
Abstract
Desoxypipradrol is a methylphenidate-like drug that has been recently found in a number of ‘legal highs’. Evidence from emergency room toxicology reports suggests that this drug might have led to a number of psychotic events in drug abusers in the UK and elsewhere. However, very little research has been done on the effects of this drug on the brain. Here we used rat brain slices from the nucleus accumbens core, which were exposed to either cocaine (1, 3 or 10 µM) or desoxypipradrol (1, 3 or 10 µM) for 60 min. Dopamine efflux was electrically evoked and recorded using fast cyclic voltammetry. Both drugs increased the peak dopamine efflux and also slowed dopamine re-uptake. Desoxypipradrol was more potent than cocaine causing a sevenfold increase in peak dopamine levels (versus a threefold increase for cocaine) and increasing dopamine re-uptake half-life 15-fold (versus fivefold for cocaine). These data suggest that desoxypipradrol is more potent than cocaine at dopamine terminals, and this could account for its psychotogenic effects.
Introduction
Desoxypipradrol (deoxypipradrol, 2-(diphenylmethyl)piperidine, 2-benzhydrylpiperidine) has a similar structure to methylphenidate, a drug commonly used for attention deficit hyperactivity disorder (ADHD) in the USA, and was developed for the treatment of ADHD and narcolepsy by CIBA (now Novartis) in the 1950s. Methylphenidate was ultimately favoured because of its shorter half-life. Desoxypipradrol has also been tested to expedite recovery from anaesthesia (Bellucci, 1955). There is a paucity of published research on this drug. Searches on Medline and PubMed revealed only six publications, and five of those are from before 1982. Most of these papers show that desoxypipradrol blocks dopamine or noradrenaline uptake, and acts as a vasoconstrictor. There are no data on the effects of this drug in vivo or in brain slices, or on dopamine efflux within the nucleus accumbens.
Desoxypipradrol is now commonly found in some pills or powders which were ‘legal highs’ or ‘research chemicals’, but perhaps more accurately described as new psychoactive substances (e.g. ‘Ivory wave’, ‘Lunar wave’ and ‘Whack’), leading to a number of medical emergencies and in November 2010 it received a UK import ban by the Home Office. The UK Advisory Council on the Misuse of Drugs (ACMD) reported that this drug can evoke psychotic symptoms (ACMD, 2010). In a case series of 30 patients presenting to the Royal Infirmary of Edinburgh with toxicity associated with desoxypipradrol, patients demonstrated a prolonged toxidrome characterised by tachycardia, tachypnoea, dystonia, leucocytosis, rhabdomyolysis, agitation, paranoia and hallucinations (unpublished data).
In an effort to shed more light on the pharmacology of this drug, in particular its potential abuse liability, we have examined the effects of desoxypipradrol on electrically stimulated dopamine efflux in the rat accumbens brain slice and compared its potency to cocaine. We used the technique of fast cyclic voltammetry (FCV), which allows us to measure dopamine release and re-uptake in real-time with sub-second time resolution.
Methods and materials
Animals
We used 8-week-old male Wistar rats kept on a 12/12 h light/dark cycle. Food and water were freely available and the rats were housed six per cage. Rats were killed by cervical dislocation (with no anaesthesia) in accordance with UK Home Office regulations.
Brain slice preparation
The brain was rapidly removed to produce a block containing the accumbens (from approximately −5 to +3 mm versus bregma) (Paxinos and Watson, 1986) that was attached to the chuck of a vibratome, using cyanoacrylate glue, and this was immersed in a bath containing ice-cold artificial cerebro-spinal fluid (aCSF). Coronal slices (400 µm) at the level of the accumbens (+1.0 to +2.2 mm versus bregma) were cut and transferred to a slice saver, comprising a plastic mesh suspended in a 500 mL container of aCSF continually bubbled with 95% O2/5% CO2. Once cut, slices were left to equilibrate for 1–4 h at room temperature (22 ± 1°C), to allow recovery from any trauma associated with slicing. Following equilibration, slices were transferred to the brain slice chamber which was heated by a thermostatically controlled circulating water bath (Grant, Cambridgeshire, UK). The slice chamber was superfused continuously with aCSF (100 mL/h) and the temperature in the slice chamber was measured before and after each experiment with a CEM DT-613 digital thermometer with K-type thermocouple probe. The temperature in the brain slice chamber was 32.5 ± 0.5°C. The slice was equilibrated for 45 min prior to electrical stimulation. The composition of aCSF (in mM) was: NaCl (126.0), KCl (2.0), KH2PO4 (1.4), MgSO4 (2.0), NaHCO3 (26.0), CaCl2 (2.4), (+)- glucose (10.0), bubbled for at least 60 min with 95% O2/5% CO2.
Fast cyclic voltammetry
Carbon fibre electrodes were made by inserting an 8 µm-diameter carbon fibre into a 10 cm-length borosilicate glass capillary (o.d: 2.0 mm, i.d.: 1.16 mm; Harvard Apparatus LTD, Kent, UK), which was pulled to a fine tip using an electrode puller (Model P-30; Sutter Instruments Co, Novato, CA, USA), such that the carbon fibre protruded from the pulled tip. The carbon fibre was then cut to a length of ~50 µm using a scalpel under microscopic guidance.
Immediately after the slice was placed in the slice chamber (i.e. at the start of the 45 min equilibration period), the tip of the microelectrode was placed into the accumbens core, at a depth of approximately 75 µm below the slice surface, using a micromanipulator and under microscopic guidance. This allowed us to monitor the stability of the brain slice, as on some occasions (e.g. poor slice health) the slice can spontaneously release large amounts of dopamine (Davidson et al., 2011). The bipolar tungsten stimulating electrode was placed ~200 µm away from the recording electrode with the tips inserted into the slice to ~100 µm using a micromanipulator. Auxiliary (stainless steel) and reference (Ag/AgCl) electrodes were placed at a convenient location in the slice chamber, away from the working electrode.
Voltammetric scans (–1.0 to +1.4 V vs Ag/AgCl, 480 V/s; see Figure 1A) were performed at a sampling frequency of 8 Hz using a Millar Voltammetric Analyser (PD Systems, West Molesey, UK), with complete voltammetric scans at the electrode displayed using a digital storage oscilloscope (Nicolet 310 DD; TekNet Electronics, Atlanta, GA, USA) and recorded digitally (Figure 1B). The signal was digitised and recorded using a CED1401 Micro3 and analysed using Spike2 v7.06 software (both Cambridge Electronic Design, Cambridge, UK). An increase in the current signal at 600–700 mV (Figure 1C), together with a corresponding reduction peak at −200 mV, were characteristic of dopamine detection in the accumbens (Robinson et al., 2003).
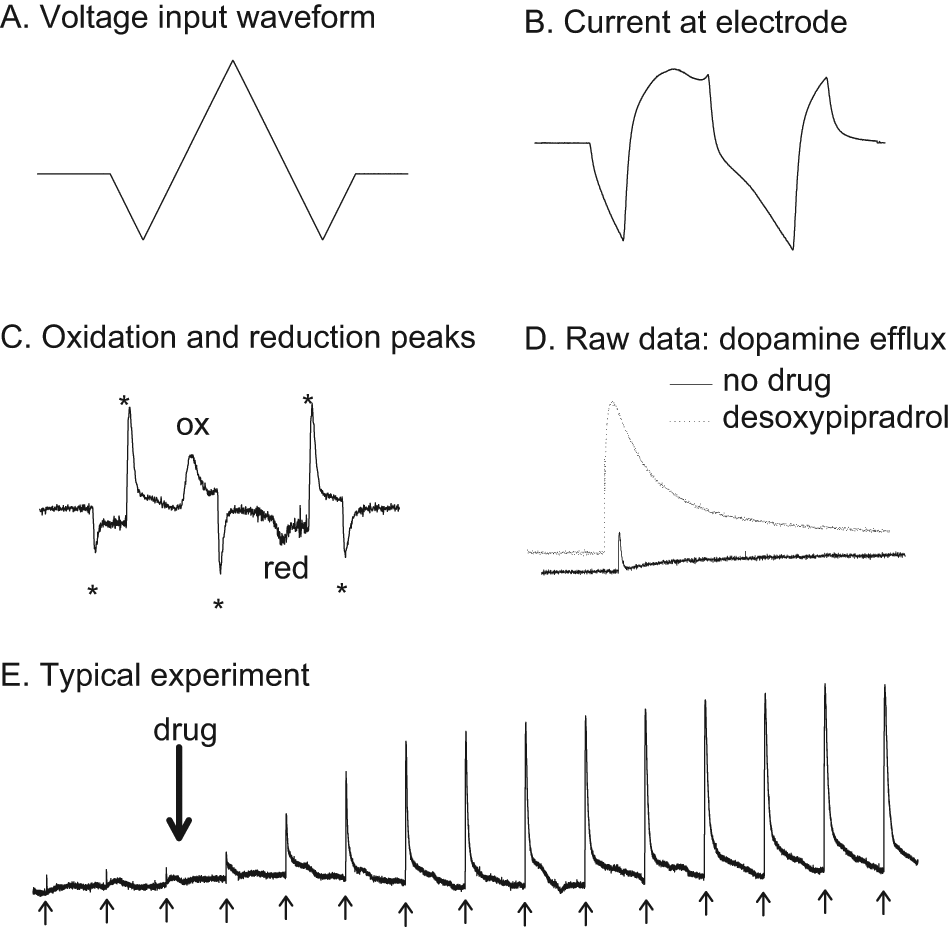
Fast cyclic voltammetry methodology and raw data. A. shows the input voltage waveform to the carbon fibre electrode. The applied voltage consists of a triangular waveform, going from 0 to −1 then up to +1.4, back to −1 and then back to 0 V. The rate of change of voltage was 480 V/s and the whole voltage scan took 20 ms. B. shows the current evoked by the input voltage at the carbon fibre electrode. When dopamine is present there is an increase in current due to the oxidation of dopamine at 600 mV. C. A subtracted voltammogram; the signal in the absence of dopamine is subtracted from the signal in the presence of dopamine (evoked dopamine efflux after 10 µM desoxypipradrol), leaving only the signal due to the oxidation and reduction of dopamine (the Faradaic current). Note the oxidation and reduction peaks at 9 ms (600 mV) and 13 ms (–200 mV). The other ‘peaks’ are just artefacts (denoted by *) that are commonly found and arise from the voltage input changing polarity. D. Typical electrically evoked dopamine release events for control (no drug) and in the presence of 1 µM desoxypipradrol. E. A typical experiment over the course of 70 min. The first three smaller events are baseline dopamine efflux events, with the drug (in this case desoxypipradrol 1 µM) added immediately after the third baseline stimulation for 60 min. We stimulated every 5 min, and the stimulations are denoted by the upward arrows. Note that desoxypipradrol has no effect on basal dopamine efflux, but does increase electrically stimulated levels.
Stimulation procedure
We used bipolar tungsten electrodes, tips about 300 µm apart to locally stimulate the core of the accumbens. Pseudo-one pulse stimulation (Singer, 1988) was used in order to avoid autoreceptor activation, which occurs at ~500 ms after striatal dopamine release (Lee et al., 2002; Phillips et al., 2002). The stimulus train was 10 × 1 ms pulses 10 ms apart (100 Hz), and we used a constant current of 10 mA with a Neurolog NL800 stimulus isolator (Warner Instruments, Hamden, CT, USA). Pulses were generated by an isolated pulse stimulator (Model 2100; A-M Systems, Sequim, WA, USA).
Fast cyclic voltammetry data analysis
Analysis of data was performed offline using Spike2 v7.06 (Cambridge Electronic Design). Prior to each experiment, the electrode was calibrated in 10 µM dopamine (made up in aCSF) and the signal measured during the experiments was then converted to dopamine concentration. We recorded two measures: 1) peak dopamine efflux (measured from pre-stimulation to dopamine peak after electrical stimulation) and 2) the re-uptake half-life, which was measured by fitting an exponential decay through the re-uptake phase of the dopamine efflux event (i.e. from peak dopamine efflux to baseline) (Yorgason et al., 2011).
Chemicals
All chemicals used were supplied by Sigma Chemicals (Poole, UK) except for racemic desoxypipradrol, which was synthesised and supplied by Dr Roland Archer, Department of Pharmacy and Chemistry, Kingston University, London, UK.
Statistics
Statistical analysis was carried out using SigmaPlot v. 11.0 (SigmaPlot, San Jose, CA, USA). For tests within either cocaine or desoxypipradrol data we used one-way ANOVA (independent variable = concentration) with post hoc Tukey’s. For comparison between cocaine and desoxypipradrol we used two-way ANOVA (independent variables were concentration and drug) followed by post hoc Tukey’s. In all graphs data are presented as means ± SEM, and significance is set at p <0.05.
Results
Baseline data
Brief (10 pulses at 100 Hz [90 ms]) local electrical stimulation reliably evoked dopamine efflux in the accumbens core region (Figure 1D, E), as previously described (Davidson and Stamford, 1993). Peak dopamine release was 367 ± 35 nM and re-uptake half-life was 0.97 ± 0.19 s (n = 33).
Basal release
Neither cocaine (data not shown) nor desoxypipradrol increased basal levels of dopamine efflux (Figure 1E); on the addition of desoxypipradrol there was no increase in the background signal (i.e. desoxypipradrol only increased electrically evoked dopamine).
Cocaine and desoxypipradrol effects on electrically evoked dopamine
We performed a one-way ANOVA on peak dopamine release data at 60 min after application of the drug (i.e. at the maximum drug effect).
Peak dopamine release after cocaine
Cocaine significantly increased peak dopamine release (F[3, 22] = 15.88, P < 0.001). Student–Newman–Keul analysis revealed that both 1 and 10 µM cocaine increased peak dopamine efflux to a greater extent than controls (P < 0.05), while 10 µM cocaine caused a significantly greater increase than both 1 and 0.1 µM cocaine (P < 0.05). No significant effect on peak dopamine efflux versus controls was recorded for 0.1 µM cocaine (P = 0.052; Figure 2A).
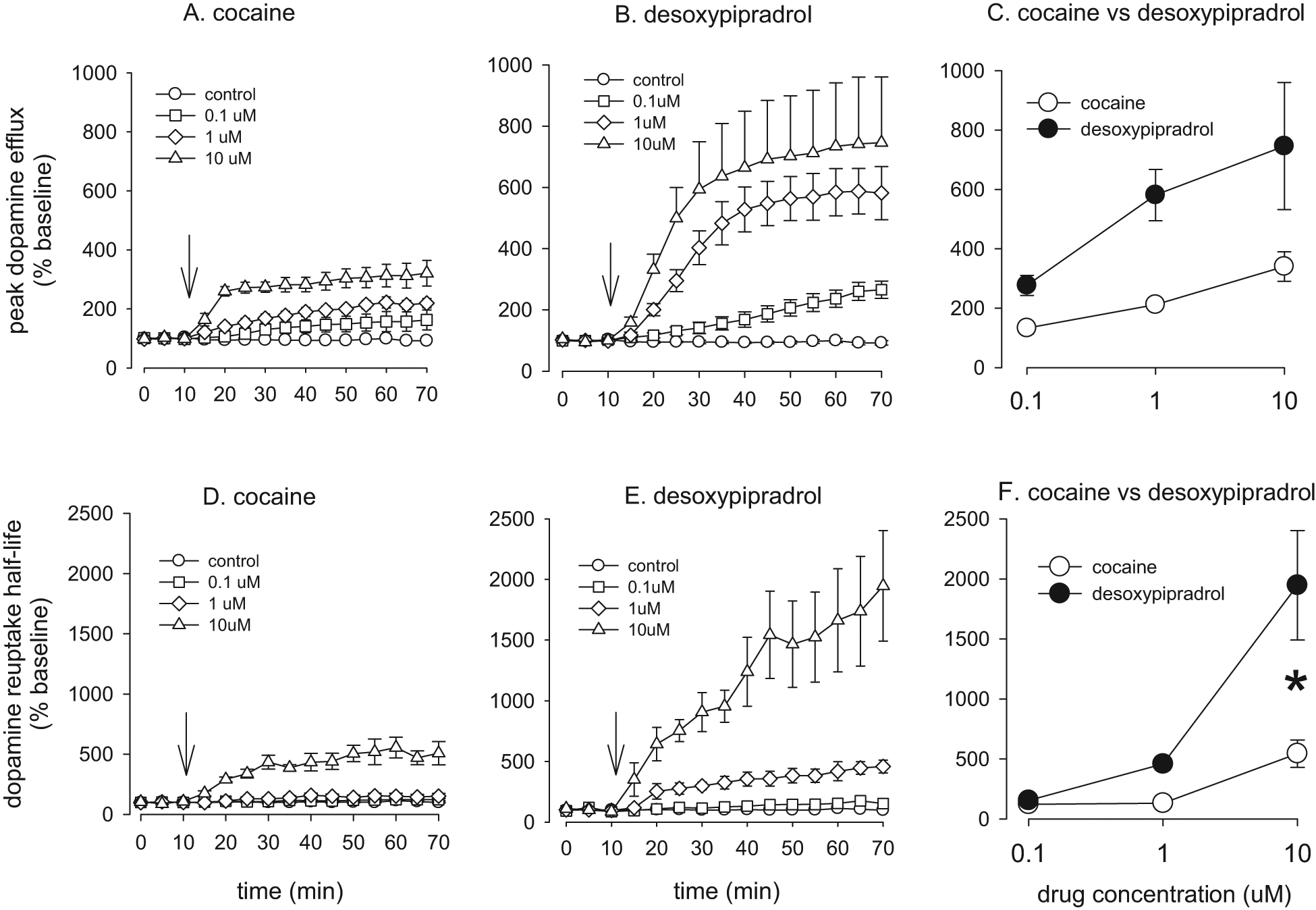
The effect of cocaine and desoxypipradrol on evoked dopamine efflux in the nucleus accumbens core. A. Effect of cocaine on peak evoked dopamine efflux over 60 min. B. Effect of desoxypipradrol on peak evoked dopamine efflux over 60 min. C. Comparison of cocaine with desoxypipradrol on peak evoked dopamine efflux. D. Effect of cocaine on dopamine uptake half-life over 60 min. E. Effect of desoxypipradrol on dopamine uptake half-life over 60 min. F. Comparison of cocaine with desoxypipradrol on dopamine uptake half-life. Values are means ± SEM, n = 5 for all groups except 10 µM desoxypipradrol (n = 3). Downward arrows denote when the drug was added (just after the third baseline stimulation).
Dopamine re-uptake half-life after cocaine
There was a significant effect of cocaine concentration on dopamine re-uptake half-life (F[3, 22] = 19.011, P < 0.001; Figure 2D). Pairwise multiple comparisons with the Student–Newman–Keul method revealed that 10 µM cocaine significantly increased dopamine re-uptake half-life versus all other concentrations (P < 0.05).
Peak dopamine release after desoxypipradrol
Desoxypipradrol significantly increased peak dopamine release (F[3, 20] = 16.68, P < 0.001). Student–Newman–Keul analysis revealed that both 1 and 10 µM desoxypipradrol increased peak dopamine efflux to a greater extent than 0.1 µM (P < 0.05) and controls (P < 0.05). 0.1 µM desoxypipradrol had no significant effect on peak dopamine efflux versus controls (P = 0.076; see Figure 2B).
Dopamine re-uptake half-life after desoxypipradrol
These data failed the normality test and so we performed a Kruskal–Wallis one-way ANOVA on ranks. There was a significant effect of desoxypipradrol concentration on dopamine re-uptake half-life (H[3] = 18.338, P < 0.001). Pairwise multiple comparisons with Dunn’s method revealed that both 1 and 10 µM desoxypipradrol significantly increased dopamine re-uptake half-life versus controls (P < 0.05; see Figure 2E).
Desoxypipradrol versus cocaine
Peak dopamine release
Two-way ANOVA revealed that there was a main effect of drug concentration (F[2, 27] = 9.85, P < 0.001) and a main effect of drug (F[1, 27] = 25.90, P < 0.001) but no significant interaction (F[2, 27] = 2.92, P = 0.075). Thus increasing concentrations of cocaine or desoxypipradrol increased peak dopamine release and desoxypipradrol increased peak dopamine release to a greater extent than cocaine (see Figure 2C).
Dopamine re-uptake half-life
Two-way ANOVA revealed that there was a main effect of drug concentration (F[2, 27] = 41.48, P < 0.001) and a main effect of drug (F[1, 27] = 34.72, P < 0.001), and a drug by concentration interaction (F[2, 27]) = 16.86, P < 0.001; Figure 2F). Thus increasing concentrations of cocaine or desoxypipradrol increased dopamine re-uptake half-life and desoxypipradrol increased dopamine re-uptake half-life to a greater extent than cocaine. Further, Tukey’s test revealed that although there was no significant difference between cocaine and desoxypipradrol at either 0.1 µM (P = 0.856) or at 1 µM (P = 0.077), 10 µM desoxypipradrol had a significantly greater effect on dopamine re-uptake than the same concentration of cocaine (P < 0.001).
Discussion
Most, if not all, drugs of abuse increase dopamine efflux in the nucleus accumbens (DiChiara and Imperato, 1988; Volkow et al., 2007), whether by a direct effect on the dopamine terminals or more indirectly at the dopamine cell body region, the ventral tegmental area. As a dopamine re-uptake inhibitor, desoxypipradrol should increase dopamine efflux directly at the dopamine terminals and we have shown that this drug is a very potent dopamine re-uptake inhibitor. By comparing the effects of desoxypipradrol with cocaine, one of the most highly addictive drugs, we have shown that desoxypipradrol is more potent than cocaine and could therefore be a highly addictive compound. The data presented here also suggest that desoxypipradrol is a ‘cocaine-like’ drug as opposed to an ‘amphetamine-like’ drug in that it has a large effect on stimulated dopamine efflux, but no effect on basal levels (see Figure 1E). Amphetamine-like drugs are able to evoke dopamine efflux in the absence of electrical stimulation in brain slices by ‘reverse transport’ (Schmitz et al., 2001). It is possible that desoxypipradrol can increase basal levels of dopamine, although if it does then these levels would be very low (<50 nM) and below the detection limit of this technique.
It could be argued that desoxypipradrol not only increases dopamine efflux but also noradrenaline efflux within the nucleus accumbens. Studies examining tissue content have found that there is negligible noradrenaline compared with dopamine within the accumbens core (Allin et al., 1988) and there are virtually no noradrenaline-containing nerve terminals within the accumbens core (Swanson and Hartman, 1975; Gaspar et al., 1985; Berridge et al., 1997). Furthermore, we have previously shown that the selective noradrenaline re-uptake inhibitor desipramine has no effect on electrically evoked amine efflux in rat accumbens core brain slices (Lee et al., 2001). There is recent evidence that the caudal accumbens shell region contains relatively high levels of noradrenaline, and amine efflux in this area can be increased by noradrenergic selective agents (Park et al., 2010); however, in the present study all the work was done in the accumbens core.
Previous research into desoxypipradrol has shown that it is a potent vasoconstrictor. Maxwell et al. (1970) found desoxypipradrol to potently inhibit noradrenaline re-uptake in a rabbit aortic strip preparation; they also found the R-(-)-enantiomer to be more potent than the S-(+)-enantiomer. Ferris et al. (1972) were the first to show that desoxypipradrol was a potent inhibitor of catecholamines in neuronal tissue, including the striatum, using rat synaptosomal preparations and labelled transmitters. However others have found pre-loading tissue with labelled transmitters to be taken up by different dopamine pools (Herdon et al., 1985). Furthermore, the ‘striatal’ samples used by Ferris and co-workers were taken from a block containing the caudate, putamen and globus pallidus. We have examined the effects of desoxypipradrol in the accumbens, where cocaine was found to exert a greater effect than in the caudate (Cass et al., 1992). Ferris and Tang (1979) also examined the effects of desoxypipradrol on [3H]dopamine and norepinephrine re-uptake using striatal and hypothalamic synaptosomal preparations, respectively. They reported an IC50 value of around 0.7 µM for the dopamine transporter compared with 20 µM for norepinephrine transporters. Using the same assay this group reported an IC50 value of 40 nM for desoxypipradrol at striatal dopamine transporters (Eckhardt et al., 1982). More recently some analogues of desoxypipradrol have been examined at wild-type and mutant dopamine transporters in HEK293 cells (Schmitt et al., 2008). These authors concluded that analogues of desoxypipradrol were cocaine-like in that they preferred the outward-facing conformational state of the dopamine transporter.
We have extended the experiments of Maxwell, Ferris and others by examining the effects of desoxypipradrol in the nucleus accumbens, a critical centre in the brain reward system, rather than more heterogenous brain areas. Another advantage is that we measure endogenous rather than pre-loaded tritiated dopamine efflux, which is not stored by normal processes. Furthermore, we have used FCV which allows us to measure dopamine efflux in real-time on a sub-second timescale and we are able to measure re-uptake directly. We have also been able to differentiate changes in basal versus stimulated efflux. Taken together our data suggest that desoxypipradrol is more cocaine-like than amphetamine-like and is also more potent than cocaine, making this drug likely to have abuse potential. Furthermore, this increased potency compared with cocaine could account for desoxypipradrol’s psychotogenic effects.
The mechanism underlying the increased potency of desoxypipradrol versus cocaine on electrically evoked dopamine efflux in the accumbens is currently unknown, but could be due to increased affinity at the dopamine transporter or binding to a different site at the transporter. Furthermore, cocaine has recently been shown to increase the immediately releasable pool of dopamine via a synapsin-dependent mechanism (Venton et al., 2006) and it is possible that desoxypipradrol also has molecular targets other than the dopamine transporter, which contributes to the effects seen on stimulated dopamine efflux.
Footnotes
Funding
This work was funded by a generous start-up grant to CD from the Division of Biomedical Science, St George’s, University of London.
Conflict of interests
CD and JR have no conflict of interests.