the target by a canonical synthetic hemodynamic response function (hrf) and its first order temporal derivative. The six movement parameters of the realignment (rigid body translation in the x-, y- and z-plane as well as rotation around the x-, y- and z-axis) were included in the design matrix as additional regressors. Data were scan-wise globally scaled to reduce globally distributed confounding effects (Kiebel and Holmes, 2004) and high-pass filtered at 1/128 Hz.
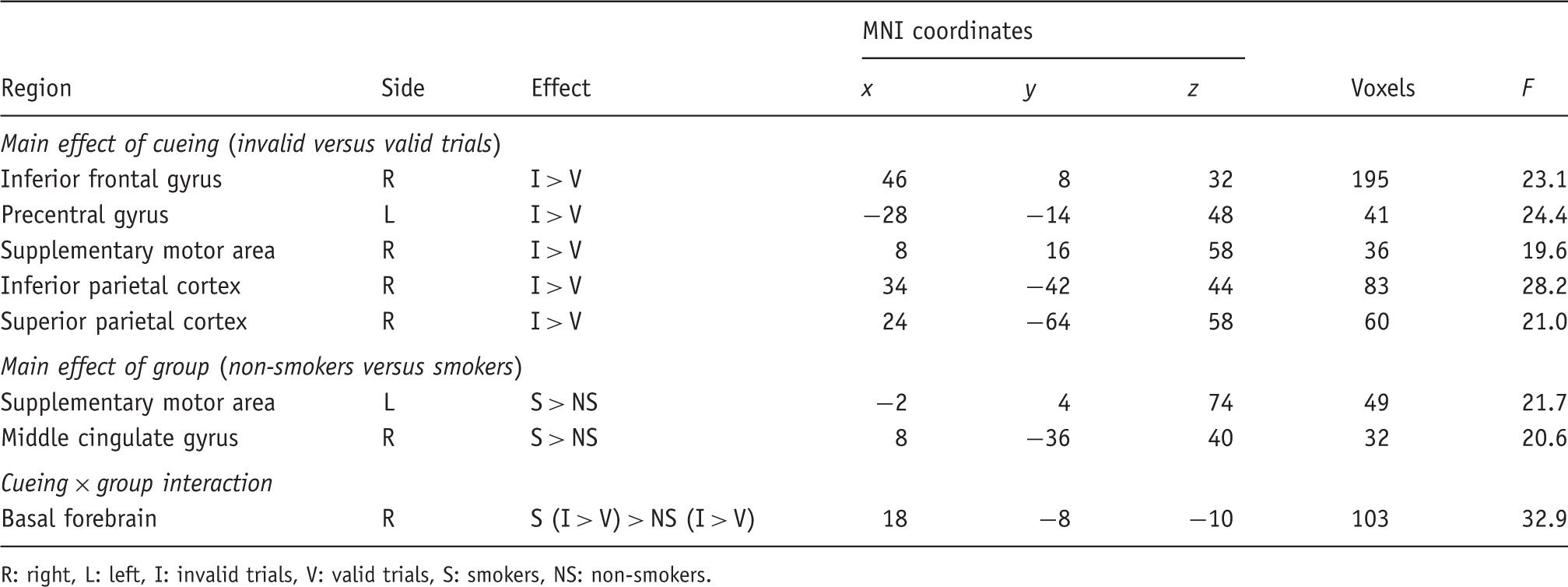
To investigate the neural correlates of spatial attention, two contrast images (valid and invalid trials vs. baseline) were created for each subject. These first-level contrast images were entered into a 2 × 2 ANOVA model with the within-subject factor ‘cueing’ (valid, invalid) and the between-subject factor ‘group’ (non-smoker, smoker). Although there were no significant differences in age (t(21) = −0.93; p = 0.362) or gender (χ2 = 0.87; p = 0.768), these variables were included as covariates in this analysis to account for the yet imperfect matching of the two groups. Inhomogeneity of variance and correlation of measurement were estimated with a Restricted Maximum Likelihood (ReML) algorithm. We tested for main effects of cueing and group, as well as for interaction effects with non-directional F-contrasts (see Figure 1). Activations are reported at a level of p < 0.001 (uncorrected). Since cluster-level inference is only implemented for t-, but not for F-contrasts in SPM (Poline et al., 1997), we employed a cluster extent threshold of more than 30 contiguous 2 × 2 × 2 mm3 voxels in order to minimize the chance of false positive findings.
(A) Neural activity related to attentional reorienting in the location-cueing paradigm (main effect of cueing). (B) Brain regions showing condition-unspecific activation differences between non-smokers and smokers (main effect of group). (C) Interaction effect of cueing and group. MNI x-, y- and z-coordinates for the activations are provided on top of the bar charts. Asterisks denote where beta parameters significantly differ from zero. v: valid; i: invalid; NS: non-smokers; S: smokers.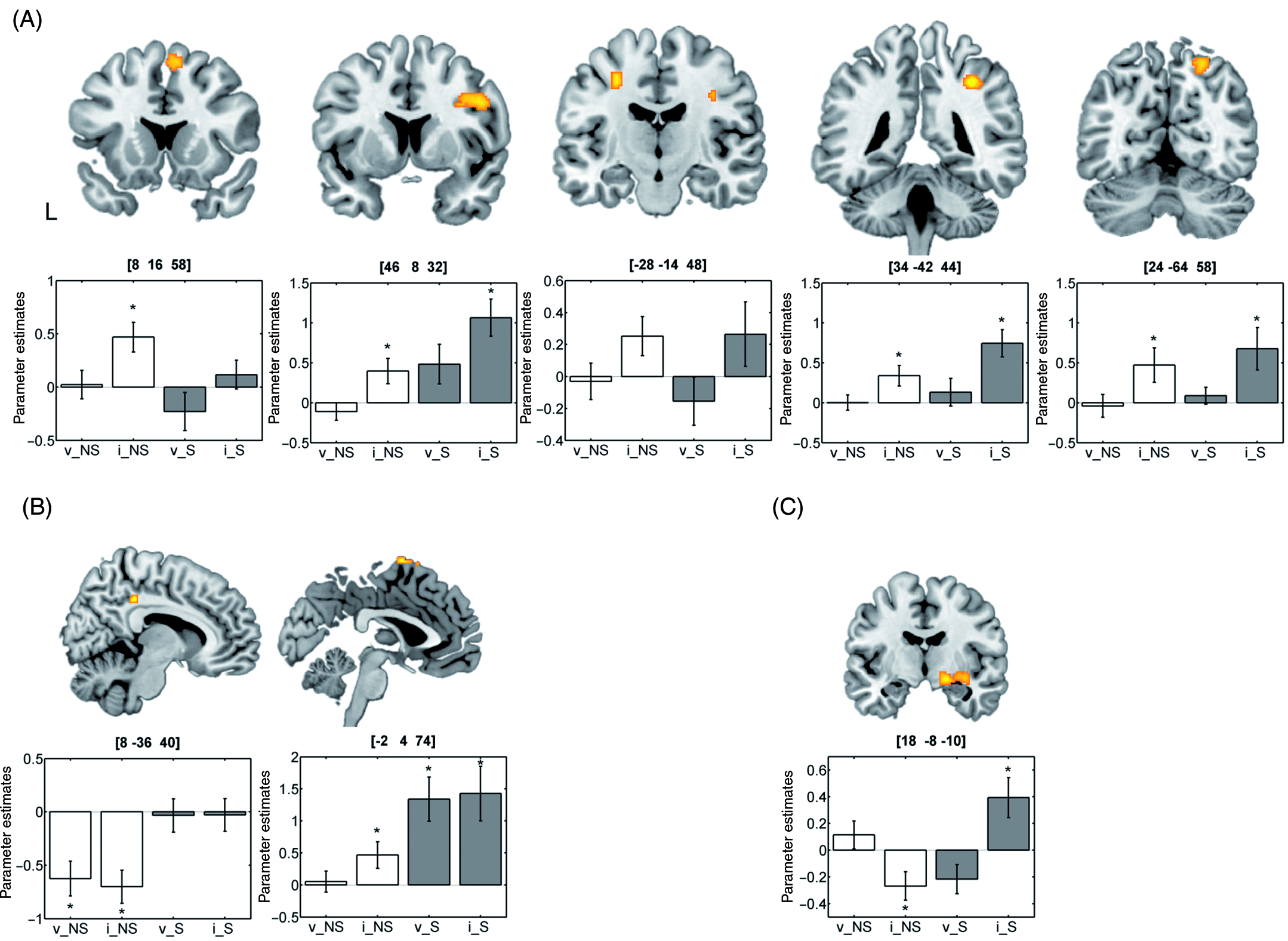
To test for linear increases and decreases of neural activity over the time of the experiment, we employed a time modulation of single-trial BOLD responses as implemented in SPM5. For this analysis, a parametric regressor was added for valid and invalid trials at the single-subject level which coded for a linear relationship of BOLD amplitudes and time (irrespective of experimental condition). Contrast images were created for this parametric time regressor for the individual subjects (one-zero contrast; time regressor vs. baseline) providing information in which voxels BOLD amplitudes are linearly associated with time. These contrast images were then entered into a second-level ANOVA with the covariates age and gender. We tested for differential effects between the two experimental groups with a non-directional F-contrast isolating brain regions that show stronger activity increases or decreases over time in smokers as compared to non-smokers. Activations from this analysis are reported at a level of p < 0.001 (uncorrected) and a cluster threshold of more than 30 contiguous voxels.
Since CO levels were significantly correlated with RTs to invalidly cued targets in smokers (see the Results section), a regression analysis between those values and neural activity during invalid trials was performed. A non-directional F-contrast was used to test for relationships between BOLD amplitudes and expired CO. Results from this analysis are reported at a significance level of p < 0.001 and a cluster threshold of more than 30 contiguous voxels. Post-hoc partial Pearson correlations (rp) were calculated for significant voxels to control for the effects of RT. Here, the voxel values (beta parameters) of the peak activation voxels were considered. Although no significant relationship between CO levels and the trial number regression weights on RTs were obtained (see below and Results section), we conducted exploratory analogical regression and correlation analyses for the parametric time regressor (significance level of p < 0.001 and a cluster threshold of more than 30 contiguous voxels).
For a further (post-hoc) analysis in the group of smokers, we tested for a relationship between basal forebrain activity (which was enhanced during invalid trials in smokers, see Results section) and the activity in other brain areas. Here, beta parameters of the regressor for invalid trials were extracted at the peak voxel showing the cueing × group interaction effect 1 for each individual smoking subject and entered as a regressor in a whole-brain random effects regression analysis. A non-directional F-contrast (thresholded at <0.001 and a cluster extent of more than 30 voxels) tested for associations between neural activity in the basal forebrain and other regions in the brain.
Statistical analysis of behavioural data
RTs faster than 100 ms (i.e. anticipated responses) (Luce, 1986) were excluded from the analysis. Median RTs were calculated separately for valid and invalid trials in each subject. Omissions were summed-up separately for valid and invalid trials and expressed as percentage values. For the ‘catch trials’, false alarm responses were determined and transformed into percentage values. To test for differences in visuospatial attention in non-smokers and smokers, median RTs and missed responses were analyzed with 2 × 2 ANOVAs with the within-subjects factor ‘cueing’ (valid, invalid) and the between-subject factor ‘group’ (non-smoker, smoker) (as well as age and gender as covariates). False alarm responses in catch trials were compared between both groups with one-way ANOVAs (including age and gender as covariates).
To assess the ability to sustain attention over the time course of the experiment, regression analyses with single-trial RTs (irrespective of condition) as dependent and trial number as independent variable were calculated in each individual subject. The resulting regression weight (beta coefficient) of the trial number variable for each subject was compared between non-smokers and smokers with a one-way ANOVA (and age and gender as covariates) 2 . In the smoker group, correlation analyses were performed between levels of expired CO and RTs, percentage of omissions and the time on task regression weights using Spearman’s rank coefficient (rs). All results are reported at a significance level of p < 0.05.
Results
Physiological and subjective measures
Cigarette craving was rated by the smoking subjects on a 7-point Likert-scale (1 (strongly disagree) to 7 (strongly agree)) in the QSU. Here, smokers rated their urge to smoke on average with 3.2 (±0.76 SD). Levels of CO in the subjects’ breath amounted on average to 17.3 ± 9.8 ppm in smokers (all values ≥ 4) and to 0.38 ± 0.9 ppm in non-smokers (CO = 0 in n = 10, CO = 1 in n = 2 and CO = 3 in n = 1 subject(s)). Blood cotinine levels amounted to 143 ng/ml ± 102.2 and cotinine levels were highly correlated with levels of expired CO in the group of smokers (rs = 0.94; p < 0.0001). None of the other correlations between levels of expired CO, blood cotinine levels, FTND score, cigarettes per day, and QSU scores reached significance. The data from three non-smokers could not be analysed for technical reasons, but cotinine levels amounted to 0 in all of the remaining non-smoking subjects (n = 10).
Behavioural data
Anticipations (RTs < 100 ms) and false alarms (catch trials) amounted to 0.00 ± 0.00% and 0.21 ± 0.36% in non-smokers versus 0.00 ± 0.00% and 0.22 ± 0.54% in smokers, respectively. No differences between the two groups were observed for anticipations and false alarm rates (ηp2 = 0.001). Missed responses in valid trials amounted to 2.24 ± 1.93% in non-smokers and to 4.58 ± 4.29% in smokers. In invalid trials, non-smokers and smokers missed 2.35 ± 2.49% and 4.72 ± 7.75% of the targets, respectively. There was a trend towards a higher percentage of missed responses in smokers as compared with non-smokers (main effect of group: F(1,19) = 3.871; p = 0.064; ηp2 = 0.169) as well as a trend towards a higher proportion of missed responses in invalid as compared with valid trials (main effect of cueing: F(1,19) = 3.864; p = 0.064; ηp2 = 0.169). The cueing × group interaction did not reach significance (ηp2 = 0.012). Averaged median RTs ± SDs in valid and invalid trials were 340.83 ± 44.8 ms and 383.52 ± 48.7 ms in non-smokers versus 337.75 ± 50.0 ms and 391.19 ± 70.0 ms in smokers, respectively. The 2 (valid, invalid) × 2 (non-smoker, smoker) ANOVA revealed a significant main effect of cueing (F(1,19) = 8.752; p = 0.008) indicating slower responses to invalidly as compared with validly cued targets. Neither the main effect of group nor the group × cueing interaction effect reached significance (ηp2 = 0.001 and 0.056, respectively).
Within the group of smokers, CO levels were significantly correlated with the percentage of missed responses in invalid trials (rs = −0.67; two-sided p = 0.034) and by trend also with RT to invalidly cued targets (rs = −0.62; two-sided p = 0.056) (see Figure 2A). No significant correlations with missed responses or RT in valid trials were obtained. When levels of cotinine were considered, significant correlations were observed for both behavioural measures (misses in invalid trials: rs = −0.68; two-sided p = 0.03; RT invalid trials: rs = −0.83; two-sided p = 0.003). No significant relationship between behavioural performance and any of the other smoking-related variables (FTND, QSU, and cigarettes per day) were observed.
(A) Relationship between CO levels and reaction times in invalid trials as well as missed responses in the group of smokers. (B) Results of the regression analysis of expired CO levels before scanning and BOLD amplitudes in invalid trials.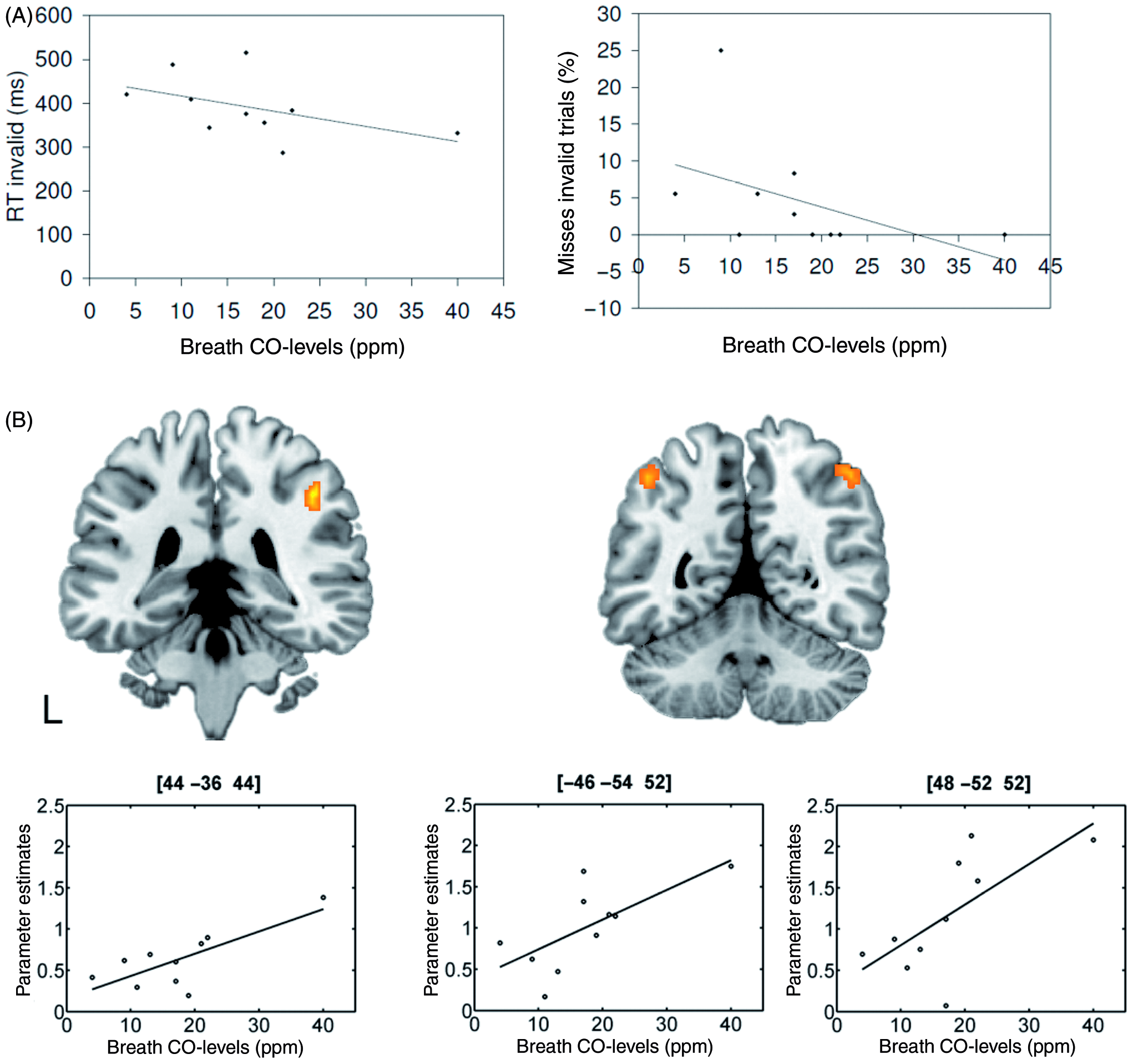
The ANOVA on the trial number regressor (coding for linear increases or decreases in RTs across the time of the experiment) revealed higher values for smokers than for non-smokers (F(1,19) = 4.423; p = 0.049). In particular, negative values close to zero (mean ± SD: −0.082 ± 0.43) indicating constant or even slightly reduced RTs with longer time on task were observed for non-smokers, while smokers showed positive values reflecting an increase in RTs towards the end of the experiment (0.307 ± 0.57) (see Figure 3A). In smokers, the correlation between CO levels and the trial number regressor did not reach significance (rs = −0.395; two-sided p = 0.260; for cotinine levels: rs = −0.43; two-sided p = 0.214). The trial number regressor was also not significantly related to other smoking-related variables such as the FTND score or the number of cigarettes per day, while a trend towards a significant positive correlation was observed with the QSU scores (rs = 0.59; two-sided p = 0.074).
(A) Mean response times and standard deviations over the time course of the experiment for non-smokers (left) and smokers (right). Note that differences in the slope of the trial number regressor were statistically significant. (B) Brain areas where neural activity decreased (blue) or increased (orange) with time in smokers more than in non-smokers.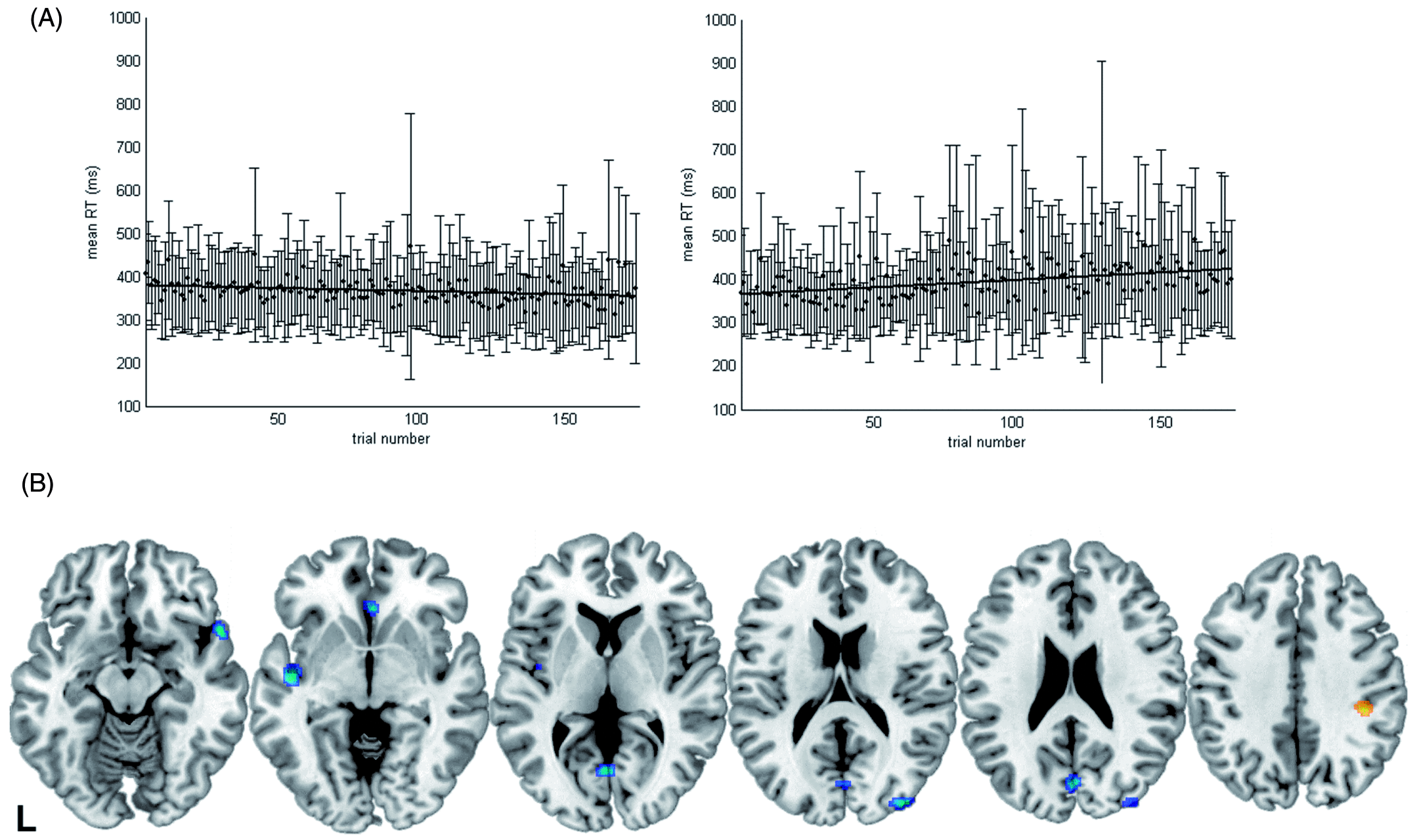
Neural data – spatial attention
Results of the cueing × group ANOVA on the fMRI data
R: right, L: left, I: invalid trials, V: valid trials, S: smokers, NS: non-smokers.
The contrast of invalidly versus validly cued targets (main effect of cueing) revealed activation within fronto-parietal brain regions predominantly in the right hemisphere (see Table 1 and Figure 1A). In all regions, neural activity was higher in invalid as compared with valid trials. Two brain areas (within the left supplementary motor area (SMA) and the right posterior middle cingulate gyrus) showed a main effect of group indicating a condition-unspecific difference in neural activity in the two experimental groups (see Table 1). While the SMA was more strongly activated, the posterior middle cingulate gyrus was less deactivated in smokers than in non-smokers (see Figure 1B). An interaction effect was observed within the right basal forebrain. Here, activity was enhanced for invalid as compared with valid trials in smokers, but not in non-smokers (see Figure 1C).
The regression analysis between neural activity in invalid trials and levels of expired CO before scanning revealed three regions within left and right parietal cortex where BOLD amplitudes were positively correlated with CO values (right angular gyrus: x = 48, y = −52, z = 52; F = 53.82; 142 voxels; left angular gyrus: x = −46, y = −54, z = 52; F = 62.94; 39 voxels; right supramarginal gyrus: x = 44, y = −36, z = 44; F = 77.28; 42 voxels; see Figure 2B). All three regions also showed significant correlation with blood cotinine levels. No negative correlations were observed. Post-hoc partial correlation analyses between the beta parameters (as indicators of BOLD amplitudes) and the CO values showed that the relationship persisted in the left angular gyrus (rp = 0.69, t(7) = 2.54, two-sided p = 0.039) and the right supramarginal gyrus (rp = 0.69, t(7) = 2.52, two-sided p = 0.04), but not in the right angular gyrus (rp = 0.59, t(7) = 1.95, two-sided p = 0.092) when the effect of RTs in invalid trials (which significantly correlated with CO levels) was controlled.
Neural data – sustained attention
Results of the time modulation comparison of neural activity in smokers versus non-smokers
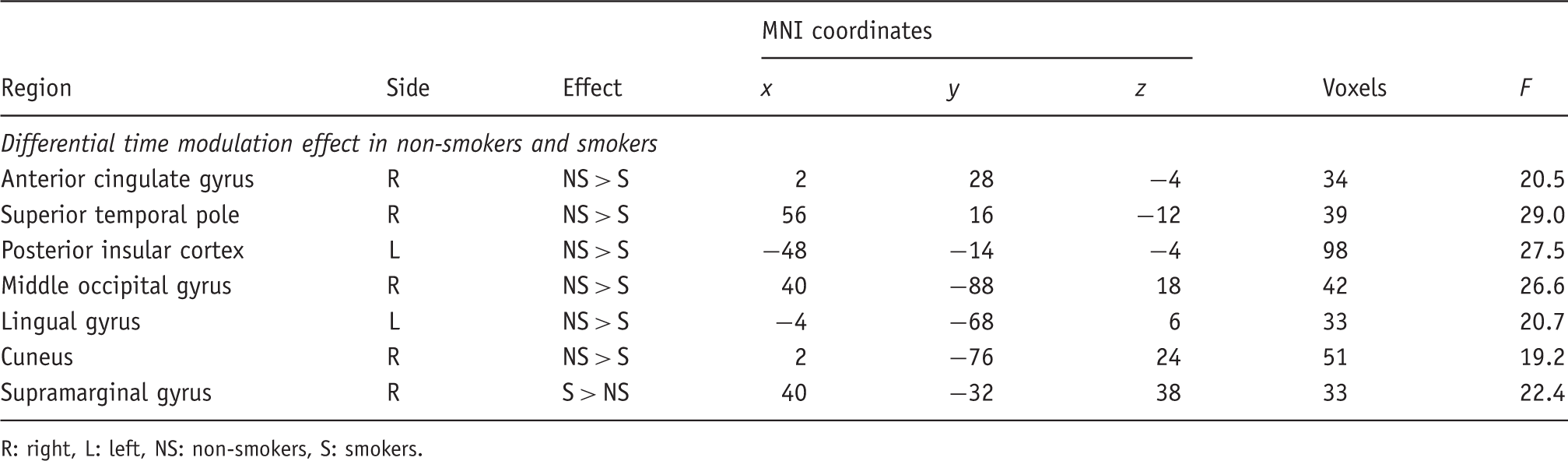
R: right, L: left, NS: non-smokers, S: smokers.
The regression analysis with expired levels of CO in smokers did not yield any significant results.
Post-hoc regression analysis in smokers
The post-hoc regression analysis testing for a relationship between the activity in the basal forebrain and the activity in other brain areas during invalid trials yielded significant correlations with (1) neighbouring regions of the basal forebrain (x = 18, y = −10, z = −10; F = 222.58; 52 voxels), (2) the right insular cortex (x = 42, y = 12, z = −2; F = 71.05; 74 voxels), and (3) the right inferior parietal cortex (x = 48, y = −58, z = 42; F = 74.18; 121 voxels) (see Figure 4). Hence, activity in these areas was higher in smokers with high basal forebrain activity during invalid trials.
Positive relationship between basal forebrain response and neural activity in the right inferior parietal cortex in invalid trials in smokers. BF: basal forebrain, IPL: inferior parietal lobe.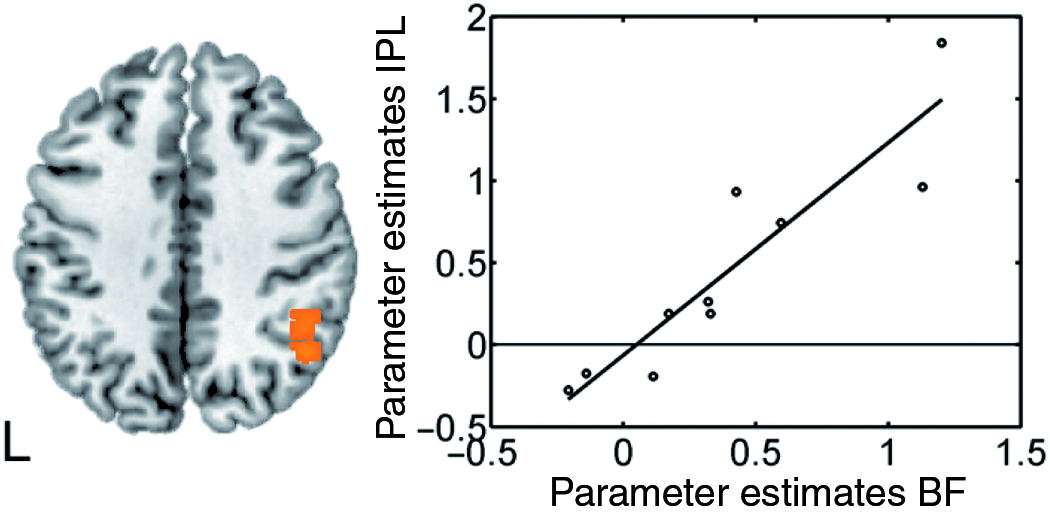
Discussion
In the present study behavioural and neural correlates of spatial and sustained attention were compared between smoking and non-smoking subjects. Related to attentional reorienting smokers did not show different RTs from non-smokers. However, the basal forebrain was more strongly recruited during invalid as compared to valid trials by smokers. Higher levels of expired CO were associated with faster RTs and higher neural activity within the bilateral parietal cortex in invalid trials in smokers. Sustained attention was reduced in smokers as compared with non-smokers as reflected in a reduced ability to maintain a constant performance level throughout the time course of the experiment. The latter effect was accompanied by both stronger reductions in neural activity in brain areas such as, for example, the visual cortex, as well as with a stronger recruitment of the right supramarginal gyrus with increasing time on task in smokers as opposed to non-smokers. Higher CO levels reduced the percentage of omissions in invalid trials, but did not significantly affect the RT slowing with increasing time on task in smokers. In sum, the current findings provide evidence for altered attentional functions in smokers which are evident at a behavioural level for sustained and at a neural level for both spatial and sustained attention.
Spatial attention
Reorienting visual attention in space activated a right-lateralized fronto-parietal network as in previous fMRI studies employing location-cueing paradigms (Giessing et al., 2006; Thiel et al., 2004, 2005; Vossel et al., 2006, 2008). These regions were likewise recruited by smokers and non-smokers. However, smokers showed less deactivation in the posterior part of the middle cingulate cortex during the task irrespective of cueing condition. This region – which was located at the border to the posterior cingulate cortex – has been implicated in the default network of the brain (see e.g. Fox and Raichle, 2007 for a review), since it usually shows decreases in activity during goal-directed actions when contrasted with passive viewing conditions. Moreover, deactivation in this region has been related to relatively faster response times, while reduced deactivation is associated with momentary lapses of attention (Weissman et al., 2006). It has also been shown that nicotine administration in smokers further deactivates areas of the default network during a visuospatial attention task (Hahn et al., 2007). The finding that this area was less deactivated during task performance in the present study may thus indicate less effective suspension of task-irrelevant processes in mildly deprived smokers.
Interestingly, differential reorienting-related activity between the two groups (i.e. a cueing × group interaction effect) was observed in the right basal forebrain where neural activity was enhanced in invalid as compared with valid trials in smokers, but not in non-smokers. The localization of this region within the basal forebrain was confirmed by cytoarchitectonic maps of this area according to which the activation cluster in the present study lay within a sphere of 5 mm from the centre of gravity of the Ch4 compartment which incorporates the nucleus basalis of Meynert (Mesulam et al., 1983; Zaborszky et al., 2008). The basal forebrain provides the major cholinergic input to the neocortex and has been related to signal detection and top-down attention on the basis of animal data (see Sarter et al., 2005 for a review). In particular, lesions of the basal forebrain result in slowed responses to invalidly cued stimuli in rats (Bushnell et al., 1998) and monkeys (Voytko et al., 1994). It was proposed that the corticopetal projections that arise from the basal forebrain influence the activity of frontal and parietal areas during attentional tasks (Sarter et al., 2001, 2005). In the present study, no significant activation differences between smokers and non-smokers were observed in the fronto-parietal reorienting network, although Lawrence et al. (2002) reported less attention task-induced activation in parietal cortex in smokers under placebo conditions. Our data thus suggest that the basal forebrain might have enhanced the activity in fronto-parietal areas in smokers during reorienting foreclosing activity differences to non-smokers. This assumption was tested with a post-hoc regression analysis. Although this analysis does not provide any information on the directionality of the effects or on the possible role of a third modulatory variable, it was observed that smokers with higher basal forebrain activity during invalid trials tended to show higher activation of the right inferior parietal cortex.
Parietal cortex activity during invalid trials in smokers was also enhanced with higher levels of expired CO before scanning resulting in faster responses to invalidly cued targets. If the relationship between basal forebrain and parietal cortex proved true, basal forebrain activation should likewise be associated with higher CO levels and parietal activity, accordingly. A post-hoc partial correlation analysis between CO levels and the basal forebrain response in invalid trials (at the voxel identified by the interaction effect of the ANOVA), controlling for the effect of RT in invalid trials, revealed a positive, but non-significant, relationship (rp = 0.55; t(7) = 1.75; two-sided p = 0.12). In sum, these effects suggest an additional recruitment of the basal forebrain in smokers during attentional reorienting which possibly counteracts differences in RTs and parietal cortex activity to non-smokers and which might depend on (or be triggered by) residual bodily levels of nicotine.
It has to be noted that the positive correlation between CO levels (as an indicator of bodily nicotine levels) and the parietal cortex response is directly opposed to the effects of nicotine administration on reorienting-related parietal activation in non-smokers (Giessing et al., 2006; Thiel et al., 2005; Thiel and Fink, 2008; Vossel et al., 2008). Here, nicotine reduces BOLD amplitudes in parietal areas during invalid trials. Thus, smokers and non-smokers seem to respond differentially to nicotinic stimulation. Possibly, the underlying mechanisms also differ, so that an enhancement of nicotine levels above the normal level in non-smokers increases information processing efficiency, whereas it prevents an attentional decline in nicotine-deprived smokers. Further evidence for enhanced attention-related parietal activity in response to nicotine in smokers is provided by the study of Lawrence et al. (2002). Here, neural activity within the bilateral parietal cortex was increased in response to nicotine administration in the sustained attention, but not in the control task in smokers.
Sustained attention
When the stability of RTs over the time course of the experiment was considered, smokers performed worse than non-smokers in that they showed an increase in RTs with increasing time on task. Hence, the ability to sustain attention over longer time periods seemed to be compromised in mildly deprived smokers. Here, the amount of missed responses in invalid trials was negatively correlated with CO levels, while no significant relationship was observed for RT increases with time. Stronger activity decreases with time than in non-smokers were observed in multiple brain areas and amongst others in the visual cortex. It could be speculated that this effect reflects difficulties in maintaining the cue-induced top-down modulation of sensory areas over time in smokers leading to the observed RT slowing. In contrast, an area within the right supramarginal gyrus was more strongly activated in smokers than in non-smokers with increasing time on task. This region – which has previously been related to sustained attention by multiple studies (see Singh-Curry and Husain, 2009 for a meta-analysis and review) – might have been more strongly recruited with increasing time on task and hence with increasing demands to maintain an attentive state in smokers.
Limitations of the study
The sample sizes in the present study were relatively small, so that the findings and particularly the correlative relationships in the group of smokers need to be replicated with larger groups. Moreover, the current study relied on levels of expired CO as an indirect indicator of bodily nicotine levels and plasma nicotine levels were not assessed. However, CO levels were highly correlated with blood levels of the nicotine metabolite cotinine and the association with behavioural and fMRI measures was consequently also observed when blood cotinine levels were considered. Moreover, previous studies have shown that levels of expired CO are highly correlated with blood nicotine levels (Herning et al., 1983; Haustein et al., 2003; Jarvik et al., 2000).
Smokers in the current study were abstinent for 2 hours prior to performing the task in the MR scanner which may have induced mild withdrawal effects. Hence, although higher CO levels are indicative of higher nicotine levels (which may retard withdrawal), they might also be related to higher smoking rates and higher nicotine dependency and thus to higher withdrawal effects in smokers. In the following we would like to speculate how the findings of the present study are related to effects of nicotine or withdrawal due to smoking deprivation, respectively. If the observed findings were mediated by higher nicotine dependency and higher withdrawal, we would expect correlations of the behavioural and fMRI measures with smoking-related variables such as the FTND score, the amount of cigarettes smoked per day, or the amount of craving assessed with the QSU. Furthermore, QSU scores in deprived smokers are usually higher than in the current study (Tiffany and Drobes, 1991; Morgan et al., 1999). In contrast to CO and cotinine levels, none of these smoking-related variables was significantly related to the reported neural and performance effects in the present study. The only possible exception to this was the measure of sustained attention, where we observed a trend towards a significant correlation with the QSU scores. Thus, the performance decrement over the time of the experiment might have been affected by beginning withdrawal in the group of smokers.
Conclusion
In the present study smokers and non-smokers showed function-specific differences in attentional processing which were evident at behavioural and neural levels and were partly modulated by residual nicotine levels in the group of smokers. During shifts of spatial attention, activity within the basal forebrain was enhanced and correlated with parietal cortex activation in smokers. Smokers showed a reduced ability to maintain a constant level of performance throughout the experiment, as well as differential increases and decreases in neural activity with time on task when compared with non-smokers. Moreover, both task performance and brain activation patterns were critically affected by residual nicotine levels as reflected by levels of expired CO in smokers.
Footnotes
Acknowledgements
We are grateful to our colleagues from the MR and Cognitive Neurology sections for valuable support and discussions.
Funding
This work was supported by the German Research Foundation (DFG, Deutsche Forschungsgemeinschaft), conducted within the framework of the Priority Program SPP1226: ‘Nicotine: Molecular and Physiological Effects in CNS’ (grant number Wi1316/7–1).
Conflict of interest statement
GW has received sponsorship to attend scientific meetings, speaker honorariums and consultancy fees from pharmaceutical companies (Johnson and Johnson/Janssen Pharmaceutica, Janssen Cilag, Pfizer Inc., AstraZenca) and GRF received honoraria from GSK and Teva as a lecturer which do not constitute a potential conflict of interest with the presented work.
1
Note that this voxel was excluded from the analysis by a mask, since here the activity perfectly correlates with the input regressor.
2
The same analyses were repeated with a separate modelling of trial number, condition (valid; invalid) and the interaction effect, to assess whether RTs to validly and invalidly cued targets differentially varied with increasing time on task between the two groups. Here, no significant interactive effects were observed, so that the condition factor was also not considered in the time modulation analyses of BOLD signals.