
Abstract
Total ginsenosides have been shown to have therapeutic actions as antidepressants. We report a major active ingredient of total ginsenosides, the ginsenoside Rb3 (Rb3), which may have antidepressant-like effects. Using the forced swim test, tail suspension test, and learned helplessness procedure, we found that Rb3 had significant anti-immobility effects in mice in the forced swim and tail suspension tests and reduced the number of escape failures in the learned helplessness procedure. In a reserpine-induced syndrome model, Rb3 attenuated hypothermia, palpebral ptosis, and akinesia. In the chronic mild stress model, chronic Rb3 administration reversed the decrease in locomotor activity, novelty-suppressed feeding, and sucrose preference. Furthermore, neurochemical tests were performed to support our hypothesis that biochemical variations (i.e. brain-derived neurotrophic factor and the monoamine neurotransmitters 5-hydroxytryptamine, dopamine, and norepinephrine) are involved in Rb3’s antidepressant-like effects. Finally, we found, using whole-cell patch-clamp recordings, that the action potential transmission in neurons within the somatosensory cortex was excited by Rb3 perfusion and blocked with Panax notoginseng total saponins extracted from leaves. This study provides evidence for the mechanism of action of the antidepressant-like effects of Rb3.
Keywords
Introduction
Depression is a widely distributed, chronic, recurrent syndrome that reflects and affects an individual’s quality of life. It is commonly considered a disorder caused by disturbances in the function of neurotransmitters (Drevets, 2001; Millan et al., 2000). Neurotransmitter-related medicines, such as serotonin reuptake inhibitors, are commonly prescribed as therapeutic drugs (Holtzheimer and Nemeroff, 2006; Hu and Fu, 2003). These medications, however, tend to be associated with certain side effects (Holtzheimer and Nemeroff, 2006), and herbal medicines, such as St. John’s wort (Hypericum perforatum L.; Bilia et al., 2002) and Withania somnifera (Bhattacharya and Muruganandam, 2003), may be better tolerated and are increasingly chosen as alternative remedies for depression.
Panax notoginseng, a species of Araliaceae, has widely served as an herbal tonic for thousands of years in ethnomedicine in China and other Asian countries for its hemostatic and cardiovascular effects (Lau et al., 2009). Its various therapeutic functions have been demonstrated in recent pharmacological research (Ng, 2006). The main ingredients responsible for the function of Panax notoginseng are ginsenosides (saponins). More than 150 ginsenosides have been isolated from Panax species, which can be classified into four types: protopanaxadiol, protopanaxatriol, ocotillol-type, and oleanolic acid (Shi et al., 2010).
Several studies have reported the clinical effects of Panax notoginseng total saponins (PNTS) on insomnia, anxiety, and neurasthenia neurosis (Tan and Tang, 1999; Zhao and Lan, 2005). Previous studies in our laboratory demonstrated the antidepressant-like effects of PNTS in several behavioral tests in mice and rats (Xiang et al., 2011). These effects may occur by increasing the levels of the brain monoamine neurotransmitters 5-hydroxytryptamine (5-HT) and norepinephrine (NE). Moreover, electrophysiological experiments have shown that PNTS modulates Na+ currents and Ca2+ concentrations, with the possible involvement of action potentials. Wang et al. (2007) reported the influence of PNTS on increasing nestin and brain-derived neurotrophic factor (BDNF) expression following focal cerebral ischemia. These results suggest a possible connection between this traditional medicine and depression, as BDNF and other neurotrophic factors are decreased in postmortem hippocampal tissue in depressed patients (Schmidt and Duman, 2007). The antidepressant effects of ginseng total saponins have also been reported by other groups (Dang et al., 2009; Hu and Fu, 2003; Kim et al., 2002).
One of the protopanaxadiol ginsenosides, ginsenoside Rb3 (Rb3), can be isolated from several different Panax species (Xu et al., 2005) and can be found at high concentration in PNTS (Wan et al., 2006). Several studies have suggested the protective effects of Rb3 on the central nervous system. Li et al. (2004) showed that Rb3 has protective effects against ischemia-induced injury in cerebral cells by enhancing antioxidant ability and lowering the amount of free radicals. Moreover, Xu et al. (2005) suggested that Rb3 protects neurons in hippocampal slices exposed to simulated ischemia by decreasing the affinity of glycine for receptors and delaying receptor activation in the hippocampus. Rb3 also shows protective effects on hypoxic brain injury, the mechanism of which may involve inhibition of γ-aminobutyric acid (GABA) exhaustion in nerve cells (Shen et al., 2005). This same research team also demonstrated Rb3’s preventive effect against glutamine-induced damage in cerebral cells by inhibiting N-methyl-D-aspartate (NMDA)-induced calcium influx (Shen et al., 2008). The involvement of glycine, GABA, NMDA, Na+ currents, and K+ currents has long been considered to play a significant role in the balance of excitation–inhibition circuits, the dysfunction of which may cause mental illness, such as depression (Stone et al., 2008). To evaluate the potential antidepressant-like effects of Rb3 and explore its possible mechanism of action, the present study examined Rb3’s effects on behavior, monoamine neurotransmitters, BDNF expression, and somatosensory cortical neuronal activity in mice.
Methods
Animals and drugs
Male NIH mice weighing 18–20 g (Guangdong Medical Laboratory Animal Center, Guangzhou, China) were group-housed under standard laboratory conditions (25 ± 2°C, 60–70% humidity, 12 h/12 h light/dark cycle, standard food and water available ad libitum). Unweaned Sprague-Dawley rats (postnatal 14–18 days old, Guangdong Medical Laboratory Animal Center) were used in the electrophysiological experiments. All experiments were performed in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH publication no. 85-23, revised 1985) and approved by the Animal Welfare Committee of Sun Yat-Sen University. All efforts were made to minimize the number of animals used and their suffering.
Ginsenoside Rb3 (purity >90%; Fengshanjian, Kunming, China) and fluoxetine (as a standard antidepressant; Patheon, Bourgoin-Jallieu, France) were dissolved in normal saline to form Rb3 solutions (3, 7.5, and 15 mg/mL) and fluoxetine solution (1 mg/mL). Drugs were intragastrically administered at 10 mL/kg body weight so that the doses of Rb3 were 30, 75, and 150 mg/kg body weight, and the dose of fluoxetine was 10 mg/kg. PNTS (Medi-World, Foshan, China) was used in the electrophysiological study.
Behavioral despair model
Open field test
Because psychostimulants exert few clinical effects as antidepressants but show antidepressant-like effects in the forced swim test (FST) (Sherman et al., 1982), the acute effects of Rb3 on locomotor activity were evaluated in the open field test (OFT) before the following behavioral tests were conducted to exclude the possibility of false positive results. Mice (n = 75) were randomly divided into five groups and intragastrically received saline (10 mL/kg), Rb3 (30, 75, and 150 mg/kg), and fluoxetine (10 mg/kg) for three consecutive days. In accordance with Boissier and Simon (1965), 1 h after the final administration the animals were individually placed into the corner of the apparatus (45 cm × 45 cm × 35 cm) and recorded by video camera for 5 min. Their movement distances were analyzed by computer software (ZH-ZFT, Zhenghua, Huaibei, China). The apparatus was cleaned after each mouse was tested. A decreased distance traveled was interpreted as a decrease in locomotor activity.
Forced swim test
The FST was performed as previously described by Porsolt et al. (1977) and Detke et al. (1995), with minor modification. Just after the OFT, mice were individually placed in a Plexiglas cylinder (19 cm diameter, 30 cm height) containing 18 cm-deep water (20 ± 1°C) and recorded by video camera for 6 min. The immobility time during the last 4 min of the 6-min trial was calculated using a time-sampling technique (ZH-QPT, Zhenghua Huaibei, China). Immobility refers to the cessation of struggling and remaining motionless with the exception of movements needed to keep the mouse’s head above the water.
Tail suspension test
The tail suspension test (TST) was performed as previously described (Steru et al., 1985). One hour after the fourth day of administration, mice were individually suspended by their tail using adhesive tape (1 cm from the tip of the tail) in a sound-proof box, with the head 15 cm from the bottom. Each mouse was recorded by video camera for 6 min, and the immobility time was recorded during the final 4 min of the total 6-min test using a time-sampling technique. Mice were considered immobile only when they hung passively and were completely motionless.
Learned helplessness procedure
In the learned helplessness procedure, mice were exposed to inescapable and unpredictable electric shocks and observed for abnormal behaviors that reflect, to a certain extent, a deteriorated emotional state in humans (Rojas-Corrales et al., 2002). The protocol was divided into two parts (Seligman and Beagley, 1975). During the first part (helplessness induction, day 1), 60 scrambled randomized inescapable electric shocks (IESs, 33 V for 15 s, 20-90 s intervals) were delivered through a stainless-steel grid floor in a two-way shuttle box (BA-200, TME Technology, Chengdu, China). Control animals were placed in boxes for the same period of time without receiving electric shock. During the second part (conditioned avoidance test, day 2-4), mice were allowed to adapt to the shuttle box for 5 min and were then exposed to 30 avoidance trials. In each trial, a light signal (conditioned stimulus) was presented during the first 3 s, followed by a 3 s electric footshock (33 V, unconditioned stimulus) administered via the grid floor of the shuttle box. A 24 s resting period then occurred. If a mouse failed to escape into the other compartment after the light signal (conditioned stimulus) and the electric footshock (unconditioned stimulus), then an escape failure was recorded. Mice (n = 90) were randomly divided into six groups and intragastrically received saline (10 mL/kg, for control and IES-Saline groups), Rb3 (30, 75, and 150 mg/kg), and fluoxetine (10 mg/kg) once on day 1 (after IESs) and twice with the same dose on subsequent days (1 h before test and 16:00 on same day).
Reserpine-induced hypothermia, palpebral ptosis, and akinesia
Mice (n = 135) were randomly divided into six groups and received saline (10 mL/kg, for control and reserpine–saline groups), Rb3 (30, 75, and 150 mg/kg), and fluoxetine (10 mg/kg) continuously for 7 days. In accordance with Bourin et al. (1983), 30 min after the final administration, mice intraperitoneally received 2.5 mg/kg reserpine (Bangmin, Guangzhou, China). Three parameters (rectal temperature, degree of palpebral ptosis, and akinesia) were recorded 1, 3, 5, and 7 h after reserpine treatment. The degree of palpebral ptosis was evaluated according to the following rating scale: 0, eyes open; 1, eyes one-quarter closed; 2, eyes half closed; 3, eyes three-quarters closed; and 4, eyes completely closed (Sánchez-Mateo et al., 2007). To measure akinesia, mice were placed in the center of a circle (7.5 cm diameter). A mouse was judged as akinetic (i.e. present or not) if it remained within the circle after 15 s (Kato et al., 1998).
Chronic mild stress procedure
Stress protocol and administration
Mice (n = 63) were exposed sequentially to the chronic mild stress (CMS) procedure, in which the mice were exposed to various unpredictable mild stressors that kept changing over weeks and caused stress-related and depression-like behavior. The protocol was adopted as described by Willner et al. (1992) with slight modifications. The unpredictable stressors in the CMS schedule consisted of 10 diverse, continuous treatments for five consecutive weeks. Antidepressant administration accompanied the stressors beginning in week 3 (Table 1). Control animals received saline and were left undisturbed in their home cages. Several behavior tests were performed each week during the experiment.
Chronic mild stress procedure and drug or saline administration.
SC, soiled cage with 250 ml water in sawdust bedding; DV, diurnal variation; BR, behavior restraint for 30 min in a tube (8 cm diameter, 20 cm length); WD, water deprivation; WN, background white noise for 30 min; CWS, cold water swimming (4°C) for 5 min; FD, food deprivation; TP, tail pinch for 1 min; TS, thermal stimulation (45°C); CT, 45° cage tilt; SPT, sucrose preference test; OFT, open field test; AT, actimeter test; NSFT, Novelty-suppressed feeding test; ad, drug or saline administration.
Locomotor activity test
Locomotor activity was assessed in either the OFT or actimeter test. In the OFT, each mouse was placed in the center of an apparatus and recorded by video camera for 7 min. Their movement distances during the last 5 min of the 7 min trial were analyzed using computer software. In the actimeter test, mice were tested in their home cage (35 cm × 24 cm × 16 cm) by placing them under the infrared probe of the actimeter (YLS-1 C, Zhenghua, Huaibei, China). The infrared detection system automatically detected the movements of the animal, and the number of movements in 5 min was recorded.
Novelty-suppressed feeding test (NSFT)
The method was performed according to Bodnoff et al. (1988) with minor modification. After 36 h food deprivation, mice were individually placed into the corner of a plastic box (56 cm × 50 cm × 35 cm) with 20 food pellets (1 cm about 1.8 g each) in the center. The first feeding latency in the 3min period (i.e. the period from entering the plastic box to biting the pellet) was recorded. Immediately after this test, the animal was transferred to its home cage, and food consumption in each mouse was measured within 5 min.
Sucrose preference test
The sucrose preference test was used as an indicator of anhedonia and consisted of training and testing phases (An et al., 2008). After acclimatization (1 week), mice were trained for 3 days to consume a 1% (w/v) sucrose solution before initiating the CMS procedure. Before each test, the animals were deprived of food and water for 12 h. Each mouse was then provided with two bottles, one containing a 1% (w/v) sucrose solution and one containing distilled water, for 24 h. Fluid consumption was measured by comparing the weight of each bottle before and after the test. Mice were monitored weekly under similar conditions throughout the entire procedure. The proportion of sucrose intake is expressed as the following: sucrose intake/(sucrose intake + water intake) × 100%.
Brain tissue collection and weighing
After the CMS procedure, mice were decapitated under brief ether anesthesia. The brains were removed, and weighed, then prefrontal cortex, total hippocampus, total amygdala were carefully and immediately dissected on glass plates over ice by a trained researcher based on the mouse brain atlas (Paxinos and Franklin, 2001). Briefly, the brain was removed and dissected into three blocks (anterior, middle, and posterior) by two coronal cuts (at the anterior edge of the optic chiasm and at the posterior edge of the mamillary body). The bilateral blocks of prefrontal cortex were dissected from the medial portion of the anterior block. The portions of the hippocampus were removed bilaterally from the dorsal-middle block. The bilateral ventral-middle blocks corresponded to the amygdala (Miura et al., 2009). All tissues were weighed and prepared for measurements of BDNF and monoamine neurotransmitters.
Measurement of BDNF and monoamine neurotransmitters in different brain regions
BDNF levels in the prefrontal cortex and hippocampus were quantified by enzyme-linked immunosorbent assay (ELISA) (Jacobsen and Mørk, 2006) using an ELISA kit (EK0476, Boster, Wuhan, China) and an enzyme-labeled meter at 450 nm (Wellscan MK3, Labsystems Dragon, Helsinki, Finland). 5-HT, dopamine, and norepinephrine levels in the prefrontal cortex, hippocampus, and amygdala were analyzed by the acid extraction of tissue and another single solvent step (Shellenberger and Gordon, 1971) that is based on the alumina method (Anton and Sayre, 1962). Briefly, after homogenization with N-butyl alcohol (Fuyu, Tianjin, China), brain tissue homogenates were rinsed in phosphate-buffered saline to extract norepinephrine and dopamine, which react with iodine reagent in alkaline condition to form indole-3-hydroxy compounds or indole-2-hydroxy compounds that are fluorescent. Another homogenate was rinsed in N-heptane and HCl to extract 5-HT, which reacts with O-phthaladehyde (Qiyun, Guangzhou, China) in acidic conditions to form a fluorescent compound. The fluorescent intensity of each compound was linearly related to its concentration within limits. Fluorescent derivatives of 5-HT, norepinephrine, and dopamine were measured using a spectrophotofluorometer (RF-5301PC, Shmadzu; 5-HT, 480/365 nm; dopamine, 375/325 nm; norepinephrine, 480/385 nm).
Whole-cell patch-clamp recordings of action potentials
The whole-cell patch-clamp recording procedure was conducted by our tested researchers and based on previously described methods (Cui et al., 2010; Wang et al., 2009; Xiang et al., 2006). Newborn Sprague-Dawley rats, of either sex, aged postnatal 14–18 days old, were decapitated, and brains were removed rapidly. Cortical brain slices (300 µm thickness) were cut using a vibratome (Campden, Loughborough, UK) and slices were incubated on cell culture inserts (8 µm pore diameter, Becton Dickinson, Franklin Lakes, NJ) covered by a thin layer of cutting solution and surrounded by a humidified 95% O2 and 5% CO2 atmosphere at room temperature (25°C). The cutting solution contains (in mmol/L) 125 NaCl, 25 NaHCO3, 2.5 KCl, 1.25 NaH2PO4, 1 CaCl2, 7 MgCl2, 20 D-glucose (pH = 7.4). After at least 1 h of incubation, slices were transferred to a submerged recording chamber with continuous flow (2 mL/min) of artificial cerebrospinal fluid (aCSF: 124 mmol/L NaCl, 26 mmol/L NaHCO3, 1.25 mmol/L NaH2PO4, 2.5 mmol/L KCl, 2 mmol/L CaCl2, 2 mmol/L MgCl2, 10 mmol/L D-glucose) and gassed with 95% O2 5% CO2 to yield pH 7.4. Whole-cell recordings from pyramidal neurons in the somatosensory cortex (layer III–V) were performed using infrared differential interference contrast microscopy and a Multiclamp 700 A patch-clamp amplifier (Axon Instruments, Foster City, CA, USA). All experiments were performed at 37 ± 1°C. In current-clamp recording mode, action potentials were induced by injecting current pulses (200 pA, 150 ms duration). After normal recording, the slices were treated with Rb3 (30 µg/mL) or PNTS (0.125 mg/mL) by perfusion for 20 min.
Statistical analyses
The data obtained by whole-cell patch-clamp recordings were obtained using an Axopatch 700 A amplifier (Axon Instruments, Burlingame, CA, USA), and digitized at sampling intervals of 10-50 μs using a Digidata 1320 AD/DA converter (Axon Instruments). Stimulation, acquisition, and data analysis were controlled with pClamp 9.0 (Axon Instruments).
Statistics and data plotting were done with Origin 7.5 (Origin Lab, Northampton, MA, USA) and Microsoft Excel 2003. All quantitative data in the text and figures are expressed as mean ± SEM unless stated otherwise. The statistical analysis of the results of akinesia induced by reserpine was performed by χ2 test, and that of electrophysiological characteristics recording by whole-cell patch-clamp were obtained by student’s t-test: all other statistical significances were determined using one-way analysis of variance (ANOVA) followed by Bonferroni post-hoc test. Values of p < 0.05 were considered statistically significant.
Results
Effect of Rb3 on behavioral despair models
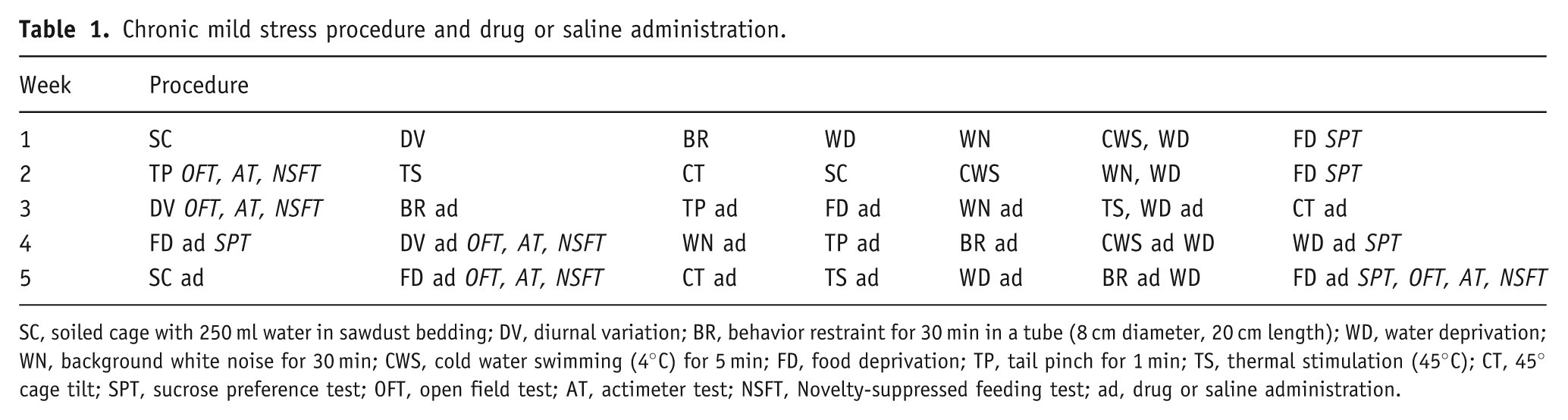
A one-way ANOVA and Bonferroni post-hoc test showed no significant differences in the distances traveled among groups with different treatments in the OFT (p > 0.05; Figure 1A). This result excludes the false positive results in the following experiments.
Effects of Rb3 and fluoxetine on acute behavior models of depression. (A) Distances traveled in the OFT. (B, C) Immobility time in the (B) FST and (C) TST. (D) Number of escape failures in the learned helplessness procedure. The number of mice in each group is shown in parentheses in each figure. Each column represents the mean ± SEM. *p < 0.05, **p < 0.01, compared with saline or IES–saline; ## p < 0.01, compared with control. OFT, open field test; FST, forced swim test; TST, tail suspension test; LH, learned helplessness; IES, inescapable electric shock; Rb3, ginsenoside Rb3; Flu, fluoxetine.
The FST and TST have long been used to screen antidepressants because these models cause a state of lower ‘mood’ in animals that may be a component of clinical depression in humans (Porsolt et al., 1977). In the present FST (Figure 1B), the group that received 75 mg/kg Rb3 exhibited significantly decreased immobility time compared with saline treatment (75 mg/kg Rb3: 69.1 ± 5.42 s; saline: 100.6 ± 14.39 s; one-wayANOVA and Bonferroni post-hoc test, p < 0.05). The immobility times in the other groups (30 and 150 mg/kg Rb3, 10 mg/kg fluoxetine) were not significantly different from saline.
In the TST (Figure 1C), the groups that received 75 mg/kg Rb3 and 10 mg/kg fluoxetine exhibited significantly reduced immobility compared with saline treatment (75 mg/kg Rb3 92.79 ± 9.734 s; fluoxetine: 93.03 ± 8.437 s; saline: 125.1 ± 11.11 s; one-way ANOVA and Bonferroni post-hoc test, p < 0.05). The other groups did not exhibit significant decreases.
Effect of Rb3 in the learned helplessness procedure
The learned helplessness procedure evaluates certain behaviors that resemble a deteriorated affective state in humans. The involvement of the hippocampal noradrenergic system in learned helplessness has been documented by several studies (Martin et al., 1986; Petty et al., 1993; Sherman et al., 1982). In the present study, mice that received IES during the induction period exhibited a significantly increased number of escape failures on all three testing days (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. control; Figure 1D), indicating a deficit in the avoidance task. Rb3 (30, 75, and 150 mg/kg) and fluoxetine (10 mg/kg) treatments, however, led to a significant attenuation of the increased number of escape failures (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. IES–saline group for the three testing days). The degree of attenuation in the number of failures positively correlated with Rb3 dose and time after administration.
Effect of Rb3 on reserpine-induced syndrome
Reserpine is an antihypertensive drug that depletes neuronal storage granules of biogenic amines in the brains of rodents and produces a clinically significant depression-like state (Goodwin and Bunney, 1971). Mice became hypothermic, akinetic, and diarrhetic, with eyelid drooping, in response to reserpine. Administration of Rb3 and fluoxetine significantly improved this condition (Figure 2).
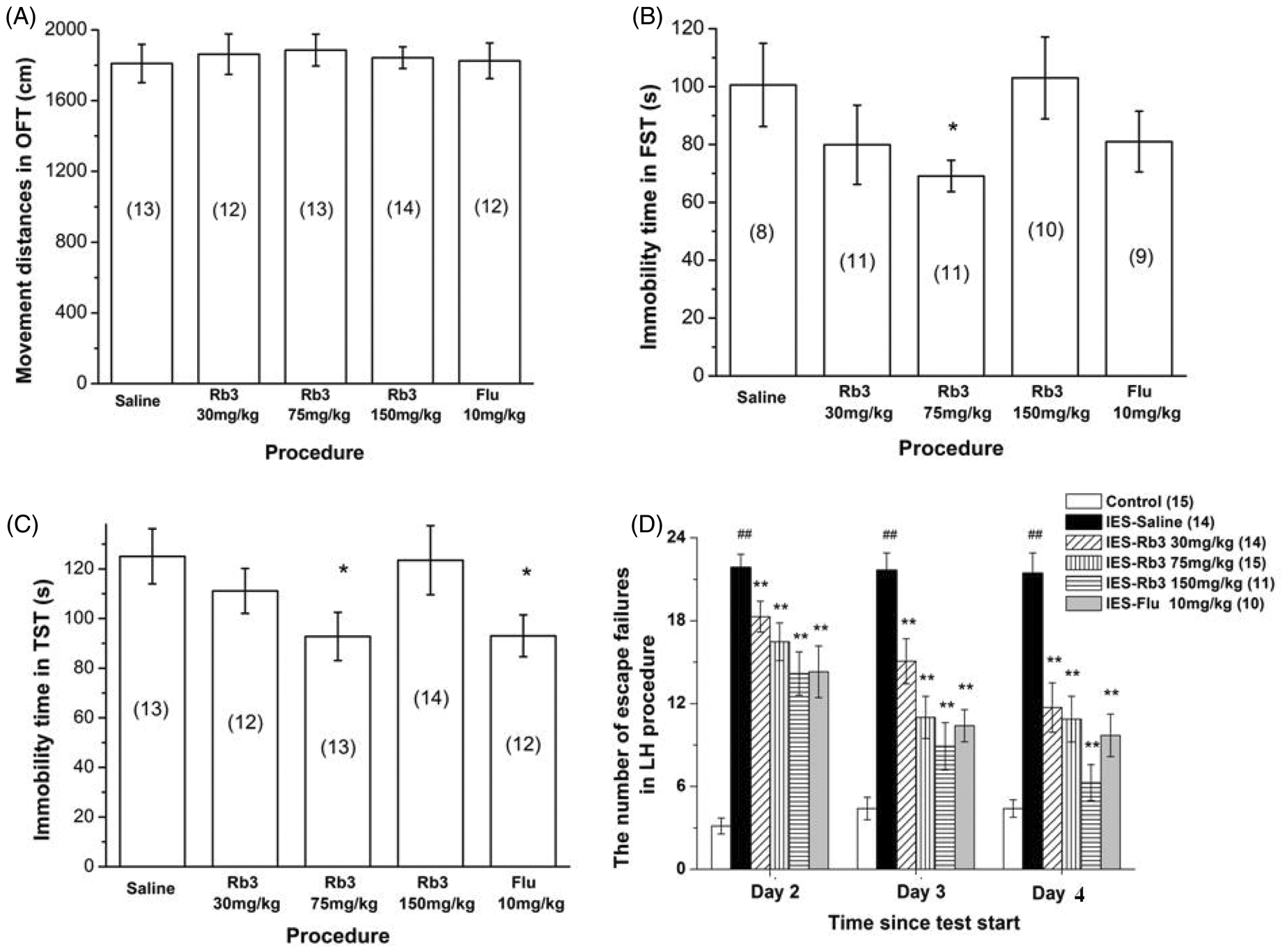
Effect of Rb3 on reserpine-induced hypothermia and palpebral ptosis. Experiments were performed before and 1, 3, 5, and 7 h after reserpine injection. (A) Rectal temperature and (B) degree of palpebral ptosis in different groups of mice. The number of mice in each group is indicated in parentheses in each figure. Each dot and column represents the mean ± SEM. *p < 0.05, **p < 0.01, compared with RES–saline. RES, reserpine; Rb3, ginsenoside Rb3; Flu, fluoxetine.
Reserpine caused a significant reduction in rectal temperature from the 1st hour to the 7th hour (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. control; Figure 2A). Pretreatment with Rb3 (150 mg/kg) and fluoxetine (10 mg/kg) attenuated the hypothermic response from the 1st hour to the 7th hour. Rectal temperature in both groups significantly increased at the 1st hour and 3rd hour (one-way ANOVA and Bonferroni post-hoc test, p < 0.05 vs. reserpine-saline) and further increased at the 5th hour and 7th hour (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. reserpine-saline). Rectal temperature in the 30 mg/kg Rb3 group significantly increased only at the 7th hour (one-way ANOVA and Bonferroni post-hoc test, p < 0.05 vs. reserpine-saline). The 75 mg/kg Rb3 group did not exhibit a significant rectal temperature increase during the entire trial.
The degree of palpebral ptosis in mice (Figure 2B) was significantly aggravated by reserpine treatment compared with control (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. control). Rb3 (150 mg/kg) and fluoxetine (10 mg/kg) significantly reversed palpebral ptosis from the 1st hour to the 7th hour (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 at the 1st hour and 3rd hour vs. reserpine-saline; p < 0.05 at the 5th hour and 7th hour vs. reserpine-saline). The 75 mg/kg Rb3 group showed a significant reduction in the degree of palpebral ptosis at the 3rd hour and 5th hour (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 at the 3rd hour; p < 0.05 at the 5th hour vs. reserpine-saline). The 30 mg/kg Rb3 group showed a significant reduction in the degree of palpebral ptosis only at the 5th hour (one-way ANOVA and Bonferroni post-hoc test, p < 0.05 vs. reserpine-saline). In the control group, all mice opened their eyes completely (data in the control group were all zero and are not included in the figure).
The χ 2 test was used to analyze the results of the akinesia test. At the 1st hour, the reserpine-saline group (n = 19) exhibited a reduction in the proportion of exiting from the circle compared with the control group (n = 15; 15.8% in reserpine-saline, 100% in control, p < 0.01). Administration of 150 mg/kg Rb3 (n = 21) and 10 mg/kg fluoxetine (n = 18) significantly reversed the reduction (57.1% in 150 mg/kg Rb3 group, p < 0.01 vs. reserpine-saline; 44.4% in fluoxetine group, p < 0.05 vs. reserpine-saline). Treatment with 30 mg/kg (n = 18) and 75 mg/kg (n = 21) Rb3 did not significantly reverse the reduction compared with the reserpine-saline group (33.3% in 30 mg/kg Rb3 group; 23.8% in 75 mg/kg Rb3 group). At the 3rd, 5th, and 7th hours, the results remained the same.
Effects of Rb3 on CMS model
Effect of Rb3 on locomotor activity
Body weights were recorded weekly during the experiment. As shown in Table 2, before the application of CMS body weights were not significantly different among groups (18-19 g average, one-way ANOVA and Bonferroni post-hoc test, p > 0.05). The CMS procedure induced a significant decrease in body weight during the second week (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. control). Administration of Rb3 (30, 75, and 150 mg/kg) and fluoxetine (10 mg/kg) in week 3 did not significantly attenuate the decrease until week 5 (one-way ANOVA and Bonferroni post-hoc test, 75 mg/kg Rb3, p < 0.01; 30 and 150 mg/kg Rb3 and fluoxetine, p < 0.05 vs. CMS-saline).
Effects of ginsenoside Rb3 and fluoxetine on behavioral test in CMS model.
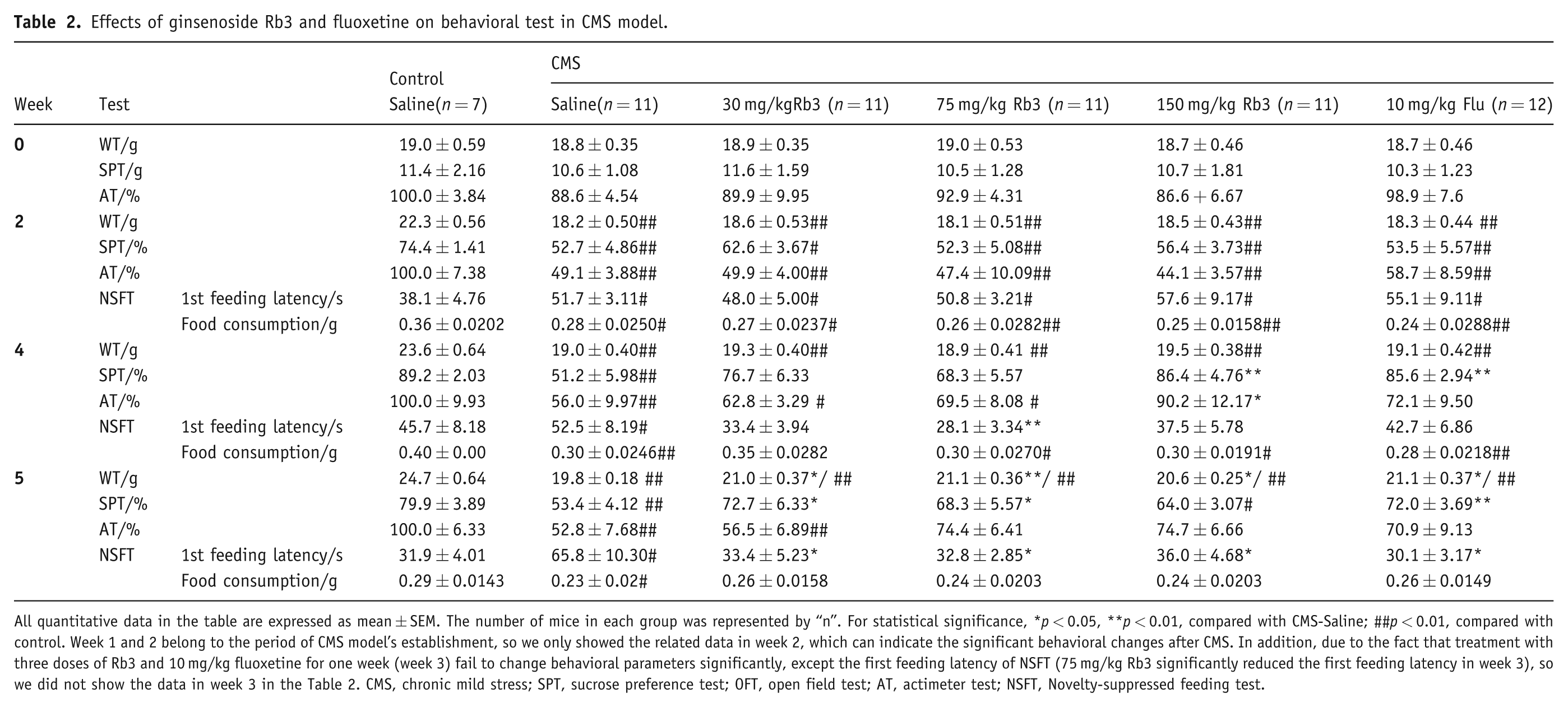
All quantitative data in the table are expressed as mean ± SEM. The number of mice in each group was represented by “n”. For statistical significance, *p < 0.05, **p < 0.01, compared with CMS-Saline; ##p < 0.01, compared with control. Week 1 and 2 belong to the period of CMS model’s establishment, so we only showed the related data in week 2, which can indicate the significant behavioral changes after CMS. In addition, due to the fact that treatment with three doses of Rb3 and 10 mg/kg fluoxetine for one week (week 3) fail to change behavioral parameters significantly, except the first feeding latency of NSFT (75 mg/kg Rb3 significantly reduced the first feeding latency in week 3), so we did not show the data in week 3 in the Table 2. CMS, chronic mild stress; SPT, sucrose preference test; OFT, open field test; AT, actimeter test; NSFT, Novelty-suppressed feeding test.
Effect of Rb3 on locomotor activity
In the OFT, the distances traveled were not significantly different among groups during each week (one-way ANOVA and Bonferroni post-hoc test, p > 0.05, data not shown). In the actimeter test (Figure 3A, Table 2), the number of movements of each mouse was recorded and normalized, with controls set to 100%. The CMS procedure caused a significant decrease in the number of movements in all groups in week 2 (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. control). Treatment with three different doses of Rb3 and fluoxetine fail to significantly increase the number of movements in week 3 (one-way ANOVA and Bonferroni post-hoc test, p > 0.05, data not shown). Treatment with 150 mg/kg Rb3 significantly increased the number of movements in week 4 compared with CMS-saline (150 mg/kg Rb3, 90.2 ± 12.17% of control; CMS-saline, 56.0 ± 9.97% of control; p < 0.05). Fluoxetine (10 mg/kg) treatment in week 4 and week 5, and 75 and 150 mg/kg Rb3 treatment in week 5 increased the number of movements up to above 70% of the control group, but no significant changes were seen in the comparisons between Rb3 (75 and 150 mg/kg) or fluoxetine (10 mg/kg) group and CMS-saline group, respectively (one-way ANOVA and Bonferroni post-hoc test, p > 0.05 vs. CMS-saline and control). The OFT results likely did not reach statistical significance because mice were moved to an unfamiliar arena, and the new experimental setting may have triggered an increase in locomotor activity. In contrast, mice remained in their home cages during the actimeter test.
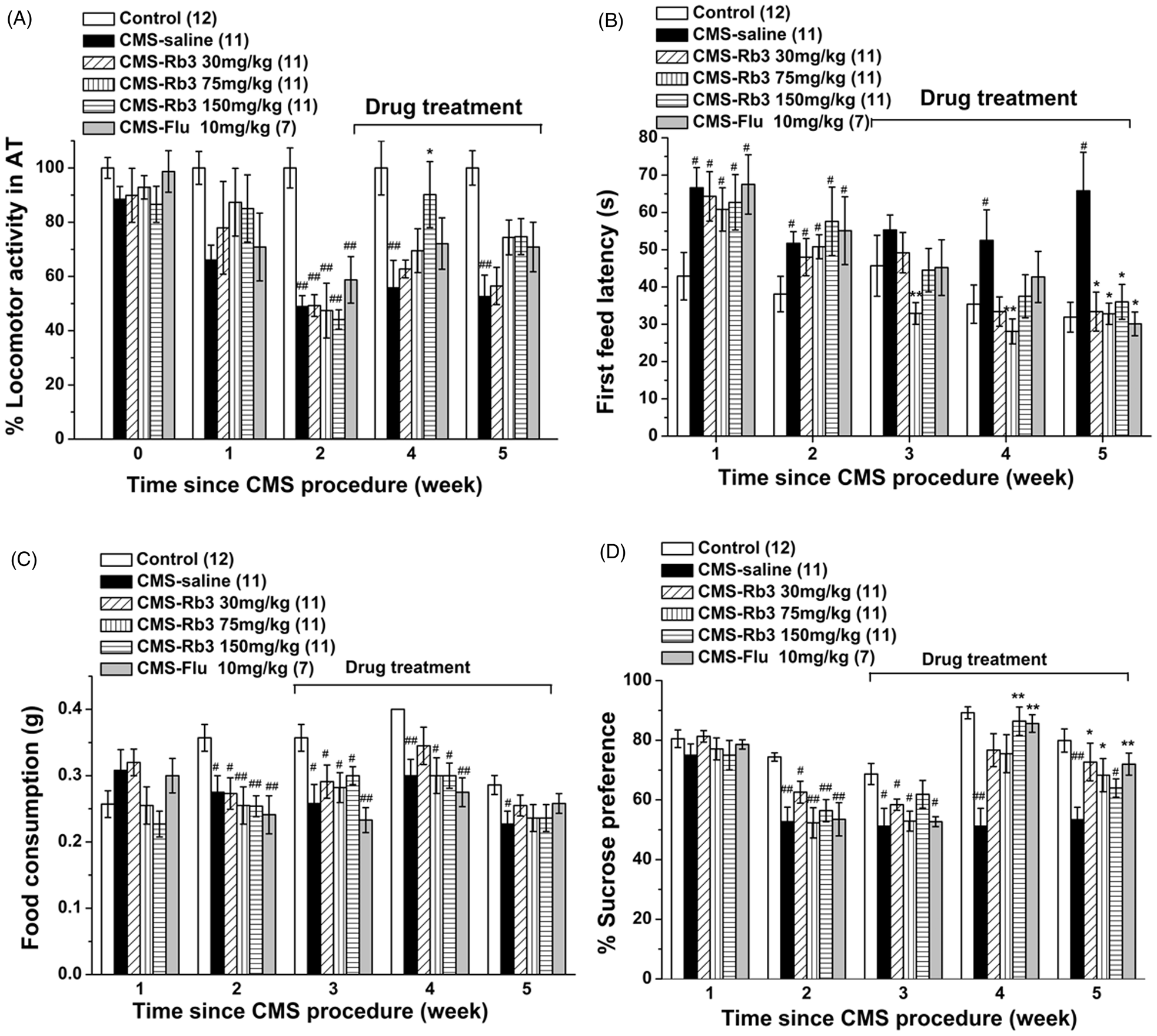
Effect of Rb3 on behaviors in the CMS procedure. Different stressors were applied for 5 successive weeks, and Rb3 was administered from week 3 onward. Several tests were performed each week. (A) Normalized number of movements of each mouse in the actimeter test (results of the control group were set to 100%). (B) First feeding latency in the novelty-suppressed feeding test in mice. (C) Food consumption in the novelty-suppressed feeding test in mice. (D) Sucrose preference in mice. The number of mice in each group is indicated in parentheses in each figure. Each column represents the mean ± SEM. *p < 0.05, **p < 0.01, compared with CMS–saline; # p < 0.05, ## p < 0.01, compared with control. AT, actimeter test; CMS, chronic mild stress; Rb3, ginsenoside Rb3; Flu, fluoxetine.
Effect of Rb3 on first feeding latency and food consumption in the NSFT
In the novelty-suppressed feeing paradigm, the first feeding latency (Figure 3B, Table 2) was prolonged significantly in mice in the CMS-saline group from week 1 to the whole procedure (one-way ANOVA and Bonferroni post-hoc test, p < 0.05 vs. control). In weeks 3 and 4 (the 1st and 2nd weeks of administration), 75 mg/kg Rb3 significantly reduced the first feeding latency, respectively (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. CMS-saline). In week 5, Rb3 (30, 75, and 150 mg/kg) and fluoxetine (10 mg/kg) significantly reversed the increase in the first feeding latency induced by CMS (one-way ANOVA and Bonferroni post-hoc test, p < 0.05 vs. CMS-saline). In contrast, food consumption (Figure 3C, Table 2) in the CMS-saline group was suppressed from week 2 to week 5 (one-way ANOVA and Bonferroni post-hoc test, p < 0.05 vs. control). In week 4, 30 mg/kg Rb3 increased food consumption to the level of control group (one-way ANOVA and Bonferroni post-hoc test, p > 0.05 vs. control). In week 5, none of the doses of Rb3 or fluoxetine significantly increased food consumption compared with the CMS-saline group (one-way ANOVA and Bonferroni post-hoc test, p > 0.05 vs. CMS-saline and control).
Effect of Rb3 on sucrose preference
Before the CMS procedure, no significant differences in sucrose preference were observed among groups (one-way ANOVA and Bonferroni post-hoc test, p > 0.05, one-way ANOVA, data not shown). After 2 weeks of exposure to different stressors, sucrose preference was significantly decreased in the CMS-saline group (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. control; Figure 3D, Table 2). In week 4, however, sucrose preference was remarkably restored by 150 mg/kg Rb3 to 86.4 ± 4.76% and by 10 mg/kg fluoxetine to 85.6 ± 2.94%, (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. CMS-saline). In week 5, sucrose preference in the 75 and 30 mg/kg Rb3 groups also increased significantly (one-way ANOVA and Bonferroni post-hoc test, p < 0.05 vs. CMS-saline), but the 150 mg/kg Rb3 group was not significantly different from the CMS-saline group (one-way ANOVA and Bonferroni post-hoc test, p > 0.05).
Effect of Rb3 on reversing hippocampal atrophy after stress
As shown in Figure 4A, CMS exposure significantly reduced hippocampal weight in the CMS-saline group (4.65 ± 0.496% of whole brain weight, one-way ANOVA and Bonferroni post-hoc test, p < 0.01) compared with control (7.07 ± 0.759% of whole brain weight). Hippocampal weight in the 75 and 150 mg/kg Rb3 and fluoxetine (10 mg/kg) groups increased to 6.15 ± 0.663%, 6.06 ± 0.311%, and 6.90 ± 0.746% of the whole brain weight, respectively (one-way ANOVA and Bonferroni post-hoc test, p < 0.05 vs. CMS-saline), but hippocampal weight in the 30 mg/kg Rb3 group did not show a significant increase.
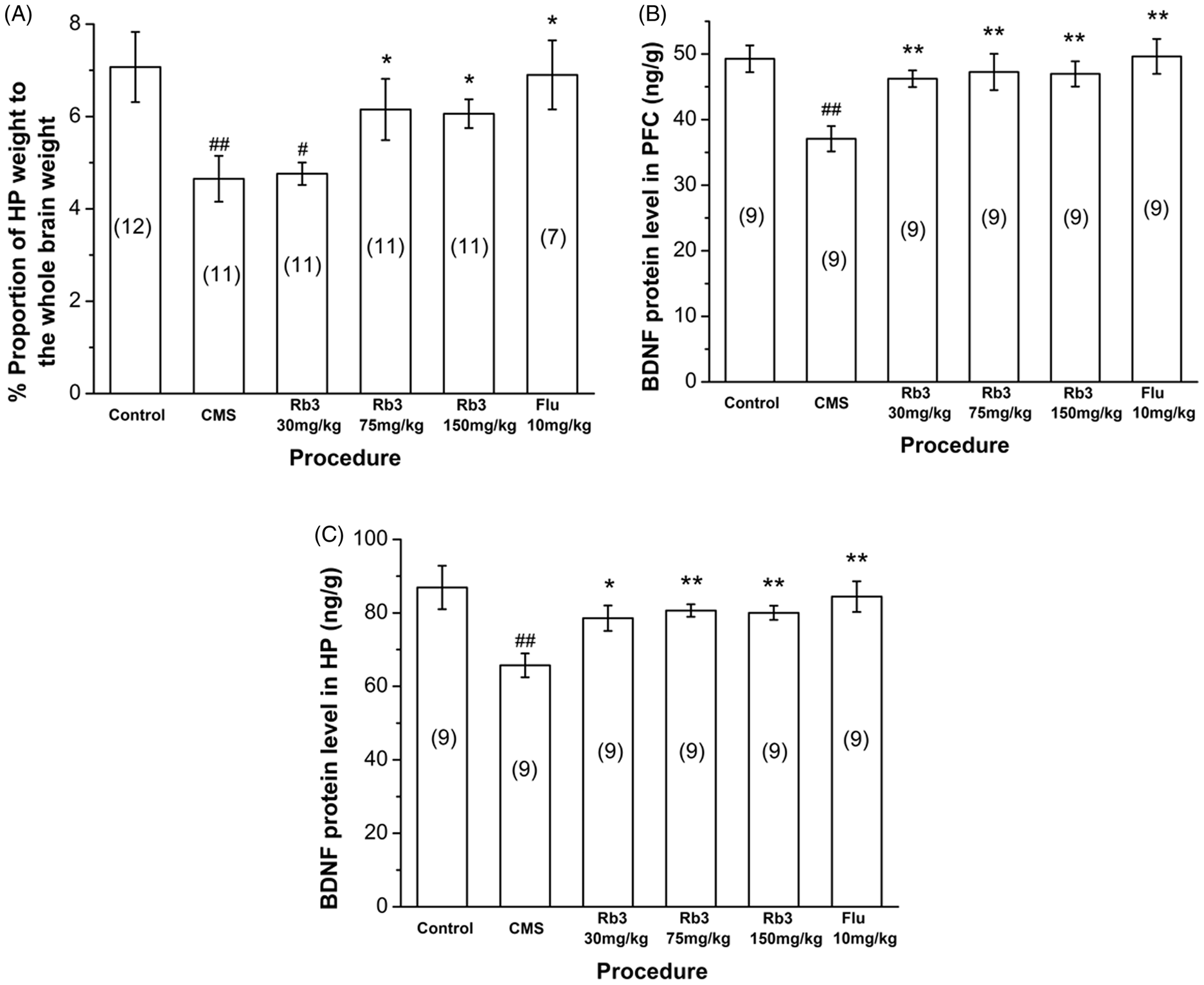
Effect of Rb3 on hippocampal weight and BDNF levels. (A) Hippocampal weight in different groups. (B, C) BDNF levels in the (B) prefrontal cortex and (C) hippocampus. The number of mice in each group is indicated in parentheses in each figure. Each column represents the mean ± SEM. *p < 0.05, **p < 0.01, compared with CMS; # p < 0.05, ## p < 0.01, compared with control. BDNF, brain-derived neurotrophic factor; HP, hippocampus; PFC, prefrontal cortex; CMS, chronic mild stress; Rb3, ginsenoside Rb3; Flu, fluoxetine.
Effects of Rb3 on BDNF protein levels
The results of the BDNF analysis are shown in Figures 4B and 4 C. The CMS procedure produced a significant decrease in BDNF levels in the prefrontal cortex and hippocampus (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. control), and drug administration elevated BDNF levels significantly in both regions (one-way ANOVA and Bonferroni post-hoc test, 30 mg/kg Rb3 in the hippocampus, p < 0.05 vs. CMS-saline; other groups, p < 0.01 vs. CMS-saline).
Effect of Rb3 on monoamine neurotransmitters in different brain regions
The levels of 5-HT, DA, and NE in different brain regions are shown in Figure 5. 5-HT levels (Figure 5A, Table 3) in the prefrontal cortex and amygdala were significantly reduced in the CMS-saline group compared with control group (one-way ANOVA and Bonferroni post-hoc test, prefrontal cortex, p < 0.05; amygdala, p < 0.01) after 5 weeks of CMS exposure. Treatment with 75 and 150 mg/kg Rb3 and 10 mg/kg fluoxetine significantly increased 5-HT levels in the prefrontal cortex by 80.8% (one-way ANOVA and Bonferroni post-hoc test, p < 0.05), 253.8% (p < 0.01), and 96.2% (p < 0.01), respectively, compared with CMS-saline group. 5-HT levels in the 150 mg/kg Rb3 group significantly exceeded control levels (one-way ANOVA and Bonferroni post-hoc test, p < 0.01) in prefrontal cortex. In the amygdala, 75 mg/kg Rb3 and fluoxetine increased 5-HT levels by 172.7% and 40.9%, respectively (p < 0.01 vs. CMS-saline), but in the 30 and 150 mg/kg Rb3 groups 5-HT levels did not markedly increase compared with CMS-saline. These results indicate a biphasic feature of Rb3 in regulating 5-HT concentrations in the amygdala. In the hippocampus, 75 mg/kg Rb3 decreased 5-HT levels (one-way ANOVA and Bonferroni post-hoc test, p < 0.05 vs. control). One-way ANOVA revealed no significant differences in 5-HT levels among the other groups (p > 0.05).
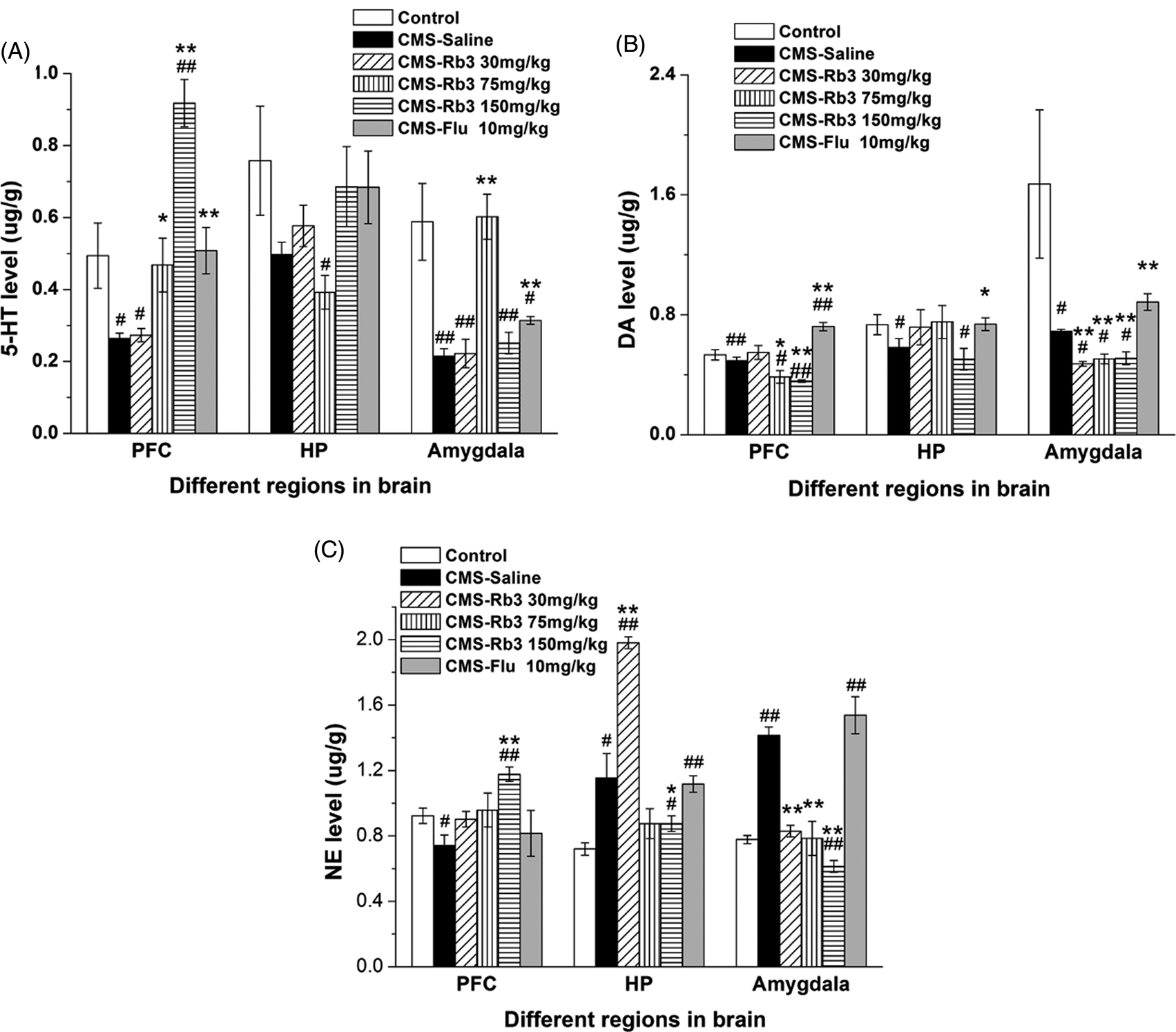
Effect of Rb3 on monoamine neurotransmitters in different brain regions after the CMS procedure in mice. (A) 5-HT levels, (B) dopamine levels, and (C) norepinephrine levels in the prefrontal cortex, hippocampus, and amygdala in different groups of mice. In each group, n = 5. Each column represents the mean ± SEM. *p < 0.05, **p < 0.01, compared with CMS–saline; # p < 0.05, ## p < 0.01, compared with control. 5-HT, 5-hydroxytryptamine; DA, dopamine; NE, norepinephrine; PFC, prefrontal cortex; HP, hippocampus; CMS, chronic mild stress; Rb3, ginsenoside Rb3; Flu, fluoxetine.
Effect of Rb3 on monoamine neurotransmitters in different brain regions after the CMS procedure in mice.
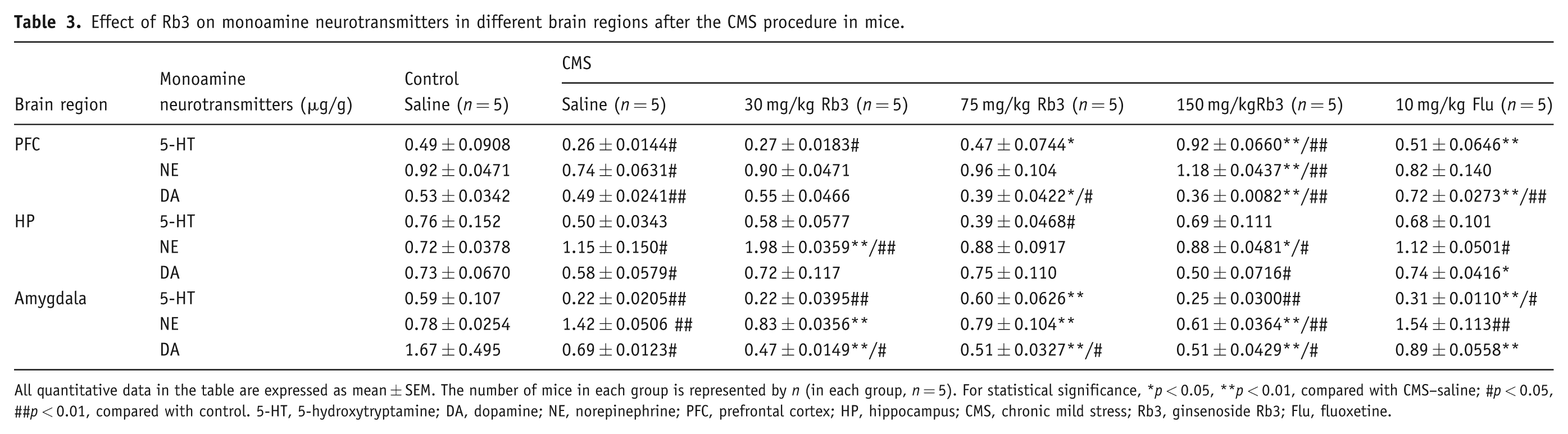
All quantitative data in the table are expressed as mean ± SEM. The number of mice in each group is represented by n (in each group, n = 5). For statistical significance, *p < 0.05, **p < 0.01, compared with CMS–saline; #p < 0.05, ##p < 0.01, compared with control. 5-HT, 5-hydroxytryptamine; DA, dopamine; NE, norepinephrine; PFC, prefrontal cortex; HP, hippocampus; CMS, chronic mild stress; Rb3, ginsenoside Rb3; Flu, fluoxetine.
Dopamine levels (Figure 5B, Table 3) in the three brain regions were significantly reduced in the CMS-saline groups compared with controls (one-way ANOVA and Bonferroni post-hoc test, prefrontal cortex, p < 0.01; amygdala and hippocampus, p < 0.05). In the prefrontal cortex, 75 and 150 mg/kg Rb3 further reduced dopamine levels by 20.4% (one-way ANOVA and Bonferroni post-hoc test, p < 0.05) and 26.5% (p < 0.01), respectively, compared with CMS-saline. However, fluoxetine increased dopamine level by 46.9% (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. CMS-saline). In the amygdala, 30, 75, and 150 mg/kg Rb3 reduced dopamine levels by 31.9%, 26.1%, and 26.1%, respectively, compared with CMS-saline (one-way ANOVA and Bonferroni post-hoc test, p < 0.01). Fluoxetine elevated dopamine levels by 29.0% (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. CMS-saline). In the hippocampus, 30 and 75 mg/kg Rb3 and fluoxetine (10 mg/kg) increased dopamine levels, but the effects were not significantly different from the CMS-saline and control groups (one-way ANOVA and Bonferroni post-hoc test, p > 0.05). The insignificant effect of 150 mg/kg Rb3 (one-way ANOVA and Bonferroni post-hoc test, p > 0.05 vs. CMS-saline; p < 0.05 vs. control) on dopamine levels appears to present a biphasic action of Rb3 on dopamine in the hippocampus.
Norepinephrine levels (Figure 5C, Table 3) in the CMS-saline group were reduced in the prefrontal cortex (one-way ANOVA and Bonferroni post-hoc test, p < 0.05 vs. control) and increased in the hippocampus (by 59.7%, p < 0.05 vs. control) and amygdala (by 82.1%, p < 0.01 vs. control). In the prefrontal cortex, 150 mg/kg Rb3 increased norepinephrine levels by 59.4% (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. CMS-saline). The 30 and 75 mg/kg Rb3 and fluoxetine (10 mg/kg) groups had increased norepinephrine levels that were not significantly different compared with the CMS-saline group or controls. In the hippocampus, 30 mg/kg Rb3 increased norepinephrine levels by 72.2% (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. CMS-saline) rather than reduced them. In contrast, 150 mg/kg Rb3 decreased norepinephrine levels by 23.5% compared with CMS-saline (one-way ANOVA and Bonferroni post-hoc test, p < 0.05). Treatment with 75 mg/kg Rb3 also decreased norepinephrine levels, but the effect was not significant compared with the CMS-saline and control groups. The effects of Rb3 on norepinephrine levels also appeared to be biphasic in the hippocampus. In the amygdala, 30, 75, and 150 mg/kg Rb3 reduced norepinephrine levels by 41.5%, 44.4%, and 57.0%, respectively (one-way ANOVA and Bonferroni post-hoc test, p < 0.01 vs. CMS-saline). Norepinephrine levels in the 150 mg/kg Rb3 group were notably lower than controls (one-way ANOVA and Bonferroni post-hoc test, p < 0.01).
Rb3 and PNTS changed the amplitude and frequency of action potentials
Figure 6 shows the action potentials of cortical neurons affected by Rb3 (30 µg/ml) and PNTS (125 µg/ml). The frequency and mean peak amplitude of the action potentials were significantly enhanced by Rb3 (amplitude from 55.52 ± 5.87 to 65.55 ± 3.43 mV, n = 4, Student’s t-test, p < 0.05; frequency from 42.60 ± 5.14 to 68.00 ± 6.48 Hz, p < 0.05 vs. control) and significantly reduced by PNTS (amplitude from 53.94 ± 3.20 to 37.63 ± 3.70 mV, n = 4, p < 0.01; frequency from 53.30 ± 7.27 to 39.14 ± 8.51 Hz, p < 0.01 vs. control).
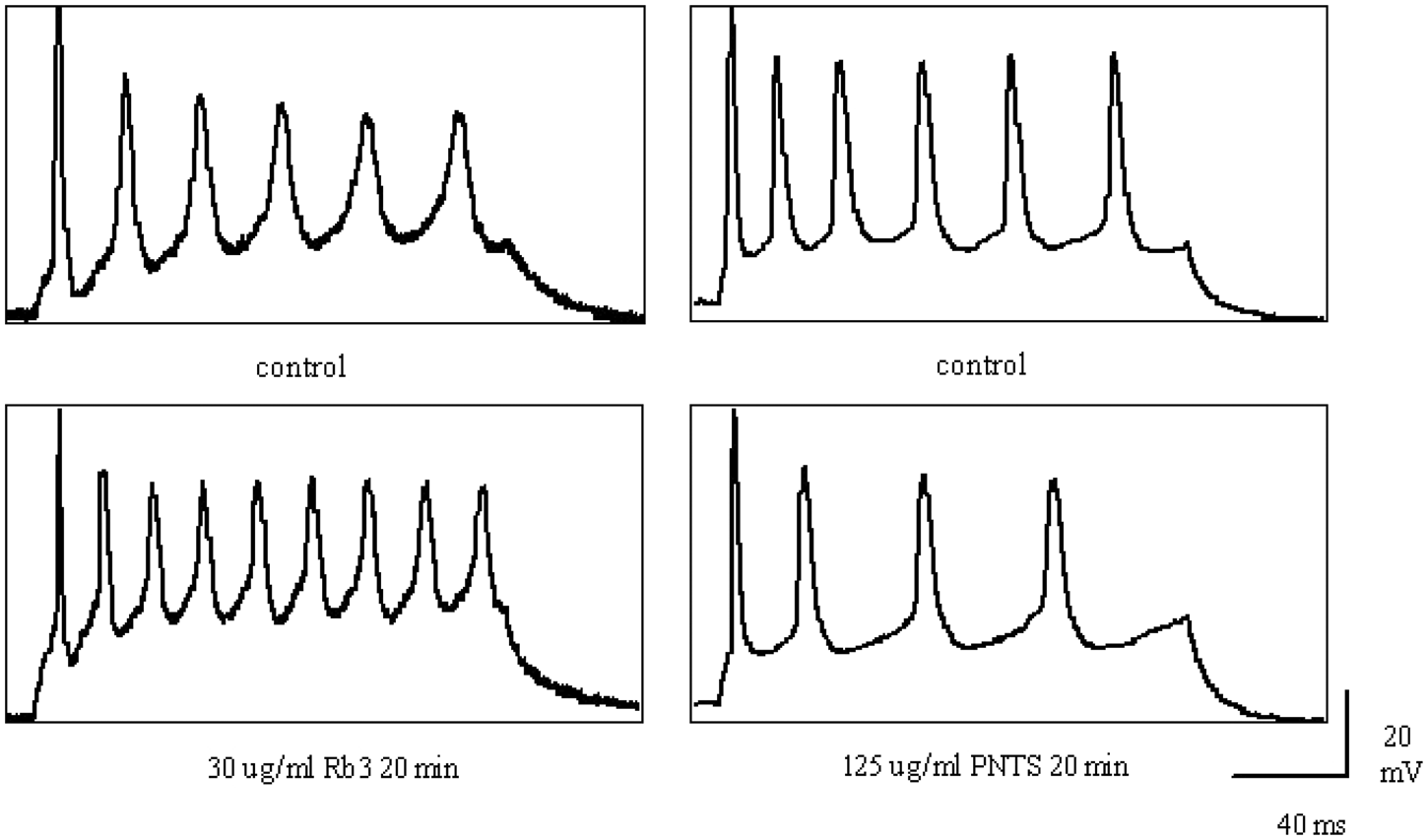
Effects of Rb3 and PNTS on action potentials in cortical neurons. Left panel: (upper) before and (lower) 20 min after 30 µg/ml Rb3. Right panel: (upper) before and (lower) 20 min after 125 µg/ml PNTS. PNTS, Panax notoginseng total saponins; Rb3, ginsenoside Rb3.
Discussion
Urgent demands exist for the development of effective and safe antidepressant medications. Ginsenosides are considered active constituents of Panax notoginseng and Panax ginseng (adaptogens for the therapy of stress disorders; Bhattacharya and Muruganandam, 2003). In the present study, one of these ginsenosides, Rb3, was explored for its potential antidepressant-like effects.
Rb3 has antidepressant effects in behavioral tests
After the preliminary OFT, which eliminated the possibility of psychomotor stimulant-like effects, acute intragastric administration of Rb3 dose-independently reduced immobility time in the FST and TST (Figure 1). Treatment with 75 mg/kg Rb3 had significant effects that were comparable with fluoxetine in the FST and TST, indicating a suitable dosage for a selective antidepressant effect of this medication. These results were consistent with previous studies which showed that ginseng total saponin decreased immobility time in the FST (Dang et al., 2009; Hu and Fu, 2003). In addition, our result concerning the antidepressant-like effect of fluoxetine in FST was not consistent with many other previous results (Dang et al., 2009). This may result from the difference of water temperature of the FST, which not only affects forced swimming behavior but also influences the individual susceptibility to antidepressant treatment (Arai et al., 2000; Bächli et al., 2008; Petit-Demouliere et al., 2005). In the present study, we controlled the water temperature at 20 ± 1°C, which differs from that in previous studies (24 ± 1°C). Our preliminary experiment found that the NIH mice showed longer immobility time with decreasing water temperature from 24°C to 20°C. To some extent, this implied that the degree of depression was more serious at 20°C than at 24°C, leading to an insignificant antidepressant-like effect of fluoxetine in the FST. Thus it could indicate that the antidepressant-like effect of 75 mg/kg Rb3 is much stronger and faster than 10 mg/kg fluoxetine in the FST. In addition, Kobayashi et al. (2008) also found that fluoxetine failed to be efficacious in the FST in CD-1 mice at 10, 20, 40 mg/kg; they attributed this result to the fact that the FST may detect compounds that are effective by altering activity in the noradrenergic system.
In the learned helplessness procedure, the degree of failure reduction positively correlated with Rb3 dose, and the drug’s effects were enhanced as the number of administration days increased (Figure 2). Furthermore, our results also demonstrated that the antidepressant-like effect of Rb3 is parallel to that of fluoxetine, as there were insignificant differences between the two.
Compared with the acute models, the CMS model has generally been thought to be the most promising model applied to the study of depression in animals because of its similarity to encounters in the daily lives of humans, as compared with the bare fears caused in acute models (Rasenick et al., 1996). In the present study, chronic treatment with Rb3 and fluoxetine markedly reversed depression-like behavior induced by CMS. Furthermore, our data in the CMS test demonstrated that more than 2 weeks of fluoxetine treatment was needed for a recovery of depression. In contrast to the gradual onset of action of fluoxetine, Rb3 (75 mg/kg) appeared to exert significant antidepressant effects after a shorter time. For the 75 mg/kg Rb3-treated CMS group, an apparent effect in the NSFT appeared as early as after 1 week of drug treatment.
Thus, the behavioral results above, either in the acute or chronic models, strongly suggest the antidepressant activity of Rb3.
Antidepressant effect of Rb3 may involve monoaminergic systems
Monoamine neurotransmitters in the central nervous system play a vital role in depression because most of the typical antidepressant medications exert their effects by preventing reuptake of neurotransmitters (mainly 5-HT) from presynaptic terminals, inhibiting monoamine oxidase from degrading neurotransmitters, or acting at receptors that modulate monoaminergic transmission (Liebelt, 2008). In the present model of the reserpine-induced syndrome, Rb3 dose-dependently reversed the decrease in body temperature, ptosis, and akathisia produced by reserpine, indicating that Rb3 could restore brain monoamine neurotransmitters. Previous studies have suggested a correlation between ptosis and monoamine depletion (Bourin et al., 1983). The biochemical tests further demonstrated that Rb3 may regulate neurotransmission. We chose the prefrontal cortex, hippocampus, and amygdala as the target sites because these regions are involved in regulating learning, memory, mood, recognition, and the hypothalamic–pituitary–adrenal axis. They are also considered the intersection of positive and negative networks that are crucially important in depression (Stone et al., 2007). Similarly, hyperforin, the main active ingredient in St. John’s wort, inhibits the reuptake of 5-HT, dopamine, norepinephrine, glutamate, and GABA (Di Carlo et al., 2001; Laakmann et al., 1998).
5-HT is involved in the regulation of several systems known to be disturbed in psychiatric disease. Central 5-HT in the nuclei raphae medianae regulates the following structures: (1) cortex, basal ganglia, and limbic system, which are all related to mood; (2) hypothalamus, which regulates appetite; and (3) hippocampus, which modulates sleep (Neumeister, 2003). Thus, reduced 5-HT neurotransmission has been reported to be one of the critical etiological factors in the development of depression, and enhanced 5-HT neurotransmission elicits a therapeutic response (Blier and de Montigny, 1994). In the present study, reversing effects of Rb3 and fluoxetine on CMS-induced reduction in 5-HT levels in the prefrontal cortex and amygdala support the hypothesis that CMS causes depression-like behavior through the 5-HT pathway. The enhancement of 5-HT by 150 mg/kg not only exceeds the 5-HT concentration in CMS-saline group, but also exceeds that in the control group. Many classical selective serotonin reuptake inhibitors (SSRIs), like fluoxetine, take part in antidepressant effects involving the downregulation of presynaptic serotonin autoreceptors and postsynaptic serotonin 5-HT2A receptors, the latter occurring when the antidepressant-like effect becomes apparent (Eison and Mullins, 1996; Gobbi et al., 1997); this is also the reason for adaptive process provoked by SSRIs. However, 150 mg/kg Rb3 may modulate different sites of the presynaptic and postsynaptic serotonergic system in a shorter time in the prefrontal cortex, leading to an over-increase of 5-HT. 5-HT in the hippocampus, however, failed to change significantly. This result is consistent with an earlier report in which no significant differences in 5-HT levels in the hippocampus, assessed by high-performance liquid chromatography, were found between the CMS and control groups (Dang et al., 2009). These researchers speculated that CMS-induced impairment of the serotonergic system may occur in other forms (for example, by affecting diverse types of receptors and transporters), rather than being based on the concentration of 5-HT between synapses.
Dopaminergic and noradrenergic systems, which perform tandem actions with the serotonergic system, also play an important role in depression-related behavior (Millan et al., 2000). Dopamine is involved in the reward system, which is closely connected with anhedonia, a typical symptom of depression. Studies have shown that antidepressants can also exert their effects through dopamine neurotransmission (Hamon and Bourgoin, 2006). Many data implicate prefrontal cortex–amygdala interactions in the regulation of affective behaviors (Pape, 2005). Dopamine is disinhibitory in the amygdala in vivo but excites local GABAergic neurons, whose activation provides inhibitory control of the amygdala (Marowsky et al., 2005). In the present study, however, continued administration with Rb3 for 3 weeks further reduced dopamine levels, which contrasts with some previous reports about ginseng total saponins on depression (Dang et al., 2009; Itoh et al., 1989). These results are consistent with Shim et al. (2000), in which ginseng total saponins completely blocked the increase in dopamine release in the nucleus accumbens induced by a systemic injection of nicotine, suggesting that Rb3 may exert antidepressant-like effects through pathways other than the dopamine–GABA pathway. Variations in norepinephrine concentrations were found in different brain regions after CMS. In the prefrontal cortex, a significant reduction in norepinephrine levels may cause memory loss and logical confusion and aggravate the depressive condition. This reduction could be reversed by Rb3, especially at the dose of 150 mg/kg, which not only returns norepinephrine levels to pre-stress levels, but actually causes levels to exceed basal values, possibly because of the compensatory increase in amine synthesis (Anisman et al., 2008). The present biochemical results also showed increased norepinephrine levels in the hippocampus and amygdala after CMS, which are not consistent with the previous study by Dang et al. (2009), who found a significant reduction of norepinephrine levels in the hippocampus reduced by CMS, but are consistent with Nemeroff (2002), who claimed that increased norepinephrine is postulated to be the most reliable neurochemical signal of depression. In addition, Tanaka et al. (2000) noted that a variety of stressful events cause a marked increase in norepinephrine release in several brain regions in rat, which suggests that an increased norepinephrine release may be closely related to the provocation of negative emotions such as anxiety and/or fear. Morilak et al. (2005) considered norepinephrine as an important modulatory factor in response to stress. We suggest that the antidepressant-like effect of Rb3 may primarily involve the function of noradrenergic pathways.
Antidepressant effect of Rb3 may involve neurotrophy and neurogenesis
Recently, the biological study of depression has moved beyond the monoamine hypothesis and to structures in different parts of the central nervous system. Neurotrophic factors have also received growing attention. Several studies have suggested that the hippocampus may be a critical structure in the etiology of depression because abnormalities in hippocampal volume have been demonstrated both experimentally and clinically (Dranovsky and Hen, 2006; Lucassen et al., 2006; Warner-Schmidt and Duman, 2006). The mechanisms of this influence may involve glucocorticoids, serotonin, and excitatory amino acids (Aan het Rot et al., 2009). Most notable with regard to neurotrophy are studies of BDNF (Aydemir and Deveci, 2009). Hippocampal BDNF mRNA levels drop significantly in CMS models. This effect can be reversed by antidepressants (Rasmusson et al., 2002; Shirayama et al., 2002), possibly through a cascade response related to phosphorylation of cyclic adenosine monophosphate response element-binding protein in the second messenger pathway (Poo, 2001). In the present study, a high dose of Rb3 (150 mg/kg) significantly recovered hippocampal volume. BDNF expression was also increased by Rb3 at all of the doses tested, similar to the effects of fluoxetine, which has been shown to upregulate BDNF expression (De Foubert et al., 2004; Molteni et al., 2006). These results are consistent with those of Dang et al. (2009), in which ginseng total saponins exerted antidepressant-like effects by increasing BDNF expression in the hippocampus.
Dose-related antidepressant-like effects of Rb3
Based on our results from the behavioral and neurochemical experiments, three doses of Rb3 showed dose-related antidepressant-like effects, including more or less linear dose-related effects and biphasic dose-related effects. The more or less linear dose-related effects primarily happened at concentrations from 30-75 mg/kg Rb3 in the FST and TST, and from 30-150 mg/kg Rb3 in the learned helplessness test and reserpine-induced test. In addition, BDNF concentration in the hippocampus and prefrontal cortex, as well as the volume of hippocampus, were dose-dependently elevated by Rb3.
Beside the linear dose-related effects, there are biphasic dose-related effects shown in our research. The biphasic feature of Rb3 in regulating 5-HT concentrations in the amygdala, NE and DA concentrations in the hippocampus and DA concentrations in the prefrontal cortex may result from many factors. Firstly, it is possible that different monoamine neurotransmitters in different brain areas were sensitive to Rb3 diversely. The results showed that the three monoamine neurotransmitters in the hippocampus all increased significantly by 30 mg/kg Rb3 and decreased significantly by 150 mg/kg Rb3. This indicated that Rb3 at low concentration primarily influences the neurochemical properties of the hippocampus, but an increase of Rb3 could impact diverse brain regions widely, which led to a decentralization of the major influence on hippocampus. Secondly, another reason may be the presence of other ingredients in the dosage form. In our research, the purity of Rb3 is 90% and some other types of ginsenoside such as Rb1, Rc are present as impurities. Yue and Zhong (2008) indicated that structurally similar ginsenosides may have different or even totally opposite effects, which may lead to the biphasic dose-related phenomenon shown in our results. Finally, Jiang et al. (2007) researched the effects of different concentrations of Rb3 on persistent Na+currents in ischemic and normal neurons, and found that Rb3 could inhibit persistent Na+ currents in a biphasic dose-related manner. Inhibiting the persistent Na+ currents could lead to a decrease in the activities of neurons, which play an important role in synaptic communication and release of neurotransmitters. Thus our result concerning the changes of 5-HT in the hippocampus by Rb3 was consistent with the previous study on the effects on the persistent Na+ currents of different concentrations of Rb3.
Antidepressant-like effect of Rb3 involves neuronal excitability
Rb3 has long been studied for its protective effect on ischemic neurons. Although its mechanism has not yet been ascertained, some researches have suggested that this protection might involve inhibition of persistent Na+ currents in rat hippocampal neurons (Jiang et al., 2007). This inhibition of Na+ currents may be achieved by NMDA-induced inhibition of Ca2+ current influx into rat hippocampal neurons (Shen et al., 2008). Accumulating evidence has demonstrated the involvement of intracellular Ca2+ in the pathophysiology of mood disorders (Galeotti et al., 2006). Holderbach et al. (2007) found a slightly increased frequency and significantly increased duration of action potentials, a measure of neuronal excitability, synaptic plasticity, and information transmission. The characteristics of action potentials depend on ion transport, especially Na+ during the depolarization period and K+ during the repolarization period, thus reflecting ion currents. In addition, previous research has shown that somatosensory cortex can respond to chronic antidepressant treatment by undergoing neuronal structural plasticity, which connects closely with neuronal network function (Guirado, et al., 2009; Lee, 2009), so we focused on the characteristics of action potentials of pyramidal neurons in the somatosensory cortex of rats for testing the antidepressant effects of Rb3 in the present research. In the present study, 30 µg/ml Rb3 increased action potential frequency and amplitude, indicating an increase in neuronal excitability and action potential transmission, which might favor synaptic plasticity. The excitement also helps to prevent the facilitation of long-term depression, which could be induced by CMS.
Rb3 and PNTS: similarities and differences
According to Li and Fitzloff (2002) and Sengupta et al. (2004), ginsenosides contain several active components that can be generally divided into two major groups: tranquilizing and exciting. Previous studies in our laboratory showed that PNTS could modulate BDNF and monoamine neurotransmitters to exert significant antidepressant-like effects in animal models including CMS and the FST (Xiang, et al., 2011). Ginsenoside Rb3, the main component of PNTS in the present study, has some similarities with PNTS in reversing the behavioral and neurochemical changes induced by depression in animal models.
With respect to ion fluxes, our research obtained opposite results from the treatment of neurons with Rb3 or PNTS with regard to action potential amplitude and frequency (Figure 6), suggesting different electrophysiological modulatory mechanisms in reversing depression. A previous study by our lab members (Xiang et al., 2011) showed that PNTS could mitigate against excitatory amino acid- or neurotoxin-induced neuronal cell damage by blocking Ca2+ overload. Ca2+ concentration contributes to the peak amplitude of action potential, so our result concerning the effects of PNTS on action potential is accordance with above conclusion; thus, it is possible that PNTS produce antidepressant-like effects through a neuroprotective role by inhibiting the action potential to decrease the release of Ca2+. On the other hand, synaptic deficiency is generally accepted to be involved in major depression, and accordingly antidepressants may exert their effects through enhancing synaptic efficiency, as determined by many factors such as action potential duration, frequency, and so on (Bean, 2007). The research by Wang et al. (2010) showed that hypericin, one of the major active constituents of extracts of St. John’s wort with antidepressive actions, could increase neuronal action potential duration to enhance synaptic efficiency, which contributed to its antidepressant effects. In our present research, it is possible that the increase the frequency and amplitude induced by Rb3 could also lead to a dramatic increase in synaptic efficiency. Treatment with Rb3 acutely could influence the function of the synaptic network to perform an antidepressant-like effect directly; in contrast, acute administration of PNTS may play a major role in sustaining the structure’s integrity which leads indirectly to reversing depression.
In summary, both Rb3 and PNTS exhibit antidepressant-like effects through multiple pathways and multiple loci. Furthermore, our findings shed new light on the pathogenic mechanism of depression in the adrenergic pathway, which may provide deeper understanding of depression prevention.
Footnotes
This work was supported by grants from the Guangdong Science-Tech Program (2001331004202516 and 2007B031400001), Cooperation of Industry, Education and Academy between Gongdong Province Government and the Chinese Government Education Department (2008B090500028), National Talented Person Foundation of China (J0730638), Science Foundation of Life Sciences School, and Opening Laboratory Fund, Sun Yat-Sen University.
The authors declare that they have no conflict of interest.