
Abstract
Reduced functioning of the dopamine transporter (DAT) has been linked to bipolar disorder (BD). Mice with reduced DAT functioning (knockdown, KD) exhibit a behavioral profile in the mouse Behavioral Pattern Monitor (BPM) consistent with patients with BD mania in the human BPM. Patients with BD also exhibit increased risk taking, which can be quantified using the Iowa Gambling Task (IGT). We hypothesized that DAT KD mice would exhibit increased risk-taking behavior in a novel mouse version of the IGT. DAT KD and wildtype (WT) littermates were trained in the mouse IGT. In session 1, KD mice initially made riskier choices, but later performed comparably to WT mice. Once trained to stable choice performance, DAT KD mice continued to exhibit a trend to choose the riskier options more than WT mice. Finally, we confirmed that these DAT KD mice also exhibited an exploratory profile in the BPM consistent with patients with BD mania, where risky choice behavior modestly correlated with specific exploration. These data demonstrate that DAT KD mice chose the riskier options more than WT mice, providing further support for the use of DAT KD mice as a model of BD mania.
Introduction
Bipolar disorder (BD) is a prevalent and debilitating psychiatric disorder, affecting 1–5% of the population from BD-I to BD-II (Merikangas et al., 2007). The prognosis of patients with BD remains poor, with 15% committing suicide, despite treatment options being available. This poor prognosis may be because treatments for BD have been discovered serendipitously, or were originally designed for other psychiatric disorders (Gould and Einat, 2007). Most current models of BD derive from observations of similarities as opposed to a rationale based on an understanding of the neuropathology of BD. For example, the amphetamine administration model of mania was first used based upon the observed behavior of rats after drug administration, not as an a priori hypothesis related to the neurobiological underpinnings of the disorder (Davies et al., 1974; Randrup and Munkvad, 1974; Rushton and Steinberg, 1963). We proposed a model of mania based on the putative reduced functioning of the dopamine transporter (DAT) in patients with BD. The DAT has been implicated in the neuropathology of BD via genetic linkage studies (Greenwood et al., 2001, 2006; Kelsoe et al., 1996), with lower DAT levels (Amsterdam and Newberg, 2007) and reduced DAT expression (Horschitz et al., 2005) being reported in patients with BD. Consistent with these neuropathological observations, mice with reduced functioning DAT levels (via either genetic or pharmacological manipulation) exhibit a profile in the mouse Behavioral Pattern Monitor (BPM) that is consistent with that of acutely manic BD patients in a human BPM (Perry et al., 2009; Young et al., 2010a, 2007b), and is exacerbated by subthreshold psychostimulant administration (Young et al., 2010b). The BPM quantifies exploration in animals across species (Geyer et al., 1986; Perry et al., 2009; Risbrough et al., 2006) along three separable factors of activity levels (diversive exploration), investigatory behavior (specific exploration), and patterns of movement (Paulus and Geyer, 1993). The abnormal exploratory behavior of patients with BD mania includes increased activity levels combined with increased specific exploration and abnormal patterns of movement as reflected in reduced spatial d (Perry et al., 2009). Most models of BD utilize hyperactivity as their only outcome measure (Einat, 2006). In contrast, the use of the BPM has revealed that hyperactivity is accompanied by increased specific exploration and reduced spatial d both in patients with BD mania and in DAT knockdown (KD) mice (Perry et al., 2009; Young et al., 2007b, 2010a). These DAT KD mice exhibit only 10% DAT levels when compared with wildtype (WT) mice (Zhuang et al., 2001). Thus the BPM may provide a behavioral profile by which novel treatments can be developed for mania that affect global behavior (see below) and not simply increased activity.
Mania is the cardinal feature of BD, as exemplified by the fact that it is a core symptom in the diagnosis of both Type I and Type II BD according to the DSM IV. While the BPM has proven to be a valuable tool to establish novel animal models of BD mania with cross-species translational validity (Perry et al., 2009; Young et al., 2007b, 2010a), the BPM only assesses exploratory behavior in a novel environment. Other aspects of mania have been identified and have been quantified utilizing neurocognitive tests with real-world translational validity. Such tests include the Iowa Gambling Task (IGT) developed by Bechara et al. (1994), which can quantify (Clark et al., 2001) the impulsive gambling trait of patients with BD that can be so detrimental to these patients (Kim et al., 2006). In brief, during the IGT the subject is asked to repeatedly choose from four decks of cards from which they may be rewarded (gain) or punished (loss). Two decks provide large immediate rewards but larger punishment, while the other two offer smaller rewards but smaller punishment. Across a series of 100 trials, healthy subjects tend to select cards from the two decks that do not provide large rewards, but also offer the lowest punishment. Patients with BD mania take longer to begin selecting from these two ‘safer’ decks (Clark et al., 2001). The IGT has provided a means by which to identify the putative cognitive construct underlying gambling behavior in BD mania (Adida et al., 2008; Yechiam et al., 2008), as well as to examine the relationship of gambling behavior to other aspects of mania (Christodoulou et al., 2006).
Given the real-world implications of assessing gambling behavior in the IGT (Buelow and Suhr, 2009), several researchers have attempted to create an animal version of the IGT (Homberg et al., 2008; Rivalan et al., 2009; van den Bos et al., 2006; Zeeb et al., 2009). These tasks have begun to be used to investigate the neurobiological underpinnings of performance in the task, including dopaminergic (Zeeb et al., 2009) and serotonergic (Homberg et al., 2008; Zeeb et al., 2009) manipulations, as well as relationships to other behaviors (Rivalan et al., 2009). We have utilized the strategy developed by Zeeb et al. (2009) to determine whether our mouse model of BD mania exhibits risk-taking behavior consistent with that of BD mania. We also examined the exploratory behavior of the same cohort of DAT KD mice in the BPM. We hypothesized that DAT KD mice would: (a) exhibit a preference for the riskier options; (b) exhibit a BD mania-like profile in the BPM; and (c) that risk preference would correlate with hole-poking, given that increased specific exploration is one of the most prominent features of abnormal BPM exploration in manic BD patients (Perry et al., 2009, 2010).
Methods
Animals
Female DAT KD and WT littermate mice (n = 15 per group) were trained in the mouse IGT. The mice were generated by inserting modified embryonic stem cells of the 129Sv/J mouse strain in C57BL/6J blastocysts. Alteration of these stem cells was detailed by Zhuang et al. (2001). DAT heterozygous breeders were sent to our laboratory from Columbia University. All the mice used resulted from a breeding colony in the vivarium at the University of California San Diego (UCSD). The DAT KD and WT mice were approximately 3–5 months old at the time of first training and weighed between 15–26 g (average 20.3 ± 0.6 SEM). No difference in weight was observed between the two genotypes.
All animals were group housed (where possible, maximum four per cage), maintained in a temperature-controlled vivarium (21 ± 1°C) on a reversed day–night cycle (lights on at 8.00 PM, off at 8.00 AM), and were tested during the dark phase of the cycle. All mice had ad libitum access to water, were food-restricted, and maintained at 85% of their free-feeding weight during the periods of testing as described below. Mice were brought to the testing area 45 min before testing between 2.00 PM and 5.00 PM. All procedures were approved by the UCSD Institutional Animal Care and Use Committee. The UCSD animal facility meets all federal and state requirements for animal care and was approved by the American Association for Accreditation of Laboratory Animal Care.
Apparatus
Mice were trained and tested in 16 five-hole operant chambers (25 cm × 25 cm × 25 cm, Med Associates Inc., St. Albans, VT, USA). Each chamber consisted of an array of five square holes (2.5 cm × 2.5 cm × 2.5 cm) arranged horizontally on a curved wall 2.5 cm above the grid floor with, on the opposite panel, a food-delivery magazine (Lafayette Instruments, Lafayette, IN, USA) at floor level and a houselight near the ceiling. The chamber was enclosed in a sound-attenuating box, ventilated by a fan that also provided a low level of background noise. An infrared camera installed in each chamber enabled the monitoring of performance during training and testing. The animals were trained to respond with a nose-poke to an illuminated LED recessed into the holes. Infrared beams, mounted vertically and located 3 mm from the opening of the hole, were used to detect the responses. The food-delivery magazine opposite to the middle hole contained a well in which liquid reinforcement utilized in the form of strawberry milkshake (Nesquik® plus non-fat milk, 30 µL) was delivered by a peristaltic pump (Lafayette Instruments, Lafayette, IN, USA). An infrared beam mounted horizontally, 5 mm from the floor and recessed 6 mm into the magazine, was used to detect the magazine entries. The control of stimuli and recording of responses were managed by a SmartCtrl Package 8-In/16-Out with additional interfacing by MED-PC for Windows (Med Associates Inc., St. Albans, VT, USA) using custom programming.
Behavioral handling and training
The mice were given a daily ration of rodent chow such that body weight was maintained at 85% of their free-feeding weight. After approximately 3 days of weighing and thus handling the mice, they were around 85% of their free-feeding weight. The day prior to the initiation of training, mice were acclimated to the food reinforcement by an overnight exposure to the strawberry milkshake in the absence of water. On training days 1–3, mice were placed in the five-hole boxes for 10 min, during which liquid reinforcement was dispensed every 15 s into the well of the magazine, while the magazine was lit (Hab1). Entry into the magazine caused the light to go out until the next reinforcement was delivered. The entries were taken as evidence of learning to associate the lit up magazine with the liquid food reinforcement. At the end of each session the wells were inspected to ensure no liquid was present. On day 4, in order to obtain reinforcement, mice were required to nose-poke in any one of the four lit holes at the side of the chamber opposite to the magazine (central hole was never lit to maintain four options for responding; maximum of 120 trials were possible). This training session (Hab2) was repeated daily (Monday–Friday) until mice were able to make 40 responses to the light cue within a 30 min session on two consecutive days (days 4–18). Upon attainment of criterion, mice were trained in Hab2 on Tuesday and Friday only in order to maintain responding while training continued for other mice. This methodology has been successfully utilized in the past to avoid over-training on what is an intermediary stage (Young et al., 2004, 2007a, 2009a). Failure to attain this criterion resulted in removal from further analyses.
Mouse Iowa Gambling Task
For the actual mouse IGT sessions, we utilized the protocol as described previously for rats (Zeeb et al., 2009). The first 3 days, mice were tested by means of a forced-choice version of the mouse IGT to acquaint the mice with the different reinforcement and punishment schedules associated with each hole. In brief, only one of the four holes was illuminated randomly, and after nose-poking this hole the appropriate reinforcement and/or punishment schedule followed. For both the mouse IGT program as well as for its forced-choice version two different forms were used to counterbalance for possible hole preferences.
The gain–loss structure in the human and mouse Iowa Gambling Tasks
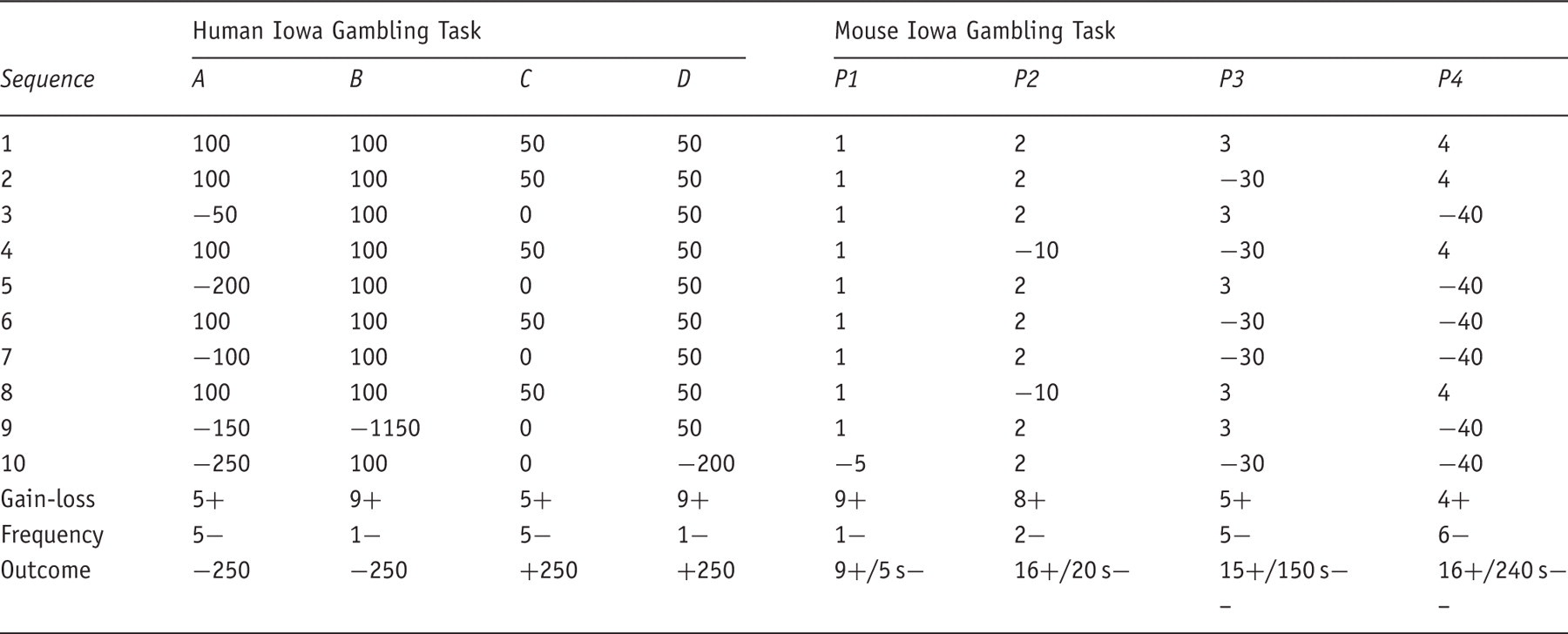
If the mouse was rewarded, the magazine light illuminated and delivered the appropriate level of liquid food reinforcement. When the mouse nose-poked to receive the food reward, a new trial was initiated automatically. If, on the other hand, a punishment occurred, no reward was given and a punishing time-out was triggered. The punishing time-out consisted of the light stimulus of the chosen hole starting to flash at a frequency of 0.5 Hz for the duration of the time-out period, during which all apertures were unresponsive. After the time-out period, the flashing light was extinguished and the magazine light lit up, giving the mouse the opportunity to start a new trial. Repeated nose-poke responses at the hole the mouse had just chosen were counted as ‘perseverative’ responses. While these were recorded, they were not punished in any way. Repeated nose-pokes in the same hole when rewarded were counted as ‘perseverative rewards’ while when punished were recorded as ‘perseverative punishments’. Finally, the times taken to choose a hole (mean choice latency; MCL) and obtain reward (mean reward latency, MRL) were also recorded. As to the different response options, data were grouped by advantageous (P1 and P2) and disadvantageous options (P3 and P4) and were measured as a percentage of the advantageous choices (P1 + P2)/(P1+P2+P3+P4)*100. Mice were trained continuously on the mouse IGT until they exhibited stability in their preference (no main effect of day when analyzed over four consecutive days).
Mouse Behavioral Pattern Monitor
Spontaneous locomotor and exploratory behavior was examined in 10 mouse BPM chambers as described previously (Risbrough et al, 2006). Each chamber consists of a 30.5 × 61 × 38 cm area with a Plexiglas holeboard floor equipped with floor holes in the front, middle, and rear parts of the floor and eight wall holes (three along each of the long walls, and two holes in the front and back walls). Hole-poking behavior was detected via infrared photobeams located at each hole. The location of the mouse was monitored from a grid of 12 × 24 infrared photobeams 1 cm above the floor (2.5 cm apart along the length and the width of the chamber; 24 × 12 X–Y array), recording the location of the mouse every 0.1 s, with its position defined across nine unequal regions (four corners, four walls and center (Flicker and Geyer, 1982; Young et al., 2010b)). Infrared photobeams (16) located 2.5 cm above the floor and aligned with the long axis of the chamber record rearing behavior. The chamber was illuminated from a single light source above the arena (producing 350 lux in the center, and 92 lux in the four corners). Mice were placed in the bottom left-hand corner of the chamber, facing the corner, and the 60-min test session began immediately. The primary dependent variables were transitions (movement across defined regions), exploratory behavior (hole-poking and rearing), and locomotor pattern (spatial d). Spatial d quantifies the geometrical structure of the locomotor path, where a value of 2 represents highly circumscribed, small-scale movements, while 1 represents straight-line distance-covering movements (Paulus and Geyer, 1991).
Statistical analysis
Operant learning during Hab2 training was analyzed utilizing a repeated measure analysis of variance (ANOVA) with session as a within-subject factor and genotype as a between-subject factor. Time taken to attain criterion was compared utilizing a t-test. Within-session performance in the mouse IGT was analyzed using a repeated measure ANOVA with session half as a within-subject factor and genotype as a between-subject factor. Stable performance in the mouse IGT was compared using a one-way ANOVA with genotype as the between-subject factor. For the mouse BPM, transitions, hole-pokes, and spatial d were subjected to one-way ANOVAs with genotype as the between-subjects factor. Tukey post hoc analyses of statistically significant main or interaction effects were performed where applicable. The level of probability for statistical significance was set at 0.05. Mouse IGT statistics and correlations were performed using SPSS (14.0, Chicago, IL, USA), while mouse BPM data were analyzed using the Biomedical Data Programs (BMDP) statistical software (Statistical Solutions Inc., Saugus, MA, USA).
Results
Operant learning in DAT KD and WT mice
We tested the genetically modified DAT KD mice to investigate the influence of reduced DAT functioning on decision-making behavior. A significant effect of day demonstrated that both KD and WT mice that met criterion learned to nose-poke for a single reward over a 15-day period (F(14,84) = 6.7, p < 0.0001; Figure 1A). Moreover, there was no effect of genotype on the number of days taken to attain criterion (> 40 responses on two consecutive days; F(1,13) = 1.2, ns; Figure 1B). Only 60% of the WT mice attained criterion within 15 days however, in comparison with 100% of the KD mice (Figure 1C).
Instrumental learning of dopamine transporter mutant mice. The learning rate of dopamine transporter (DAT) knockdown (KD) and wildtype (WT) mice were compared. The mice were trained to nose-poke in one of four holes at the rear of an operant chamber with responses measured across sessions (A). No differences in days to criterion were observed (B), although not all WT mice achieved criterion in the allotted time-frame (C). Data presented as mean ± SEM.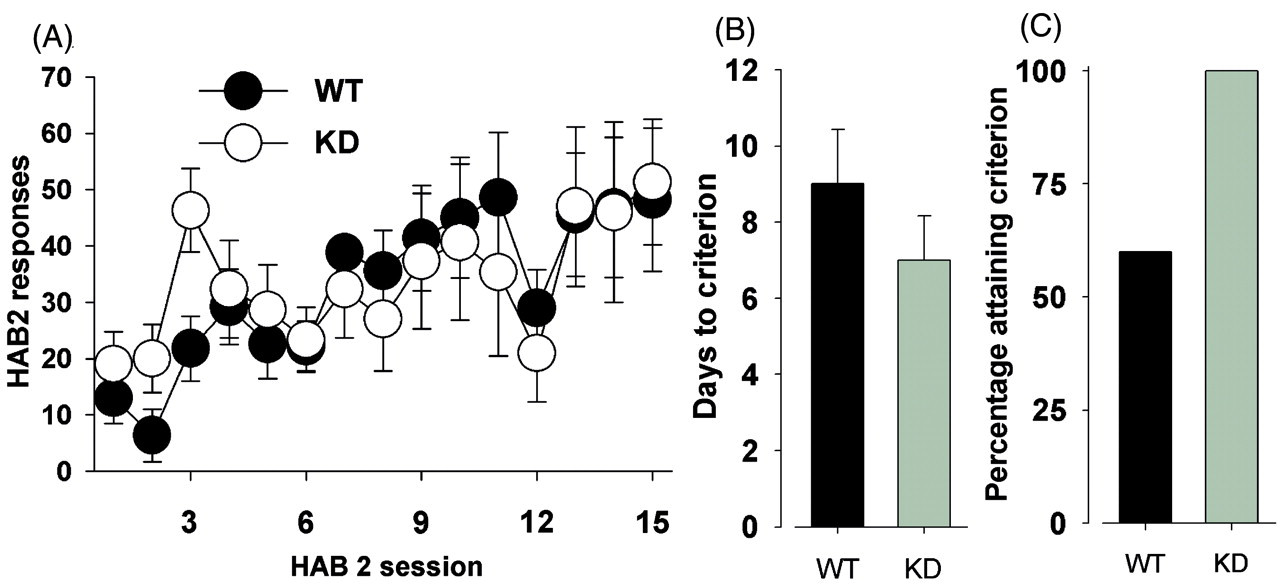
Initial risk preference of DAT KD and WT mice in the mIGT
When the percentage of advantageous choice performance for session 1 was divided by session-half, we observed a significant genotype by session-half interaction (F(1,18) = 7.0, p < 0.05; Figure 2). Post hoc analyses revealed that DAT KD mice exhibited a preference for the ‘riskier’ cues during the first half of training compared with WT littermates (p < 0.05), but by the second session-half they selected the ‘safe’ cues as often as their littermates (p > 0.1). Neither group exhibited an overall preference that differed from chance, however (p > 0.2). No main effect of genotype was observed for premature responses (F < 1, ns), MCL (F < 1, ns), perseveration during punishment (F < 1, ns), perseveration during rewards (F(1,18) = 1.5, ns), % omissions (F < 1, ns), MRL (F(1,18) = 1.4, ns), or total trials (F(1,18) = 1.8, ns) during session 1.
Initial learning of the mouse Iowa Gambling Task. The risk preferences of dopamine transporter wildtype (WT) and knockdown (KD) mice were assessed in session 1 of the mouse Iowa Gambling Task. KD mice exhibited increased risk preference compared with WT mice in the first half of session 1. By the second half, however, the risk preferences of the groups were no longer different. Data presented as mean ± SEM, *denotes p < 0.05 when compared with WT mice within that session half.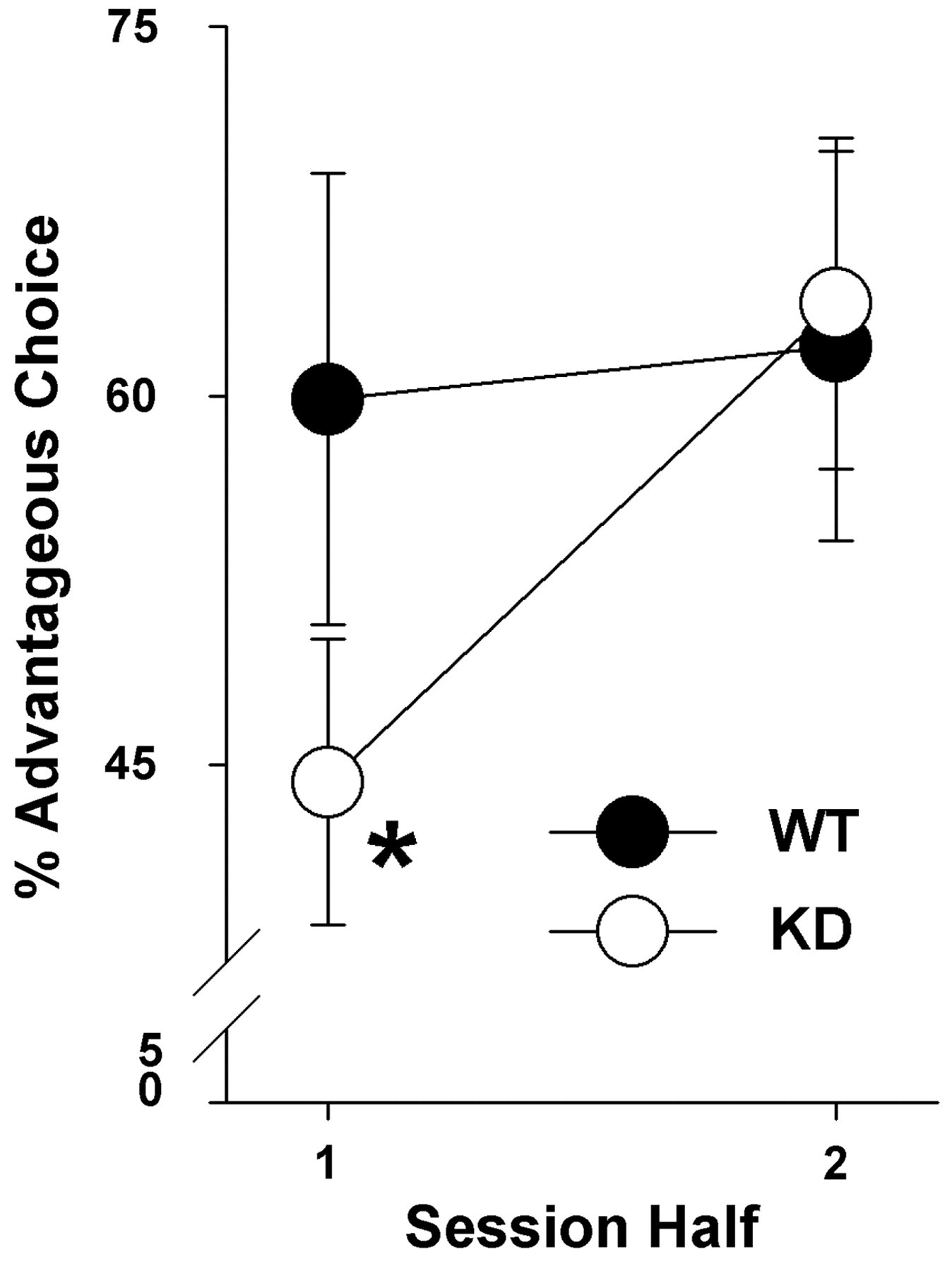
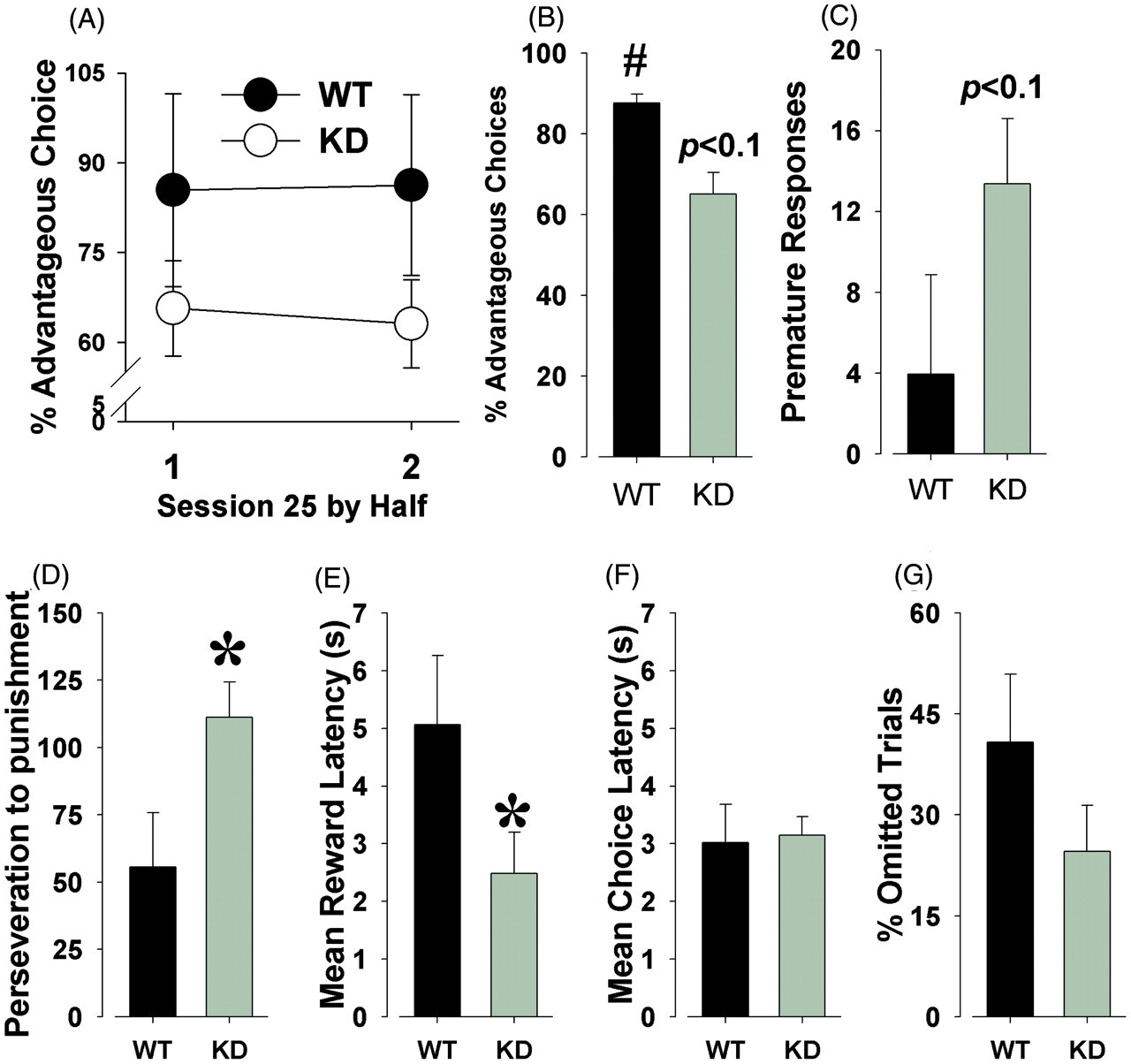
Stable risk preference of DAT KD and WT mice in the mouse IGT
Training in the task continued (Figure 3) until a stable cue preference was observed. Once stable, we again examined performance by session-half. At this stage, no genotype by session-half interaction was observed for % advantageous choice (F(1,18) < 1, ns; Figure 4A). When performance was examined over the entire session, DAT KD mice exhibited a trend toward risky choice when compared with WT mice (65% vs. 88% advantageous choice, respectively; F(1,18) = 2.4, p < 0.1; Figure 4B). Further examination revealed that while WT mice exhibited significantly increased performance when compared with chance (50%; t = 7.3, p < 0.0001), KD mice only exhibited a trend toward a preference for the advantageous holes when compared with chance (t = 1.8, p = 0.096). KD mice also exhibited a trend toward more premature responses compared with the WT mice, although this was not significant (12 vs. 4; F(1,18) = 2.6, p < 0.1; Figure 4C). KD mice exhibited increased perseverative responses after punishments compared with their WT littermates (F(1,18) = 5.4, p < 0.05; Figure 4D), while only a trend was observed for perseverations after a reward (F(1,18) = 3.0, p = 0.10). DAT KD mice exhibited a lower mean reward latency than WT mice (F(1,18) = 6.1, p < 0.05; Figure 4E). No effect of genotype was observed for choice latency, however (F(1,18) < 1, ns; Figure 4F). No effect of genotype was observed for % omitted trials (F(1,18) = 1.8, ns; Figure 4G), or total trials completed (F(1,18) = 1.0, ns).
Alterations in risk preference with continued training. Dopamine transporter (DAT) wildtype (WT) and knockdown (KD) mice were trained in the Iowa Gambling Task for a further 24 days. While the risk preference of DAT WT and KD mice did not differ greatly after training session 1, as training continued WT mice exhibited increased preference for the safer options compared to KD mice. Data presented as mean ± SEM. Stable performance of mice in the Iowa Gambling Task. After 24 sessions of training, the stable risk preferences of dopamine transporter wildtype (WT) and knockdown (KD) mice were compared. Within the final session, WT and KD mice exhibited stable performance from the first to second halves of the session (A). A trend toward riskier performance in KD mice was observed over the entire session (B), as was premature responses (C). KD mice exhibited increased perseverative responses to punishing stimuli (D), as well as shorter mean reward latencies (E). Despite shorter reward latencies, KD did not differ from WT mice in choice latency (F), nor were differences observed in % omitted trials (G). Data presented as mean ± SEM, *denotes p < 0.05 when compared with WT, # denotes p < 0.05 when compared with chance.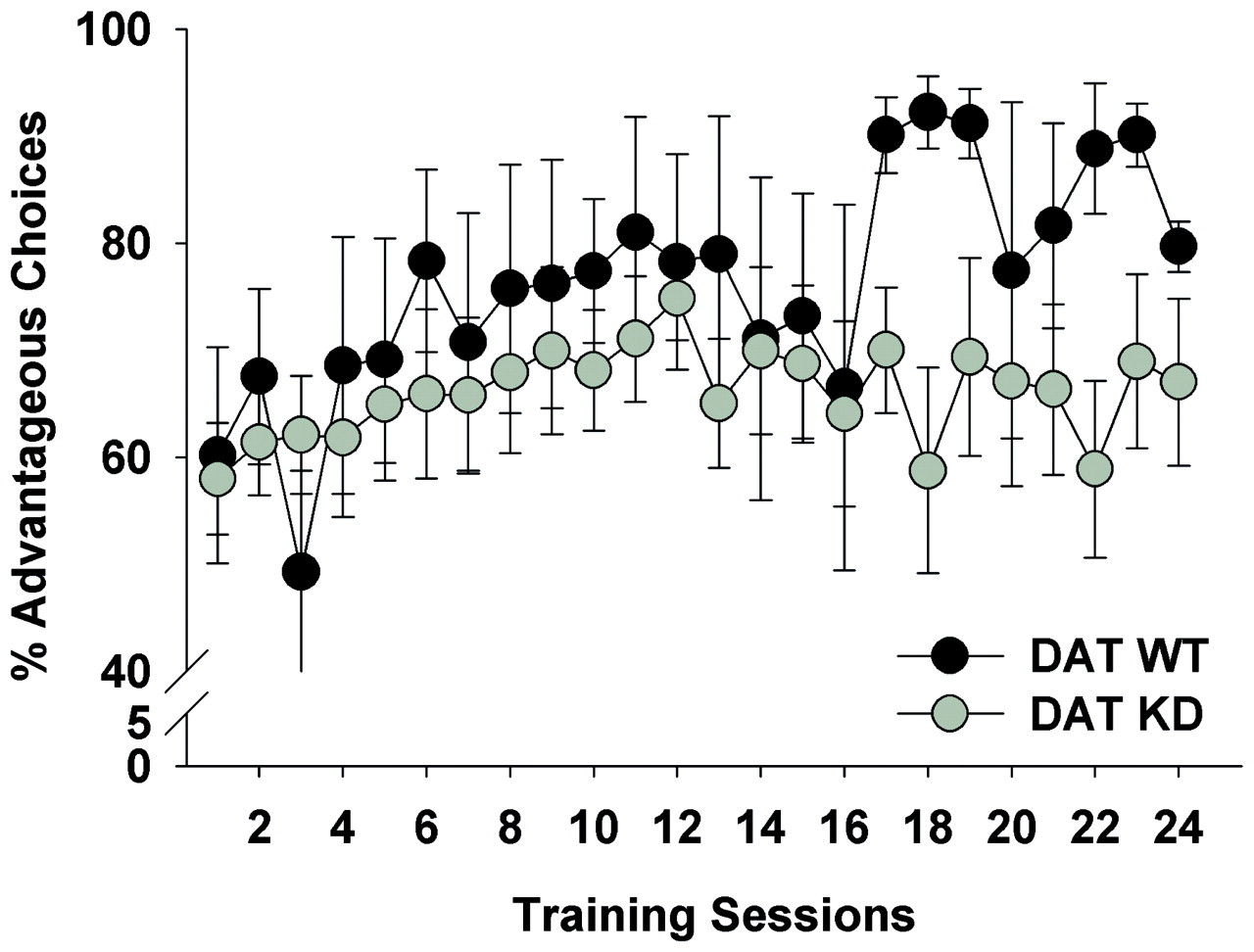
Mania-like exploratory profile of DAT KD mice
Finally, we also tested the exploratory behavior of these mice in the mouse BPM. Consistent with previous reports, DAT KD mice exhibited increased activity as measured by transitions, when compared with WT littermates (F(1,23) = 16.0, p < 0.001; Figure 5A). Moreover, DAT KD mice also exhibited increased specific exploration as measured by hole-pokes (F(1,23) = 8.7, p < 0.01; Figure 5B). Finally, a trend toward reduced spatial d locomotor pattern was also observed in DAT KD mice compared with their WT littermates (F(1,23) = 3.8, p = 0.06; Figure 5C). Choosing the riskier option did not correlate with transitions, spatial d, or weight (p > 0.1) in the BPM, but did exhibit a trend for a modest correlation with specific exploration (hole-poking; r = 0.36, p = 0.084).
Mania-like exploratory profile of dopamine transporter knockdown mice in the behavioral pattern monitor. The exploratory behavior of dopamine transporter wildtype (WT) and knockdown (KD) mice was assessed in the behavioral pattern monitor. Consistent with previous reports, KD mice exhibited increased transitions (A), increased hole-poking (B), and reduced spatial d (C), when compared to WT littermate mice. Data presented as mean + SEM, *denotes p < 0.05 when compared with WT mice.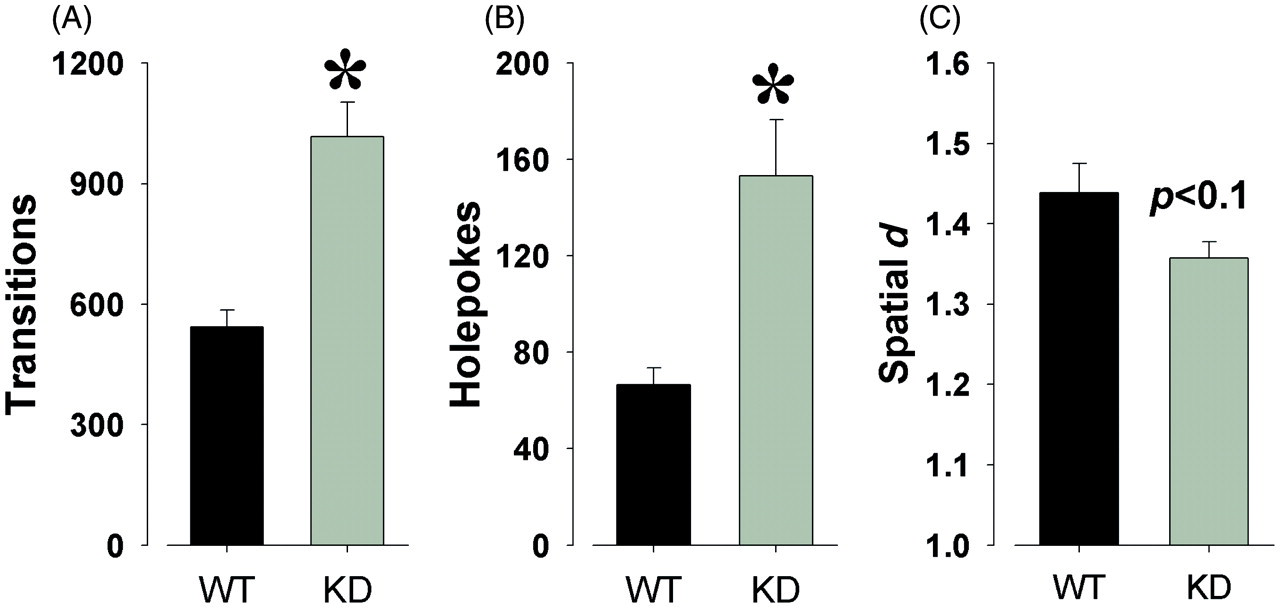
Discussion
DAT KD mice exhibited increased risky behavior in the mouse IGT. This risky behavior was measured by an increased preference for disadvantageous choices. This increase in risky behavior was observed during initial learning in session 1, with a trend toward risk preference even after prolonged training. Moreover, consistent with previous reports, these DAT KD mice exhibited an exploratory profile in the mouse BPM consistent with that exhibited by patients with BD mania. Thus, these data provide further support for the use of DAT KD mice as a mouse model of BD mania.
The present data are consistent with previous reports that DAT KD mice exhibit normal learning in operant-based tasks (Hab2 performance (Cagniard et al., 2006)). In contrast to previous reports, however, WT mice exhibited poorer operant learning than KD mice. The reasons for poorer operant learning in WT mice remain unclear, but could reflect reduced activity in these mice. DAT KD mice backcrossed onto C57BL/6J mice are now available (Tilley et al., 2007) and future studies will examine operant learning in these mice. When the mice in the present study were switched from simple responding to the IGT, the performance of DAT KD and WT mice also differed. From session 1, KD mice exhibited a preference for the disadvantageous holes, only learning to select from the advantageous holes by the second half of the session. While deficient learning in DAT KD mice compared with WT mice could contribute to this observation, we feel that previous reports of normal learning elsewhere in these DAT KD mice (Cagniard et al., 2006), as well as superior learning during the Hab2 phase in the present studies, support the interpretation that these mice select the high-risk holes due to preference for larger rewards. Further, these data are consistent with risk preference in the IGT in patients with BD mania (Adida et al., 2008; Christodoulou et al., 2006; Clark et al., 2001; Yechiam et al., 2008). With a further 24 days of continued training however, DAT KD mice acquired a preference for the advantageous holes, although this preference was only a trend toward being significantly different from chance. Moreover, DAT KD mice still exhibited a lower preference for advantageous holes when compared with WT mice. These findings were not due to changes in preference over a session as seen in session 1. Furthermore, WT mice exhibited a preference for the advantageous holes that differed from chance. Thus, even with continued training, DAT KD mice continued to exhibit a preference for the riskier choices. Therefore, it appears that chronic increased extracellular dopamine levels increases risk preference.
The present study explored risk preference in female DAT KD mice. Previous studies supporting these mice as a model for BD mania have investigated both male and female mice and found no sex effects or interactions in spontaneous exploration reported (Perry et al., 2009; Young et al., 2010b). Similarly, the spontaneous exploration of BD mania patients was unaffected by gender (Perry et al., 2009, 2010). Gender can interact with genes to influence performance in the IGT, although this gender effect was only apparent in the first stage of the IGT where decisions were made under ambiguity and not risk (Stoltenberg and Vandever, 2010). Gender was not reported to influence BD mania performance in the IGT, however (Adida et al., 2008; Clark et al., 2001). In a direct gender comparison in the IGT, it was noted that men chose the safe options more rapidly than women (Weller et al., 2010), results which have been reflected in earlier rodent IGT versions (van den Bos et al., 2006). Thus the learning of the mice in the current study could reflect slower learning rates in WT female mice, in part due to slower learning rates in females, although this slower rate can be influenced by anxiety levels (de Vissier et al., 2010). Another putative confound of using female mice in the present study is that the estrus cycle can affect performance of rodents in operant-based tasks such as cocaine (Roberts et al., 1989) or alcohol (Roberts et al., 1998) self-administration. Further, food deprivation can alter the estrus cycle of rodents that vary by background strain (Tropp and Markus, 2001). The effect of food deprivation on the estrus cycle of DAT KD mice is currently unknown, as is the effect of the estrus cycle on performance of the IGT reported here. Figure 3 details the development of preference toward safe options during training, with no obvious differences between WT and KD mice on specific days. The mice in the present study were all housed close together, thus their estrus cycle was likely synchronized. Given the possible complication of estrus cycle, however, and the better within-session learning exhibited by men in the human IGT, future studies will focus on assessing risk preference in male DAT KD mice in the mouse IGT. Moreover, altering reinforcement and punishment schedules may provide a better opportunity to observe within-session learning, as has recently been described for another rat IGT version (Rivalan et al., 2009).
It was interesting to note that DAT KD mice also exhibited a trend toward more impulsive-like behavior in the task, as measured by premature responses. Previous reports suggest that this measure is linked to decreased 5-HT levels as observed in rats performing the 5-choice serial reaction time task (5CSRTT) (Dalley et al., 2002). This increased motor impulsivity could, however, simply reflect the increased activity in these DAT KD mice, because psychostimulants such as amphetamine also increase measures of motor impulsivity of rats in the 5CSRTT (Robbins, 2002). Such motor impulsivity could also be linked to the increased perseverative responding observed in these mice (Fineberg et al., 2010; Robbins, 2002). Future studies could determine the full extent of impulsive behaviors in these mice (Evenden, 1999).
DAT KD mice did not exhibit shorter mean choice latencies when compared with WT mice, despite increased activity levels. Thus it would appear that during goal-directed behavior, DAT KD mice respond as quickly as their WT littermates. It is interesting to note, therefore, that DAT KD mice were significantly faster than their WT littermates when returning to the magazine to gain their reward. Since this reward latency has been linked to motivational factors for food reward (Robbins, 2002), the data support previous reports of increased motivation in DAT KD mice (Cagniard et al., 2006). This hedonic behavior supports the use of DAT KD mice as a mouse model of BD mania, because patients with mania also exhibit hedonic behavior (Cassidy et al., 2002).
Finally, the DAT KD mice used in the current study exhibited an exploratory profile consistent with that of previous reports and patients with BD mania (Perry et al., 2009; Young et al., 2007b, 2010a). Thus the IGT behavior presented here may well reflect the behavior of patients with BD during their manic phase. Increased risk preference is also observed in patients with attention deficit hyperactivity disorder (ADHD) (Malloy-Diniz et al., 2007). In contrast with ADHD but consistent with BD mania, subthreshold doses of stimulants reinstate the hyperactive phenotype of DAT KD mice (Young et al., 2010b). These converging lines of evidence from the BPM and IGT provide further support for the use of these mice as a model of BD mania (Young et al., 2010b). Moreover, these findings suggest that chronic reductions in DAT functioning result in a preference for riskier options, and may therefore support the idea that reduced DAT functioning may contribute to the risky behavior exhibited by BD manic patients (Amsterdam and Newberg, 2007; Horschitz et al., 2005). The observed relationship between risk preference in the IGT and hole-poking in the BPM suggests that targeting treatments that reduce hole-poking in the BPM may provide a high-throughput screen to develop treatments to improve a broad range symptomatology in BD. It would prove useful in future studies to examine impulsive and cognitive performance of these mice in cross-species translational tasks (Young et al., 2009b) to further assess the relevance of DAT KD mice as a model of BD mania. Tasks assaying executive functioning, such as the attentional set-shifting task (Birrell and Brown, 2000; Young et al., 2010c), or vigilance and response inhibition, such as the 5-choice continuous performance test (Young et al., 2009a) may prove useful given the deficits in performance observed in BD patients in these tasks. Future studies will also begin to examine the effects of current treatments on the exploratory profile of these mice in the BPM.
Footnotes
Acknowledgements
We thank Richard Sharp and Mahálah Buell for their support.
Funding
This study was supported by a NARSAD Young Investigator Award, as well as the NIH (grant numbers R21-MH085221 and R01-MH071916), and the Veteran's Administration VISN 22 Mental Illness Research, Education, and Clinical Center.
Conflict of interest
The authors report no conflict of interest.