
Abstract
Several phosphodiesterase inhibitors (PDEis) improve cognition, suggesting that an increase in brain cAMP and cGMP facilitates learning and memory. Since extinction of drug-seeking behavior requires associative learning, consolidation and formation of new memory, the present study investigated the efficacy of three different PDEis in the extinction of cocaine-induced conditioned place preference (CPP) in B6129S mice. Mice were conditioned by escalating doses of cocaine which was resistant to extinction by free exploration. Immediately following each extinction session mice received (a) saline/vehicle, (b) rolipram (PDE4 inhibitor), (c) BAY-73-6691 (PDE9 inhibitor) or (d) papaverine (PDE10A inhibitor). Mice that received saline/vehicle during extinction training showed no reduction in CPP for >10 days. BAY-73-6691 (a) dose-dependently increased cGMP in hippocampus and amygdala, (b) significantly facilitated extinction and (c) diminished the reinstatement of cocaine CPP. Rolipram, which selectively increased brain cAMP levels, and papaverine which caused increases in both cAMP and cGMP levels, had no significant effect on the extinction of cocaine CPP. The results suggest that increase in hippocampal and amygdalar cGMP levels via blockade of PDE9 has a prominent role in the consolidation of extinction learning.
Introduction
The administration of a drug that changes the affective state of the organism in a specific context triggers an associative learning process and the formation of long-term memory (LTM). The expression of conditioned place preference (CPP) is viewed as a test for reactivity to a drug-associated conditioned stimulus (CS); this test has validity for cue-reactivity in human drug users. The CPP paradigm has also been used to investigate extinction of ‘drug-seeking behavior’ and reinstatement of conditioned response (Aguilar et al., 2009; Itzhak and Martin, 2002; Mueller and Stewart, 2000; Mueller et al., 2002; Parker and McDonald, 2000). Interestingly, reinstatement of cocaine CPP following extinction is a drug-specific phenomenon that can be triggered only by drugs that share a similar mechanism of action with that of cocaine (Itzhak and Martin, 2002). Therefore, the reinstatement of place preference, like the reinstatement of drug self-administration, presents a meaningful resource for the study of relapse. Animal and human studies suggest that re-exposure to a low dose of psychostimulants, opiates or alcohol, following abstinence or extinction of drug use, may cause relapse (Shaham et al., 2003). It is therefore critical to develop pharmacotherapies and behavioral practices by which extinction of drug-seeking behavior will ultimately result in resistance to both drug-associated cues and drug-priming.
Extinction learning by ‘exposure therapy’ is thought to be essential for the management of drug addiction (Carter and Tiffany, 1999; Powell et al., 1993; Siegel and Ramos, 2002). Extinction typically requires long or multiple re-exposures to a CS (Nader, 2003; Power et al., 2006). Results from fear conditioning studies suggest that the extinction process does not eliminate or cause ‘unlearning’ of the initial conditioned response; rather, the organism learns that the CS no longer elicits the previous stimulus (Bouton, 2002, 2004; Havermans and Jansens, 2003). Thus, extinction requires associative learning, consolidation and formation of new memory (Milad and Quirk, 2002; Santini et al., 2001).
Through the activation of their respective kinases (PKA and PKG), cyclic nucleotide (cAMP and cGMP) signaling pathways are important regulators of neural function and synaptic homeostasis (Bales et al., 2010). While adenylyl cyclase (AC) and guanylyl cyclase (GC) generate the second messengers cAMP and cGMP respectively, phosphodiesterases (PDEs) hydrolyze these cyclic nucleotides into their inactive monophosphates, 5’-AMP and 5’-GMP, and thereby contribute to the regulation of their intracellular levels (Essayan, 2001). Eleven different families of mammalian PDEs have been identified in the central nervous system (CNS) and periphery. All neurons express multiple PDEs which differ in cyclic nucleotide specificity, affinity, regulatory control and subcellular distribution (Bender and Beavo, 2006; Blokland et al., 2006; Boswell-Smith et al., 2006; Menniti et al., 2006).
The differential localization of PDEs in the CNS and periphery determine how effective phosphodiesterase inhibitors (PDEis) are at regulating different processes. Brain PDEs include PDE1, PDE2, PDE4, PDE5, PDE9, PDE10 and PDE11. PDE4 is widely distributed throughout the brain (Bender and Beavo, 2006). The PDE4 inhibitor rolipram has been used as an anti-inflammatory and has shown antipsychotic-like therapeutic effects (Kelly et al., 2007). Rolipram enhances learning and memory in various paradigms (Cheng et al., 2010; Monti et al., 2006; Rose et al., 2005; Tully et al., 2003; Zhang and O’Donnell, 2000), but unexpectedly it disturbed expression and extinction of conditioned fear in mice (Mueller et al., 2010). PDE9 is highly localized in all sub-areas of the hippocampus (van Staveren et al., 2002, 2004; Reyes-Irisarri et al., 2007) and the specific PDE9 inhibitor BAY-73-6691 improves learning and memory in rodents (van der Staay et al., 2008). PDE10A is densely localized in the striatum but less in the hippocampus (Seeger et al., 2003). Papaverine is a specific inhibitor of PDE10A that increased levels of cAMP and cGMP (Siuciak et al., 2006) and improved phencyclidine-induced cognitive deficits in rats (Rodefer et al., 2005).
The use of selective PDEis as potential cognitive enhancers is suggested by studies in which PDEis facilitated learning and memory in animal models with experimentally induced learning and memory deficits (Bender and Beavo, 2006; Blokland et al., 2006; Boswell-Smith et al., 2006; Menniti et al., 2006). However, it is unclear whether selective PDEis facilitate learning and memory in subjects with no cognitive impairments. Therefore, the effects of the PDEis on extinction learning may be different than their effects on improving learning following cognitive deficits.
The hippocampus and amygdala are implicated in spatial/contextual and emotional/cued memory, respectively. We hypothesized that increases in hippocampal and amygdalar cyclic nucleotide levels through the action of PDEis will facilitate extinction learning of cocaine-induced place preference. We sought to investigate PDEis with different specificities to cAMP and cGMP in order to determine which if any group of PDEis has a more prominent role in the consolidation of extinction learning. We first determined how cyclic nucleotide levels were affected in the hippocampus and amygdala in response to a PDE4 (cAMP specific) inhibitor rolipram, a PDE9 (cGMP specific) inhibitor BAY-73-6691 and a PDE10A (dual specificity) inhibitor papaverine. Then, the efficacies of these PDEis to extinguish and prevent reinstatement of cocaine CPP were investigated. We report that of the three PDEis only BAY-73-6691, which increased hippocampal and amygdalar cGMP levels, accelerated the extinction and attenuated the reinstatement of cocaine place preference.
Materials and methods
Animals
Male B6129S adult mice (8–10 weeks old, weighing 25–35 g) were supplied from breeding colonies in our facilities at the University of Miami, Miller School of Medicine, Miami, FL, as described previously in detail (Balda et al., 2006). Following weaning (postnatal day 21), mice were housed in single-sex groups; males were used for the current study. The ‘litter effect’ was negated by grouping mice from 4–5 different litters into each cage. Animals were housed in a temperature (22 ± 0.5 °C) and humidity (50%) controlled room and maintained on a 12-hour light/dark schedule with food and water ad lib except during training and testing. Animal care was in accordance with the Guide for the Care and Use of Laboratory Animals (National Research Council and National Academy Press, 1996) and approved by the University of Miami Animal Care and Use Committee.
Measurement of cyclic nucleotide levels
Our studies focused on the effects of the PDEis on cyclic nucleotides levels particularly in hippocampus and amygdala because these brain regions are involved in associative learning (Robbins et al., 2008). Papaverine (Sigma, St Louis, MO) was dissolved in saline (0.9% NaCl); rolipram (Sigma, St Louis, MO) was dissolved in 2% DMSO (Sigma, St Louis, MO); BAY-73-6691 (Sigma, St Louis, MO) was dissolved in 10% Solutol® HS 15 (BASF, Ludwigshafen, Germany) solution (vehicle). To determine the dose effects of the PDEis, mice received single intraperitoneal (IP) injections of (a) rolipram (0.05, 0.25, 1 mg/kg; n = 3/group), (b) BAY-73-6691 (0.03, 0.3 and 3 mg/kg; n = 3/group), (c) papaverine (10, 20, 40 mg/kg; n = 3–4/group) or (d) saline/vehicle (n = 3/group). Thirty minutes later, mice were sacrificed by cervical dislocation and brains were immediately removed and placed in ice-cold saline. The hippocampus and amygdala were dissected on an ice-cold surface according to The Mouse Brain atlas (Paxinos and Franklin, 2001), snap-frozen on dry ice and stored at −80°C. Levels of cGMP (sensitivity, 25 fmol/mL) and cAMP (sensitivity, 0.39 pmol/mL) were quantified with direct EIA kits (Enzo Life Sciences International, Inc., PA) as described in the manufacturer’s protocols and determined by spectrophotometry (OD450 nm). The optimal doses of the PDEis for the behavioral studies were determined from these experiments.
Place conditioning apparatus
An Opto-Max Activity Meter (Colombus Instruments, Columbus, Ohio, USA) was used to monitor place preference. The training context consisted of two compartments separated by a removable divider. One compartment had black walls and smooth black floor while the other compartment had white walls with a floor covered with sandpaper (fine grit 150C, Norton; Stephenville, TX) thus providing distinct visual and tactile cues. Each compartment was scanned by seven infrared beams at a rate of 10 Hz (2.54 cm intervals). The horizontal sensors were mounted alongside opposing lengths of each compartment. A null zone of 8 cm was assigned at the interface of the two compartments to ensure that only full entry into one compartment or the other was registered as distinct time spent on each side. Time spent in each compartment and locomotor activity were recorded and analyzed by the Opto-max interface and software.
Conditioning
Mice were trained and tested in a room separate from the housing room as described previously (Itzhak and Anderson, 2012). The testing room was equipped with a fluorescent lamp strategically positioned to create a dimmed lighting environment. On the first day of each experiment, between 12:00 and 14:00, mice were habituated for 20 minutes to the training context; time spent in each compartment was recorded. Extremely biased mice (about 10%) were eliminated from the study. About 50% of the remaining animals had slight preference for the black compartment and the other 50% had slight preference for the white compartment. Accordingly, the assignment criterion was such that mice were conditioned by cocaine in their least preferred compartment. Thus, the training procedure was counterbalanced where half the mice were trained with cocaine in the black compartment while the other half of the mice were trained with cocaine in the white compartment. Mice were trained with IP injections of saline during the morning (10:00–12:00) session and cocaine during the afternoon (14:00–16:00) session. Each conditioning session lasted 30 minutes.
Mice were conditioned with ascending dosages (3, 6, 12, 24 mg/kg) of cocaine over 4 days (Table 1). This regimen of escalating dosage caused a CPP that was of higher magnitude and longer lasting than the CPP that resulted from a fixed daily dose; this enhanced CPP was also resistant to extinction by free exploration (Itzhak and Anderson, 2012). Hence, we hypothesized that investigating extinction in a model of robust conditioning is more significant to the real-life human situation of escalating drug use than the typical model of conditioning (fixed daily dose of cocaine) which affords relatively quick extinction.
Schematic presentation of the timeline and different phases of the extinction experiment.

CPP, conditioned place preference; PDE, phosphodiesterase
Cocaine was administered immediately before the animal was placed into its respective compartment. To maintain a consistent environment for each mouse, the sandpaper was removed and the cages thoroughly cleaned with dilute laboratory-grade detergent followed by water and then dried, following each training session. The locomotor activity in response to the different doses of cocaine was recorded daily.
CPP test
The expression of place preference was tested 3 days after the final training session in order to eliminate possible contributions from residual cocaine administration on the last training session and also to allow the consolidation of long-term memory of drug reward. Similar to habituation, CPP testing was carried out between 12:00 and 14:00.
Extinction and reinstatement
Conditioned mice were allowed free exploration for 20 minutes in the training context 24 hours after the CPP test. Since we aimed to facilitate the consolidation of extinction, immediately following each extinction session mice received single IP injections of either (a) vehicle (n = 5; n = 8)/saline (n = 5), (b) rolipram (0.25 mg/kg, n = 6), (c) BAY-73-6691 (3 mg/kg, n = 10), or (d) papaverine (20 mg/kg, n = 6) and were returned to their home cages. Rates of extinction, herein defined as a significant reduction in the magnitude of CPP compared with CPP test, were recorded daily. Mice received a 2-day break from extinction training after 4 days. Training resumed the following day and continued for the next 3 days (Table 1).
Twenty-four hours after the last extinction session, mice received a priming dose of cocaine (12 mg/kg; the average daily dose during conditioning) in the absence of the PDEi. Time spent in each compartment as well as locomotor activity was recorded for 20 minutes.
Statistics
Changes in levels of cyclic nucleotides in response to different doses of a PDEi were analyzed by one-way analysis of variance (ANOVA) supplemented by Holm–Sidak method for post hoc analysis. Results of CPP are presented as mean ± standard error of the mean (SEM) of the difference in time spent in cocaine- and saline-paired compartments. The overall effect on acquisition, extinction and reinstatement of cocaine-induced place preference was analyzed by two-way ANOVA (group × time) general linear model supplemented with post hoc multiple pair-wise comparisons using the Tukey test. Further, unpaired Student’s t-test or Mann–Whitney rank sum test where appropriate were used to compare differences in the magnitude of CPP upon cocaine priming versus the final day of extinction training. For all measures, significance was considered from a value of p < 0.05.
Results
Effects of PDEis on cAMP and cGMP levels
The PDE4 inhibitor rolipram dose dependently increased levels of cAMP in both the hippocampus and amygdala (Figure 1A) but had no effect on cGMP levels (data not shown). Rolipram treatment resulted in an overall significant increase in cAMP in both the hippocampus (F[3,8] = 18.463, p < 0.001) and amygdala (F[3,8] = 17.824, p < 0.001). At doses of 0.25 and 1 mg/kg rolipram administration resulted in about two-fold increase in cAMP in the amygdala (t = 6.102, p < 0.001 and t = 6.166, p < 0.001, respectively) and hippocampus (t = 6.072, p < 0.001 and t = 4.858, p < 0.001, respectively) compared with vehicle controls; a dose of 0.05mg/kg rolipram also significantly increased levels of cAMP in the amygdala (t = 2.606, p = 0.031). A dose of 0.25 mg/kg rolipram was used for the subsequent behavioral experiment. The PDE9 inhibitor BAY-73-6691 dose dependently increased levels of cGMP in both the hippocampus (F[3,8] = 27.052, p < 0.001) and amygdala (F[3,8] = 7.941, p < 0.001) (Figure 1B) but had no effect on levels of cAMP (data not shown). A dose of 3 mg/kg of BAY-73-6691 was used in the extinction experiments because this dose resulted in the highest increase in levels of cGMP (t = 4.208, p = 0.003 and t = 8.549, p < 0.001) for amygdala and hippocampus, respectively. Significant increases in cGMP were also observed in the hippocampus for dosages of 0.03 mg/kg (t = 3.157, p = 0.013) and 0.3 mg/kg (t = 5.969, p < 0.001) and amygdala at 0.3 mg/kg (t = 3.272, p = 0.011). An overall significant effect of papaverine on levels of cAMP was observed in the hippocampus (F[3,8] = 6.531, p = 0.015) but not in the amygdala (F[3,9] = 1.225, p = 0.356). Significant increases in cGMP in both the hippocampus (F[3,10] = 7.613, p = 0.006) and amygdala (F[3,10] = 4.929, p = 0.024) were observed. Holm–Sidak post hoc analysis showed that the PDE10A inhibitor papaverine at 10, 20 and 40 mg/kg significantly increased levels of cAMP in the hippocampus (t = 3.997, p = 0.004; t = 3.640, p = 0.007; and t = 2.674, p = 0.028, respectively) but not in the amygdala (Figure 1C). However, post hoc comparisons showed that papaverine increased cGMP in both the hippocampus at 10, 20 and 40 mg/kg (t = 2.984, p = 0.014; t = 4.286, p = 0.002; and t = 4.077, p = 0.002, respectively) and the amygdala at 40 mg/kg (t = 3.715, p = 0.004) (Figure 1D). A dose of 20 mg/kg was chosen for further experiments because (a) unpaired Student’s t-test showed a significant difference between controls and mice that received 20 mg/kg papaverine (t[5] = −9.542, p < 0.001) and (b) the high dose of 40 mg/kg resulted in ataxia in mice. Differences in control levels of cAMP in Figure 1A compared with Figure 1C may have arisen because two different vehicles were used, 2% DMSO for rolipram experiment versus saline for papaverine experiment. Similarly, in Figure 1B, the vehicle for BAY-73-6691 was Solutol HS 15 while that for papaverine in Figure 1D was saline.
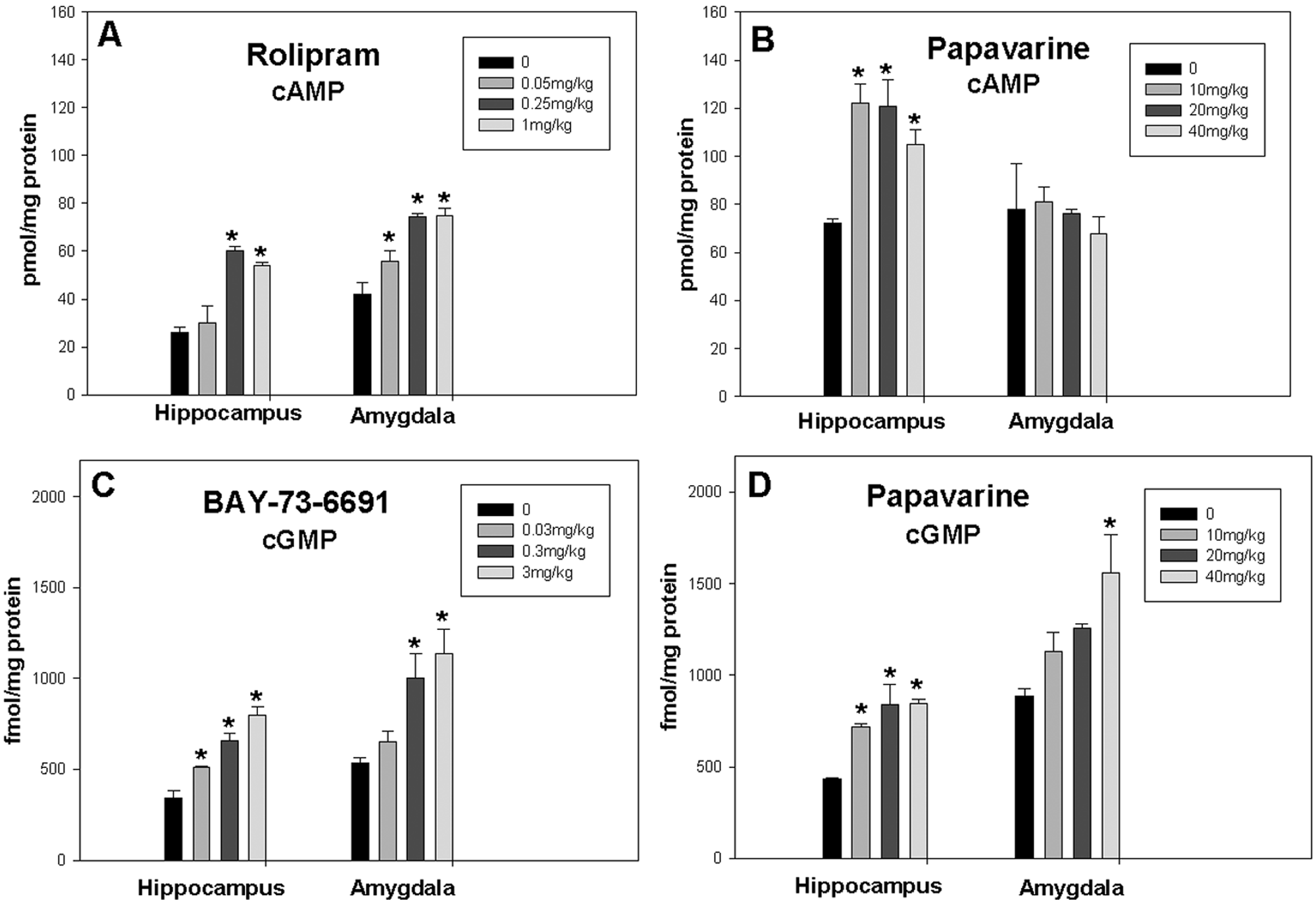
Effect of different doses of phosphodiesterase (PDE) inhibitors on levels of cAMP and cGMP in the hippocampus and amygdala. (A) Rolipram (n = 3/group) significantly increased levels of cAMP in the hippocampus (F[3,8] = 18.463, p < 0.001) and amygdala (F[3,8] = 17.824, p < 0.001). (B) BAY-73-6691 (n = 3/group) dose dependently increased levels of cGMP in the hippocampus (F[3,8] = 27.052, p < 0.001) and amygdala (F[3,8] = 7.941, p < 0.001). (C) Papaverine (n = 3–4/group) significantly increased levels of cAMP in the hippocampus (F[3,8] = 6.531, p = 0.015) but not in the amygdala (F[3,9] = 1.225, p = 0.356). (D) Papaverine significantly increased levels of cGMP in the hippocampus (F[3,10] = 7.613, p = 0.006) and amygdala (F[3,10] = 4.929, p = 0.024). Data are presented as mean ± SEM (*p < 0.05 compared with control groups, Holm–Sidak method post hoc).
Extinction of cocaine-induced place preference
Mice that received saline or vehicle during extinction training did not show ‘extinction’ (Figure 2). These results are consistent with a previous study showing that conditioning by escalating doses of cocaine resulted in extinction-resistant CPP (Itzhak and Anderson, 2012). Administration of rolipram immediately after each extinction session for a total of 8 days had no effect on the magnitude of place preference compared with the vehicle group (Figure 2A). In a separate experiment, prolonged extinction training (12 days) followed by rolipram administration had also no effect (data not shown). Since there was no evidence of extinction in both groups, a test for reinstatement was not performed.
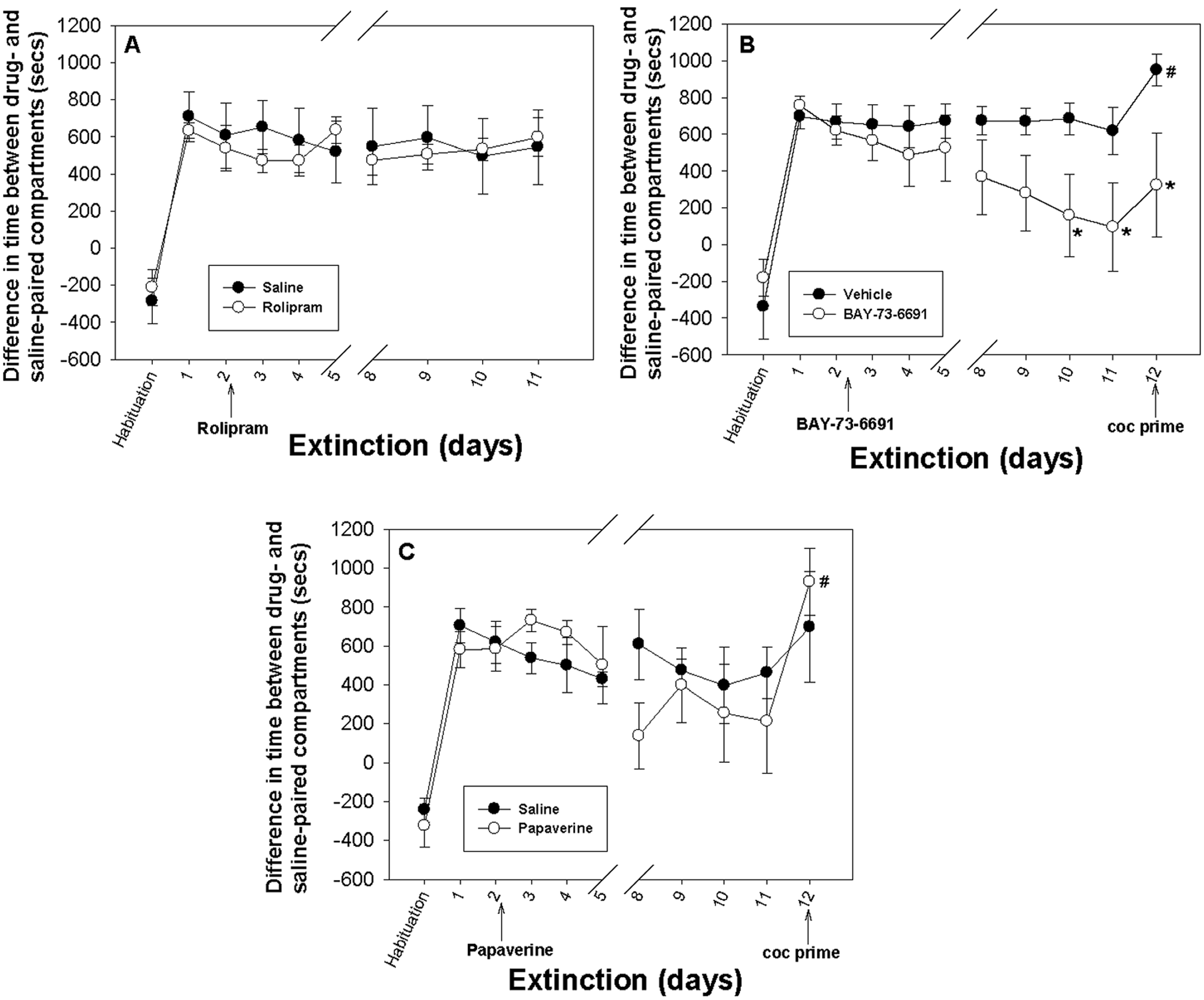
Effect of different phosphodiesterase (PDE) inhibitors on extinction of cocaine-induced conditioned place preference (CPP). The y-axes represent the mean ± SEM of difference in time spent on the drug-paired versus the saline-paired compartment. The x-axis shows the timeline for the extinction experiment. Day 1 represents the CPP test. The test drugs and vehicles were administered from the second day (first extinction training session) and for 7 more days with a 2-day break as illustrated between days 5 and 8. (A) Rolipram (n = 6) had no significant effect on extinction of cocaine-induced place preference. (B) BAY-73-6691 (n = 10) significantly reduced the magnitude of CPP over time (F[1,176] = 12.168, p < 0.001 two-way analysis of variance [ANOVA; group × time] general linear model). Post hoc comparisons by Tukey test showed significant differences between the two groups on days 10 (q = 3.307, *p = 0.019), 11 (q = 3.402, *p = 0.016) and 12 (q = 3.932, *p = 0.005). The vehicle-treated group showed a significant increase in place preference upon cocaine priming (t[14] = −2.314; #p = 0.036). (C) There was no significant difference in the magnitude of CPP between the papaverine-treated (n = 6) and the saline-treated (n = 5) groups over time (F[1,99] = 0.380, p = 0.539, two-way ANOVA). The papaverine-treated group showed a significant increase in place preference upon cocaine priming (T = 98, #p < 0.003, Mann–Whitney rank sum test) compared with day 11 which was the final day of extinction training.
Administration of BAY-73-6691 immediately after each extinction session resulted in significant reduction in the magnitude of CPP compared with controls (F[1,176] = 12.168, p < 0.001, two-way ANOVA) (Figure 2B). Post hoc comparison by Tukey test showed significant differences between the two groups on days 10 (q = 3.307, p = 0.019), 11 (q = 3.402, p = 0.016) and 12 (q = 3.932, p = 0.005). On day 11, the difference between times spent in drug- and saline-paired compartments was 95 ± 162 seconds (Figure 2B), suggesting extinction of CPP since this was significantly different from CPP test (T = 133, p = 0.038, Mann–Whitney rank sum test ). BAY-73-6691 also attenuated the reinstatement of CPP; the difference between the magnitude of CPP on the final day of extinction (day 11) and the day of cocaine priming (day 12) was not significant (T = 90, p = 0.273, Mann–Whitney rank sum test). However, the vehicle treated group showed a significant increase in the magnitude of CPP following cocaine priming compared with the final day of extinction (t[14] = −2.314, p = 0.038) (Figure 2B). This finding suggests that BAY-73-6691 may have provided partial resistance against reinstatement of cocaine CPP. The differences in extinction rate and resistance to reinstatement in mice treated with BAY-73-6691 versus vehicle control mice were not due to differences in locomotor behavior. Results of locomotor activity, which was recorded during each session, showed that during the reinstatement test, ambulatory counts in control and BAY-73-6691 groups were 1560 ± 143 and 1642 ± 139 counts/20 min, respectively.
Finally, administration of papaverine immediately after each extinction session had no significant effect on extinction (F[1,99] = 0.539, p > 0.05) (Figure 2C). The magnitude of CPP in the papaverine group fluctuated over time. Although a reduction in CPP was observed on days 8–11 (Figure 2C), the differences between the control and the papaverine group did not reach statistical significance. However, because there was a trend of reduction in the magnitude of CPP in both groups, challenge cocaine (12 mg/kg) was given on day 12. A significant increase in CPP was observed in the papaverine-treated group (T = 98, p < 0.003, Mann–Whitney rank sum test) upon cocaine priming. However, the increase in CPP following a priming injection to the control group was not statistically significant (Figure 2C).
Discussion
The current study employed an escalating dose schedule for cocaine-induced CPP as opposed to a typical fixed schedule used in other studies. Mice conditioned by ascending doses of cocaine maintain a higher magnitude of CPP and show greater resistance to extinction compared with mice conditioned by a fixed daily dose of cocaine (Itzhak and Anderson, 2012). Furthermore, an increasing dose schedule more closely resembles human drug use, in which the quantity of drug increases over time. Indeed results depicted in Figure 2 indicate that cocaine CPP was long lasting in control groups that received saline/vehicle during post-extinction training. Along this vein, a PDEi that successfully extinguishes place preference at its highest degree, and provides resistance to cocaine reinstatement, may be particularly valuable relative to its effect on extinction of a weaker conditioned response that results from conditioning by a fixed dose of cocaine.
The purpose of this study was to determine how different PDEi with differential specificity for elevating levels of cAMP and cGMP influence extinction learning. Our studies focused on the effects of the PDEi on the levels of cyclic nucleotides in the hippocampus and amygdala because these brain regions have a role in spatial and emotional learning, respectively, associated with the CPP paradigm.
Since PDE4 is highly expressed in the hippocampus, investigation of a specific PDE4 inhibitor seemed feasible to study consolidation of extinction. Here we show that although acute injection of rolipram increased levels of cAMP in both the hippocampus and amygdala, it did not influence the extinction of cocaine-induced place preference. Treatment with rolipram during or 30 min prior to but not following cocaine administration attenuated CPP (Thompson et al., 2004) and reduced cocaine-induced behavioral sensitization (Janes et al., 2009). This suggests that rolipram is not effective in reducing cocaine effects if given after drug administration. Therefore, it appears that an increase in cAMP may not facilitate extinction of cocaine CPP. Although the present study was not focused on determination of compensatory mechanism associated with repeated rolipram administration, it has been reported that 2 weeks administration of rolipram resulted in a 17-fold increase in PDE4D3 and a down-regulation of PDE4A1 (30%) and PDE4A5 (37%) in rat hippocampus (Dlaboga et al., 2006). The latter may also explain why rolipram did not accelerate the extinction of place preference.
We next investigated whether selective increases in brain cGMP facilitate extinction learning. BAY-73-6691 is a selective inhibitor of PDE9 and may be a potential therapeutic for Alzheimer’s disease (Bender and Beavo, 2006). PDE9 is one of the most recently discovered PDE families with the highest affinity for cGMP. In rat and mouse brain, high densities of PDE9 were found in regions containing soluble guanlyl cyclase (sGC) and neuronal nitric oxide synthase (nNOS) including the hippocampus, amygdala, olfactory bulb, allocortex and basal ganglia (Andreeva et al., 2001; van Staveren et al., 2002). BAY-73-6691 has been shown to improve learning and memory in rodents (van der Staay et al., 2008).
In the present study, acute injection of BAY-73-6691 increased levels of cGMP in the hippocampus and amygdala, accelerated extinction of cocaine-induced place preference and attenuated place preference reinstatement upon cocaine priming. A plausible explanation for BAY-73-6691 accelerating extinction learning is the involvement of cGMP in the NO/sGC/PKG/CREB signaling pathway. It has been reported that elevated cGMP levels in the hippocampus and amygdala, brain regions known to be important for learning and memory, enhances synaptic plasticity through NO which acts both pre- and post-synaptically to promote LTP (Arancio et al., 2001; Kleppisch and Feil, 2009; Lu et al., 1999; Ota et al., 2010; Son et al., 1998).
The PDE5 inhibitor sildenafil (Viagra), which also increases cGMP in the CNS and periphery (Rutten et al., 2005), reversed memory deficits due to nNOS inhibition in rats (Devan et al., 2006). In the present study, BAY-73-6691 was chosen because of its high selectivity for PDE9. Also, PDE9 is distributed in brain regions involved in learning and memory whereas PDE5 is expressed in the cerebellum and to a lesser degree the hippocampus (Menniti et al., 2006).
The K m of PDE9 is in the range of 170 nM for cGMP and the Vmax is about 4.9 nM/min/µg of recombinant protein which is about twice as fast as the Vmax of PDE4 for cAMP (Fisher et al., 1998). This accelerated rate of cGMP hydrolysis may partially explain why BAY-73-6691 was more successful at facilitating extinction than rolipram. By blocking the catalytic activity of PDE9, the rapid increase in cGMP may have more significant effects on downstream molecules that are involved in memory consolidation (e.g. CREB). The impact of other PDEs that degrade cGMP may be less significant because the pharmacokinetics of PDE9 may overshadow the kinetics of the other PDEs.
After considering PDEs that specifically increases cAMP (rolipram) or cGMP (BAY-73-6691) we investigated whether a dual specificity PDEi which elevates levels of both cAMP and cGMP would accelerate extinction of place preference. Papaverine is a specific inhibitor of PDE10A that increased levels of cAMP and cGMP (Siuciak et al., 2006) and improved phencyclidine-induced cognitive deficits in rats (Rodefer et al., 2005). We found that acute injection of papaverine significantly elevated cGMP levels in both the hippocampus and amygdala, but it only increased levels of cAMP in the hippocampus. However, papaverine had no significant effect on extinction of CPP (Figure 2C).
Studies have shown that PDE10A is densely localized in the striatum but less so in the hippocampus (Seeger et al., 2003), while PDE9 is highly localized in all sub-areas of the hippocampus (van Staveren et al., 2002, 2004; Reyes-Irisarri et al., 2007). Given the role of hippocampus in extinction learning, it is likely that inhibiting PDE9 has a more profound effect on hippocampal functions than inhibiting PDE10A. This was manifested through the behavioral experiment where BAY-73-6691 but not papaverine, facilitated new learning and thereby accelerated extinction of cocaine-induced place preference. Another possible explanation for the failure of papaverine to accelerate extinction is that PDE9 metabolizes cGMP at a very high rate. Owing to this, while papaverine blocks hydrolysis of cGMP by PDE10A, PDE9 activity may increase and quickly reduce levels of cGMP. In addition, PDE10A has a much higher affinity for cAMP than cGMP (Boswell-Smith et al., 2006; Francis et al., 2010) and it does not hydrolyze cGMP as efficiently as PDE9. This may also explain the finding that blockade of PDE9 (BAY-73-6691) afforded accelerated extinction while blockade of PDE10A (papaverine) did not (Figure 2B and C). However, further studies are required to determine the long-term effects of repeated administration of PDEis; this will help to elucidate whether a compensatory action of PDEs could explain differential efficacies of the different PDEis.
In summary, the present study suggests that inhibitors of PDE9 have a prominent role in the consolidation of extinction learning. It also appears that targeting a specific PDE is more critical than targeting any PDE which metabolizes cGMP.
Footnotes
Acknowledgements
We are grateful to LiZi Tan for her assistance in data collection.
Funding
This work was supported by the National Institute on Drug Abuse, National Institutes of Health, USA (grant numbers R01DA026878 and R21DA029404).
Conflict of interest
The authors declare that they do not have any conflict of interest.